

Лекция 1.ppt_Биология
- Количество слайдов: 10
 Лекция 1 Ископаемые свидетельства о жизни минувших эпох. Геохронология и стратиграфия. География и климат прошлого. Начальные этапы развития жизни на Земле.
Лекция 1 Ископаемые свидетельства о жизни минувших эпох. Геохронология и стратиграфия. География и климат прошлого. Начальные этапы развития жизни на Земле.
 В пределах данного курса на кафедре Зоологии будет рассматриваться один из разделов общей биологии – палеозоология, являющаяся составной частью палеонтологии. Палеонтология (палеобиология) – наука о живых организмах и экосистемах прошлых геологических эпох, в противоположность неонтологии. Основателями палеонтологии считаются Ж. Кювье (в области изучения вымерших позвоночных животных, Ж. –Б. Ламарк (беспозвоночных) и А. Броньяр (растений). Основы современной эволюционной палеонтологии заложены В. Ковалевским, и в нынешнем виде палеонтология неразрывно связана с теорией эволюции и эмбриологией. Палеонтология судит о жизни вымерших существ не по ним самим, а по оставленным ими “вещественным доказательствам”. Иногда это лишь следы жизни – биоглифы, или ихнофоссилия. Известны отпечатки ног самых разных животных, следы ползанья и зарывания, надгрызенная пища, окаменевший помет. Комплексы следов жизни называют ихноценозами. Ископаемые остатки самих организмов, как правило, сохраняются в осадочных породах. Это имеет место в водных бассейнах – морях, озерах, реках. Чем быстрее труп окажется в воде и чем быстрее в ней идет накопление осадка, тем выше шанс сохранения. Поэтому большая часть окаменелостей – это остатки водных животных, и притом живших во внутренних водоемах и по берегам морей. Из наземных животных лучше всего сохраняются обитатели речных долин. В то же время об обитателях гор и морских глубин мало что известно. Чем тонкозернистее осадок, тем детальнее окаменелость. От животных обычно сохраняются детали минерального скелета, поэтому в знаниях о мягкотелых беспозвоночных так же очень много пробелов. Ископаемые остатки могут сохранять свой первичный состав или подвергаться фоссилизации – замещению окружающей минеральной породой. Виды ископаемых остатков: слепок, внутреннее ядро (оно не передает внешних особенностей оригинала), внешнее ядро (не передает внутренних особенностей). Редкий случай фоссилизации, при которой первоначальное захоронение происходит без участия осадочных пород – занесение организмов на суше песком. Таковы некоторые находки из пустыни Гоби. Другие пути сохранения организмов, не связанные напрямую с занесением осадком как способом захоронения – инклюзы в смоле (янтарь), захоронение в озокерите, вечной мерзлоте, мумификация; при этом не происходит замещение тканей минеральной породой. Раздел палеонтологии, изучающий закономерности захоронения, называется тафономией. Биоценоз (присущая определенному месту совокупность живых организмов) переходит в танатоценоз (совокупность этих организмов после смерти). Затем их останки заносятся осадком или захораниваются иным образом и образуют тафоценоз, а когда происходит их минерализация, становятся ориктоценозом. Чаще всего в захоронениях встречается мешанина остатков, принесенных водой из разных мест. Другой тип тафоценоза – захоронениея частей одного организма и разных организмов в прижизненной взаимосвязи, что гораздо ценнее. Иногда находят поломанные зубы хищников вблизи костей их жертв, скелеты жертв или плодов в полости тела, или даже сцепившиеся скелеты животных, погибших во время схватки. Обстоятельства смерти животных можно выяснить, опираясь на условия захоронения, состояние самих остатков и другие сопутствующие признаки; а по характеру смерти во многом можно судить и о характере жизни животного. Представить себе прижизненный облик и образ жизни многих ископаемых организмов было бы почти невозможно, если бы не были известны их современные родственники, часто имеющие с ними довольно отдаленное внешнее сходство. Классический пример – динозавры, на протяжении полутора сотен неверно реконструировавшиеся наподобие гигантских ящериц, пока не стало известно, что их ближайшие родственники – птицы.
В пределах данного курса на кафедре Зоологии будет рассматриваться один из разделов общей биологии – палеозоология, являющаяся составной частью палеонтологии. Палеонтология (палеобиология) – наука о живых организмах и экосистемах прошлых геологических эпох, в противоположность неонтологии. Основателями палеонтологии считаются Ж. Кювье (в области изучения вымерших позвоночных животных, Ж. –Б. Ламарк (беспозвоночных) и А. Броньяр (растений). Основы современной эволюционной палеонтологии заложены В. Ковалевским, и в нынешнем виде палеонтология неразрывно связана с теорией эволюции и эмбриологией. Палеонтология судит о жизни вымерших существ не по ним самим, а по оставленным ими “вещественным доказательствам”. Иногда это лишь следы жизни – биоглифы, или ихнофоссилия. Известны отпечатки ног самых разных животных, следы ползанья и зарывания, надгрызенная пища, окаменевший помет. Комплексы следов жизни называют ихноценозами. Ископаемые остатки самих организмов, как правило, сохраняются в осадочных породах. Это имеет место в водных бассейнах – морях, озерах, реках. Чем быстрее труп окажется в воде и чем быстрее в ней идет накопление осадка, тем выше шанс сохранения. Поэтому большая часть окаменелостей – это остатки водных животных, и притом живших во внутренних водоемах и по берегам морей. Из наземных животных лучше всего сохраняются обитатели речных долин. В то же время об обитателях гор и морских глубин мало что известно. Чем тонкозернистее осадок, тем детальнее окаменелость. От животных обычно сохраняются детали минерального скелета, поэтому в знаниях о мягкотелых беспозвоночных так же очень много пробелов. Ископаемые остатки могут сохранять свой первичный состав или подвергаться фоссилизации – замещению окружающей минеральной породой. Виды ископаемых остатков: слепок, внутреннее ядро (оно не передает внешних особенностей оригинала), внешнее ядро (не передает внутренних особенностей). Редкий случай фоссилизации, при которой первоначальное захоронение происходит без участия осадочных пород – занесение организмов на суше песком. Таковы некоторые находки из пустыни Гоби. Другие пути сохранения организмов, не связанные напрямую с занесением осадком как способом захоронения – инклюзы в смоле (янтарь), захоронение в озокерите, вечной мерзлоте, мумификация; при этом не происходит замещение тканей минеральной породой. Раздел палеонтологии, изучающий закономерности захоронения, называется тафономией. Биоценоз (присущая определенному месту совокупность живых организмов) переходит в танатоценоз (совокупность этих организмов после смерти). Затем их останки заносятся осадком или захораниваются иным образом и образуют тафоценоз, а когда происходит их минерализация, становятся ориктоценозом. Чаще всего в захоронениях встречается мешанина остатков, принесенных водой из разных мест. Другой тип тафоценоза – захоронениея частей одного организма и разных организмов в прижизненной взаимосвязи, что гораздо ценнее. Иногда находят поломанные зубы хищников вблизи костей их жертв, скелеты жертв или плодов в полости тела, или даже сцепившиеся скелеты животных, погибших во время схватки. Обстоятельства смерти животных можно выяснить, опираясь на условия захоронения, состояние самих остатков и другие сопутствующие признаки; а по характеру смерти во многом можно судить и о характере жизни животного. Представить себе прижизненный облик и образ жизни многих ископаемых организмов было бы почти невозможно, если бы не были известны их современные родственники, часто имеющие с ними довольно отдаленное внешнее сходство. Классический пример – динозавры, на протяжении полутора сотен неверно реконструировавшиеся наподобие гигантских ящериц, пока не стало известно, что их ближайшие родственники – птицы.
 Ископаемые остатки организмов, а также следы их жизни, сохранившиеся в последовательных слоях осадочных пород, составляют палеонтологическую летопись. Только лишь благодаря ей картина развития жизни на Земле воссоздана быть не может, так как эта летопись очень неполна. Мало что остается от мягкотелых животных, от некоторых организмов (особенно растений) остаются лишь разрозненные части. Возможность сохранения зависит и от образа жизни животных и их обилия. Поэтому многие группы могут надолго исчезать из летописи, а потом опять появляться. Например, морские ежи многочисленны в мелководных морских отложениях мела, не обнаружены в третичном периоде (возможно, также они было глубоководными) и изобильны сейчас. Щитни известны из триаса, не обнаружены в юре, найдены в мелу, не известны в третичном периоде и обычны сейчас. Считалось, что белемниты вымерли в мелу, но теперь найдены эоценовые представители этой группы. Целаканты считались мезозойскими рыбами, а в ХХ веке были найдены живьём. Проблема в том, что нет таких сред, в которых осадконакопление шло бы очень долго и непрерывно. Этим объясняется и то, что обычно виды появляются в летописи “готовыми”, исчезают и заменяются новыми “готовыми”, давая козырь креационистам. (Реальное превращение одних видов в другие все же было доказано благодаря длительному накоплению осадка в долго существовавших озерных экосистемах). В летописи сохраняются остатки обычных, широко распространенных видов, а они как раз наименее интересны для теории эволюции. Эволюционные преобразования должны происходить, в первую очередь, в малочисленных локальных популяциях, так как в них мутации легче могут перейти в гомозиготное состояние и стать материалом для отбора. Последовательность отложений, которую изучает раздел геологии, называемый стратиграфией, отражается в геохронологической шкале. Относительная шкала устанавливает лишь, что древнее (глубже), а что моложе (ближе к поверхности). Изотопная шкала устанавливает абсолютный возраст пород по содержанию продуктов радиоактивного распада в минералах. Уран-свинцовый метод основан на распаде U-235 и U-238, рубидий-стронциевый – Rb-87, калийаргоновый - К-40, самарий-неодимовый – Sm-147, радиоуглеродный – С-14 (для последних 60 тысяч лет). Для сопоставления возраста отложений из разных мест используют руководящее ископаемое. Это остатки организмов с широким распространением и узким вертикальным залеганием. Таковы виды фораминифер (с девона по современность), археоциат и плеченогих (палеозой), трилобитов (нижний палеозой), аммонитов (девон – мел), белемнитов (юра – мел), граптолитов (нижний палеозой), высших растений, в том числе их спор и пыльцы (с девона), двустворчатых моллюсков (с карбона), остракод (с девона), земноводных (пермь – триас), пресмыкающихся (пермь – мел), млекопитающих (с мела). При сопоставлении с изотопной шкалой оказывается, что руководящим ископаемым можно верить не всегда. Так, в Европе исчезновение гиппариона служит границей между и плиоценом и плейстоценом, тогда как в Африке он жил и в плейстоцене, аналогично мастодону в Северной Америке.
Ископаемые остатки организмов, а также следы их жизни, сохранившиеся в последовательных слоях осадочных пород, составляют палеонтологическую летопись. Только лишь благодаря ей картина развития жизни на Земле воссоздана быть не может, так как эта летопись очень неполна. Мало что остается от мягкотелых животных, от некоторых организмов (особенно растений) остаются лишь разрозненные части. Возможность сохранения зависит и от образа жизни животных и их обилия. Поэтому многие группы могут надолго исчезать из летописи, а потом опять появляться. Например, морские ежи многочисленны в мелководных морских отложениях мела, не обнаружены в третичном периоде (возможно, также они было глубоководными) и изобильны сейчас. Щитни известны из триаса, не обнаружены в юре, найдены в мелу, не известны в третичном периоде и обычны сейчас. Считалось, что белемниты вымерли в мелу, но теперь найдены эоценовые представители этой группы. Целаканты считались мезозойскими рыбами, а в ХХ веке были найдены живьём. Проблема в том, что нет таких сред, в которых осадконакопление шло бы очень долго и непрерывно. Этим объясняется и то, что обычно виды появляются в летописи “готовыми”, исчезают и заменяются новыми “готовыми”, давая козырь креационистам. (Реальное превращение одних видов в другие все же было доказано благодаря длительному накоплению осадка в долго существовавших озерных экосистемах). В летописи сохраняются остатки обычных, широко распространенных видов, а они как раз наименее интересны для теории эволюции. Эволюционные преобразования должны происходить, в первую очередь, в малочисленных локальных популяциях, так как в них мутации легче могут перейти в гомозиготное состояние и стать материалом для отбора. Последовательность отложений, которую изучает раздел геологии, называемый стратиграфией, отражается в геохронологической шкале. Относительная шкала устанавливает лишь, что древнее (глубже), а что моложе (ближе к поверхности). Изотопная шкала устанавливает абсолютный возраст пород по содержанию продуктов радиоактивного распада в минералах. Уран-свинцовый метод основан на распаде U-235 и U-238, рубидий-стронциевый – Rb-87, калийаргоновый - К-40, самарий-неодимовый – Sm-147, радиоуглеродный – С-14 (для последних 60 тысяч лет). Для сопоставления возраста отложений из разных мест используют руководящее ископаемое. Это остатки организмов с широким распространением и узким вертикальным залеганием. Таковы виды фораминифер (с девона по современность), археоциат и плеченогих (палеозой), трилобитов (нижний палеозой), аммонитов (девон – мел), белемнитов (юра – мел), граптолитов (нижний палеозой), высших растений, в том числе их спор и пыльцы (с девона), двустворчатых моллюсков (с карбона), остракод (с девона), земноводных (пермь – триас), пресмыкающихся (пермь – мел), млекопитающих (с мела). При сопоставлении с изотопной шкалой оказывается, что руководящим ископаемым можно верить не всегда. Так, в Европе исчезновение гиппариона служит границей между и плиоценом и плейстоценом, тогда как в Африке он жил и в плейстоцене, аналогично мастодону в Северной Америке.


 Используемая ныне геохронологическая шкала выглядит следующим образом: Эра Период Кайнозой Эпоха Четвертичный Третичный Голоцен Плейстоцен Плиоцен Миоцен Олигоцен Эоцен Палеоцен Начало, млн. лет назад 0, 01 1 5 25 38 55 65 Мел Юра Триас 140 205 245 Палеозой Пермь Карбон Девон Силур Ордовик Кембрий 290 360 408 436 505 570 Протерозой Венд Рифей Карелий 670 1650 3500 Мезоз ой Азой 4600
Используемая ныне геохронологическая шкала выглядит следующим образом: Эра Период Кайнозой Эпоха Четвертичный Третичный Голоцен Плейстоцен Плиоцен Миоцен Олигоцен Эоцен Палеоцен Начало, млн. лет назад 0, 01 1 5 25 38 55 65 Мел Юра Триас 140 205 245 Палеозой Пермь Карбон Девон Силур Ордовик Кембрий 290 360 408 436 505 570 Протерозой Венд Рифей Карелий 670 1650 3500 Мезоз ой Азой 4600
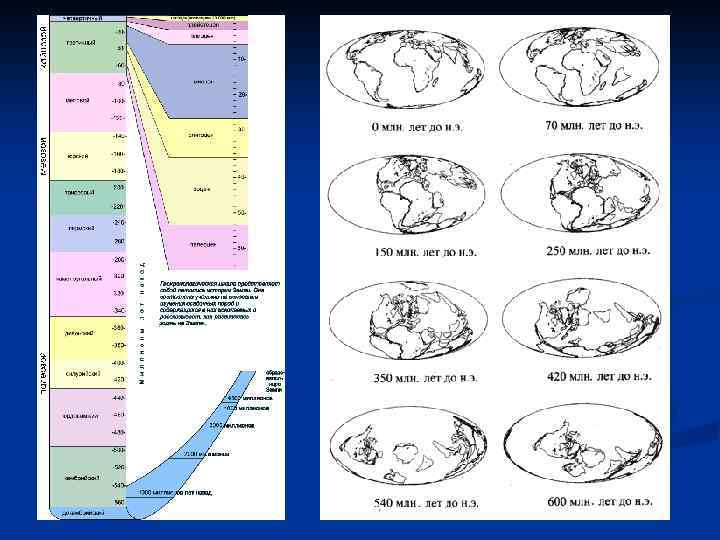
 Живые организмы неотделимы от среды обитания, а климат и расположение континентов не всегда были такими, как теперь. Земная кора находится в постоянном движении. Некоторые материки, если взглянуть на карту, выглядят как раздвинутые куски мозаики. Теперь известно, что на дне океанов имеются срединно-океанические хребты, по длине которые расположены вулканы. Породы, слагающие эти хребты, очень молоды, а с боков они все древнее и древнее. Новая кора возникает в гребне хребта и расползается по сторонам. Кора Земли состоит из нескольких плит, надстраивающихся по одному из краев, и разъезжающихся. Когда разъезжающиеся края встречаются, одна плита подныривает под другую, и разрушается в недрах Земли. Так ведут себя плиты, сложенные из океанического типа коры. Океанические хребты – их зона нарастания, океанические желоба – их зона разрушения. И то, и другое – источник вулканов и землетрясений. Континентальная кора легче океанической, она плавает по ней, как печенье по меду. Когда континенты доезжают до зоны разрушения, они останавливаются, так как не могут погрузиться в недра Земли, и сминаются, вздыбливая между собой горы. Так Гималаи возникли при столкновении Индии с Азией. Но положение континентальных плит и положение суши – это не одно и то же, так как уровень моря в разные времена колебался. Границы суши можно установить по осадочным породам, по ним же разгадывается и климат, так как сохраняются песчаные дюны пустынь, следы от капель дождя на глине, последствия ветров и наводнений и так далее. Изначально Южная Америка, Африка, Индия, Антарктида и Австралия составляли континент Гондвана, располагавшийся у Южного полюса. Азия была разделена на две части, находившиеся на экваторе. Северная Америка и Европа представляли собой цепь островов. Между Гондваной и Восточной Европой находился океан Прото-Тетис, а между Северной Америкой и Северной Европой – Япетус (будущий Атлантический океан). Во второй половине протерозоя многократно происходило горообразование. Оледенения случались в докембрии 2300, 900, 800 и 600 млн. лет назад. В раннем палеозое края материков были залиты мелководными морями. В позднем палеозое Япетус изчез, так как Северная Америка столкнулась со Скандинавией, образовав Каледонские озера. Снесенные реками осадки стали заполнять мелководные моря. Дельты и болота покрыли большие территории. В конце карбона Гондвана переехала на север и столкнулась с Северной Америкой и Европой, составлявшими тогда Лавразию. В перми все материки слились в Пангею. Мелководные моря по краям слившихся материков высохли, изменилась циркуляция воздуха и климат стал засушливым; случилось сильное похолодание (из-за континентального климата) и оледенение. В начале триаса продолжала существовать Пангея. Пермские горы сгладились до холмов. Затем Пангея раскололась на Гондвану и Лавразию. Обе Америки были пустынными, Африка, Европа и Сибирь – лесистыми. В юре море наступало и отступало, но в целом его уровень стал выше. Пангея продолжала раскалываться. Появилась рифтовая долина между Северной Америкой и Африкой. Климат стал влажнее и менее сезонным. В поздней юре мелководные моря проникли глубоко внутрь материков, Европа стала морем с островами, в Сибири образовался Обский бассейн. Климат был теплый, влажный и стабильный. Повсеместно распространились леса и речные заливные луга.
Живые организмы неотделимы от среды обитания, а климат и расположение континентов не всегда были такими, как теперь. Земная кора находится в постоянном движении. Некоторые материки, если взглянуть на карту, выглядят как раздвинутые куски мозаики. Теперь известно, что на дне океанов имеются срединно-океанические хребты, по длине которые расположены вулканы. Породы, слагающие эти хребты, очень молоды, а с боков они все древнее и древнее. Новая кора возникает в гребне хребта и расползается по сторонам. Кора Земли состоит из нескольких плит, надстраивающихся по одному из краев, и разъезжающихся. Когда разъезжающиеся края встречаются, одна плита подныривает под другую, и разрушается в недрах Земли. Так ведут себя плиты, сложенные из океанического типа коры. Океанические хребты – их зона нарастания, океанические желоба – их зона разрушения. И то, и другое – источник вулканов и землетрясений. Континентальная кора легче океанической, она плавает по ней, как печенье по меду. Когда континенты доезжают до зоны разрушения, они останавливаются, так как не могут погрузиться в недра Земли, и сминаются, вздыбливая между собой горы. Так Гималаи возникли при столкновении Индии с Азией. Но положение континентальных плит и положение суши – это не одно и то же, так как уровень моря в разные времена колебался. Границы суши можно установить по осадочным породам, по ним же разгадывается и климат, так как сохраняются песчаные дюны пустынь, следы от капель дождя на глине, последствия ветров и наводнений и так далее. Изначально Южная Америка, Африка, Индия, Антарктида и Австралия составляли континент Гондвана, располагавшийся у Южного полюса. Азия была разделена на две части, находившиеся на экваторе. Северная Америка и Европа представляли собой цепь островов. Между Гондваной и Восточной Европой находился океан Прото-Тетис, а между Северной Америкой и Северной Европой – Япетус (будущий Атлантический океан). Во второй половине протерозоя многократно происходило горообразование. Оледенения случались в докембрии 2300, 900, 800 и 600 млн. лет назад. В раннем палеозое края материков были залиты мелководными морями. В позднем палеозое Япетус изчез, так как Северная Америка столкнулась со Скандинавией, образовав Каледонские озера. Снесенные реками осадки стали заполнять мелководные моря. Дельты и болота покрыли большие территории. В конце карбона Гондвана переехала на север и столкнулась с Северной Америкой и Европой, составлявшими тогда Лавразию. В перми все материки слились в Пангею. Мелководные моря по краям слившихся материков высохли, изменилась циркуляция воздуха и климат стал засушливым; случилось сильное похолодание (из-за континентального климата) и оледенение. В начале триаса продолжала существовать Пангея. Пермские горы сгладились до холмов. Затем Пангея раскололась на Гондвану и Лавразию. Обе Америки были пустынными, Африка, Европа и Сибирь – лесистыми. В юре море наступало и отступало, но в целом его уровень стал выше. Пангея продолжала раскалываться. Появилась рифтовая долина между Северной Америкой и Африкой. Климат стал влажнее и менее сезонным. В поздней юре мелководные моря проникли глубоко внутрь материков, Европа стала морем с островами, в Сибири образовался Обский бассейн. Климат был теплый, влажный и стабильный. Повсеместно распространились леса и речные заливные луга.
 В раннем мелу начала разрушаться Гондвана. Расширилась рифтовая долина между Южной Америкой и Африкой. Уровень моря упал, и Европейские острова увеличились, а Обский бассейн уменьшился. В мезозое северный и южный полюса находились в зонах, связанных морскими течениями с теплыми районами, поэтому общий климат был теплым. В позднем мелу Пангея разрушилась, уровень моря поднялся необычно высоко – воду вытеснили высокие океанические хребты. Между Африкой и Южной Америкой стремительно расширялся Атлантический океан (увеличивается он и сейчас). В Северной Америке образовалось море Ниобрара, вся Европа находилась под водой, кроме Скандинавии, и поперек Африки тоже разлилось море. Циркуляция воды и воздуха изменилась, климат стал прохладнее и зональнее. Раздвижение континентов привело к изоляции наземных экосистем. Австралия и Антарктида отделились друг от друга, первая отправилась на север, а вторая – к южному полюсу. В кайнозое Антарктида покрылась льдами и сохранила жизнь лишь на побережье. Климат Австралии становился все более засушливым (она и сейчас движется к экватору), что привело к вымиранию в конце третичного – начале четвертичного периода многих животных, связанных с лесными экосистемами. Несмотря на мозаичность, в которой выражается неполнота летописи, в целом можно сказать, что, чем дальше вглубь времен, тем меньше нам известно. О том, как вообще на Земле появилась жизнь, точных данных нет. Гипотезы есть следующие: Креационизм – творение со стороны некоего духовного начала – высшего интеллекта, то есть Бога. Впрочем, если считать, что Бог – это совокупность всех объективно действующих законов мироздания, то есть что он – везде и во всем, то эта гипотеза не противоречит любым другим. Панспермия – занесение жизни извне. Прямых доказательств этому нет, но, учитывая размеры Вселенной, представляется практически невероятным, чтобы жизнь существовала только на Земле. Сомнительные, но следы жизни найдены недавно на Марсе. Теоретически некоторые споры бактерий и вирусы могли бы вынести путешествие в космосе. Косвенно в пользу этой гипотезы говорит то, что расчеты показывают недостаточность предполагаемого времени существования Земли для возникновения и развития жизни на ней путем случайных мутация. Возникновение жизни на Земле, согласно мнению А. Опарина и Дж Холдейна.
В раннем мелу начала разрушаться Гондвана. Расширилась рифтовая долина между Южной Америкой и Африкой. Уровень моря упал, и Европейские острова увеличились, а Обский бассейн уменьшился. В мезозое северный и южный полюса находились в зонах, связанных морскими течениями с теплыми районами, поэтому общий климат был теплым. В позднем мелу Пангея разрушилась, уровень моря поднялся необычно высоко – воду вытеснили высокие океанические хребты. Между Африкой и Южной Америкой стремительно расширялся Атлантический океан (увеличивается он и сейчас). В Северной Америке образовалось море Ниобрара, вся Европа находилась под водой, кроме Скандинавии, и поперек Африки тоже разлилось море. Циркуляция воды и воздуха изменилась, климат стал прохладнее и зональнее. Раздвижение континентов привело к изоляции наземных экосистем. Австралия и Антарктида отделились друг от друга, первая отправилась на север, а вторая – к южному полюсу. В кайнозое Антарктида покрылась льдами и сохранила жизнь лишь на побережье. Климат Австралии становился все более засушливым (она и сейчас движется к экватору), что привело к вымиранию в конце третичного – начале четвертичного периода многих животных, связанных с лесными экосистемами. Несмотря на мозаичность, в которой выражается неполнота летописи, в целом можно сказать, что, чем дальше вглубь времен, тем меньше нам известно. О том, как вообще на Земле появилась жизнь, точных данных нет. Гипотезы есть следующие: Креационизм – творение со стороны некоего духовного начала – высшего интеллекта, то есть Бога. Впрочем, если считать, что Бог – это совокупность всех объективно действующих законов мироздания, то есть что он – везде и во всем, то эта гипотеза не противоречит любым другим. Панспермия – занесение жизни извне. Прямых доказательств этому нет, но, учитывая размеры Вселенной, представляется практически невероятным, чтобы жизнь существовала только на Земле. Сомнительные, но следы жизни найдены недавно на Марсе. Теоретически некоторые споры бактерий и вирусы могли бы вынести путешествие в космосе. Косвенно в пользу этой гипотезы говорит то, что расчеты показывают недостаточность предполагаемого времени существования Земли для возникновения и развития жизни на ней путем случайных мутация. Возникновение жизни на Земле, согласно мнению А. Опарина и Дж Холдейна.
 Так или иначе, где бы ни возникла жизнь, как-то она должна была возникнуть. Чем же живое отличается от неживого? Живые системы – это организованные системы, поддержание которых зависит от программ, репликации и открытости. Они потребляют пищевые ресурсы из окружающей среды и используют их так, что это обеспечивает их выживание и плодовитость. Организмы запрограммированы. Этим программам подчинены рабочие системы, которые открыты для поступления вещества и энергии. Упорядоченные системы организмов постоянно подвергаются атакам энтропии, но их разрушающиеся компоненты могут замещаться в соответствии с программой, то есть организмы должны быть открытыми системами, поглощать материал и энергию, выделять отходы и лишнюю энергию. Эволюция путем естественного отбора – это неизбежное следствие возобновления систем за счет репликации. Геном не всегда реплицируется точно. Изменчивость генетической программы влечет за собой изменчивость фенотипов, и программы, лучше всего обеспечивающие выживание и плодовитость в данной среде, станут встречаться чаще. Поскольку мир конечен и ресурсы ограничены, эти программы будут иметь тенденцию замещать другие, менее выигрышные. Приспособленность представляет собой свойство одного генетически детерминированного признака распространяться в популяции за счет других, а это требует как выживание, так и плодовитости. Главный вопрос – как молекулы организовались в самореплицирующиеся системы? О первоначальной земной атмосфере известно мало, но, возможно, она возникла в ходе дегазации Земли и была похожа на газы, выделяемые вулканами и нынче. Если это так, то в ней не было кислорода. Эксперименты показали, что в таких условиях любое энергетическое воздействие – освещение, ударные волны, радиация или тепло, - привело к синтезу разнообразных органических веществ: сахаров, аминокислот и даже нуклеотидов. При благоприятных обстоятельствах, например, при высокой концентрации неорганических полифосфатов, они могут полимеризоваться. Все эти вещества, вероятно, накапливались в древнем океане, образуя “первичный бульон”. В этом добиологическом мире тоже шел отбор, так как молекулы, полимеризовавшиеся быстрее, и/или оказавшиеся более стабильными, эффективнее поглощали исходные вещества, и получали большее распространение. Это вряд ли могло привести к возникновению принципиально новых структур, так как образование каждого мономера происходило независимо и не фиксировалось в генетической памяти. Но некоторые полимеры, однажды образовавшись, влияют на образование других. Полинуклеотиды определяют последовательность нуклеотидов, действуя как матрица, для полимеризации. Если один полинуклеотид служит матрицей для комплементарного ему другого, и т. д. , то уже можно говорить о генетической памяти, а полинуклеотиды, воспроизводящиеся наиболее эффективно, имеют селективное преимущество, конкурируя с другими за ограниченное количество мономерных блоков. При нормальной температуре такая реакция протекала бы медленно и со множеством ошибок, но включение в процесс белка – каталазы ее бы ускорило. Как это произошло, неясно, возникнув, такая система получила преимущество. Должны быть преимущества и от окружения матрицы и репликазы оболочкой, чтобы выгодой от их взаимодействия не могли воспользоваться другие матрицы – конкуренты. Мембрана клетки, представляющая собой двойной слой фофсолипидов, могла возникнуть при обратном падении капли “бульона”. Те примитивные клетки, у которых взаимодействие между генотипом и фенотипом стимулировало скорость репликации и ее точность, распространялись быстрее других. При этом почти наверняка эукариоты – это результат эндосимбиоза (или неудачного поедания друга) прокариот. Почему сейчас на Земле не происходит зарождения жизни? Потому, что накопившийся кислород делает невозможным существование “первичного бульона”.
Так или иначе, где бы ни возникла жизнь, как-то она должна была возникнуть. Чем же живое отличается от неживого? Живые системы – это организованные системы, поддержание которых зависит от программ, репликации и открытости. Они потребляют пищевые ресурсы из окружающей среды и используют их так, что это обеспечивает их выживание и плодовитость. Организмы запрограммированы. Этим программам подчинены рабочие системы, которые открыты для поступления вещества и энергии. Упорядоченные системы организмов постоянно подвергаются атакам энтропии, но их разрушающиеся компоненты могут замещаться в соответствии с программой, то есть организмы должны быть открытыми системами, поглощать материал и энергию, выделять отходы и лишнюю энергию. Эволюция путем естественного отбора – это неизбежное следствие возобновления систем за счет репликации. Геном не всегда реплицируется точно. Изменчивость генетической программы влечет за собой изменчивость фенотипов, и программы, лучше всего обеспечивающие выживание и плодовитость в данной среде, станут встречаться чаще. Поскольку мир конечен и ресурсы ограничены, эти программы будут иметь тенденцию замещать другие, менее выигрышные. Приспособленность представляет собой свойство одного генетически детерминированного признака распространяться в популяции за счет других, а это требует как выживание, так и плодовитости. Главный вопрос – как молекулы организовались в самореплицирующиеся системы? О первоначальной земной атмосфере известно мало, но, возможно, она возникла в ходе дегазации Земли и была похожа на газы, выделяемые вулканами и нынче. Если это так, то в ней не было кислорода. Эксперименты показали, что в таких условиях любое энергетическое воздействие – освещение, ударные волны, радиация или тепло, - привело к синтезу разнообразных органических веществ: сахаров, аминокислот и даже нуклеотидов. При благоприятных обстоятельствах, например, при высокой концентрации неорганических полифосфатов, они могут полимеризоваться. Все эти вещества, вероятно, накапливались в древнем океане, образуя “первичный бульон”. В этом добиологическом мире тоже шел отбор, так как молекулы, полимеризовавшиеся быстрее, и/или оказавшиеся более стабильными, эффективнее поглощали исходные вещества, и получали большее распространение. Это вряд ли могло привести к возникновению принципиально новых структур, так как образование каждого мономера происходило независимо и не фиксировалось в генетической памяти. Но некоторые полимеры, однажды образовавшись, влияют на образование других. Полинуклеотиды определяют последовательность нуклеотидов, действуя как матрица, для полимеризации. Если один полинуклеотид служит матрицей для комплементарного ему другого, и т. д. , то уже можно говорить о генетической памяти, а полинуклеотиды, воспроизводящиеся наиболее эффективно, имеют селективное преимущество, конкурируя с другими за ограниченное количество мономерных блоков. При нормальной температуре такая реакция протекала бы медленно и со множеством ошибок, но включение в процесс белка – каталазы ее бы ускорило. Как это произошло, неясно, возникнув, такая система получила преимущество. Должны быть преимущества и от окружения матрицы и репликазы оболочкой, чтобы выгодой от их взаимодействия не могли воспользоваться другие матрицы – конкуренты. Мембрана клетки, представляющая собой двойной слой фофсолипидов, могла возникнуть при обратном падении капли “бульона”. Те примитивные клетки, у которых взаимодействие между генотипом и фенотипом стимулировало скорость репликации и ее точность, распространялись быстрее других. При этом почти наверняка эукариоты – это результат эндосимбиоза (или неудачного поедания друга) прокариот. Почему сейчас на Земле не происходит зарождения жизни? Потому, что накопившийся кислород делает невозможным существование “первичного бульона”.