Prezentatsia_BGTs_2_kurs_Ekologi.ppt
- Количество слайдов: 111



Курс Общей экологии

Структура курса Блоки курса «Общая экология» • Введение в курс «Общая экология» • Экология организмов (аутэкология) • Экология популяций (демэкология) • Биогеосистемная экология

Структура курса «Общая экология» По семестрам Часов 2 семестр 3 семестр 4 семестр Зач Экз Курс Все Ауд. ет аме раб. го занят ы ны ий Самост. работа Лек ЛПЗ ций 2 100 36 3 9 200 18 Полевая практика (зачет) 16 30 Полевая практика (зачет)

Биогеосистемная экология 2 курс Экологи

Биогеосистемная экология Вопросы Исторический экскурс в становление биогеоценологической мысли в России и за рубежом l Соотношение понятий экосистема, биогеоценоз, биогеосистема l Современное представление о пространственно-временной структуре биогеоценоза l Функиональная организация биогеоценоза l Динамика биогеоценозов: а) цикличные (обратимые) изменения - сезонные - флуктуации б) сукцессионные (необратимые) изменения - автогенные сукцессии - аллогенные сукцессии в) климаксы (концепции, типы, критерии) l Планетарная роль биогеоценотического покрова l

1. Исторический экскурс в становление биогеоценологической мысли в России и за рубежом Ж. Кювье, 1801 (определение понятия «жизнь» ) l Ч. Р. Дарвин (1837, 1881) l Э. Геккель, 1866, 1868 l К. Мебиус, 1877 – биоценоз l С. Форбс, 1887 – биоценотический микрокосм l К. Ф. Рулье «Сомнения в зоологии» , 1848 – урочище l

Степи и леса как база развития биогеоценологической мысли в России l Ю. К. Пачоски (1864 -1942) – фитосоциология l В. В. Докучаев, 1892 – «Наши степи прежде и теперь» l Г. Ф. Морозов (1867 -1920), «Учение о лесе» Георгий Федорович Морозов l Василий Васильевич Докучаев П. Н. Крылов, 1898 - «Тайга с естественно-исторической точки зрения как общественное явление»

Оформление биогеоценологического знания и сознания l Артур Тенсли, 1935 – Экосистема l Р. Линдеманн, 1942 l В. Н. Сукачев, 1940, 1942 l Н. В. Дылис, 1978 l Ю. Одум, 1986 Артур Тенсли Владимир Николаевич Сукачев
, * биосистема (Thienemann,")
Многоликость понятий В зарубежной литературе предложены понятия: * экосистема (Tensley, 1935), * биосистема (Thienemann, 1941), * холоцен (Friederichs, 1930) и др. l В отечественной литературе: * эпиморфа (Аболин, 1914), * биогеоценоз (Сукачев, 1942), * биоэкос (Нестеров, 1954) и др. l l Самые распространенные понятия "биогеоценоз" и "экосистема". Эти два понятия на 1 Х международном ботаническом конгрессе в 1959 г. в Канаде были признаны синонимами, однако различие в них все же есть. Объединяет их общая биокосная природа.

2. Соотношение понятий экосистема, биогеоценоз, биогеосистема l Экосистема Биогеоценоз биоценоз ЗЦ биосистема физиосистема ФЦ эдафотоп Микросистема мезосистема макросистема (Дювиньо, Танг, 1973) МЦ климатоп экотоп Биогеосистемы (Бяллович, 1960 )

Биогеосистема Ландшафт вещество, энергия, информация БГЦ вещество, энергия, информация дрифт БГЦ вещество, энергия, информация Биогеосистемы (Бяллович, 1960 )
 А. Пространственно-временная структура биогеоценозов")
2. Структура биогеоценозов (синморфология) А. Пространственно-временная структура биогеоценозов
 l Напочвенный (подстилка, войлок) l Почвенный")
Экологические ярусы l Надпочвенный(геоботанические ярусы с их обитателями) l Напочвенный (подстилка, войлок) l Почвенный
 Видовая структура l l l Эдификаторы Доминанты Сопутствующие К. Раункиер Кривая Раункиера число")
а) Видовая структура l l l Эдификаторы Доминанты Сопутствующие К. Раункиер Кривая Раункиера число видов 10 300 600 численность
 Биологическая структура * Спектры жизненных форм, % (Уиттекер, 1980) Биом Фанерофи ты Хамефиты")
б) Биологическая структура * Спектры жизненных форм, % (Уиттекер, 1980) Биом Фанерофи ты Хамефиты Гемикрипт Геофиты Терофиты (криптофи ты) Тропическ ий лес 96 2 0 Лес умеренной зоны 10 17 54 12 7 Тундра 1 22 60 15 2 Степь 1 12 63 10 14 Пустыня 0 4 17 6 73 53 0 Травяные 0 0 47 болота* *по: Сеняниновой-Корчагиной М. В. , 1949

Географические закономерности распределения жизненных форм Пояс Климат Преобладающая жизненная форма Тропический Жаркий, влажный фанерофиты Умеренный, гемикриптофиты континентальный Субарктический Холодный, арктический хамефиты
 Промежуточные группы: Гигромезо-")
* Спектры экологических групп ( по отношению к l Гидрофиты воде) Промежуточные группы: Гигромезо- l Гигрофиты Мезогигро. Мезоксеро- l Мезофиты Ксеромезо. Стипаксеро- l Ксерофиты

Спектры экологических групп по отношению к субстрату Участки: l Глинистые эремо l Песчаные псаммо l Каменистые петро - бионты l по отношению к геоботаническому ярусу Деревья дендро- l Кустарники тамно- l Травы хорто- l Подстилка герпето- l Почва гео- бионты

* Спектры фитоценотических групп Лесные l Лугово-лесные l Степные l Лугово-степные l Болотные l Лугово-болотные l Пустынные l Долевое участие (%) ФЦГ в сложении фитоценоза – отражает динамическое состояние ценоза

* Спектры фитоценотипов Раменский Л. Г. , 1938 l Фитоценотипы – группы организмов, различающиеся по особенностям их отношений к другим компонентам и способности к совместному произрастанию в фитоценозах Фитоценотипы l Виоленты (силовики), «львы» еловый лес Патиенты Эксплеренты (выносливцы) (выполняющие) «верблюды» «шакалы» кочедыжник кислица первоцветы на www. botsad. ru

Вертикальная структура Еловый лес умеренных широт степь континуальность луг
 зоогенная l Мозаичность l фитогенная")
Горизонтальная структура Синузии (Х. Гамс, 1918) зоогенная l Мозаичность l фитогенная

Фитогенное поле А. А. Уранов Фитогенное поле – территория, в пределах которой отдельное растение оказывает воздействие на условия своего существования (Уранов, 1965)

Модуль 4. Биотические отношения l Биотические отношения – это прямые или косвенные взаимоотношения организмов между собой, осуществляющихся на внутри- или межвидовом уровнях. Связи на трофической основе Автотрофы Фитофаги Зоофаги Микробы-редуценты Паразиты Сверхпаразиты
 Выделяются высшие растения Действуют на…. колины высшие")
Связи на химической основе Аллелопатия (Молиш, 1937) Выделяются высшие растения Действуют на…. колины высшие растения фитонциды маразмины микроорганизмы антибиотики микроорганизмы феромоны животные телергоны животные
 Эффект группы")
Значение внутригрупповых отношений l l Трофика Панмиксия Эффект плотности (Чэпмен, биологический потенциал) Эффект группы (Грэссе, Чэйвин, 1944) В зоологии: В ботанике: Уваров, А. А. Уранов И. В. Стебаев Титов • Фитогенное поле (Уранов, 1965)

по: В. Н. Беклемишеву Биотические отношения форические трофические топические фабрические симбиотические антибиотические нейтрализм (0 0) l Мутуализм (+ +) Хищничество(+ -) Протокооперация (+ +) Паразитизм (+ -) Комменсализм (+ 0) Каннибализм (+ -) Конкуренция (- -) Аменсализм (- 0) «Рабовладельчество» (+ -)

Форические отношения Зоохория – перенос животными семян, спор, пыльцы растений. [Энциклопедия «В мире дикой природы» . , 2006 ] Блоха крысиная Форезия – перенос животными более мелких животных. [Энциклопедия «В мире дикой природы» . , 2006 ] Мышь - полевка

Фабрические связи Это такой тип биоценотических отношений, в которые вступает вид, используя для своих сооружений (фабрикации) продукты выделения или мертвые остатки, или даже живых особей другого вида, а также один вид может использовать другой вид для строительства жилья, [Cтепановский А. С. , 2001] гнезд, убежищ и т. п. [Энциклопедия «В мире дикой природы» . , 2006 ] Грач использует ветви и стебли травы для строительства гнезда [Энциклопедия «В мире дикой природы» . , 2006 ] Большая синица использует дупло в стволе дерева и перо и пух для строительства гнезда

Топические связи один вид создает среду для другого формируют субстрат, на котором поселяются или, наоборот, избегают селиться представители других видов влияют на движение воды, воздуха, изменение температуры, освещенности окружающего пространства насыщают среду продуктами выделения и т. п. Топические связи – характеризуют любое, физическое или химическое изменение условий обитания одного вида в результате жизнедеятельности другого.

Топические отношения морской желудь и кожа кита лишайник на стволе дерева

Трофические отношения А. Симбиотические отношения: мутуализм, комменсализм, протокооперация, . Мутуализм - это форма облигатного симбиоза, при которой организмы двух и более видов возлагают друг на друга регуляцию своих отношений с внешней средой и не могут жить самостоятельно.
![Мутуализм [Серия «Эрудит» . Мир животных. , 2006] [Машанова О. Г. , Евстафьев В.](https://present5.com/presentation/138345737_41907021/image-33.jpg "Мутуализм [Серия «Эрудит» . Мир животных. , 2006] [Машанова О. Г. , Евстафьев В.")
Мутуализм [Серия «Эрудит» . Мир животных. , 2006] [Машанова О. Г. , Евстафьев В. В. , 1997] Термиты вырабатывают фермент глюкозидазу, переводящий клетчатку древесины в сахара [Энциклопедический словарь юного биолога. , 1986 ] Биотические отношения Жгутиконосец в кишечнике термита

комменсализм Комменсализм – одностороннее использование одного вида другим без нанесения ему вреда. Рыбы-прилипалы, присосавшиеся к телу черепахи Рыбы амфиприоны плавают среди жгучих щупалец актинии
![Комменсализм [Резник Дж. П. , 1995] Тигровая акула и рыба-прилипала Средиземноморский карапус Это форма](https://present5.com/presentation/138345737_41907021/image-35.jpg "Комменсализм [Резник Дж. П. , 1995] Тигровая акула и рыба-прилипала Средиземноморский карапус Это форма")
Комменсализм [Резник Дж. П. , 1995] Тигровая акула и рыба-прилипала Средиземноморский карапус Это форма симбиоза, при которой взаимоотношения между организмами двух видов, когда один из партнеров системы (комменсал) возлагает на другого (хозяина) регуляцию своих отношений с внешней средой, но не вступает с ним в тесные отношения. [Машанова О. Г. , Евстафьев В. В. , 1997] Голотурии Обитают в полости тела; используют как убежище [Энциклопедия «В мире дикой природы» . , 2006 ]
 Квартирантство")
Виды комменсализма: Нахлебничество потребление остатков пищи хозяев: (лев и гиена, волк и шакал) Квартирантство Использование убежищ либо в постройках, либо на телах других видов (Водная черепаха и рыба-прилипала)

Протокооперация Это форма симбиоза, при которой совместное существование организмов выгодно обоим видам, но не обязательно для них. [Энциклопедия «В мире дикой природы» . , 2006 ] Актиния и рак - отшельник [Энциклопедический словарь юного биолога. , 1986 ] Морские ежи и рыбы чистильщики

Протокооперация Муравей и тля Рак отшельник и актиния

Б. Антибиотические отношения Антибиоз - это различные формы совместного существования организмов при которой, обе взаимодействующих популяции или одна из них испытывают отрицательное влияние. Отрицательные взаимоотношения могут принимать следующие формы: хищничество, паразитизм, конкуренция, каннибализм. Это форма антибиоза при которой, межвидовые отношение связанны с активным поиском и энергичными способами овладения сопротивляющейся и убегающей добычей. Хищничество [Машанова О. Г. , Евстафьев В. В. , 1997] [Резник Дж. П. , 1995] Североамериканская красная рысь на охоте Хищничество встречается во всех группах эукариотических организмов.
![Хищничество [Вилли К. А. , 1985] Орел с пойманной добычей Жаба с пойманным дождевым](https://present5.com/presentation/138345737_41907021/image-40.jpg "Хищничество [Вилли К. А. , 1985] Орел с пойманной добычей Жаба с пойманным дождевым")
Хищничество [Вилли К. А. , 1985] Орел с пойманной добычей Жаба с пойманным дождевым червем [Энциклопедия «В мире дикой природы» . , 2006 ] Сова с полевкой

По способу охоты хищников делят на: Подстерегателей Преследователей Полярные волки на охоте [Резник Дж. П. , 1995] Леопард на охоте Хищничеству принадлежит важная роль в регуляции численности организмов более низкого трофического уровня, что способствует сохранению их высокого [Машанова О. Г. , Евстафьев В. В. , 1997] видового разнообразия.

Взаимодействие между хищниками и их жертвами приводит к тому, что эволюция хищников и жертв происходит сопряжено, в процессе ее хищники совершенствуют способы нападения, а жертвы – способы защиты. [Энциклопедия «В мире дикой природы» . , 2006 ] Грифы [Энциклопедия «В мире дикой природы» . , 2006 ] Гиены Встречаются также переходы между хищничеством и питанием трупами [Машанова О. Г. , Евстафьев В. В. , 1997] животных (некрофагия).

Каннибализм Богомол самец и самка H H Балхашский окунь Самец американской саламандры ttp: //fishing. kz/index. p

Конкуренция - это форма антибиоза при которой, взаимоотношения между особями или популяциями одного и того же вида или разных видов соревнующимися за одни и те же ресурсы внешней среды при их ограниченном количестве. [Вилли К. А. , 1985] Морские слоны в борьбе за территорию [Большая иллюстрированная энциклопедия животных. , 2004] Гиены прогнали гепарда и приступают к трапезе
 p: //www. deltanewsweb. com/news/archives. html p: //www. deltanewsweb. com/news/ http: //www. photosight.")
Конкуренция (скворцы) p: //www. deltanewsweb. com/news/archives. html p: //www. deltanewsweb. com/news/ http: //www. photosight. ru/photos/2102590/ http: //www. photosight. ru/ photos/2102590/

Паразитизм Если представители одного вида обитают внутри или на поверхности тела другого и используют его вещество и энергию, то такой тип взаимоотношения называют паразитизмом. [Вилли К. А. , 1985] Присосавшись к жертве, пиявка прокусывает ее кожу и высасывает тканевую жидкость [Энциклопедия «В мире дикой природы» . , 2006 ] [Энциклопедический словарь юного биолога. , 1986 ] Спорынья

Паразитизм известен на всех уровнях организации живого Типичные паразиты птиц [Энциклопедический словарь юного биолога. , 1986 ] Паразиты принимают участие в регуляции численности хозяев, а иногда определяют направленность микроэволюционных процессов. [Энциклопедический словарь юного биолога. , 1986 ]

Характерная особенность паразитов - редукция у них одних систем органов (напр. , пищеварительной системы, органов чувств, конечностей) и усложнение других (половой системы, органов прикрепления) [Энциклопедический словарь юного биолога. , 1986 ] [Энциклопедия «В мире дикой природы» . , 2006 ] [Энциклопедический словарь юного биолога. , 1986 ] Ленточный червь - солитер

В. Нейтрализм Если ни одна популяция не испытывает влияние другой, т. е. между ними нет взаимодействия, то ситуация именуется нейтрализмом. Пеликаны и бегемоты Зебры и бородавочники

Г. Амменсализм l Для одного из двух взаимодействующих видов последствия взаимодействия отрицательны, а для другого – безразличны. (г. Новосибирск, Первомайский сквер, 2008 г. , фото Глебовой А. Е. )

Оптимумы вида Физиологический Синэкологический Это благоприятное для вида сочетание всех абиотических факторов, при котором возможны наиболее быстрые темпы роста и размножения Это такое биотическое окружение, при котором вид испытывает наименьшее давление со стороны врагов, конкурентов, что позволяет ему успешно размножаться
 Физиологический и синэкологический оптимумы не всегда совпадают. Например, массовое")
(Чернова Н. М. , 2) Физиологический и синэкологический оптимумы не всегда совпадают. Например, массовое размножение вредителя зерновых культур, гессенского комарика, после особенно суровых зим, которые должны неблагоприятно сказываться на численности этого насекомого; нашествие саранчи. гессенский комарик рожь саранча

Концепции экологической ниши
 Одним из")
1. История формирования понятия экологическая ниш Пространственная ниша Дж. Гринелла (Grinnell, 1917) Одним из первых термин "ниша" употребил Дж. Гриннел (1917 год). Гриннел считал нишу поведенческой единицей и подчеркивал, что она определяет распределение вида в пространстве, т. е. Гриннел трактовал нишу как пространственную единицу. Джозеф Гринелл Трофическая ниша Ч. Элтона (Elton, 1927) Нишу он понимал как "функциональный статус" вида в сообществе, т. е. что "делает" вид в сообществе. Так по Ч. Элтону, под нишей животного разумеется образ жизни и в особенности способ питания. Другими словами "экологическая ниша" Элтоном трактуется как трофическая ниша. Чарльз Элтон
: По Е. Одуму, экологическая ниша -")
Юджин Одум Eugene P. Odum (1959 г. ): По Е. Одуму, экологическая ниша - это "положение, или статус, организма в сообществе и экосистеме, вытекающее из его структурных адаптаций, физиологических реакций и специфического поведения. " Одум подчеркивает, что "экологическая ниша зависит не только от того, где он живет, но и от того, что он делает". По его образному выражению "экологическая ниша - это адрес и профессия вида". Роберт Уиттекер Robert H. Whittaker (1980 г. ): «. . . ниша – это термин, употребляемый для обозначения специализации популяции вида внутри сообщества» .
: «. . . ниша экологическая – место вида в природе,")
Николай Федорович Реймерс (1990): «. . . ниша экологическая – место вида в природе, включающее не только положение вида в пространстве, но и функциональную роль его в сообществе (например, трофический статус) и его положение относительно абиотических условий существования (температуры, влажности и т. п. )» . Понятие «экологическая ниша» следует отличать от понятия «местообитание» . Под местообитанием подразумевается та часть пространства, которая заселена видом и которая обладает необходимыми абиотическими условиями для его существования. Экологическая ниша вида зависит не только от абиотических условий среды, но и в не меньшей мере от его биоценотического окружения.
 Д.")
2. Развитие представлений о многомерной нише Многомерная экологическая ниша Дж. Хатчинсона (Hutchinson, 1957) Д. Хатчинсон (1957) определил экологическую нишу как многомерное пространство или гиперобъем, в пределах которого условия среды позволяют неопределенно длительное время существовать особи или виду. Хатчинсон не включает в понятие ниша поведенческую характеристику особи (вида). Ниша - это прежде всего место в пространстве, подобное микроместообитанию (стации). Джордж Хатчинсон Рис. Модель двумерной (а) и трехмерной экологической ниши (b); Fi – отдельные факторы пространства экологических ниш
 (реализованная) Жизненная стратегия")
Экологическая ниша – адрес и профессия вида Ниша Фундаментальная Функциональная (потенциальная) (реализованная) Жизненная стратегия = ПН - РН
 который можно представить как \"протяженность гиперпространства\"")
Мерность ниш – это ширина ниши (ее размер) который можно представить как "протяженность гиперпространства" реализованной (функциональной) ниши. Специализация вида по питанию, использованию пространства, времени активности и другим условиям характеризуется как сужение его экологической ниши, обратные процессы — как ее расширение. Примеры узкой и широкой экологической ниши Узкую нишу занимает панда Широкую нишу занимает бурый медведь

Внутривидовая конкуренция и межвидовая конкуренция по-разному влияют на ширину ниши. Внутривидовая конкуренция ведёт к расширению ниши, а межвидовая к её сужению.

Взаимодействие ниш Две фундаментальные ниши могут быть идентичными, полностью соответствуя другу. Такая экологическая ситуация мало вероятна. Если два вида занимают идентичную экологическую нишу (в данном случае хищники), то более конкурентноспособный вид вытеснит менее конкурентноспособный, или же максимум численности одного вида будет приходиться на минимум численности другого. Когда ниша вида А 1 находится внутри ниши вида А 2 возможно два исхода: а) Если вид А 2 обладает превосходством (прерывистая линия), то он будет сосуществовать с видом А 1 при неполном использовании общих ресурсов; б) Если превосходством обладает вид А 1 (сплошная линия), то он будет использовать весь градиент ресурсов, а вид А 2 будет вытеснен.

Две фундаментальные ниши могут перекрываться не полностью, когда часть нишевого пространства занята обеими видами, а другая часть - только одним. В этом случае каждый вид имеет "убежище" в виде неоспариваемой части нишевого пространства и неизбежно сосуществование, при котором доминирующий вид занимает зону перекрывания ниш. Возможно два варианта: а) Перекрывание ниш равной ширины. Конкуренция одинакова в обоих направлениях. b) Перекрывание ниш неравной ширины. Конкуренция неодинакова в двух направлениях, т. к. часть нишевого пространства, входящая в область перекрывания у вида А 2 больше, чем у вида А 1. Таким образом, экологическую нишу, действительно, следует рассматривать как пространственнофункциональную единицу. Многоаспектный характер взаимодействия экологических нищ обусловливает широкую экологическую лабильность вида.

Сформулированное Г. Ф. Гаузе правило конкурентного исключения для близких по экологии видов может быть выражено таким образом, что два вида не уживаются в одной экологической нише. Выход из конкуренции достигается расхождением требований к среде, изменению образа жизни Механизмы выхода из конкуренции и разграничения экологических ниш при этом следующие: Ш Специализация морфологии и поведения в соответствии с родом пищи; Ш Пространственная дифференциация. Вертикальное и горизонтальное разделение; Ш Различия во времени активности. Пищевые угодья у различных видов синиц (по Е. А. Криксунову и др. , 1995)

Правило обязательности заполнения экологической ниши. Пустующая экологическая ниша всегда и обязательно бывает естественно заполнена. В насыщенных биогеоценозах жизненные ресурсы используются наиболее полно - в них все экологические ниши заняты. В ненасыщенных биогеоценозах жизненные ресурсы утилизируются частично, для них характерно наличие свободных экологических ниш. Рис. Избирательное использование травяного покрова животными А, Б—саванна, В — степь
 Виды, объединенные в сообщество (экосистему),")
ПРИНЦИП "ПЛОТНОЙ УПАКОВКИ" ЭКОЛОГИЧЕСКИХ НИШ Мак-Артура (дифференциации экологических ниш) Виды, объединенные в сообщество (экосистему), стремятся использовать все возможности для существования, представляемые средой и биотическим окружением, и максимизировать биопродуктивность в конкретном биотопе. Роберт Мак-Артур Robert H. Mc. Arthur (1930 -1972) Рис. Схема, иллюстрирующая процесс внедрения в сообщество новых видов и их плотной упаковки
, «. . . если на градиент ресурса мы добавим")
Как подчеркивает Р. Уиттекер (1980), «. . . если на градиент ресурса мы добавим другой вид, ширина ниш уже имеющихся здесь видов будет сокращена. . , что позволяет сделать некоторые заключения: • виды эволюционируют в направлении специализации к разным частям градиента ресурса, что уменьшает конкуренцию между ними; • в любой период виды стремятся к равномерному распределению адаптивных центров по градиенту; • последовательности ниш могут удлиняться за счет внедрения новых видов между старыми, что сокращает ширину ниш ранее существовавших видов; • имеются, вероятно, пределы количества видов, которые могут, таким образом, "упаковываться" в последовательность видов вдоль данного градиента» .

Экологические ниши видов изменчивы в пространстве и во времени.

Экологическое дублирование - занятие освободившейся экологической ниши другим видом, способным выполнять в сообществе те же функции, что и исчезнувший вид. При экологическом дублировании предполагается, что исчезнувший из сообщества вид замещается (занимает экологическую нишу) экологически эквивалентным. Экологическими эквивалентами могут быть даже не близкородственные таксономически виды. Кактус калифорнийский Молочай блестящий ( Cylindropuntia californica ) ( Euphorbia fulgens)

Функциональная организация экосистем

l Функциональные группы l Трофические уровни – группа организмов одного типа питания l Трофические цепи – передача вещества и энергии от звена производителя к звену потребителей пастбищные l Трофические цепи детритные начинаются с зеленого растения начинаются с отторгнутого орг. вещества или трупа

Функциональные нагрузки подсистем БГЦ l Растительная подсистема: * первичный синтез биомассы из неорганических веществ * создание исходного негэнтропийного материала * системообразующая роль Животная подсистема: Консументы Редуценты l *Прогрессивное преобразование биомассы *Повышение оструктуренности * Санация Вырабатывают энергию в себе * Утилизация * Зацикливание круговорота *Повышение негэнтропии • Человеческая подсистема: вырабатывает энергию машинами для себя, прессируя естественные БГЦ

Перенос потенциальной энергии пищи, созданной растениями, через ряд организмов путем поедания одних видов другими называется цепью питания, или пищевой цепью, а каждое ее звено - трофическим уровнем. Поток энергии через типичную пищевую цепь

Пастбищная трофическая цепь п к 1 к 2 к 3 к 4 Трава кузнечик зяблик уж сова 100% 10 1 0. 01 к 5 R жук-мертвоед бактерии 0. 001 0. 0001 КПД(ф) = 0. 1 - 3% Закон 10% !!!!

Примеры типов пищевых цепей

http: //tana. ucoz. ru/load/345 -2 -2 http: //www. vistavoda. ru/clearvod. php http: //www. labirint. ru/news/3776/

Пищевые сети. Пищевая сеть и направление потока вещества в водных экосистемах Пищевая сеть и направление потока вещества в наземных экосистемах (по Е. А. Криксунову и др. , 1995)

Пирамиды: * численности * биомассы * энергии
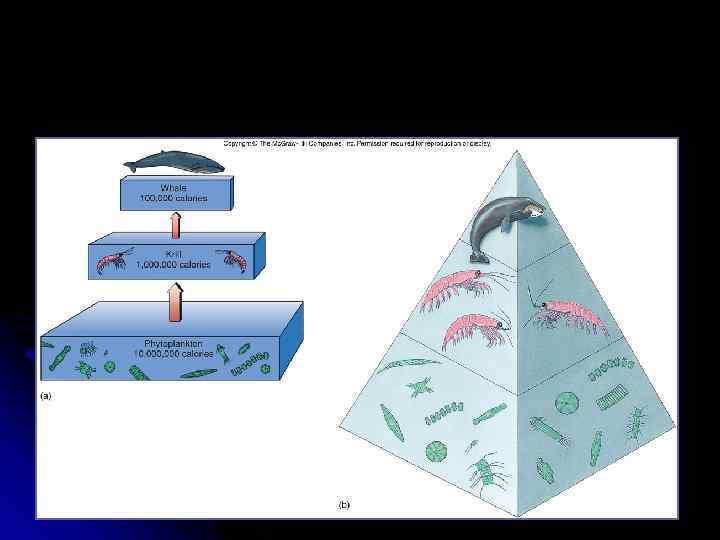

Слева изображена прямая пирамида биомасс, справа – перевёрнутая Пример сезонного изменения в пирамиде биомассы.
 Чистая первичная продуктивность (ЧПП) Вторичная продуктивность ВПП")
Продуктивность экосистемы Валовая первичная продуктивность (ВПП) Чистая первичная продуктивность (ЧПП) Вторичная продуктивность ВПП примерно на 20 % выше, чем ЧПП, так как часть энергии растения тратят на дыхание. Поток энергии через пастбищную пищевую цепь. Все цифры даны в к. Дж на метр в квадрате умноженное на год.
 • Чистая первичная продуктивность (ЧПП) ЧПП= ВПП")
Продуктивность экосистемы • Валовая первичная продуктивность (ВПП) • Чистая первичная продуктивность (ЧПП) ЧПП= ВПП - Д • ВПП примерно на 20 % выше, чем ЧПП, так как часть энергии растения тратят на дыхание. • Вторичная продуктивность
 изменения а) сезонные изменения Общая закономерность: -")
5. Динамика биогеоценозов 1. Цикличные (обратимые) изменения а) сезонные изменения Общая закономерность: - зависимость от температурного и водного режимов - сопровождается сменой аспектов цветения растений в зависимости от биологии видов б) флуктуации Флуктуации (Глизон, 1939) = разногодичная изменчивость (Работнов, 1955) Флуктуации - колебания растительных сообществ около средних значений за определенный период времени, вызванные колебаниями погодных условий от года к году.

Динамика биогеоценозов « Сукцессия должна занимать в экологии такое же место, как эволюция в общей биологии» (Маргалеф) 2. Необратимые изменения (сукцессии) l Каулс (Cowles, 1899) - работы по сукцессиям растительности песчаных дюн на побережье озера Мичиган (США), которые были признаны классическими. Ф. Е. Клементс l Клементс (1904, 1936): разработана стройная теория сукцессий Дал обзор литературы по этому вопросу начиная с 1685 года до 1916 в монографии «Plant succеssion. An analysis of the development of vegetation»
 Аллогенные (вторичные) Автоаллогенные гейтогенез гологенез причины Внутренние причины (эндогенез)")
Классификация сукцессий Сукцессии Автогенные (первичные) Аллогенные (вторичные) Автоаллогенные гейтогенез гологенез причины Внутренние причины (эндогенез) Внешние причины (экзогенез)
 сукцессии l Возникают на: • скальных породах • песчаных дюнах, •")
Автогенные ( первичные) сукцессии l Возникают на: • скальных породах • песчаных дюнах, • лавовых плато • терриконниках Фредерик Клементс Горный Алтай, 3000 м над у. м. , фото Глебовой А. Е. , 2008 г. • конусах выноса оврагов • эродированные почвы • ледниковые щиты • территории освободившиеся от ледника Игорь Васильевич Стебаев Процесс зарастания песков в пустыне
 сукцессии» http: //www. sakhalin. ru/Re gion/korni/sea/fakt 18. htm http: //www.")
«Пионерные (вечно молодые) сукцессии» http: //www. sakhalin. ru/Re gion/korni/sea/fakt 18. htm http: //www. volganet. ru/irj/avo. html? Navigation. Target=ROLES: //portal_content/News http: //yusha. ucoz. ru/forum/5 -9 -9

Тенденции автогенных сукцессий: • Увеличение видового разнообразия • Укомплектовка экологических ниш • Переход от трофических цепей к сетевому распределению вещества и энергии • Увеличение информативной ёмкости системы • Расширение локального БИК • Увеличение продуктивности • Увеличение степени закрытости системы для сингенеза других видов
 сукцессии l распространены значительно шире, чем первичные l происходят там, где растительность")
Аллогенные (вторичные) сукцессии l распространены значительно шире, чем первичные l происходят там, где растительность уже существовала и сформировалась почва l резкое улучшение обеспечения растений элементами минерального питания l увеличение сомкнутости и высоты растительности l вызваны воздействием внешних по отношению к ним факторов, как природных, так и антропогенных l могут быть скоротечными и весьма длительными По причинам они могут быть: - климатогенные - эдафогенные - антропогенные Ярошенко, 1961 Л. Г. Раменский

http: //www. wagner. pp. ru/~vitus/phot o/2004. 09. 11 -ploskoe/ http: //www. geonature. ru/worldgeo/05 -8 -23. htm Сосняк-зеленомошник Сосняк-беломошник http: //www. ecosystema. ru/08 n ature/world/ker/21. htm Сосняк высокотравный

Аллогенные сукцессии l Гейтогенез – сукцессия внутри фитоценоза, вызванная внешними факторами, часто антропогенного характера регрессивные восстановительные демутационные ценакулярные НСО, 2007, фото Глебовой А. Е. Демутационная сукцессия: зарастание берега реки при снижении уровня воды до нормального (после осеннего паводка). постмортальные

Демутационная сукцессия Г. Н. Высоцкий http: //www. booksite. ru/fulltext/mel/eho/voc/3. htm НСО, 2008. фото Глебовой А. Е. Зарастание берега реки при снижении уровня воды до нормального (после осеннего паводка). Тихон Александрович Работнов http: //www. vistavoda. ru/clearvod. php Александр Петрович Шенников Фото с сайта www. yuko-travel. ru

Ценакулярные сукцессии http: //www. speleo. lt/forum/viewtopic. php? p=8403&sid= d 172982 ebf 0 ae 8 a 9591996 b 931 ef 8971 Пастбище после выпаса http: //www. photohost. ru/showpicture. php ? pictureid=78710&galleryid=6467&user= 0&days=&sti= http: //puteshestvenniki. ru/story/id/3301

Постмортальные сукцессии Труп суслика www. migranov. ru http: //pavlyuk. livejournal. com/95768. html Выброшенные на берег водоросли http: //pochemuchek. net/files/01_animals/12_anima ls_plants/them_006/004. html

Регрессивные сукцессии http: //bva. wmsite. ru/fotoalbom/prpark-numto/-27 http: //www. yandex. cc/meteo/63375402894/ http: //www. ya-online. ru/news. php? id=225404
 Растения каждой предыдущей стадии создают")
Модели протекания сукцессий l А. Благоприятствования (освоение скальных пород) Растения каждой предыдущей стадии создают благоприятные условия для последующих. l Б. Толерантности (сукцессия на вырубке) Высокотравье: Иван-чай узколистный, Лабазник вязолистный Минерализация лесной подстилки и корней вырубленных деревьев Злаково-разнотравные ценозы: Вейник, пырей ползучий (элитригия), ежа сборная Вынос ЭМП Мелколистные злаки: Душистый колосок, полевица тонкая, белоус Дальнейшее обеднение сообщества

Модели протекания сукцессий А. Благоприятствования Возникновение термальных карстовых озер, вследствие провалов земли вулканы

l Модели протекания сукцессий В. Модель ингибирования Сукцессия может приостановиться, не достигнув терминальной стадии климакса Сукцессия на вырубках Сукцессия нв гарях: кукушкин лен • Г. Модель нейтральности • Крайне редки; сукцессия протекает как популяционный процесс, а роль взаимодействия популяций очень незначительна • Гроссгейм, 1929

Б. Гологенетические сукцессии l l Это сопряженные изменения растительности ландшафта под влиянием внешних причин. типы природные антропогенные Смена растительности при: * Развитие речной долины * Меандрирование * Смена ландшафтов в геологическом аспекте времени Смена растительности при: *Строительстве гидросооружений *Техногенных изменениях ландшафта
 – Уитфорд, Каулс, 1901 Климакс – устойчивое")
Климаксы l Климакс ( англ. - climax) – Уитфорд, Каулс, 1901 Климакс – устойчивое сообщество, которое находится в равновесном состоянии с условиями абиогенной среды. Концепции климаксов: l Концепция климатического моноклимакса (Хульт, Е. Варминг, Клементс) Ведущий фактор - климат Время протекания - геологически длительный отрезок времени Критерии: *устойчивость *конвергенция *доминантность *климакс как суперорганизм
")
Климаксы l Концепция эдафического поликлимакса ( Николс, 1917; Тенсли, 1935; Селлик, 1960; Кершоу, 1974) Ведущий фактор – почвы Время протекания – относительно быстротечно Критерии: * устойчивость * дискретность
")
Климаксы l Концепция эдафического поликлимакса ( Николс, 1917; Тенсли, 1935; Селлик, 1960; Кершоу, 1974) Ведущий фактор – почвы Время протекания – относительно быстротечно Критерии: * устойчивость * дискретность
 * Дискретность – паттерность (pattern) l")
Климаксы Концепция климакса-континуума, или климаксамозаики (Р. Уиттекер, 1953) * Дискретность – паттерность (pattern) l * Принцип Раменского-Глизона (Раменский, 1924, Глизон, 1926)) Климакс – не остановка, а медленное движение при угасании автогенного компонента сукцессионного тренда, но при продолжающихся процессах микроэволюции и аллогенных реакций на изменение климата. l Концепция «скользящего климакса»

Концепция «скользящего климакса»
 l l l Время генерации доминантов меньше цикла изменений")
Типы климаксов Аклимакс (пример: планктон) l l l Время генерации доминантов меньше цикла изменений условий среды Флуктуации сообществ постоянны Климаксы и серии неразличимы зоопланктон фитопланктон

Типы климаксов Циклоклимакс l l Цикл генерации доминантов совпадает с годичными колебаниями условий среды Климаксы как и серии характеризуются главными доминантами – эфемерами или ранневесенними видами

Типы климаксов Катаклимакс l l Генерация доминантов происходит между катастрофами (пожары, сели, оползни и др. ) Климаксы и серии более менее различимы
 l l l Генерация доминантов длительна, изменения среды незначительны")
Типы климаксов Суперклимакс (например, тундра) l l l Генерация доминантов длительна, изменения среды незначительны Биомасса низка, популяции видов более менее стабильны Климаксы и серии формируются одними и теме же видами, отлична только структура
 l l l Генерация доминантов длительна и их")
Типы климаксов Эуклимакс (пример: лес, тайга) l l l Генерация доминантов длительна и их состав меняется более менее непрерывно в ходе сукцессии Биомасса высокая Серийные сообщества хорошо отличимы от климакса

Критерии климаксов 1. Флористический * Видовое разнообразие * Средообразующая роль доминантов * Повторяемость комбинаций видов в сообществах одного и того же типа климаксов * Укомплектовка ниш * Нормальная половозрелая структура доминантов * Закрытость системы для сингенеза
 Соотношение ежегодного прироста биомассы и мортомассы")
Критерии климаксов l 2. Продукционно-энергетический Уиттекер, Вудвелл (1972) Соотношение ежегодного прироста биомассы и мортомассы равны Маккинтош (1974) Отношение продуктивности к дыханию уменьшается и приближается к единице (Р = Д) Фиксируется системой максимальное количество вещества и энергии Цикл обмена веществом и потоки распределения энергии замедляются Одум (1975) Отношение валовой продукции к чистой убывает Линейные пастбищные цепи заменяются сетевыми, детритными

Планетарная роль биогеоценотического покрова
Prezentatsia_BGTs_2_kurs_Ekologi.ppt