ДНК окончательный.ppt
- Количество слайдов: 49

 Краткая история открытия двойной спирали ДНК впервые выделил из гноя в 1869 г. Иоганн Фридрих Мишер. 1889 Альтманом выделена чистая ДНК. На рубеже 40 -х годов Дж. Бидл (род. в 1903 г. ) и Э. Тейтум (1909— 1975) заложили основы биохимической генетики. Они показали, что мутации у хлебной плесени Neurospora crassa блокируют различные этапы клеточного метаболизма и высказали предположение о том, что гены контролируют биосинтез ферментов. Иоганн Фридрих Мишер
Краткая история открытия двойной спирали ДНК впервые выделил из гноя в 1869 г. Иоганн Фридрих Мишер. 1889 Альтманом выделена чистая ДНК. На рубеже 40 -х годов Дж. Бидл (род. в 1903 г. ) и Э. Тейтум (1909— 1975) заложили основы биохимической генетики. Они показали, что мутации у хлебной плесени Neurospora crassa блокируют различные этапы клеточного метаболизма и высказали предположение о том, что гены контролируют биосинтез ферментов. Иоганн Фридрих Мишер
 О. Эвери К. Мак-Леод М. Мак-Карти В 1944 г. американцы О. Эвери, К. Мак-Леод и М. Мак-Карти доказали генетическую роль нуклеиновых кислот в экспериментах по трансформации признаков у микроорганизмов — пневмококков. Они идентифицировали природу трансформирующего агента как молекулы ДНК. Это открытие символизировало возникновение нового этапа в генетике — рождение молекулярной генетики, которая легла в основу целого ряда революционизирующих открытий в биологии XX века.
О. Эвери К. Мак-Леод М. Мак-Карти В 1944 г. американцы О. Эвери, К. Мак-Леод и М. Мак-Карти доказали генетическую роль нуклеиновых кислот в экспериментах по трансформации признаков у микроорганизмов — пневмококков. Они идентифицировали природу трансформирующего агента как молекулы ДНК. Это открытие символизировало возникновение нового этапа в генетике — рождение молекулярной генетики, которая легла в основу целого ряда революционизирующих открытий в биологии XX века.
 Эксперимент Эвери, Мак. Леода и Мк. Карти
Эксперимент Эвери, Мак. Леода и Мк. Карти
 «До Уотсон-Криковская» история В начале 50 -х годовx XX века профессор биохимии Колумбийского университета Эрвин Чаргафф определил состав ДНК с гораздо большей точностью по сравнению с предыдущими исследователями. Он обнаружил, что содержание 4 типов азотистых оснований не соответствует соотношению 1: 1: 1: 1, как предполагали ранее. Количество аденина всегда равно количеству тимина, а содержание гуанина - содержанию цитозина. Эта закономерность соотношения А -Т и Г – Ц получила название правило Чаргаффа Эрвин Чаргафф На основе правил Чаргаффа Уотсон и Крик постулировали «комплементарность» взаимодействия азотистых оснований в составе молекулы ДНК.
«До Уотсон-Криковская» история В начале 50 -х годовx XX века профессор биохимии Колумбийского университета Эрвин Чаргафф определил состав ДНК с гораздо большей точностью по сравнению с предыдущими исследователями. Он обнаружил, что содержание 4 типов азотистых оснований не соответствует соотношению 1: 1: 1: 1, как предполагали ранее. Количество аденина всегда равно количеству тимина, а содержание гуанина - содержанию цитозина. Эта закономерность соотношения А -Т и Г – Ц получила название правило Чаргаффа Эрвин Чаргафф На основе правил Чаргаффа Уотсон и Крик постулировали «комплементарность» взаимодействия азотистых оснований в составе молекулы ДНК.
 «До Уотсон-Криковская» история В 1950 году английский физик М. Уилкинс получил рентгенограмму ДНК. Рентгенограммы, полученные на высокоочищенной ДНК, позволили Розалинд Франклин увидеть четкий крестообразный рисунок – опознавательный знак двойной спирали. Розалинд Франклин умерла в 1958 г.
«До Уотсон-Криковская» история В 1950 году английский физик М. Уилкинс получил рентгенограмму ДНК. Рентгенограммы, полученные на высокоочищенной ДНК, позволили Розалинд Франклин увидеть четкий крестообразный рисунок – опознавательный знак двойной спирали. Розалинд Франклин умерла в 1958 г.
 Приоритет в расшифровке структуры молекулы ДНК принадлежит американскому вирусологу Дж. Уотсону (род. в 1928 г. ) и английскому физику Ф. Крику (род. в 1916 г. ), опубликовавшим в 1953 г. структурную модель этого полимера.
Приоритет в расшифровке структуры молекулы ДНК принадлежит американскому вирусологу Дж. Уотсону (род. в 1928 г. ) и английскому физику Ф. Крику (род. в 1916 г. ), опубликовавшим в 1953 г. структурную модель этого полимера.
 1962 год – Нобелевская премия
1962 год – Нобелевская премия
 Компоненты нуклеиновых кислот При полном гидролизе нуклеиновых кислот образуются: 1. азотистые основания (пурины и пиримидины); 2. остатки сахаров (рибозы – от РНК; дезоксирибозы – от ДНК); 3. фосфорная кислота. При неполном гидролизе НК образуются: 1. нуклеотиды; 2. нуклеозиды.
Компоненты нуклеиновых кислот При полном гидролизе нуклеиновых кислот образуются: 1. азотистые основания (пурины и пиримидины); 2. остатки сахаров (рибозы – от РНК; дезоксирибозы – от ДНК); 3. фосфорная кислота. При неполном гидролизе НК образуются: 1. нуклеотиды; 2. нуклеозиды.
 Мононуклеотиды Фосфат Пуриновые или пиримидиновые азотистые основания Пентоза
Мононуклеотиды Фосфат Пуриновые или пиримидиновые азотистые основания Пентоза
 Азотистые основания Пиримидин Пурин
Азотистые основания Пиримидин Пурин
 Пурины Аденин Гуанин
Пурины Аденин Гуанин
 Пиримидины Цитозин Тимин Урацил
Пиримидины Цитозин Тимин Урацил
 β-D-рибоза
β-D-рибоза
 Структура азотистых оснований и нуклеотидов
Структура азотистых оснований и нуклеотидов
 фосфорной кислоты.") Фосфатные остатки Нуклеотиды – это нуклеозиды соединенные сахаро-фосфатной связью с остатком (остатками) фосфорной кислоты.
Фосфатные остатки Нуклеотиды – это нуклеозиды соединенные сахаро-фосфатной связью с остатком (остатками) фосфорной кислоты.
 Рибонуклеотиды АМФ ГМФ УМФ СМФ
Рибонуклеотиды АМФ ГМФ УМФ СМФ
 Дезоксирибонуклеотиды АМФ ГМФ ТМФ СМФ
Дезоксирибонуклеотиды АМФ ГМФ ТМФ СМФ
. • Сигнальная (вторичные мессенджеры: ГТФ, ц. ГМФ, ЦАМФ). •") Функции нуклеотидов • Энергетическая (АТФ). • Сигнальная (вторичные мессенджеры: ГТФ, ц. ГМФ, ЦАМФ). • Мономеры коферментов (НАД, НАДФ, ФАД, ко-энзим А, метионин-аденозин). • Мономеры нуклеиновых кислот (РНК, ДНК). • Активация при метаболизме липидов и моносахаридов (УТФ, СТФ). • Участие в дезактивации различных спиртов и фенолов (УДФ-глюкуроновая кислота).
Функции нуклеотидов • Энергетическая (АТФ). • Сигнальная (вторичные мессенджеры: ГТФ, ц. ГМФ, ЦАМФ). • Мономеры коферментов (НАД, НАДФ, ФАД, ко-энзим А, метионин-аденозин). • Мономеры нуклеиновых кислот (РНК, ДНК). • Активация при метаболизме липидов и моносахаридов (УТФ, СТФ). • Участие в дезактивации различных спиртов и фенолов (УДФ-глюкуроновая кислота).
 Первичная структура нуклеиновых кислот • Азотистые основания прикреплены к сахарофосфатному остову. • В сахаро-фосфатном остове чередуются остатки мосахарида (пентозы) и фосфорной кислоты. • Остатки пентозы и сахаро-фосфатного остова соединяются фосфо-эфирной связью. • Молекула ассиметрична, есть 5’ и 3’ концы.
Первичная структура нуклеиновых кислот • Азотистые основания прикреплены к сахарофосфатному остову. • В сахаро-фосфатном остове чередуются остатки мосахарида (пентозы) и фосфорной кислоты. • Остатки пентозы и сахаро-фосфатного остова соединяются фосфо-эфирной связью. • Молекула ассиметрична, есть 5’ и 3’ концы.
 Различия РНК и ДНК РНК ДНК рибоза 2’-дезоксирибоза А, Г, У, Ц А, Г, Т, Ц Вторичная структура одноцепочечная молекула сложной вторичной структурой двухцепочечная спираль Функция транспортная каталитическая матрица для синтеза белка сохранение и передача генетической информации Пентоза Азотистые основания
Различия РНК и ДНК РНК ДНК рибоза 2’-дезоксирибоза А, Г, У, Ц А, Г, Т, Ц Вторичная структура одноцепочечная молекула сложной вторичной структурой двухцепочечная спираль Функция транспортная каталитическая матрица для синтеза белка сохранение и передача генетической информации Пентоза Азотистые основания
 Вторичная структура ДНК Две спиральные полинуклеотидные цепи закручены вокруг общей оси. Цепи направлены в противоположные стороны, то есть антипараллельны. Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы – снаружи. Плоскости оснований перпендикулярны оси спирали. Плоскости остатков сахара расположены почти под прямым углом к основаниям. Диаметр спирали 20 А. Расстояние между соседними основаниями вдоль оси спирали 0, 34 нм, они повернуты относительно друга на 36°. Таким образом, на один виток спирали каждой из цепей приходится 10 нуклеотидов, что соответствует 3, 4 нм. Две цепи удерживаются вместе водородными связями между парами оснований. Аденин всегда спаривается с тимином, гуанин - с цитозином. На последовательность оснований в полинуклеотидной цепи не накладывается никаких ограничений. Определенная последовательность оснований несет конкретную генетическую информацию.
Вторичная структура ДНК Две спиральные полинуклеотидные цепи закручены вокруг общей оси. Цепи направлены в противоположные стороны, то есть антипараллельны. Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы – снаружи. Плоскости оснований перпендикулярны оси спирали. Плоскости остатков сахара расположены почти под прямым углом к основаниям. Диаметр спирали 20 А. Расстояние между соседними основаниями вдоль оси спирали 0, 34 нм, они повернуты относительно друга на 36°. Таким образом, на один виток спирали каждой из цепей приходится 10 нуклеотидов, что соответствует 3, 4 нм. Две цепи удерживаются вместе водородными связями между парами оснований. Аденин всегда спаривается с тимином, гуанин - с цитозином. На последовательность оснований в полинуклеотидной цепи не накладывается никаких ограничений. Определенная последовательность оснований несет конкретную генетическую информацию.
 Полинуклеотиды Одно из упрощенных изображений структуры полинуклеотидной цепи:
Полинуклеотиды Одно из упрощенных изображений структуры полинуклеотидной цепи:
 ВАЖНО!!! конец
ВАЖНО!!! конец
 Нуклеиновые кислоты ? ?
Нуклеиновые кислоты ? ?
 Уотсон-Криковский тип спаривания
Уотсон-Криковский тип спаривания
 Модель В-формы ДНК
Модель В-формы ДНК
 Структура различных форм ДНК
Структура различных форм ДНК
 1. Две спиральные полинуклеотидные цепи закручены вокруг общей оси. Цепи направлены в противоположные стороны. 2. Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы – снаружи. 3. Диаметр спирали 20 А. Расстояние между соседними основаниями вдоль оси спирали 3, 4 А, они повернуты относительно друга на 36°. Таким образом, на один виток спирали каждой из цепей приходится 10 нуклеотидов, что соответствует 34 А. 4. Две цепи удерживаются вместе водородными связями между парами оснований: А - Т, Г-Ц. 5. На последовательность оснований в полинуклеотидной цепи не накладывается никаких ограничений. Определенная последовательность оснований несет конкретную генетическую информацию.
1. Две спиральные полинуклеотидные цепи закручены вокруг общей оси. Цепи направлены в противоположные стороны. 2. Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы – снаружи. 3. Диаметр спирали 20 А. Расстояние между соседними основаниями вдоль оси спирали 3, 4 А, они повернуты относительно друга на 36°. Таким образом, на один виток спирали каждой из цепей приходится 10 нуклеотидов, что соответствует 34 А. 4. Две цепи удерживаются вместе водородными связями между парами оснований: А - Т, Г-Ц. 5. На последовательность оснований в полинуклеотидной цепи не накладывается никаких ограничений. Определенная последовательность оснований несет конкретную генетическую информацию.
 Репликация ДНК
Репликация ДНК
 • Репликация - это основной генетический процесс, в ходе которого родительские молекулы ДНК удваиваются и затем распределяются между потомками.
• Репликация - это основной генетический процесс, в ходе которого родительские молекулы ДНК удваиваются и затем распределяются между потомками.
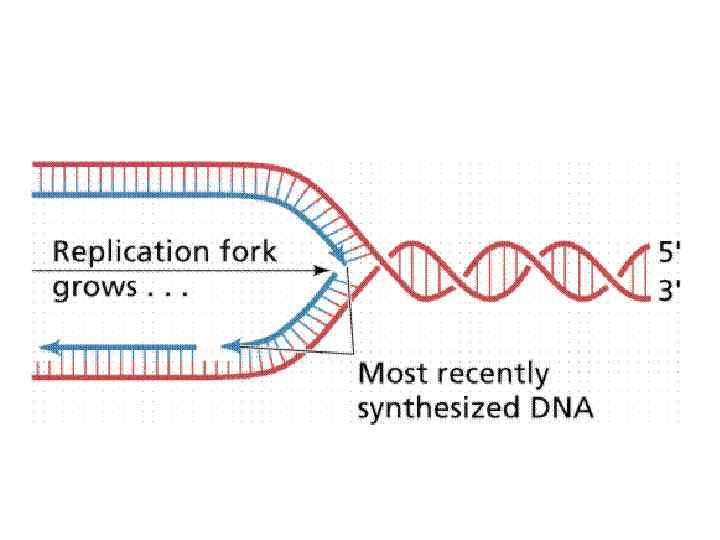
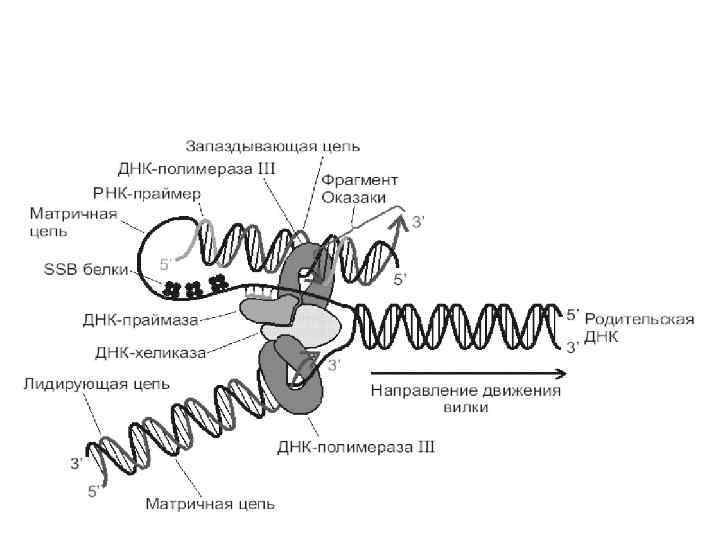
 Основной принцип репликации: • Удвоение ДНК происходит вследствие того, что цепи расходятся, а потом каждая цепь служит матрицей, на которой собирается комплементарная ей новая цепь ДНК. • В результате образуются две дочерние, двуспиральные, неотличимые по строению от родительской ДНК молекулы.
Основной принцип репликации: • Удвоение ДНК происходит вследствие того, что цепи расходятся, а потом каждая цепь служит матрицей, на которой собирается комплементарная ей новая цепь ДНК. • В результате образуются две дочерние, двуспиральные, неотличимые по строению от родительской ДНК молекулы.
. ORI;") Правила репликации: 1. Репликация всегда осуществляется с исключение – искусственные системы (ПЦР). ORI;
Правила репликации: 1. Репликация всегда осуществляется с исключение – искусственные системы (ПЦР). ORI;
 Правила репликации: 2. Репликация полуконсервативна.
Правила репликации: 2. Репликация полуконсервативна.
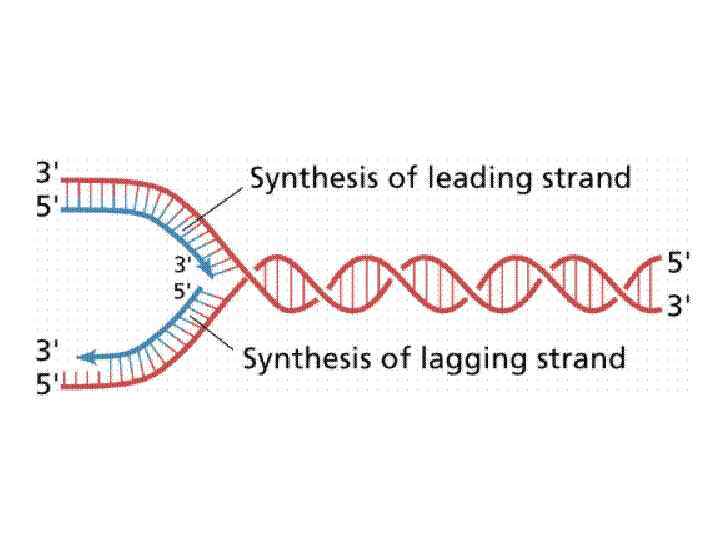
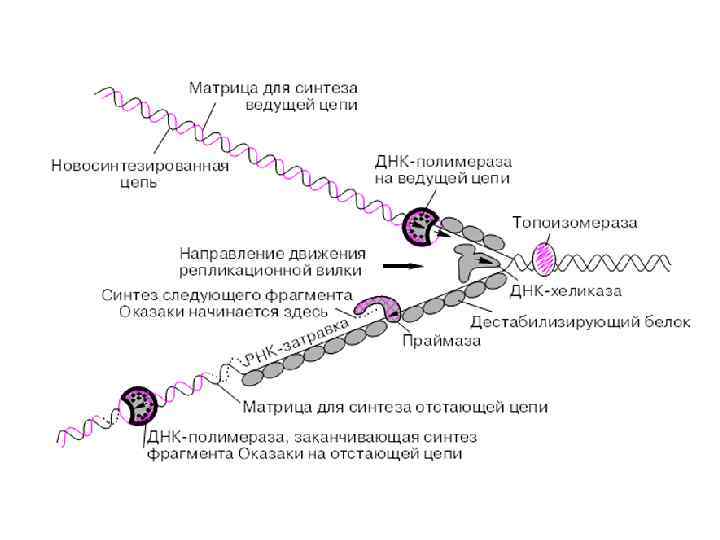
 Правила репликации: Синтез ДНК всегда осуществляется в направлении 5’ – 3’. Это связано с тем фактом, что присоединение к 3’ – ОН энергетически более выгодно, а также с активностью коррекции присоединения нуклеотидов.
Правила репликации: Синтез ДНК всегда осуществляется в направлении 5’ – 3’. Это связано с тем фактом, что присоединение к 3’ – ОН энергетически более выгодно, а также с активностью коррекции присоединения нуклеотидов.
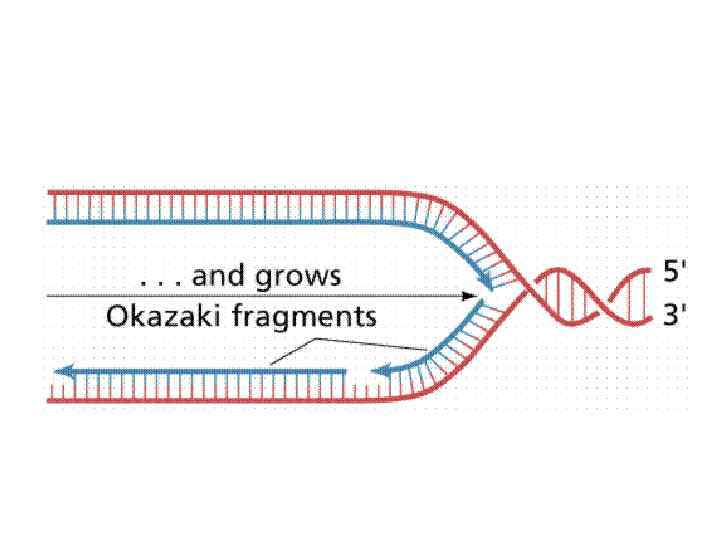
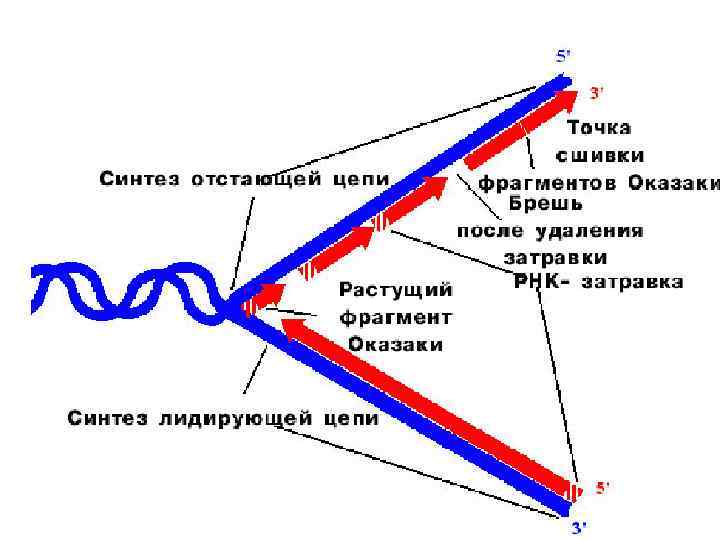
 Правила репликации: Синтез ДНК у всех организмов и в искусственных системах требует затравки в отличие от синтеза РНК. Затравка может быть белковая (вирусы), РНК и ДНК.
Правила репликации: Синтез ДНК у всех организмов и в искусственных системах требует затравки в отличие от синтеза РНК. Затравка может быть белковая (вирусы), РНК и ДНК.
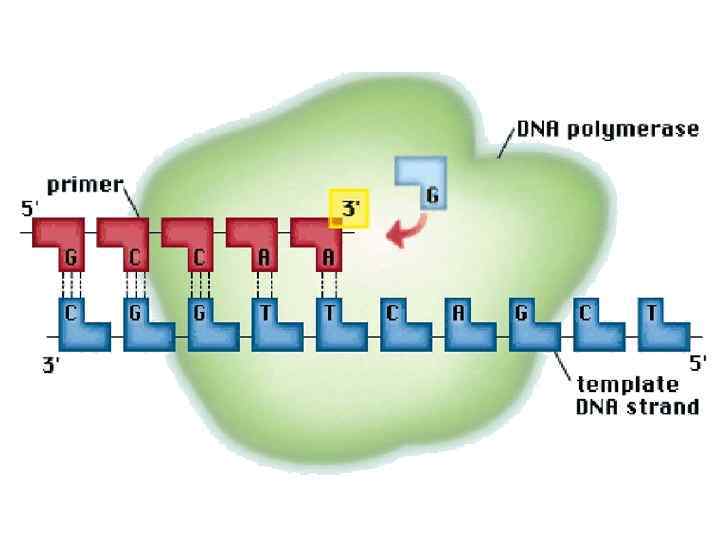
 Ферменты репликации Основным ферментом репликации является ДНК-полимераза, этот ДНК-полимераза фермент впервые был выделен в 1956 г. из Esherichia coli. ДНК-полимераза осуществляет полимеризацию нуклеотидов в цепь, комплементарную матричной. Субстратом являются тринуклеотиды. Коферментом двухвалентные ионы Zn, обеспечивающие правильную ориентацию 3’гидроксила альфа-фосфата и нуклеотида матрицы Для полимеризации необходимы ионы Mg, которые образуют комплекс с нуклеотид-трифосфатами и уже этот комплекс поступает в активный центр фермента. Энергия необходимая для катализа выделяется при расщеплении макроэргической связи между альфа и бета фосфатными группами.
Ферменты репликации Основным ферментом репликации является ДНК-полимераза, этот ДНК-полимераза фермент впервые был выделен в 1956 г. из Esherichia coli. ДНК-полимераза осуществляет полимеризацию нуклеотидов в цепь, комплементарную матричной. Субстратом являются тринуклеотиды. Коферментом двухвалентные ионы Zn, обеспечивающие правильную ориентацию 3’гидроксила альфа-фосфата и нуклеотида матрицы Для полимеризации необходимы ионы Mg, которые образуют комплекс с нуклеотид-трифосфатами и уже этот комплекс поступает в активный центр фермента. Энергия необходимая для катализа выделяется при расщеплении макроэргической связи между альфа и бета фосфатными группами.
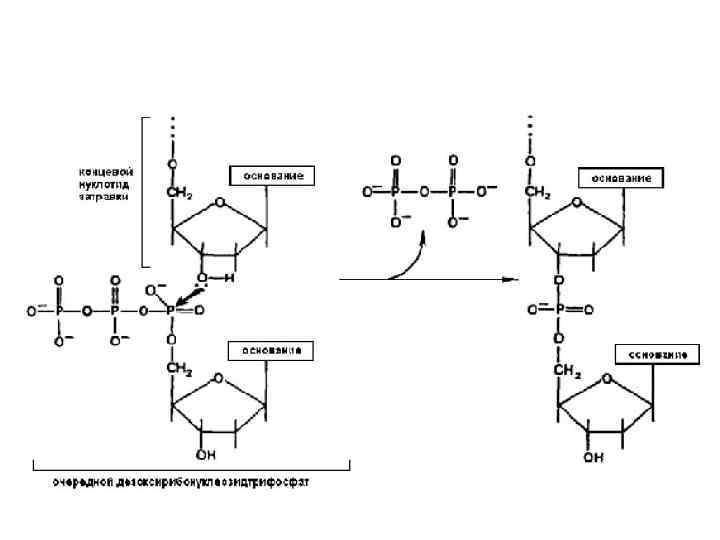
 поступает в активный центр фермента и образует водородные связи с") Этапы полимеризации: Комплекс (Mg/дезоксирибонуклеотидтрифосфат) поступает в активный центр фермента и образует водородные связи с нуклеотидом матрицы. Если нуклеотиды комплементарны, то образуется комплекс, в котором нуклеотид матрицы, 3’ концевой нуклеотид нарастающей цепи и новый нуклеотидтрифосфат образуют комплекс с правильной ориентацией всех молекул. После образования комплекса происходит нуклеофильная атака кислородом 3’ концевого гидроксила, 5’-концевого альфа-фосфата, следующего нуклеотида; в результате образуется фосфоэфирная связь и выщепляется пирофосфат. После присоединения нового нуклеотида происходит второй этап коррекции полимеризации; если новоприсоединенный нуклеотид не комплементарен, то следующий не поступает в активный центр фермента и ДНК полимераза своей 3’-5’ нуклеазной активностью отщепляет неправильный нуклеотид. Далее цикл повторяется.
Этапы полимеризации: Комплекс (Mg/дезоксирибонуклеотидтрифосфат) поступает в активный центр фермента и образует водородные связи с нуклеотидом матрицы. Если нуклеотиды комплементарны, то образуется комплекс, в котором нуклеотид матрицы, 3’ концевой нуклеотид нарастающей цепи и новый нуклеотидтрифосфат образуют комплекс с правильной ориентацией всех молекул. После образования комплекса происходит нуклеофильная атака кислородом 3’ концевого гидроксила, 5’-концевого альфа-фосфата, следующего нуклеотида; в результате образуется фосфоэфирная связь и выщепляется пирофосфат. После присоединения нового нуклеотида происходит второй этап коррекции полимеризации; если новоприсоединенный нуклеотид не комплементарен, то следующий не поступает в активный центр фермента и ДНК полимераза своей 3’-5’ нуклеазной активностью отщепляет неправильный нуклеотид. Далее цикл повторяется.
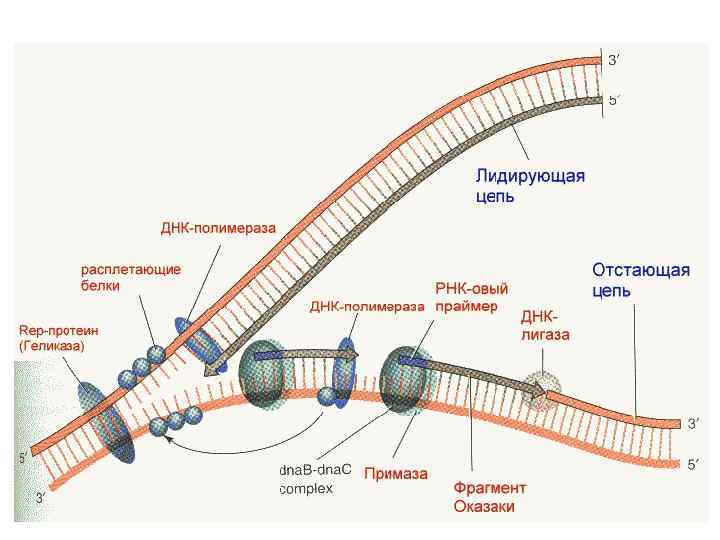
 Начало репликации Геликаза Репликационная вилка
Начало репликации Геликаза Репликационная вилка