

Лекция 9 биологи.pptx
- Количество слайдов: 108
 Конкуренция И СОСУЩЕСТВОВАНИЕ Competition and coexistence
Конкуренция И СОСУЩЕСТВОВАНИЕ Competition and coexistence
 Два способа достичь сосуществования: 1. Виды расходятся по разным экологическим нишам: соответственно, конкуренция между ними отсутствует или же резко ослаблена.
Два способа достичь сосуществования: 1. Виды расходятся по разным экологическим нишам: соответственно, конкуренция между ними отсутствует или же резко ослаблена.
 Механизм победы второго вида совершенно аналогичен механизму победы первого вида Побеждает вид 2 (независимо от начальных условий) 2 1 Как объяснить сильное воздействие второго вида на первый и слабое воздействие первого вида на второй в терминах ниши? Ниша вида 2 включает нишу вида 1 (т. е. вид 2 более эврибионтен, например, имеет более разнообразные места для гнездования или для нерестилищ) – поэтому вид 2 побеждает. Эти неравенства выполняются, когда коэффициент воздействия второго вида на первый, α 12, большой, а коэффициент воздействия первого вида на второй, α 21, - маленький. (Почему? ) Неудивительно, что в результате сильного воздействия второго вида на первый побеждает второй вид! F 2 Вид 1 F 1
Механизм победы второго вида совершенно аналогичен механизму победы первого вида Побеждает вид 2 (независимо от начальных условий) 2 1 Как объяснить сильное воздействие второго вида на первый и слабое воздействие первого вида на второй в терминах ниши? Ниша вида 2 включает нишу вида 1 (т. е. вид 2 более эврибионтен, например, имеет более разнообразные места для гнездования или для нерестилищ) – поэтому вид 2 побеждает. Эти неравенства выполняются, когда коэффициент воздействия второго вида на первый, α 12, большой, а коэффициент воздействия первого вида на второй, α 21, - маленький. (Почему? ) Неудивительно, что в результате сильного воздействия второго вида на первый побеждает второй вид! F 2 Вид 1 F 1
 Классическая модель конкуренции Побеждает вид 1 Побеждает вид 2 2 1 Анализ УСТОЙЧИВОГО равновесия 1 Равновесие устойчивое 2 2 Равновесие неустойчивое 1 В этом случае виды подавляют друга слабо, так что возможно сосуществование
Классическая модель конкуренции Побеждает вид 1 Побеждает вид 2 2 1 Анализ УСТОЙЧИВОГО равновесия 1 Равновесие устойчивое 2 2 Равновесие неустойчивое 1 В этом случае виды подавляют друга слабо, так что возможно сосуществование
 Равновесие устойчивое 1 F 2 Вид 1 Вид 2 2 F 1
Равновесие устойчивое 1 F 2 Вид 1 Вид 2 2 F 1
 Два способа достичь сосуществования: 2. Виды занимают одну экологическую нишу, конкурируют за общие ресурсы, но вытеснение одного вида другим откладывается на долгое время, прерывается извне или же не допускается путем интерференции (ингибирования одних видов другими).
Два способа достичь сосуществования: 2. Виды занимают одну экологическую нишу, конкурируют за общие ресурсы, но вытеснение одного вида другим откладывается на долгое время, прерывается извне или же не допускается путем интерференции (ингибирования одних видов другими).
 Способы сосуществования видов, занимающих одну и ту же экологическую нишу: 1. Ограничение разными ресурсами. 2. Смена конкурентного преимущества по мере расходования лимитирующего ресурса или же, наоборот, дополнительного его поступления в среду. 3. Прерывание процесса вытеснения нарушениями извне за счет абиотических (пожары, осыпи и т. п. ) или биотических факторов (выедание, вытаптывание). 4. Интерференция – ингибирование одних видов другими за счет специально выделяемых веществ. 5. Конвергенция разных видов, направленная на возрастание экологического сходства (сосуществование за счет «отложенного вытесенения» ). 6. Мутуалистические взаимоотношения между видами, иногда опосредованные (например, через микоризу). 7. Гетерогенная среда (эфемерный биотоп)
Способы сосуществования видов, занимающих одну и ту же экологическую нишу: 1. Ограничение разными ресурсами. 2. Смена конкурентного преимущества по мере расходования лимитирующего ресурса или же, наоборот, дополнительного его поступления в среду. 3. Прерывание процесса вытеснения нарушениями извне за счет абиотических (пожары, осыпи и т. п. ) или биотических факторов (выедание, вытаптывание). 4. Интерференция – ингибирование одних видов другими за счет специально выделяемых веществ. 5. Конвергенция разных видов, направленная на возрастание экологического сходства (сосуществование за счет «отложенного вытесенения» ). 6. Мутуалистические взаимоотношения между видами, иногда опосредованные (например, через микоризу). 7. Гетерогенная среда (эфемерный биотоп)
 *что можно рассматривать и как разные ниши") 1. Ограничение разными ресурсами* (или разными хищниками) *что можно рассматривать и как разные ниши
1. Ограничение разными ресурсами* (или разными хищниками) *что можно рассматривать и как разные ниши
 (resource-based competition theory) (1) В теории в явном") Ресурсная теория конкуренции Тильмана (David Tilman) (resource-based competition theory) (1) В теории в явном виде присутствуют ресурсы (или ресурс, если конкуренция за один ресурс) – в классической теории конкуренции Вольтерры-Лотки ресурсов нет! (2) Успех вида в конкуренции определяется пороговой концентрацией ресурса для этого вида, а возможность сосуществования с др. видами – скоростью потребления лимитирующего ресурса (наклоном вектора потребления в точке равновесия) (3) Это – графическая теория. Графическая теория не заменяет строгой математической теории. Однако теория, изложенная на графическом языке, часто (хотя не всегда!) обладает большей общностью и всегда обладает большей наглядностью, что облегчает ее восприятие и открывает возможность для экспериментальной проверки
Ресурсная теория конкуренции Тильмана (David Tilman) (resource-based competition theory) (1) В теории в явном виде присутствуют ресурсы (или ресурс, если конкуренция за один ресурс) – в классической теории конкуренции Вольтерры-Лотки ресурсов нет! (2) Успех вида в конкуренции определяется пороговой концентрацией ресурса для этого вида, а возможность сосуществования с др. видами – скоростью потребления лимитирующего ресурса (наклоном вектора потребления в точке равновесия) (3) Это – графическая теория. Графическая теория не заменяет строгой математической теории. Однако теория, изложенная на графическом языке, часто (хотя не всегда!) обладает большей общностью и всегда обладает большей наглядностью, что облегчает ее восприятие и открывает возможность для экспериментальной проверки
 Пороговая концентрация ресурса – концентрация, при которой рождаемость потребителя равна его смертности, так что скорость роста численности равна нулю и численность постоянна Удельная скорость роста r Удельная смертность m Удельная рождаемость b r Пороговая концентрация ресурса R* Временной ход обилия потребителя и ресурса
Пороговая концентрация ресурса – концентрация, при которой рождаемость потребителя равна его смертности, так что скорость роста численности равна нулю и численность постоянна Удельная скорость роста r Удельная смертность m Удельная рождаемость b r Пороговая концентрация ресурса R* Временной ход обилия потребителя и ресурса
 Два ресурса: Изоклины нулевого роста численности r = 0 в пространстве двух ресурсов R 2 Полностью взаимозаменяемые ресурсы R 2 R*2 r > 0 r = 0 0 Концентрация 2 -го ресурса Незаменимые ресурсы r = 0 R*2 r < 0 R*1 R 1 Концентрация 1 -го ресурса Замечание. Эти рисунки представляют иллюстрацию того, что R* - это нижняя граница зоны толерантности (в терминах концепции ниши) R*1 R 1 Концентрация 1 -го ресурса при (Потребление ресурса 2) = (Потребление ресурса 1)
Два ресурса: Изоклины нулевого роста численности r = 0 в пространстве двух ресурсов R 2 Полностью взаимозаменяемые ресурсы R 2 R*2 r > 0 r = 0 0 Концентрация 2 -го ресурса Незаменимые ресурсы r = 0 R*2 r < 0 R*1 R 1 Концентрация 1 -го ресурса Замечание. Эти рисунки представляют иллюстрацию того, что R* - это нижняя граница зоны толерантности (в терминах концепции ниши) R*1 R 1 Концентрация 1 -го ресурса при (Потребление ресурса 2) = (Потребление ресурса 1)
 Два ресурса: Вектора C и U Вектор потребления C, «вектор снабжения» U и ресурсная точка (S 1, S 2) Вектор потребления C – это вектор, проекции которого на оси ресурсов представляют собой скорости потребления соответствующих ресурсов при данной концентрации ресурсов (т. е. в данной точке пространства (R 1, R 2)). Таким образом, наклон вектора потребления (точнее, тангенс угла наклона) – это отношение скорости потребления ресурса 2 к скорости потребления ресурса 1 Вектор поступления ресурсов U и ресурсная точка (S 1, S 2) Вектор потребления C и вектор удельного R 2 потребления Вектор U всегда направлен в точку (S 1, S 2) R 2 или S 2 (S 1, S 2) α∙(S 2 - R 2’) N*c 1 C R 2 ’ U α∙(S 1 - R 1’) N*c 2 R 1 ’ R 1 или S 1
Два ресурса: Вектора C и U Вектор потребления C, «вектор снабжения» U и ресурсная точка (S 1, S 2) Вектор потребления C – это вектор, проекции которого на оси ресурсов представляют собой скорости потребления соответствующих ресурсов при данной концентрации ресурсов (т. е. в данной точке пространства (R 1, R 2)). Таким образом, наклон вектора потребления (точнее, тангенс угла наклона) – это отношение скорости потребления ресурса 2 к скорости потребления ресурса 1 Вектор поступления ресурсов U и ресурсная точка (S 1, S 2) Вектор потребления C и вектор удельного R 2 потребления Вектор U всегда направлен в точку (S 1, S 2) R 2 или S 2 (S 1, S 2) α∙(S 2 - R 2’) N*c 1 C R 2 ’ U α∙(S 1 - R 1’) N*c 2 R 1 ’ R 1 или S 1
") Один потребитель и два ресурса Когда система находится в равновесии? В случае, если: (1) Потребитель находится на нулевой изоклине, то есть в положении равновесия (r = 0) (2) Вектор потребления равен вектору снабжения, то есть ресурс также находится в положении равновесия (d. R/dt = 0) В точке E равновесие Точка E – положение равновесия локально устойчивое R 2 или S 2 (S 1, S 2) U 1 (S 1, S 2) C 1 E UE E CE CE UE U 2 C 2 R 1 или S 1
Один потребитель и два ресурса Когда система находится в равновесии? В случае, если: (1) Потребитель находится на нулевой изоклине, то есть в положении равновесия (r = 0) (2) Вектор потребления равен вектору снабжения, то есть ресурс также находится в положении равновесия (d. R/dt = 0) В точке E равновесие Точка E – положение равновесия локально устойчивое R 2 или S 2 (S 1, S 2) U 1 (S 1, S 2) C 1 E UE E CE CE UE U 2 C 2 R 1 или S 1
 Изоклины пересекаются: Объяснение исхода в области I, II и IV R 2 или S 2 I A II B III VI SI SII V CB IV CA SIV I SI R 1 или S 1 SI – при такой максимальной концентрации ресурсов не может существовать ни один из 2 -х видов SII или SIV – при такой максимальной концентрации ресурсов может существовать только вид A – для случая SII или вид B – для случая SIV
Изоклины пересекаются: Объяснение исхода в области I, II и IV R 2 или S 2 I A II B III VI SI SII V CB IV CA SIV I SI R 1 или S 1 SI – при такой максимальной концентрации ресурсов не может существовать ни один из 2 -х видов SII или SIV – при такой максимальной концентрации ресурсов может существовать только вид A – для случая SII или вид B – для случая SIV
 Изоклины пересекаются: Объяснение исхода в области III и V R 2 или S 2 A II B SIII I SIII или SV – при такой максимальной концентрации VI U SV V CB IV CA I ресурсов в среде могут существовать оба вида. Кто из них победит? Рассмотрим ситуацию в случае SIII. Поскольку система должна прийти в состояние равновесия, в принципе возможны три исхода: равновесная точка для вида B, если бы вида A не было (на изоклине вида В – обозначена розовым цветом); равновесная точка для вида А, если бы вида B не было (на изоклине вида А – обозначена коричневым цветом); равновесная точка по численностям обоих видов (на пересечении их изоклин – обозначена зеленым цветом), в которой, однако, виды не находятся в равновесии с ресурсами. R 1 или S 1 Как Вы думаете, какой исход будет реализован на самом деле (рассматриваем область III)? Ответ: победит вид А. Почему? Вид В не может удержаться в своей равновесной точке (потому что вид A продолжает расти). Виды A и В не могут удержаться в общей равновесной точке (ОРТ), потому что в этой точке в систему поступает много ресурса 2 и мало ресурса 1. (Почему? Ответ дан на рисунке в виде разложения вектора снабжения U на составляющие. ) Однако в ОРТ вид A лимитирован ресурсом 2, а вид B – ресурсом 1. Поэтому в этой точке вид A получает преимущество над видом B. В итоге вид А «сваливается» в свою равновесную точку. Эта точка является устойчивой для вида A, поскольку численность вида B в окрестности этой точки падает, и он не может понизить концентрацию ресурса ниже лимитирующей для вида A.
Изоклины пересекаются: Объяснение исхода в области III и V R 2 или S 2 A II B SIII I SIII или SV – при такой максимальной концентрации VI U SV V CB IV CA I ресурсов в среде могут существовать оба вида. Кто из них победит? Рассмотрим ситуацию в случае SIII. Поскольку система должна прийти в состояние равновесия, в принципе возможны три исхода: равновесная точка для вида B, если бы вида A не было (на изоклине вида В – обозначена розовым цветом); равновесная точка для вида А, если бы вида B не было (на изоклине вида А – обозначена коричневым цветом); равновесная точка по численностям обоих видов (на пересечении их изоклин – обозначена зеленым цветом), в которой, однако, виды не находятся в равновесии с ресурсами. R 1 или S 1 Как Вы думаете, какой исход будет реализован на самом деле (рассматриваем область III)? Ответ: победит вид А. Почему? Вид В не может удержаться в своей равновесной точке (потому что вид A продолжает расти). Виды A и В не могут удержаться в общей равновесной точке (ОРТ), потому что в этой точке в систему поступает много ресурса 2 и мало ресурса 1. (Почему? Ответ дан на рисунке в виде разложения вектора снабжения U на составляющие. ) Однако в ОРТ вид A лимитирован ресурсом 2, а вид B – ресурсом 1. Поэтому в этой точке вид A получает преимущество над видом B. В итоге вид А «сваливается» в свою равновесную точку. Эта точка является устойчивой для вида A, поскольку численность вида B в окрестности этой точки падает, и он не может понизить концентрацию ресурса ниже лимитирующей для вида A.
 Изоклины пересекаются: Объяснение исхода в области VI R 2 или S 2 Равновесие устойчивое B A VI III I II SVI U V CB IV CA I R 1 или S 1 SVI – ресурсная точка находится между лучами – продолжениями векторов потребления, и каждый вид преимущественно потребляет тот ресурс, который в точке пересечения изоклин является для этого вида лимитирующим. (Убедитесь в этом, глядя на рисунок!) Система имеет 3 потенциальных равновесных точки: равновесная точка для вида B, если бы вида A не было (на изоклине вида В – обозначена розовым цветом); равновесная точка для вида А, если бы вида B не было (на изоклине вида А – обозначена коричневым цветом); равновесная точка по численностям обоих видов (на пересечении их изоклин – обозначена зеленым цветом), в которой, однако, виды не находятся в равновесии с ресурсами. Как Вы думаете, какой исход будет реализован на самом деле? Ответ: Сосуществование видов (то есть система окажется в общей равновесной точке – ОРТ), причем это равновесие будет устойчивым. Почему? Вид A не может удержаться в своей равновесной точке (потому что вид B продолжа-ет при этом расти). Вид B не может удержаться в своей равновесной точке (потому что вид A продолжает при этом расти). Виды A и В могут удержаться в общей равновесной точке, пото-му что равнодействующая потребления и поступления ресурсов (зеленая стрелка на рисунке) ведет систему в область, где численность вида A падает, а численность вида B растет. Но вид A преимущественно потребляет ресурс 2, поэтому концентрация ресурса 2 будет расти. С другой стороны, вид B преимущественно потребляет ресурс 1, поэтому концентрация ресур-са 1 будет падать. В итоге система возвращается в положение равновесия – равновесие устойчивое!
Изоклины пересекаются: Объяснение исхода в области VI R 2 или S 2 Равновесие устойчивое B A VI III I II SVI U V CB IV CA I R 1 или S 1 SVI – ресурсная точка находится между лучами – продолжениями векторов потребления, и каждый вид преимущественно потребляет тот ресурс, который в точке пересечения изоклин является для этого вида лимитирующим. (Убедитесь в этом, глядя на рисунок!) Система имеет 3 потенциальных равновесных точки: равновесная точка для вида B, если бы вида A не было (на изоклине вида В – обозначена розовым цветом); равновесная точка для вида А, если бы вида B не было (на изоклине вида А – обозначена коричневым цветом); равновесная точка по численностям обоих видов (на пересечении их изоклин – обозначена зеленым цветом), в которой, однако, виды не находятся в равновесии с ресурсами. Как Вы думаете, какой исход будет реализован на самом деле? Ответ: Сосуществование видов (то есть система окажется в общей равновесной точке – ОРТ), причем это равновесие будет устойчивым. Почему? Вид A не может удержаться в своей равновесной точке (потому что вид B продолжа-ет при этом расти). Вид B не может удержаться в своей равновесной точке (потому что вид A продолжает при этом расти). Виды A и В могут удержаться в общей равновесной точке, пото-му что равнодействующая потребления и поступления ресурсов (зеленая стрелка на рисунке) ведет систему в область, где численность вида A падает, а численность вида B растет. Но вид A преимущественно потребляет ресурс 2, поэтому концентрация ресурса 2 будет расти. С другой стороны, вид B преимущественно потребляет ресурс 1, поэтому концентрация ресур-са 1 будет падать. В итоге система возвращается в положение равновесия – равновесие устойчивое!
 Конкуренция двух видов за два ресурса: Как определить, является ли равновесие устойчивым? При отклонении от положения равновесия надо задать следующие вопросы: Какой ресурс растет? Какой ресурс падает? Какой потребитель растет? Какой потребитель падает? Оказывается, что положение равновесия устойчиво, если при отклонении от положения равновесия (а) растет потребитель, который преимущественно потребляет тот ресурс, которого стало больше (б) падает потребитель, который преимущественно потребляет тот ресурс, которого стало меньше Если равновесие устойчивое, то при отклонении от него в рост должен пойти тот вид, который в большей мере потребляет тот ресурс, численность которого при отклонении от положения равновесия возросла, так что численность этого ресурса станет падать. Напротив, на спад должен пойти тот вид, который в большей мере потребляет тот ресурс, численность которого упала. Соответственно, потребление этого ресурса уменьшится, и его численность станет расти. В итоге система вернется в положение равновесия.
Конкуренция двух видов за два ресурса: Как определить, является ли равновесие устойчивым? При отклонении от положения равновесия надо задать следующие вопросы: Какой ресурс растет? Какой ресурс падает? Какой потребитель растет? Какой потребитель падает? Оказывается, что положение равновесия устойчиво, если при отклонении от положения равновесия (а) растет потребитель, который преимущественно потребляет тот ресурс, которого стало больше (б) падает потребитель, который преимущественно потребляет тот ресурс, которого стало меньше Если равновесие устойчивое, то при отклонении от него в рост должен пойти тот вид, который в большей мере потребляет тот ресурс, численность которого при отклонении от положения равновесия возросла, так что численность этого ресурса станет падать. Напротив, на спад должен пойти тот вид, который в большей мере потребляет тот ресурс, численность которого упала. Соответственно, потребление этого ресурса уменьшится, и его численность станет расти. В итоге система вернется в положение равновесия.
 Примеры сосуществования многих видов на ограниченном числе ресурсов Планктонный парадокс Хатчинсона В верхнем освещенном и перемешиваемом слое водной толщи обитает множество организмов фитопланктона, ограниченных в своем развитии одними и теми же немногими ресурсами. Данная ситуация противоречит принципу конкурентного исключения Гаузе О планктонном парадоксе см. А. М. Гиляров. Популяционная экология. 1990, с. 163, 170 Из лекций А. М. Гилярова
Примеры сосуществования многих видов на ограниченном числе ресурсов Планктонный парадокс Хатчинсона В верхнем освещенном и перемешиваемом слое водной толщи обитает множество организмов фитопланктона, ограниченных в своем развитии одними и теми же немногими ресурсами. Данная ситуация противоречит принципу конкурентного исключения Гаузе О планктонном парадоксе см. А. М. Гиляров. Популяционная экология. 1990, с. 163, 170 Из лекций А. М. Гилярова

 Видовое богатство растений 40 по Tilman, 1982 20 3 1 2 4 Богатство почвы
Видовое богатство растений 40 по Tilman, 1982 20 3 1 2 4 Богатство почвы
 Сосуществование многих видов и парадокс обогащения: Объяснение, предложенное Тильманом Основано на ресурсной теории конкуренции и пространственной неоднородности ресурсов (S 1, S 2) – «ресурсная точка» данного местообитания или микроместообитания Круги – контуры местообитаний с данной пространственной неоднородностью ресурсов Концентрация ресурса 2, R 2 S – максимальная концентрация ресурса в данном местообитании, «емкость среды» Следствия: 1) В бедной среде всего на двух ресурсах может сосуществовать много видов 2) С обогащением среды число видов уменьшается Концентрация ресурса 1, R 1 3) Видовой состав зависит от того, в каком направлении идет обогащение среды Семь видов (a, b, c, d, e, f, g) конкурируют за два ресурса (1 и 2). Линии, образующие прямой угол, представляют нулевые изоклины для каждого из семи видов в пространстве ресурсов. Наклонные линии представляют продолжения векторов потребления в точке равновесия пары видов. Сектора (a+b и т. д. ) есть множество точек (совокупность местообитаний), в которых «емкость среды» такова, что в них возможно сосуществование соответствующей пары видов (таких секторов шесть). Из Tilman 1982.
Сосуществование многих видов и парадокс обогащения: Объяснение, предложенное Тильманом Основано на ресурсной теории конкуренции и пространственной неоднородности ресурсов (S 1, S 2) – «ресурсная точка» данного местообитания или микроместообитания Круги – контуры местообитаний с данной пространственной неоднородностью ресурсов Концентрация ресурса 2, R 2 S – максимальная концентрация ресурса в данном местообитании, «емкость среды» Следствия: 1) В бедной среде всего на двух ресурсах может сосуществовать много видов 2) С обогащением среды число видов уменьшается Концентрация ресурса 1, R 1 3) Видовой состав зависит от того, в каком направлении идет обогащение среды Семь видов (a, b, c, d, e, f, g) конкурируют за два ресурса (1 и 2). Линии, образующие прямой угол, представляют нулевые изоклины для каждого из семи видов в пространстве ресурсов. Наклонные линии представляют продолжения векторов потребления в точке равновесия пары видов. Сектора (a+b и т. д. ) есть множество точек (совокупность местообитаний), в которых «емкость среды» такова, что в них возможно сосуществование соответствующей пары видов (таких секторов шесть). Из Tilman 1982.
 2. Смена конкурентного преимущества по мере расходования лимитирующего ресурса или дополнительного поступления его в среду (Сосуществование за счет «отложенного вытеснения» )
2. Смена конкурентного преимущества по мере расходования лимитирующего ресурса или дополнительного поступления его в среду (Сосуществование за счет «отложенного вытеснения» )
 B R 2 A R 1
B R 2 A R 1
 или биотических факторов") 3. Прерывание процесса вытеснения нарушениями извне за счет абиотических (пожары, осыпи) или биотических факторов (выедание) (Сосуществование за счет «отложенного вытеснения» ; бурая водоросль и мидии; деревья в тайге)
3. Прерывание процесса вытеснения нарушениями извне за счет абиотических (пожары, осыпи) или биотических факторов (выедание) (Сосуществование за счет «отложенного вытеснения» ; бурая водоросль и мидии; деревья в тайге)
 4. Интерференция – ингибирование одних видов другими за счет специально выделяемых веществ
4. Интерференция – ингибирование одних видов другими за счет специально выделяемых веществ
 обычный (sensitive) 2) убийца (killer) 3) устойчивый (resistant) завоевывают поверхность") Три клона бактерий: 1) обычный (sensitive) 2) убийца (killer) 3) устойчивый (resistant) завоевывают поверхность питательной среды (один ресурс)
Три клона бактерий: 1) обычный (sensitive) 2) убийца (killer) 3) устойчивый (resistant) завоевывают поверхность питательной среды (один ресурс)
 данных штаммов: 1) Sensitive - r 2) Resistant") Скорость популяционного роста ( r ) данных штаммов: 1) Sensitive - r 2) Resistant – ½ r 3) Killer – ¼ r
Скорость популяционного роста ( r ) данных штаммов: 1) Sensitive - r 2) Resistant – ½ r 3) Killer – ¼ r
 Rock - scissors - paper Камень-ножницы-бумага
Rock - scissors - paper Камень-ножницы-бумага
 обычного (S) устойчивому (R) Обычный (S) устойчивого киллеру") Штамм: Выигрывает у: Проигрывает: Киллер (K) обычного (S) устойчивому (R) Обычный (S) устойчивого киллеру (R) (K) Устойчивый (R) киллера (K) обычному (S)
Штамм: Выигрывает у: Проигрывает: Киллер (K) обычного (S) устойчивому (R) Обычный (S) устойчивого киллеру (R) (K) Устойчивый (R) киллера (K) обычному (S)
 5. Конвергенция разных видов, направленная на возрастание экологического сходства
5. Конвергенция разных видов, направленная на возрастание экологического сходства
 Сосуществующие виды могут быть ЭКОЛОГИЧЕСКИ БЛИЗКИМИ. Это сходство может быть результатом отбора, протекавшего в сходных условиях и приведшего к конвергенции, а не дивергенции ниш
Сосуществующие виды могут быть ЭКОЛОГИЧЕСКИ БЛИЗКИМИ. Это сходство может быть результатом отбора, протекавшего в сходных условиях и приведшего к конвергенции, а не дивергенции ниш
. На") Часть страницы из журнала Nature V. 417. P. 480 -481 (30 May 2002). На фото: слева - Graham Bell, справа - Stephen Hubbell
Часть страницы из журнала Nature V. 417. P. 480 -481 (30 May 2002). На фото: слева - Graham Bell, справа - Stephen Hubbell
 Гипотеза нейтральности Стивена Хаббела
Гипотеза нейтральности Стивена Хаббела
 Начало редакционной статьи Nature от 30 мая 2002 г.
Начало редакционной статьи Nature от 30 мая 2002 г.
 Гипотеза нейтральности Стивена Хаббела Виды сосуществуют потому, что в расчете на одну особь имеют примерно равные вероятности рождения, гибели и миграции Экологическая идентичность (или точнее - экологическая эквивалентность) видов, проявляется в сходстве их демографических характеристик
Гипотеза нейтральности Стивена Хаббела Виды сосуществуют потому, что в расчете на одну особь имеют примерно равные вероятности рождения, гибели и миграции Экологическая идентичность (или точнее - экологическая эквивалентность) видов, проявляется в сходстве их демографических характеристик
 Хаббел и его сторонники вовсе не отрицают того, что разные виды в сообществе могут занимать разные ниши. Они только замечают, что есть и другой способ достичь сосуществования, — быть максимально похожими по демографическим характеристикам, иметь сходные удельные (в расчете на одну особь) скорости популяционного роста и заселения освободившихся участков
Хаббел и его сторонники вовсе не отрицают того, что разные виды в сообществе могут занимать разные ниши. Они только замечают, что есть и другой способ достичь сосуществования, — быть максимально похожими по демографическим характеристикам, иметь сходные удельные (в расчете на одну особь) скорости популяционного роста и заселения освободившихся участков
 Нейтральная модель не отрицает существования разных экологических ниш. Но она утверждает, что предположение о расхождении ВСЕХ видов по разным нишам не является обязательным, чтобы объяснить их сосуществование. Предположение о НЕЙТРАЛЬНОСТИ должно приниматься как нуль-гипотеза при анализе всех многовидовых сообществ.
Нейтральная модель не отрицает существования разных экологических ниш. Но она утверждает, что предположение о расхождении ВСЕХ видов по разным нишам не является обязательным, чтобы объяснить их сосуществование. Предположение о НЕЙТРАЛЬНОСТИ должно приниматься как нуль-гипотеза при анализе всех многовидовых сообществ.
 и относительной частотой встречаемости") Предполагаемые взаимосвязи между средней удельной скоростью роста популяции (ось Y) и относительной частотой встречаемости того же вида в сообществе (ось Х). Peter B. Adler, Janneke Hille, Ris Lambers and Jonathan M. Levine. A niche for neutrality // Ecology Letters. 2007. V. 10 (2). P. 95– 104
Предполагаемые взаимосвязи между средней удельной скоростью роста популяции (ось Y) и относительной частотой встречаемости того же вида в сообществе (ось Х). Peter B. Adler, Janneke Hille, Ris Lambers and Jonathan M. Levine. A niche for neutrality // Ecology Letters. 2007. V. 10 (2). P. 95– 104
 Сходство дем хар-к Модель, предложенная в работе: Peter B. Adler, Janneke Hille, Ris Lambers and Jonathan M. Levine. A niche for neutrality // Ecology Letters. 2007. V. 10 (2). P. 95– 104 Разделение ниш
Сходство дем хар-к Модель, предложенная в работе: Peter B. Adler, Janneke Hille, Ris Lambers and Jonathan M. Levine. A niche for neutrality // Ecology Letters. 2007. V. 10 (2). P. 95– 104 Разделение ниш
 и степени разделения ниш") Рисунок показывает, как комбинация сходства демографических характеристик видов (ось Y) и степени разделения ниш (ось Х) может обеспечивать сосуществование видов. Разделение ниш характеризуется «степенью стабилизации» . Чем более схожи виды по скорости популяционного роста (значения по оси Y ближе к 1), тем меньше потребность в стабилизирующем механизме для достижения сосуществования. Если же виды по скорости популяционного роста сильно различаются (значения по оси Y ближе к 0), то их сосуществование возможно только за счет различия ниш (большей стабилизации). Область сосуществования — правая верхняя часть поля графика, а область конкурентного исключения — нижняя левая. Ситуации, складывающиеся при разном сочетании величин, отложенных по осям, отражены на графиках, вынесенных на поля. Механизму нейтральности соответствуют точки, лежащие в левой верхней части графика, а механизму разделения ниш — в правой нижней.
Рисунок показывает, как комбинация сходства демографических характеристик видов (ось Y) и степени разделения ниш (ось Х) может обеспечивать сосуществование видов. Разделение ниш характеризуется «степенью стабилизации» . Чем более схожи виды по скорости популяционного роста (значения по оси Y ближе к 1), тем меньше потребность в стабилизирующем механизме для достижения сосуществования. Если же виды по скорости популяционного роста сильно различаются (значения по оси Y ближе к 0), то их сосуществование возможно только за счет различия ниш (большей стабилизации). Область сосуществования — правая верхняя часть поля графика, а область конкурентного исключения — нижняя левая. Ситуации, складывающиеся при разном сочетании величин, отложенных по осям, отражены на графиках, вынесенных на поля. Механизму нейтральности соответствуют точки, лежащие в левой верхней части графика, а механизму разделения ниш — в правой нижней.
") 6. Мутуалистические взаимоотношения между видами, иногда опосредованные (например, через микоризу)
6. Мутуалистические взаимоотношения между видами, иногда опосредованные (например, через микоризу)
 Net transfer of carbon between ectomycorrhizal tree species in the field SUZANNE W. SIMARD et al. , // Nature V. 388, pp. 579 - 582 (07 August 1997)
Net transfer of carbon between ectomycorrhizal tree species in the field SUZANNE W. SIMARD et al. , // Nature V. 388, pp. 579 - 582 (07 August 1997)
 и псевдотсуги") Сьюзан Симард и ее коллеги экспериментировали с саженцами березы бумажной (Betula papyrifera) и псевдотсуги Мензиса (Pseudotsuga menziesii), которые высаживались смешанными группами, так, что расстояния между растениями составляли около 0, 5 м. Опыты проводили в течение двух сезонов с деревьями, соответственно, 2=х и 3=х летнего возраста. Для того, чтобы выяснить, возможен ли обмен веществом между этими растениями, им давали порцию диоксида углерода СО 2, меченного для одного вида изотопом 13 С, а для другого - 14 С. Данную процедуру проводили, заключая отдельные растения в пластиковые мешки и подавая в них СО 2, содержащий тот или иной изотоп. Через 1, 5 недели после "подкормки" растений СО 2 их изымали и оценивали содержание каждого из использованных изотопов в тканях каждого растения отдельно. В ходе эксперимента было обнаружено, что между березой и псевдотсугой происходил взаимный обмен продуктами фотосинтеза, причем если в первый год он был примерно одинаков в обоих направлениях, то во второй год псевдотсуга была в явном выигрыше, получая от 2, 5 до 10% больше углерода, фиксированного обоими растениями. В некоторых вариантах опыта псевдотсугу до подкормки СО 2 в течение 4 -6 недель выдерживали в затенении, и именно в этом случае особенно возрастало потребление ею углерода, фиксированного березой. Результаты данного исследования невольно порождают ряд вопросов. Прежде всего, - можно ли рассматривать обмен продуктами фотосинтеза как свидетельство взаимопомощи между деревьями разных видов? Если да, то как можно объяснить его возникновение в процессе эволюции, не вступая в противоречие с современным дарвинизмом и концепцией "эгоистичного гена"? Один из вариантов объяснения предполагает, что такой обмен есть всего-навсего побочный продукт коэволюции высших растений и грибов, а некоторая часть продукции, отдаваемая одним растением другому (который, возможно, является конкурентом), - это своего рода незначительная плата за громадные преимущества, предоставляемые микоризой. Не следует сбрасывать со счета и то, что донор может превратиться в рецепиента, если условия его существования ухудшаться.
Сьюзан Симард и ее коллеги экспериментировали с саженцами березы бумажной (Betula papyrifera) и псевдотсуги Мензиса (Pseudotsuga menziesii), которые высаживались смешанными группами, так, что расстояния между растениями составляли около 0, 5 м. Опыты проводили в течение двух сезонов с деревьями, соответственно, 2=х и 3=х летнего возраста. Для того, чтобы выяснить, возможен ли обмен веществом между этими растениями, им давали порцию диоксида углерода СО 2, меченного для одного вида изотопом 13 С, а для другого - 14 С. Данную процедуру проводили, заключая отдельные растения в пластиковые мешки и подавая в них СО 2, содержащий тот или иной изотоп. Через 1, 5 недели после "подкормки" растений СО 2 их изымали и оценивали содержание каждого из использованных изотопов в тканях каждого растения отдельно. В ходе эксперимента было обнаружено, что между березой и псевдотсугой происходил взаимный обмен продуктами фотосинтеза, причем если в первый год он был примерно одинаков в обоих направлениях, то во второй год псевдотсуга была в явном выигрыше, получая от 2, 5 до 10% больше углерода, фиксированного обоими растениями. В некоторых вариантах опыта псевдотсугу до подкормки СО 2 в течение 4 -6 недель выдерживали в затенении, и именно в этом случае особенно возрастало потребление ею углерода, фиксированного березой. Результаты данного исследования невольно порождают ряд вопросов. Прежде всего, - можно ли рассматривать обмен продуктами фотосинтеза как свидетельство взаимопомощи между деревьями разных видов? Если да, то как можно объяснить его возникновение в процессе эволюции, не вступая в противоречие с современным дарвинизмом и концепцией "эгоистичного гена"? Один из вариантов объяснения предполагает, что такой обмен есть всего-навсего побочный продукт коэволюции высших растений и грибов, а некоторая часть продукции, отдаваемая одним растением другому (который, возможно, является конкурентом), - это своего рода незначительная плата за громадные преимущества, предоставляемые микоризой. Не следует сбрасывать со счета и то, что донор может превратиться в рецепиента, если условия его существования ухудшаться.
 Растения няни Alfonso Valiente-Banuet, Miguel Verdú Facilitation can increase the phylogenetic diversity of plant communities // Ecology Letters. 2007. V. 10 (11). P. 1029– 1036 Синопсис статьи смотрите здесь: «Растения-няни помогают поддерживать высокое разнообразие растительных сообществ» (http: //elementy. ru/news/430616)
Растения няни Alfonso Valiente-Banuet, Miguel Verdú Facilitation can increase the phylogenetic diversity of plant communities // Ecology Letters. 2007. V. 10 (11). P. 1029– 1036 Синопсис статьи смотрите здесь: «Растения-няни помогают поддерживать высокое разнообразие растительных сообществ» (http: //elementy. ru/news/430616)
. Железное дерево") Скопление растенийнянь и их подопечных в пустыне в Аризоне (район Tucson Mountains). Железное дерево Olneya tesota своей кроной дало приют «зеленому дереву» , Cercidium microphyllum (на переднем плане), а оно в свою очередь является няней для проростков кактуса сагуаро (Carnegia gigantea). Впрочем, скоро кактусы перерастут своих нянь
Скопление растенийнянь и их подопечных в пустыне в Аризоне (район Tucson Mountains). Железное дерево Olneya tesota своей кроной дало приют «зеленому дереву» , Cercidium microphyllum (на переднем плане), а оно в свою очередь является няней для проростков кактуса сагуаро (Carnegia gigantea). Впрочем, скоро кактусы перерастут своих нянь

 0 Комменсализм Хищничество Детритофагия Паразитизм Нейтрализм 0 (в т.") + + Мутуализм (протокооперация, симбиоз) 0 Комменсализм Хищничество Детритофагия Паразитизм Нейтрализм 0 (в т. ч. условный) Аменсализм (аллелопатия, антибиоз) Конкуренция
+ + Мутуализм (протокооперация, симбиоз) 0 Комменсализм Хищничество Детритофагия Паразитизм Нейтрализм 0 (в т. ч. условный) Аменсализм (аллелопатия, антибиоз) Конкуренция
 Predation, parasitism, plant herbivores") Хищничество (эксплуатация, фитофагия, паразитизм) Predation, parasitism, plant herbivores
Хищничество (эксплуатация, фитофагия, паразитизм) Predation, parasitism, plant herbivores
 ОПРЕДЕЛЕНИЕ • Хищничество – взаимодействие в ходе которого видэксплуататор использует живых особей другого вида в качестве пищевого ресурса. Это приводит к обратной связи их организменных или популяционных характеристик dr 1 d. N 2 0 dr 2 d. N 1 0
ОПРЕДЕЛЕНИЕ • Хищничество – взаимодействие в ходе которого видэксплуататор использует живых особей другого вида в качестве пищевого ресурса. Это приводит к обратной связи их организменных или популяционных характеристик dr 1 d. N 2 0 dr 2 d. N 1 0
 и детритофагия (включая") • Отношения «эксплуататор жертва» в широком смысле, (включая отчасти комменсализм) и детритофагия (включая редуцентов) – основные пути передачи вещества и энергии в экосистемах
• Отношения «эксплуататор жертва» в широком смысле, (включая отчасти комменсализм) и детритофагия (включая редуцентов) – основные пути передачи вещества и энергии в экосистемах
 экосистемное определение • В экосистемах хищники выполняют функции консументов всех порядков
экосистемное определение • В экосистемах хищники выполняют функции консументов всех порядков
 Варианты хищничества Число единиц ресурса потребленного за жизнь хищника Живой") Классификация хищничества (Thompson, 1982) Варианты хищничества Число единиц ресурса потребленного за жизнь хищника Живой ресурс поедается. . . Живой ресурс в результате. . . Истинные хищники много Целиком, связь не тесная гибнет Пастбищные хищники много Частично, связь не тесная Не гибнет Паразиты Жертва одна или несколько Частично, связь тесная Обычно не гибнет Паразитоиды (взрослые) много Связь не тесная Не гибнет Паразитоиды (личинки) Жертва одна Целиком или частично Гибнет, но не сразу
Классификация хищничества (Thompson, 1982) Варианты хищничества Число единиц ресурса потребленного за жизнь хищника Живой ресурс поедается. . . Живой ресурс в результате. . . Истинные хищники много Целиком, связь не тесная гибнет Пастбищные хищники много Частично, связь не тесная Не гибнет Паразиты Жертва одна или несколько Частично, связь тесная Обычно не гибнет Паразитоиды (взрослые) много Связь не тесная Не гибнет Паразитоиды (личинки) Жертва одна Целиком или частично Гибнет, но не сразу
 Истинные хищники - львы, насекомоядные растения, планктоноядные киты, семеноядные грызуны и птицы и т. д. Пастбищные хищники - травоядные, насекомые-фитофаги, мухи-кровососы, пиявки и т. д. Паразиты - многие круглые и ленточные черви, патогенные бактерии и простейшие, тли и гусеницы, связанные с одним растением, и т. д.
Истинные хищники - львы, насекомоядные растения, планктоноядные киты, семеноядные грызуны и птицы и т. д. Пастбищные хищники - травоядные, насекомые-фитофаги, мухи-кровососы, пиявки и т. д. Паразиты - многие круглые и ленточные черви, патогенные бактерии и простейшие, тли и гусеницы, связанные с одним растением, и т. д.
 «Неэффективный хищник» X") численности видов Х 2 X *1 Х 1 X 1 (t) «Неэффективный хищник» X 2 (t) время, t
численности видов Х 2 X *1 Х 1 X 1 (t) «Неэффективный хищник» X 2 (t) время, t
 X 2 (t) Х 1 «Эффективный хищник,") численности видов Х 2 X 1 (t) X 2 (t) Х 1 «Эффективный хищник, r-стратег» время, t 0
численности видов Х 2 X 1 (t) X 2 (t) Х 1 «Эффективный хищник, r-стратег» время, t 0

 X*2 X 1(t) X*1 Х 1 X*2 время, t") Х 2 X 2(t) X*2 X 1(t) X*1 Х 1 X*2 время, t
Х 2 X 2(t) X*2 X 1(t) X*1 Х 1 X*2 время, t


 и опунция http: //en. wikipedia. org/wiki/Cactoblastis_cactorum") Кактусовая огневка (Cactoblastis cactorum) и опунция http: //en. wikipedia. org/wiki/Cactoblastis_cactorum
Кактусовая огневка (Cactoblastis cactorum) и опунция http: //en. wikipedia. org/wiki/Cactoblastis_cactorum
 X 2 X 1 X*2 X*1 X 1 X*2 X 2 время, t
X 2 X 1 X*2 X*1 X 1 X*2 X 2 время, t
 X*1 X 2(t) T t") X*2 X 1(t) X*1 X 2(t) T t
X*2 X 1(t) X*1 X 2(t) T t

 Huffaker, C. B. 1958. Experimental studies on predation: dispersion factors and predator-prey oscillations. Hilgardia 27(14): 343 -383 Predator Typhlodromus occidentalis http: //www. goodbugs. org. au/BCAs%20 available/occidentalis. htm Prey Eotetranychus sexmaculatus http: //www. uae. gov. ae/uaeagricent/qgcc/application/insects/A 1/IA 1 -52. htm
Huffaker, C. B. 1958. Experimental studies on predation: dispersion factors and predator-prey oscillations. Hilgardia 27(14): 343 -383 Predator Typhlodromus occidentalis http: //www. goodbugs. org. au/BCAs%20 available/occidentalis. htm Prey Eotetranychus sexmaculatus http: //www. uae. gov. ae/uaeagricent/qgcc/application/insects/A 1/IA 1 -52. htm


 Примеры циклических колебаний численности природных популяций Процесс, протекающий не только во времени, но и в пространстве
Примеры циклических колебаний численности природных популяций Процесс, протекающий не только во времени, но и в пространстве
 http: //www. anselm. edu/homepage/jpitocch/genbios/52 -19 a-Pop. Cycle. Hare. Lynx. Phot. jpg
http: //www. anselm. edu/homepage/jpitocch/genbios/52 -19 a-Pop. Cycle. Hare. Lynx. Phot. jpg


 Krebs et al. , 2005
Krebs et al. , 2005
") Лиственничная листовертка (Zeiraphera diniana)
Лиственничная листовертка (Zeiraphera diniana)
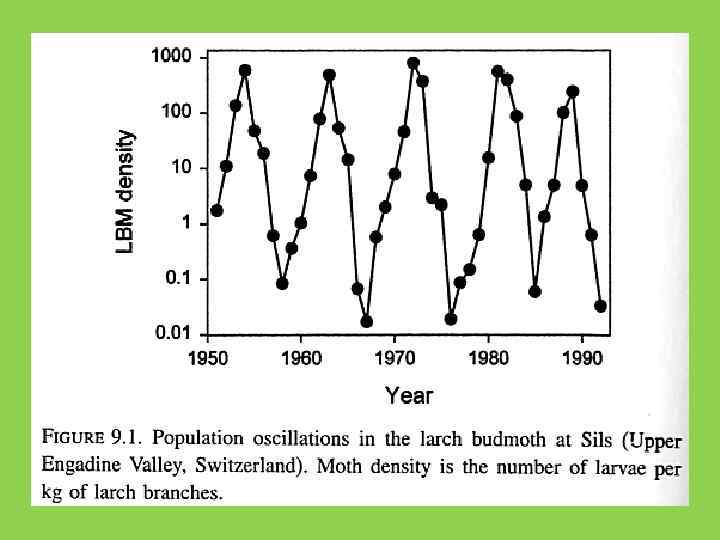

 Figure 1. Historical records of defoliation caused by the larch budmoth in the European Alps (1961 to 1998). Red dots correspond to areas with one or more years of detectable defoliation. The grid shows the location of the 20 km × 20 km aggregates used in the analyses (18). O. N. Bjø rnstad et al. , Science 298, 1020 -1023 (2002) Published by AAAS
Figure 1. Historical records of defoliation caused by the larch budmoth in the European Alps (1961 to 1998). Red dots correspond to areas with one or more years of detectable defoliation. The grid shows the location of the 20 km × 20 km aggregates used in the analyses (18). O. N. Bjø rnstad et al. , Science 298, 1020 -1023 (2002) Published by AAAS

 – численность жертвы, X 2 (t) – хищника;") Модель А. Н. Колмогорова. Х 1(t) – численность жертвы, X 2 (t) – хищника; Вi – абсолютная рождаемость, Di – абсолютная скорость элиминации: d. X 1/dt = B 1 (Х 1, Х 2) – D 1 (Х 1, Х 2) d. Х 2/dt = B 2 (Х 1, Х 2) – D 2 (Х 1, Х 2) если d. X 1/dt = f 1 (Х 1, Х 2), d. X 2/dt = f 2 (Х 1, Х 2), то обязательные условия для этих функций: df 2 /d. Х 1 > 0, df 1 /d. Х 2 < 0. Если уд. рождаемость жертвы b 1 зависит от ее плотности b 1 = b 1(X 1), то абсолютная скорость размножения жертвы: B 1 = b 1(X 1) Х 1. При Х 2 =0 cмертность жертвы зависит только от ее плотности: d 1 = d 1(X 1), при Х 2 ≠ 0 эксплуатация особей жертвы в расчете на единицу эксплуататора за единицу времени зависит от текущей численности жертвы и выражается функцией выедания - скоростью потребления пищи эксплуататором λ(Х 1). Она равна нулю при Х 1 =0 и возрастает до предельного значения с ростом плотности жертвы. Абсолютная смертность жертвы: D 1 = d 1 (Х 1) X 1 + λ(Х 1) X 2
Модель А. Н. Колмогорова. Х 1(t) – численность жертвы, X 2 (t) – хищника; Вi – абсолютная рождаемость, Di – абсолютная скорость элиминации: d. X 1/dt = B 1 (Х 1, Х 2) – D 1 (Х 1, Х 2) d. Х 2/dt = B 2 (Х 1, Х 2) – D 2 (Х 1, Х 2) если d. X 1/dt = f 1 (Х 1, Х 2), d. X 2/dt = f 2 (Х 1, Х 2), то обязательные условия для этих функций: df 2 /d. Х 1 > 0, df 1 /d. Х 2 < 0. Если уд. рождаемость жертвы b 1 зависит от ее плотности b 1 = b 1(X 1), то абсолютная скорость размножения жертвы: B 1 = b 1(X 1) Х 1. При Х 2 =0 cмертность жертвы зависит только от ее плотности: d 1 = d 1(X 1), при Х 2 ≠ 0 эксплуатация особей жертвы в расчете на единицу эксплуататора за единицу времени зависит от текущей численности жертвы и выражается функцией выедания - скоростью потребления пищи эксплуататором λ(Х 1). Она равна нулю при Х 1 =0 и возрастает до предельного значения с ростом плотности жертвы. Абсолютная смертность жертвы: D 1 = d 1 (Х 1) X 1 + λ(Х 1) X 2
 Рождаемость эксплуататора прямо пропорциональна удельной скорости потребления им пищи") Модель А. Н. Колмогорова (продолжение) Рождаемость эксплуататора прямо пропорциональна удельной скорости потребления им пищи λ(Х 1) и зависит от эффективности ее использования ν. Тогда абсолютная скорость размножения в популяции эксплуататора: B 2 = ν λ(Х 1) X 2 где ν – коэффициент эффективности (как увеличивается популяция эксплуататора при увеличении скорости потребления пищи на единицу). Если в популяции эксплуататора нет внутривидовой конкуренции и смертность эксплуататора d 2 = d 2(X 1) зависит от плотности жертвы, то абсолютная скорость элиминации эксплуататора: D 2= d 2(X 1) X 2 Тогда: d. X 1/dt = (b 1(X 1) - d 1(Х 1)) X 1 - λ(Х 1) X 2 d. X 2/ dt = (ν λ(Х 1) - d 2(X 1)) X 2 Вводя обозначения r 1 и r 2 получим: d. X 1/ dt = r 1(X 1) X 1 - λ(Х 1) X 2 d. X 2/ dt = r 2(X 1) X 2
Модель А. Н. Колмогорова (продолжение) Рождаемость эксплуататора прямо пропорциональна удельной скорости потребления им пищи λ(Х 1) и зависит от эффективности ее использования ν. Тогда абсолютная скорость размножения в популяции эксплуататора: B 2 = ν λ(Х 1) X 2 где ν – коэффициент эффективности (как увеличивается популяция эксплуататора при увеличении скорости потребления пищи на единицу). Если в популяции эксплуататора нет внутривидовой конкуренции и смертность эксплуататора d 2 = d 2(X 1) зависит от плотности жертвы, то абсолютная скорость элиминации эксплуататора: D 2= d 2(X 1) X 2 Тогда: d. X 1/dt = (b 1(X 1) - d 1(Х 1)) X 1 - λ(Х 1) X 2 d. X 2/ dt = (ν λ(Х 1) - d 2(X 1)) X 2 Вводя обозначения r 1 и r 2 получим: d. X 1/ dt = r 1(X 1) X 1 - λ(Х 1) X 2 d. X 2/ dt = r 2(X 1) X 2
 использовалась") Модель Лотки –Вольтерра. Численность хищника - Х 2, жертвы Х 1. Допущения: 1) использовалась простейшая однородная линейная функция выедания: Ψ(Х 1) =λ 1 X 1, где λ 1 = const > 0. 2) рождаемость жертвы b 1 и смертности жертвы и эксплуататора d 1, d 2 принимались постоянными положительными величинами, т. е. внутривидовая конкуренция в популяции жертвы отсутствовала. В отсутствие эксплуататора численность жертвы растет экспоненциально: d. Х 1/dt = r 1 Х 1 Поскольку жертвы выедаются хищником со скоростью пропорциональной численности обеих популяций, то с учетом функции выедания жертв хищником и без зависимости от собственной плотности, динамика популяции жертвы будет равна ее экспоненциальному росту минус потребление хищником: d. Х 1/ dt = r 1 Х 1 – λ 1 Х 1 Х 2
Модель Лотки –Вольтерра. Численность хищника - Х 2, жертвы Х 1. Допущения: 1) использовалась простейшая однородная линейная функция выедания: Ψ(Х 1) =λ 1 X 1, где λ 1 = const > 0. 2) рождаемость жертвы b 1 и смертности жертвы и эксплуататора d 1, d 2 принимались постоянными положительными величинами, т. е. внутривидовая конкуренция в популяции жертвы отсутствовала. В отсутствие эксплуататора численность жертвы растет экспоненциально: d. Х 1/dt = r 1 Х 1 Поскольку жертвы выедаются хищником со скоростью пропорциональной численности обеих популяций, то с учетом функции выедания жертв хищником и без зависимости от собственной плотности, динамика популяции жертвы будет равна ее экспоненциальному росту минус потребление хищником: d. Х 1/ dt = r 1 Х 1 – λ 1 Х 1 Х 2
 плато насыщения плотность жертвы, Х ^Х (плотность насыщения)") скорость питания хищника, λ (особь/ особь*время) плато насыщения плотность жертвы, Х ^Х (плотность насыщения)
скорость питания хищника, λ (особь/ особь*время) плато насыщения плотность жертвы, Х ^Х (плотность насыщения)
 В отсутствие жертвы (голодание) численность хищника будет снижаться экспоненциально: d.") Модель Лотки –Вольтерра (продолжение) В отсутствие жертвы (голодание) численность хищника будет снижаться экспоненциально: d. Х 2 /dt = -d 2 Х 2, где d 2 – постоянная смертность хищника. В присутствии жертвы убыль хищника начнет снижаться за счет роста его рождаемости со скоростью, которая зависит от: 1) скорости потребления жертвы (λ 1 Х 1 Х 2) и 2) эффективности перехода пищи в потомство хищника ν. Т. о. рождаемость хищника будет равна ν λ 1 Х 1 Х 2, а его динамика численности: d. Х 2 /dt = ν λ 1 Х 2 Х 1 - d 2 Х 2 Или: d. X 1/ dt = r 1 X 1 - λ 1 X 1 X 2 d. X 2/ dt = ν λ 1 X 1 X 2 - d 2 X 2
Модель Лотки –Вольтерра (продолжение) В отсутствие жертвы (голодание) численность хищника будет снижаться экспоненциально: d. Х 2 /dt = -d 2 Х 2, где d 2 – постоянная смертность хищника. В присутствии жертвы убыль хищника начнет снижаться за счет роста его рождаемости со скоростью, которая зависит от: 1) скорости потребления жертвы (λ 1 Х 1 Х 2) и 2) эффективности перехода пищи в потомство хищника ν. Т. о. рождаемость хищника будет равна ν λ 1 Х 1 Х 2, а его динамика численности: d. Х 2 /dt = ν λ 1 Х 2 Х 1 - d 2 Х 2 Или: d. X 1/ dt = r 1 X 1 - λ 1 X 1 X 2 d. X 2/ dt = ν λ 1 X 1 X 2 - d 2 X 2
 Х 2 r 1/ λ 1 Х 1 d 2/ νλ 1 d. X 1/ dt = r 1 X 1 - λ 1 X 1 X 2 = 0 Х 2 = r 1/ λ 1 Х 2 (Х 01, Х 0 2) r 1/ λ 1 Х 1 d 2/ νλ 1
Х 2 r 1/ λ 1 Х 1 d 2/ νλ 1 d. X 1/ dt = r 1 X 1 - λ 1 X 1 X 2 = 0 Х 2 = r 1/ λ 1 Х 2 (Х 01, Х 0 2) r 1/ λ 1 Х 1 d 2/ νλ 1
 Х 2 С Х*2 Х 1 Х*1
Х 2 С Х*2 Х 1 Х*1
 • Модели показывают характерную особенность систем «эксплуататор- жертва» возникновение сопряженных колебаний численностей в результате запаздывания во времени
• Модели показывают характерную особенность систем «эксплуататор- жертва» возникновение сопряженных колебаний численностей в результате запаздывания во времени
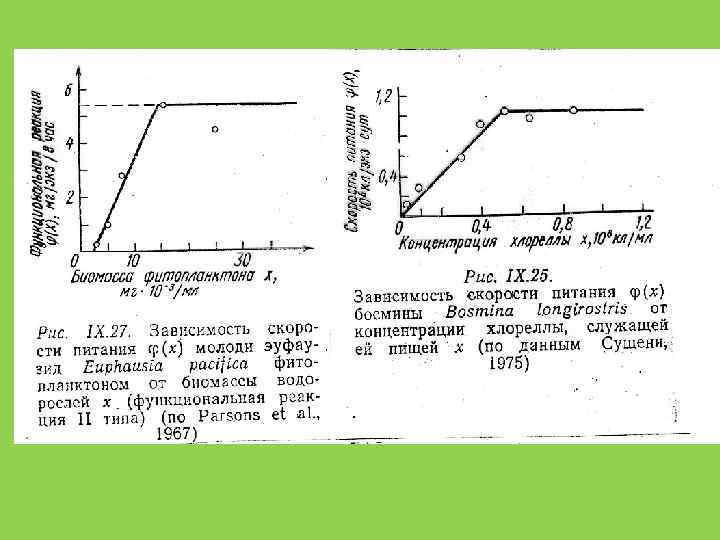
 и численная реакция (numerical response) хищника в ответ на увеличение") Функциональная реакция (functional response) и численная реакция (numerical response) хищника в ответ на увеличение численности жертв
Функциональная реакция (functional response) и численная реакция (numerical response) хищника в ответ на увеличение численности жертв
") скорость питания хищника, λ Holling, 1965 плато насыщения плотность жертвы, Х ^Х (плотность насыщения)
скорость питания хищника, λ Holling, 1965 плато насыщения плотность жертвы, Х ^Х (плотность насыщения)
") скорость питания хищника, λ плато насыщения плотность жертвы, Х ^Х (плотность насыщения)
скорость питания хищника, λ плато насыщения плотность жертвы, Х ^Х (плотность насыщения)
 скорость питания хищника, λ плато насыщения λ 1/2 λ плотность жертвы, Х β 0 К
скорость питания хищника, λ плато насыщения λ 1/2 λ плотность жертвы, Х β 0 К
 скорость питания хищника, λ плато насыщения плотность жертвы, Х
скорость питания хищника, λ плато насыщения плотность жертвы, Х

 численная реакция короткохвостой бурозубки (1), обыкновенной бурозубки (2) и") а – функциональная и (б) численная реакция короткохвостой бурозубки (1), обыкновенной бурозубки (2) и оленьей «мыши» (3) на увеличение численности коконов пилильщика
а – функциональная и (б) численная реакция короткохвостой бурозубки (1), обыкновенной бурозубки (2) и оленьей «мыши» (3) на увеличение численности коконов пилильщика
 Gilg, O. et al. Functional and numerical responses of four lemming predators in high arctic Greenland. Oikos 113, 193– 216 (2006)
Gilg, O. et al. Functional and numerical responses of four lemming predators in high arctic Greenland. Oikos 113, 193– 216 (2006)
 Gilg, O. et al. Functional and numerical responses of four lemming predators in high arctic Greenland. Oikos 113, 193– 216 (2006)
Gilg, O. et al. Functional and numerical responses of four lemming predators in high arctic Greenland. Oikos 113, 193– 216 (2006)
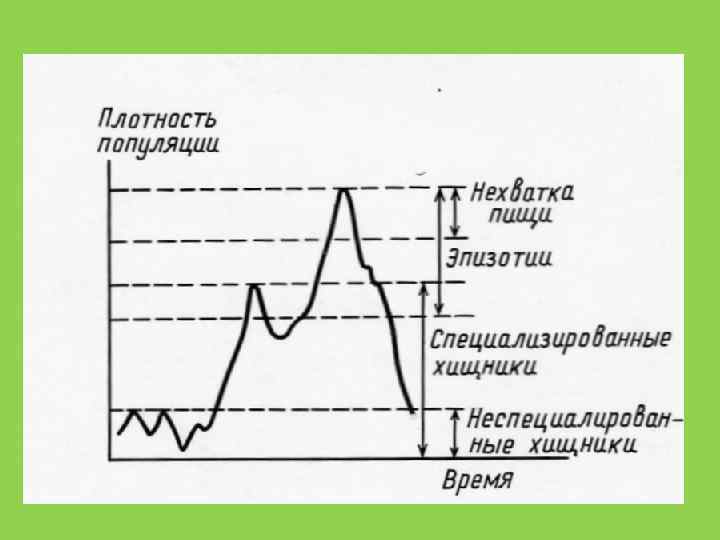
 Адаптивные эффекты в паре «хищник-жертва» • Действие хищничества может быть компенсировано снижением внутривидовой конкуренции жертв и их смертности, или ростом плодовитости жертвы • Специализация хищников (полифагия и монофагия, аггрегирующий ответ и переключение, самоограничение хищника) • Компенсация у растений
Адаптивные эффекты в паре «хищник-жертва» • Действие хищничества может быть компенсировано снижением внутривидовой конкуренции жертв и их смертности, или ростом плодовитости жертвы • Специализация хищников (полифагия и монофагия, аггрегирующий ответ и переключение, самоограничение хищника) • Компенсация у растений
 способствует разнообразию видов в сообществе за счет повышенного давления") Гипотеза Джензена-Коннелла • Хищничество (паразитизм) способствует разнообразию видов в сообществе за счет повышенного давления на жертв-доминантов
Гипотеза Джензена-Коннелла • Хищничество (паразитизм) способствует разнообразию видов в сообществе за счет повышенного давления на жертв-доминантов
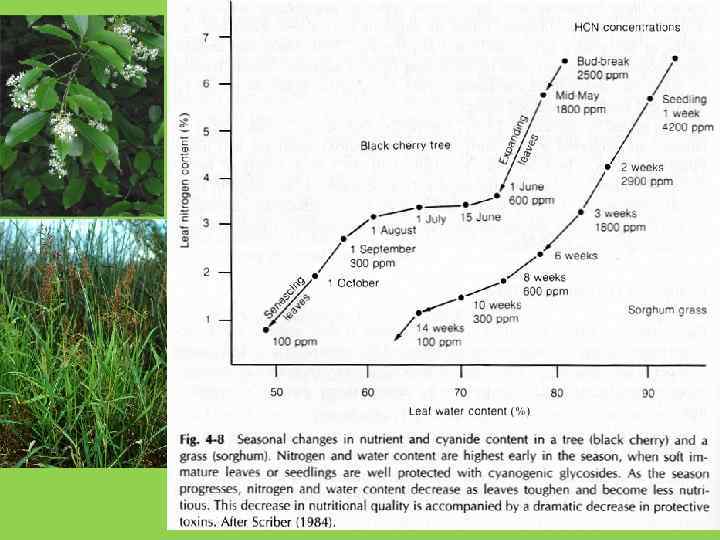
 Средства защиты растений от поедания животными-фитофагами: 1. Механические (разнообразные выросты поверхности – шипы, колючки, жгучие волоски) 2. Сложные полимеры (целлюлоза, лигнин) и кристаллы окиси кремния. Действуют в больших количествах, затрудняя переваривание пищи 3. Токсины (алкалоиды, токсичные аминокислоты, цианогены, терпеноиды и т. п. ) – убивают или отпугивают травоядных животных. Действуют в очень небольших количествах.
Средства защиты растений от поедания животными-фитофагами: 1. Механические (разнообразные выросты поверхности – шипы, колючки, жгучие волоски) 2. Сложные полимеры (целлюлоза, лигнин) и кристаллы окиси кремния. Действуют в больших количествах, затрудняя переваривание пищи 3. Токсины (алкалоиды, токсичные аминокислоты, цианогены, терпеноиды и т. п. ) – убивают или отпугивают травоядных животных. Действуют в очень небольших количествах.


 Вещества, препятствующие перевариванию Углеводный полимер Может быть переварена только с помощью кишечной микрофлоры Гемицеллюлоза Углеводный Целлюлоза (1 основной тип) полимер Может быть переварена только с помощью кишечной микрофлоры Лигнины Фенольные полимеры Связываются с белками и углеводами Фенольные полимеры Связываются с белками Минеральные кристаллы Не перевариваются (множество) Танины (множество) Кремнезем (1 основной тип)
Вещества, препятствующие перевариванию Углеводный полимер Может быть переварена только с помощью кишечной микрофлоры Гемицеллюлоза Углеводный Целлюлоза (1 основной тип) полимер Может быть переварена только с помощью кишечной микрофлоры Лигнины Фенольные полимеры Связываются с белками и углеводами Фенольные полимеры Связываются с белками Минеральные кристаллы Не перевариваются (множество) Танины (множество) Кремнезем (1 основной тип)
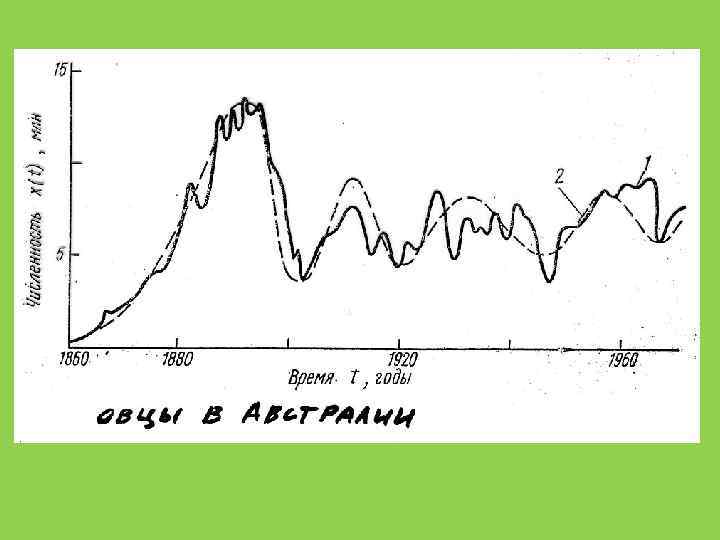
 В одном миллилитре содержимого рубца овцы содержится: 1. 61 × 1010 бактерий 106 жгутиковых 3. 3 × 105 инфузорий объем желудка овцы 6 литров коровы – около 80 литров
В одном миллилитре содержимого рубца овцы содержится: 1. 61 × 1010 бактерий 106 жгутиковых 3. 3 × 105 инфузорий объем желудка овцы 6 литров коровы – около 80 литров
 Гетероциклически Множественное; е азотсодержащие некоторые") Токсины, действующие в малых количествах (не менее 20 000) Гетероциклически Множественное; е азотсодержащие некоторые блокируют синтез ДНК или РНК соединения Токсические аминокислоты Аналоги белковых Конкурируют с белковыми аминокислотами аминокислот Алкалоиды (260) канаванин Цианогены Глюкозиды с HCN (не менее 23) Глюкозинолаты Азотсодержащие Останавливают митохондри-альное дыхание Множественное; нарушения эндокринной системы (80) соли калия Ингибиторы протеиназы Белки или полипептиды в субъединицах Связываются с активными центрами ферментов Полимеры с С 5 единицами Множественное; некоторые останавливают дыхание (неопределенно много) Терпеноиды (более 100 000)
Токсины, действующие в малых количествах (не менее 20 000) Гетероциклически Множественное; е азотсодержащие некоторые блокируют синтез ДНК или РНК соединения Токсические аминокислоты Аналоги белковых Конкурируют с белковыми аминокислотами аминокислот Алкалоиды (260) канаванин Цианогены Глюкозиды с HCN (не менее 23) Глюкозинолаты Азотсодержащие Останавливают митохондри-альное дыхание Множественное; нарушения эндокринной системы (80) соли калия Ингибиторы протеиназы Белки или полипептиды в субъединицах Связываются с активными центрами ферментов Полимеры с С 5 единицами Множественное; некоторые останавливают дыхание (неопределенно много) Терпеноиды (более 100 000)

") Клоп-солдатик (Pyrrhocoris apterus)
Клоп-солдатик (Pyrrhocoris apterus)