

Биология_ 3_Клетка.pptx
- Количество слайдов: 41
 Клетка Строение и некоторые функции
Клетка Строение и некоторые функции
 Что такое клетка? • Если бы мы полностью, во всей неисчерпаемой сложности живой клетки, узнали, наконец, что эта клетка собой представляет – из чего состоит и как работает – то многие, важнейшие и трудно разрешимые проблемы, стоящие перед человечеством, стали бы предельно просто решаемыми. К примеру – если бы мы знали, как клетка растущего под солнцем растения создаёт все вещества, необходимые для существования других, нерастительных, гетеротрофных, организмов, то нам бы не было необходимости заботиться о развитии, например, сельского хозяйства или с тревогой думать о лёгких планеты. Почти всё необходимое человеку для нормальной жизни, мы бы производили на соответствующих предприятиях, работающих на принципах функционирования растительной клетки, а заповедные участки дикой природы оставили бы лишь для прогулок. Однако на самом деле это лишь мечта, область фантастики. • Клетка неисчерпаемо сложна, и наука её изучающая – цитология – ещё не скоро, а скорее всего, никогда не поставит последней точки. По словам из доклада молодого учёного Евгения Шеваля, доступного в Интернете, учёные чуть ли не каждый день продолжают открывать новые составляющие клетки – органеллы. • Можно согласиться с утверждением, что клетка – это жизнь, а жизнь возможна только в виде клетки. Правда, ещё совсем недавно высказывались возражения, что поми-мо клетки жизнь существует в виде вирусов или, скажем, прионов, однако и они проявля-ют признаки жизни лишь внутри клетки, вне клетки их ни по каким показателям нельзя назвать живыми.
Что такое клетка? • Если бы мы полностью, во всей неисчерпаемой сложности живой клетки, узнали, наконец, что эта клетка собой представляет – из чего состоит и как работает – то многие, важнейшие и трудно разрешимые проблемы, стоящие перед человечеством, стали бы предельно просто решаемыми. К примеру – если бы мы знали, как клетка растущего под солнцем растения создаёт все вещества, необходимые для существования других, нерастительных, гетеротрофных, организмов, то нам бы не было необходимости заботиться о развитии, например, сельского хозяйства или с тревогой думать о лёгких планеты. Почти всё необходимое человеку для нормальной жизни, мы бы производили на соответствующих предприятиях, работающих на принципах функционирования растительной клетки, а заповедные участки дикой природы оставили бы лишь для прогулок. Однако на самом деле это лишь мечта, область фантастики. • Клетка неисчерпаемо сложна, и наука её изучающая – цитология – ещё не скоро, а скорее всего, никогда не поставит последней точки. По словам из доклада молодого учёного Евгения Шеваля, доступного в Интернете, учёные чуть ли не каждый день продолжают открывать новые составляющие клетки – органеллы. • Можно согласиться с утверждением, что клетка – это жизнь, а жизнь возможна только в виде клетки. Правда, ещё совсем недавно высказывались возражения, что поми-мо клетки жизнь существует в виде вирусов или, скажем, прионов, однако и они проявля-ют признаки жизни лишь внутри клетки, вне клетки их ни по каким показателям нельзя назвать живыми.
 Клетка прокариот Морфологические особенности прокариотной клетки • Науку о клетке, её ещё часто называют биологией клетки, очень старая, ей примерно три столетия. Сейчас она естественным образом делится на два крупных раздела в соответствии с характером организации генетического материала – хромосом: клетки прокариот и клетки эвкариот. • По форме клеток, организмы этого домена не столь уж разнообразны (рис. 3. 1. ), они представлены: кокками – шаровидная форма, такую форму могут иметь как археи, так и бактерии; диплококками – округлые бактерии (кокки), обычно встречающиеся парами, примерами могут служить бактерии рода Neisseria: менингококк, гонококк, а также пневмококк; стрептококки – шаровидные или овоидные (яйцеподобные) бактерии; колонии кокков; палочковидные бактерии (одиночная клетка и цепочка клеток); спириллы – спирально изогнутые клетки; бактерии, имеющие форму замкнутого или незамкнутого кольца; бактерии, образующие выросты (простеки); бактерия червеобразной формы; бактериальная клетка в форме шестиугольной звезды, а также бактерии или археи, имеющие всевозможные жгутики, нитчатые клетки или клетки с шипами. • Прокариоты сферической формы, называемые кокками, могут после деления не расходиться. Если деление происходит в одной плоскости, образуются пары клеток (ди-плококки) или цепочки (стрептококки). В том случае, когда деление происходит относи-тельно равномерно в трех взаимно перпендикулярных направлениях, и клетки после деления остаются соединенными друг с другом, возникают пакеты правильной формы (сарцины) или колонии сферической формы.
Клетка прокариот Морфологические особенности прокариотной клетки • Науку о клетке, её ещё часто называют биологией клетки, очень старая, ей примерно три столетия. Сейчас она естественным образом делится на два крупных раздела в соответствии с характером организации генетического материала – хромосом: клетки прокариот и клетки эвкариот. • По форме клеток, организмы этого домена не столь уж разнообразны (рис. 3. 1. ), они представлены: кокками – шаровидная форма, такую форму могут иметь как археи, так и бактерии; диплококками – округлые бактерии (кокки), обычно встречающиеся парами, примерами могут служить бактерии рода Neisseria: менингококк, гонококк, а также пневмококк; стрептококки – шаровидные или овоидные (яйцеподобные) бактерии; колонии кокков; палочковидные бактерии (одиночная клетка и цепочка клеток); спириллы – спирально изогнутые клетки; бактерии, имеющие форму замкнутого или незамкнутого кольца; бактерии, образующие выросты (простеки); бактерия червеобразной формы; бактериальная клетка в форме шестиугольной звезды, а также бактерии или археи, имеющие всевозможные жгутики, нитчатые клетки или клетки с шипами. • Прокариоты сферической формы, называемые кокками, могут после деления не расходиться. Если деление происходит в одной плоскости, образуются пары клеток (ди-плококки) или цепочки (стрептококки). В том случае, когда деление происходит относи-тельно равномерно в трех взаимно перпендикулярных направлениях, и клетки после деления остаются соединенными друг с другом, возникают пакеты правильной формы (сарцины) или колонии сферической формы.
 и цианобактерии (б): 1, 2 – бациллы; 3 – спирохеты;") Строение прокариот Бактерии (а) и цианобактерии (б): 1, 2 – бациллы; 3 – спирохеты; 4 – кокки; 5 – спириллы; 6 – вибрионы; 7 – стрептококки и диплококки; 8 – сарцины; 9 – нитевидные формы; 10 – жгутиковые формы; 11 – реснитчатые формы; 12 – хроококк; 13 – нить ностока; 14 – осциллатория (справа в увеличенном виде).
Строение прокариот Бактерии (а) и цианобактерии (б): 1, 2 – бациллы; 3 – спирохеты; 4 – кокки; 5 – спириллы; 6 – вибрионы; 7 – стрептококки и диплококки; 8 – сарцины; 9 – нитевидные формы; 10 – жгутиковые формы; 11 – реснитчатые формы; 12 – хроококк; 13 – нить ностока; 14 – осциллатория (справа в увеличенном виде).
 Размер и свойства прокариотных клеток • Размер различных представителей прокариотов существенно разнятся. Самая маленькая бактерия – это паразитическая микоплазма (она живет внутри клеток эвкариот) и имеет размер 0, 1 мкм. Самые большие представители прокариот видны невооруженным глазом (граница видимости – 70 -80 мкм), эта спирохета имеет длину 250 мкм. Типичный же представитель прокариот имеет размер 0, 5 мкм в ширину и 2 мкм в длину. • Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации. Структуры, расположенные снаружи от цитоплазматической мембраны (ЦПМ) – клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки – называют обычно поверхностными структурами. Термином "клеточная оболочка" часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. • У прокариотических клеток есть цитоплазматическая мембрана, также как и у эв-кариотических. У бактерий мембрана двуслойная (липидный бислой), у архей мембрана довольно часто бывает однослойной. Мембрана архей состоит из веществ, отличных от тех, из которых состоит мембрана бактерий, это одна тех особенностей архей, которая даёт основание считать, что исторически археи не могли иметь общего предка с бактериями.
Размер и свойства прокариотных клеток • Размер различных представителей прокариотов существенно разнятся. Самая маленькая бактерия – это паразитическая микоплазма (она живет внутри клеток эвкариот) и имеет размер 0, 1 мкм. Самые большие представители прокариот видны невооруженным глазом (граница видимости – 70 -80 мкм), эта спирохета имеет длину 250 мкм. Типичный же представитель прокариот имеет размер 0, 5 мкм в ширину и 2 мкм в длину. • Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации. Структуры, расположенные снаружи от цитоплазматической мембраны (ЦПМ) – клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки – называют обычно поверхностными структурами. Термином "клеточная оболочка" часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. • У прокариотических клеток есть цитоплазматическая мембрана, также как и у эв-кариотических. У бактерий мембрана двуслойная (липидный бислой), у архей мембрана довольно часто бывает однослойной. Мембрана архей состоит из веществ, отличных от тех, из которых состоит мембрана бактерий, это одна тех особенностей архей, которая даёт основание считать, что исторически археи не могли иметь общего предка с бактериями.
 Свойства клеток прокариот • Клеточное ядро, такое, как у эвкариот, у прокариот отсутствует. ДНК находится внутри клетки, упорядоченно свернутая и поддерживаемая белками, но этот компартмент не отделён от остального протопласта мембраной. Этот ДНКбелковый комплекс называется нуклеоид. У бактерий белки, которые поддерживают ДНК, отличаются от гистонов, которые образуют нуклеосомы у эвкариот. А у архей гистоны есть, и этим они похожи на эвкариот. Внутри клетки могут находиться газовые пузырьки, запасные вещества в виде гранул полифосфатов, гранул углеводов, жировых капель. Могут присутствовать включения серы (образующейся, например, в результате бескислородного фотосинтеза). У фотосинтетических бактерий имеются складчатые структуры, называемые тилакоидами, на которых идет фотосинтез. Таким образом, у прокариот, в принципе, имеются те же самые элементы, что и у эвкариотов, но без перегородок, без внутренних мембран. Те перегородки, которые имеются, являются выростами клеточной мембраны. • Клеточная стенка • Под электронным микроскопом клеточная стенка грамположительных бактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм.
Свойства клеток прокариот • Клеточное ядро, такое, как у эвкариот, у прокариот отсутствует. ДНК находится внутри клетки, упорядоченно свернутая и поддерживаемая белками, но этот компартмент не отделён от остального протопласта мембраной. Этот ДНКбелковый комплекс называется нуклеоид. У бактерий белки, которые поддерживают ДНК, отличаются от гистонов, которые образуют нуклеосомы у эвкариот. А у архей гистоны есть, и этим они похожи на эвкариот. Внутри клетки могут находиться газовые пузырьки, запасные вещества в виде гранул полифосфатов, гранул углеводов, жировых капель. Могут присутствовать включения серы (образующейся, например, в результате бескислородного фотосинтеза). У фотосинтетических бактерий имеются складчатые структуры, называемые тилакоидами, на которых идет фотосинтез. Таким образом, у прокариот, в принципе, имеются те же самые элементы, что и у эвкариотов, но без перегородок, без внутренних мембран. Те перегородки, которые имеются, являются выростами клеточной мембраны. • Клеточная стенка • Под электронным микроскопом клеточная стенка грамположительных бактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм.
 Строение грамположительной и грамотрицательной бактерии Пептидогликан— гетерополимер Важнейший компонент клеточной стенки бактерий, выполняющий механические функции, осмотической защиты клетки, выполняет антигенные функции. Характерен только для бактерий.
Строение грамположительной и грамотрицательной бактерии Пептидогликан— гетерополимер Важнейший компонент клеточной стенки бактерий, выполняющий механические функции, осмотической защиты клетки, выполняет антигенные функции. Характерен только для бактерий.
 Строение и свойства клеток прокариот • В состав клеточной стенки бактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных бактерий он составляет основную массу вещества клеточной стенки (от 40 до 90 %), у грамотрицательных – содержание пептидогликана значительно меньше (1 -10 %). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных бактерий, содержит от 20 до 50 % этого гетерополимера. • У грамотрицательных бактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2 -3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8 -10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных бактерий получил название наружной мембраны. • Клеточная стенка грамположительных бактерий плотно прилегает к ЦПМ, в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электроннопрозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической: и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм.
Строение и свойства клеток прокариот • В состав клеточной стенки бактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных бактерий он составляет основную массу вещества клеточной стенки (от 40 до 90 %), у грамотрицательных – содержание пептидогликана значительно меньше (1 -10 %). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных бактерий, содержит от 20 до 50 % этого гетерополимера. • У грамотрицательных бактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2 -3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8 -10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных бактерий получил название наружной мембраны. • Клеточная стенка грамположительных бактерий плотно прилегает к ЦПМ, в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электроннопрозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической: и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм.
 Функции клеточной стенки прокариот • • Клеточная стенка прокариот выполняет разнообразные функции: механически защищает клетку от воздействий окружающей среды, обеспечивает поддержание ее внеш-ней формы, дает возможность клетке существовать в гипотонических растворах. В первую очередь в этом "заслуга" пептидогликана. Структурная дифференцировка клеточной стенки у грамотрицательных эубактерий, приведшая к формированию дополнительного слоя в виде наружной мембраны, зна-чительно расширила круг функций клеточной стенки. Прежде всего, это связано с проблемами проницаемости и транспорта веществ в клетку. Наружная мембрана имеет специфические и неспецифические каналы (поры) для пассивного транспорта веществ и ионов, необходимых клетке, т. е. осуществляет функции молекулярного "сита". Наружная мембрана также препятствует проникновению в клетку токсических веществ, что находит отражение в большей устойчивости грамотрицательных эубактерий (сравнительно с грамположительными) к действию некоторых ядов, химических веществ, ферментов и антибиотиков. Появление у грамотрицательных эубактерий дополнительной мембраны в составе клеточной стенки фактически привело к созданию обособленной полости (периплазмати-ческого пространства), отграниченной от цитоплазмы и внешней среды специфическими мембранами и несущей важную функциональную нагрузку.
Функции клеточной стенки прокариот • • Клеточная стенка прокариот выполняет разнообразные функции: механически защищает клетку от воздействий окружающей среды, обеспечивает поддержание ее внеш-ней формы, дает возможность клетке существовать в гипотонических растворах. В первую очередь в этом "заслуга" пептидогликана. Структурная дифференцировка клеточной стенки у грамотрицательных эубактерий, приведшая к формированию дополнительного слоя в виде наружной мембраны, зна-чительно расширила круг функций клеточной стенки. Прежде всего, это связано с проблемами проницаемости и транспорта веществ в клетку. Наружная мембрана имеет специфические и неспецифические каналы (поры) для пассивного транспорта веществ и ионов, необходимых клетке, т. е. осуществляет функции молекулярного "сита". Наружная мембрана также препятствует проникновению в клетку токсических веществ, что находит отражение в большей устойчивости грамотрицательных эубактерий (сравнительно с грамположительными) к действию некоторых ядов, химических веществ, ферментов и антибиотиков. Появление у грамотрицательных эубактерий дополнительной мембраны в составе клеточной стенки фактически привело к созданию обособленной полости (периплазмати-ческого пространства), отграниченной от цитоплазмы и внешней среды специфическими мембранами и несущей важную функциональную нагрузку.
 Капсулы, слизистые слои и чехлы • Снаружи клеточная стенка прокариот часто бывает окружена слизистым веще -ством. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоев или чехлов. • Под капсулой понимают слизистое образование, обволакивающее клетку, сохраняющее связь с клеточной стенкой и имеющее аморфное строение. Если толщина образования меньше 0, 2 мкм и, следовательно, оно может быть обнаружено только с помощью электронного микроскопа, то говорят о микрокапсуле. Если больше 0, 2 мкм, говорят о макрокапсуле. • В отличие от капсул чехлы имеют тонкую структуру. Нередко в них обнаруживают несколько слоев с разным строением. Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами. Между этими структурами у прокариот обнаружено много переходных форм, так что иногда нельзя четко отграничивать капсулу от слизистых клеточных выделений или капсулу от чехла. • На клеточной поверхности многих прокариот имеются структуры, определяющие способность клетки к движению в жидкой среде. Это – жгутики. Если жгутики находятся у полюсов или в полярной области клетки, говорят об их полярном или субполярном расположении, если – вдоль боковой поверхности, говорят о латеральном расположении, число жгутиков может достигать 1000 на клетку.
Капсулы, слизистые слои и чехлы • Снаружи клеточная стенка прокариот часто бывает окружена слизистым веще -ством. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоев или чехлов. • Под капсулой понимают слизистое образование, обволакивающее клетку, сохраняющее связь с клеточной стенкой и имеющее аморфное строение. Если толщина образования меньше 0, 2 мкм и, следовательно, оно может быть обнаружено только с помощью электронного микроскопа, то говорят о микрокапсуле. Если больше 0, 2 мкм, говорят о макрокапсуле. • В отличие от капсул чехлы имеют тонкую структуру. Нередко в них обнаруживают несколько слоев с разным строением. Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами. Между этими структурами у прокариот обнаружено много переходных форм, так что иногда нельзя четко отграничивать капсулу от слизистых клеточных выделений или капсулу от чехла. • На клеточной поверхности многих прокариот имеются структуры, определяющие способность клетки к движению в жидкой среде. Это – жгутики. Если жгутики находятся у полюсов или в полярной области клетки, говорят об их полярном или субполярном расположении, если – вдоль боковой поверхности, говорят о латеральном расположении, число жгутиков может достигать 1000 на клетку.
 Поверхностные структуры • Большие успехи достигнуты в расшифровке механизма движения прокариот, имеющих жгутики. Если в клетке много жгутиков, все они при движении собираются в пучок, вращаясь в одном направлении. Вращение жгутиков передается клетке и обеспечивает эффективное движение (плавание) в жидкой среде и более медленное перемещение по поверхности твердых сред. • Подвижные бактерии активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие направленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай – аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис. • К поверхностным структурам бактериальной клетки относятся также ворсинки (фимбрии, пили). Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм. Ворсинки построены из одного вида белка – пилина – и представляют собой прямые белковые цилиндры, отходящие от поверхности клетки. Они, как правило, тоньше жгутиков (диаметр в пределах 5 – 10 нм, длина 0, 2 – 2, 0 мкм), расположены на боковых поверхностях клеток или полярно. Ворсинки общего типа придают бактериям свойство гидрофобности, обеспечивают их прикрепление к клеткам растений, грибов и неорганическим частицам, принимают участие в транспорте метаболитов. Через ворсинки в клетку могут проникать вирусы.
Поверхностные структуры • Большие успехи достигнуты в расшифровке механизма движения прокариот, имеющих жгутики. Если в клетке много жгутиков, все они при движении собираются в пучок, вращаясь в одном направлении. Вращение жгутиков передается клетке и обеспечивает эффективное движение (плавание) в жидкой среде и более медленное перемещение по поверхности твердых сред. • Подвижные бактерии активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие направленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай – аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис. • К поверхностным структурам бактериальной клетки относятся также ворсинки (фимбрии, пили). Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм. Ворсинки построены из одного вида белка – пилина – и представляют собой прямые белковые цилиндры, отходящие от поверхности клетки. Они, как правило, тоньше жгутиков (диаметр в пределах 5 – 10 нм, длина 0, 2 – 2, 0 мкм), расположены на боковых поверхностях клеток или полярно. Ворсинки общего типа придают бактериям свойство гидрофобности, обеспечивают их прикрепление к клеткам растений, грибов и неорганическим частицам, принимают участие в транспорте метаболитов. Через ворсинки в клетку могут проникать вирусы.
 – обязательным структурным") Мембраны • Содержимое клетки отделяется от клеточной стенки цитоплазматической мембраной (ЦПМ) – обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8 -15 % сухого вещества клеток. У большинства прокариотных клеток ЦПМ – единственная мембрана. • Химический состав мембран прокариотов • ЦПМ – белково-липидный комплекс, в котором белки составляют 50 -75 %, липиды – от 15 до 45 %. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95 % и больше вещества мембран. • Главным липидным компонентом бактериальных мембран являются фосфолипиды – производные 3 -фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. • Помимо обычных жирных кислот, т. е. обнаруживаемых и в клетках эукариот, в составе мембранных липидов эубактерий находят и кислоты, не встречающиеся, как правило, в мембранах эукариот. Набор жирных кислот в мембранных липидах также чрезвычайно видоспецифичен. Особый состав липидов обнаружен в мембранах архебактерий. У них не найдены типичные для эубактерий эфиры глицерина и жирных кислот, но присутствуют эфиры глицерина и высокомолекулярных С 20 -, С 40 -спиртов, а также нейтральные изопреноидные С 20 – С 30 -углеводороды. • На долю белков приходится больше половины сухой массы мембран. К мембранам с наиболее высоким содержанием белка относятся бактериальные ЦПМ. В некоторых бактериальных мембранах в значительных количествах обнаружены углеводы. Повидимому, они содержатся не в свободном состоянии, а входят в состав гликолипидов и гликопротеинов.
Мембраны • Содержимое клетки отделяется от клеточной стенки цитоплазматической мембраной (ЦПМ) – обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8 -15 % сухого вещества клеток. У большинства прокариотных клеток ЦПМ – единственная мембрана. • Химический состав мембран прокариотов • ЦПМ – белково-липидный комплекс, в котором белки составляют 50 -75 %, липиды – от 15 до 45 %. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95 % и больше вещества мембран. • Главным липидным компонентом бактериальных мембран являются фосфолипиды – производные 3 -фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. • Помимо обычных жирных кислот, т. е. обнаруживаемых и в клетках эукариот, в составе мембранных липидов эубактерий находят и кислоты, не встречающиеся, как правило, в мембранах эукариот. Набор жирных кислот в мембранных липидах также чрезвычайно видоспецифичен. Особый состав липидов обнаружен в мембранах архебактерий. У них не найдены типичные для эубактерий эфиры глицерина и жирных кислот, но присутствуют эфиры глицерина и высокомолекулярных С 20 -, С 40 -спиртов, а также нейтральные изопреноидные С 20 – С 30 -углеводороды. • На долю белков приходится больше половины сухой массы мембран. К мембранам с наиболее высоким содержанием белка относятся бактериальные ЦПМ. В некоторых бактериальных мембранах в значительных количествах обнаружены углеводы. Повидимому, они содержатся не в свободном состоянии, а входят в состав гликолипидов и гликопротеинов.
 Функции цитоплазматической мембраны прокариот • ЦПМ прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ. • Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза, в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии используемой затем в клетке по разным каналам, в том числе и для образования химической энергии (АТФ). ЦПМ принимает участие в репликации и последующем разделении хромосомы прокариотной клетки. • В последнее время выявляется еще одна функциональная грань клеточных мембран – их интегрирующая роль в организме, вполне сочетающаяся с давно установленной разъединяющей (барьерной) функцией.
Функции цитоплазматической мембраны прокариот • ЦПМ прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ. • Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза, в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии используемой затем в клетке по разным каналам, в том числе и для образования химической энергии (АТФ). ЦПМ принимает участие в репликации и последующем разделении хромосомы прокариотной клетки. • В последнее время выявляется еще одна функциональная грань клеточных мембран – их интегрирующая роль в организме, вполне сочетающаяся с давно установленной разъединяющей (барьерной) функцией.
 Функции мембран • Клетка – единое целое. В обеспечении этого принципа клеточной организации важная роль принадлежит мембранам. Показан перенос электрохимической энергии и электронов вдоль мембран. Последние рассматриваются так же как возможные пути транспорта жирорастворимых субстратов и молекулярного кислорода. • ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов. Осуществляется это с использованием разных механизмов мембранного транспорта. • Рис. (Справа): 1 -молекулы липидов: а-гидрофильная голова, б-гидрофобный хвост; 2 -молекулы белков: в-интегральная, г- периферическая, д-поверхностная. • (В центре) липосахарид: 1 -липид, 2 -внутреннее полисахаридное ядро, 3 наружное полисахаридное ядро, 4 -антиген. (Слева) 2 -пептидогликан, 3 -периплазматическое пространство, 5 -фосфолипид, 6 -полисахарид. Гидроф. ч. заштрих.
Функции мембран • Клетка – единое целое. В обеспечении этого принципа клеточной организации важная роль принадлежит мембранам. Показан перенос электрохимической энергии и электронов вдоль мембран. Последние рассматриваются так же как возможные пути транспорта жирорастворимых субстратов и молекулярного кислорода. • ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов. Осуществляется это с использованием разных механизмов мембранного транспорта. • Рис. (Справа): 1 -молекулы липидов: а-гидрофильная голова, б-гидрофобный хвост; 2 -молекулы белков: в-интегральная, г- периферическая, д-поверхностная. • (В центре) липосахарид: 1 -липид, 2 -внутреннее полисахаридное ядро, 3 наружное полисахаридное ядро, 4 -антиген. (Слева) 2 -пептидогликан, 3 -периплазматическое пространство, 5 -фосфолипид, 6 -полисахарид. Гидроф. ч. заштрих.
 Цитозоль и рибосомы • Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: внутрицитоплазматическими мембранами (если они есть), генетическим аппаратом, рибосомами и включениями разной химической природы и функционального назначения. • Рибосомы – место синтеза белка – рибонуклеопротеиновые частицы размером 15 -20 нм. Их число в клетке зависит от интенсивности процессов белкового синтеза и колеблется от 5000 до 90 000. Общая масса рибосом может составлять примерно 1/4 клеточной массы, а количество рибосомальной РНК (р. РНК) – 80 -85 % всей бактериальной РНК. • Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул ин -формационной и транспортных РНК и так называемыми полирибосомами, или полисома-ми. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами.
Цитозоль и рибосомы • Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: внутрицитоплазматическими мембранами (если они есть), генетическим аппаратом, рибосомами и включениями разной химической природы и функционального назначения. • Рибосомы – место синтеза белка – рибонуклеопротеиновые частицы размером 15 -20 нм. Их число в клетке зависит от интенсивности процессов белкового синтеза и колеблется от 5000 до 90 000. Общая масса рибосом может составлять примерно 1/4 клеточной массы, а количество рибосомальной РНК (р. РНК) – 80 -85 % всей бактериальной РНК. • Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул ин -формационной и транспортных РНК и так называемыми полирибосомами, или полисома-ми. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами.
 Генетический аппарат прокариотов • Строение генетического аппарата прокариотов долгое время было предметом жарких дискуссий, суть которых сводилась к тому, есть у них такое же ядро, как у эвкариот, или нет. Установлено, что генетический материал прокариотных организмов, как и эвкариотных, представлен ДНК, но имеются существенные различия в его структурной организации. У прокариотов ДНК представляет собой более или менее компактное образование, занимающее определенную область в цитоплазме и не отделенное от нее мембраной, как это имеет место у эвкариот. Чтобы подчеркнуть структурные различия в генетическом аппарате прокариотных и эвкариотных клеток, предложено у первых его называть нуклеоидом в отличие от ядра у вторых. • При электронно-микроскопическом наблюдении видно, что нуклеоид прокариот, несмотря на отсутствие ядерной мембраны, довольно четко отграничен от цитоплазмы, занимает в ней, как правило, центральную область и заполнен нитями ДНК диаметром около 2 нм. Не исключено, что на выявляемую в электронном микроскопе организацию прокариотной хромосомы большое влияние оказывают условия фиксации препарата. По имеющимся наблюдениям, в живой клетке нуклеоид занимает больше места в цитоплазме.
Генетический аппарат прокариотов • Строение генетического аппарата прокариотов долгое время было предметом жарких дискуссий, суть которых сводилась к тому, есть у них такое же ядро, как у эвкариот, или нет. Установлено, что генетический материал прокариотных организмов, как и эвкариотных, представлен ДНК, но имеются существенные различия в его структурной организации. У прокариотов ДНК представляет собой более или менее компактное образование, занимающее определенную область в цитоплазме и не отделенное от нее мембраной, как это имеет место у эвкариот. Чтобы подчеркнуть структурные различия в генетическом аппарате прокариотных и эвкариотных клеток, предложено у первых его называть нуклеоидом в отличие от ядра у вторых. • При электронно-микроскопическом наблюдении видно, что нуклеоид прокариот, несмотря на отсутствие ядерной мембраны, довольно четко отграничен от цитоплазмы, занимает в ней, как правило, центральную область и заполнен нитями ДНК диаметром около 2 нм. Не исключено, что на выявляемую в электронном микроскопе организацию прокариотной хромосомы большое влияние оказывают условия фиксации препарата. По имеющимся наблюдениям, в живой клетке нуклеоид занимает больше места в цитоплазме.
 Генетический аппарат 2 • Считалось, что у всех прокариотов вся наследственная информация организована в виде одной, замкнутой в кольцо хромосомы. Однако новые исследования внесли в это представление существенные поправки: «Азбучная истина гласит, что прокариоты имеют гаплоидные, простые кольцевые хромосомы, в то время как у геномов эукариот, сильно различающихся по плоидности, гены распределены между множеством линейных хромо-сом. И хотя такие геномные формы, по-видимому, действительно доминируют, на самом деле разнообразие геномов выходит далеко за рамки такого простого дихотомического разделения. В частности, у многих прокариот имеется несколько хромосом, в отдельных случаях – линейных. Более того, вопреки распространённому заблуждению, у прокариот большинство клеток не гаплоидные, то есть они содержат несколько копий генома» . Длина молекулы ДНК в развернутом виде может составлять более 1 мм, т. е. почти в 1000 раз превышать длину самой бактериальной клетки. Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Однако если исходить из того, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хромосом по дочерним клеткам. Специальные исследования показали, что на самом деле хромосомы прокариот представляют собой высокоупорядоченную структуру. В обеспечении суперспирализованной организации хромосом участвуют молекулы РНК.
Генетический аппарат 2 • Считалось, что у всех прокариотов вся наследственная информация организована в виде одной, замкнутой в кольцо хромосомы. Однако новые исследования внесли в это представление существенные поправки: «Азбучная истина гласит, что прокариоты имеют гаплоидные, простые кольцевые хромосомы, в то время как у геномов эукариот, сильно различающихся по плоидности, гены распределены между множеством линейных хромо-сом. И хотя такие геномные формы, по-видимому, действительно доминируют, на самом деле разнообразие геномов выходит далеко за рамки такого простого дихотомического разделения. В частности, у многих прокариот имеется несколько хромосом, в отдельных случаях – линейных. Более того, вопреки распространённому заблуждению, у прокариот большинство клеток не гаплоидные, то есть они содержат несколько копий генома» . Длина молекулы ДНК в развернутом виде может составлять более 1 мм, т. е. почти в 1000 раз превышать длину самой бактериальной клетки. Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Однако если исходить из того, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хромосом по дочерним клеткам. Специальные исследования показали, что на самом деле хромосомы прокариот представляют собой высокоупорядоченную структуру. В обеспечении суперспирализованной организации хромосом участвуют молекулы РНК.
 Рост и способы размножения прокариот • Под ростом прокариотной клетки понимают согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регуляторным контролем, и приводит к увеличению массы (а следовательно, и размеров) клетки. Но рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению. • Для подавляющего большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток. При таком способе деления имеет место симметрия в отношении продольной и поперечной оси. У большинства грамположительных эубактерий и нитчатых цианобактерий деление происходит путем синтеза поперечной перегородки, идущего от периферии к центру. • Вариантом бинарного деления является почкование, которое можно рассматривать как неравновеликое бинарное деление. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная стенка почки полностью синтезируется заново.
Рост и способы размножения прокариот • Под ростом прокариотной клетки понимают согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регуляторным контролем, и приводит к увеличению массы (а следовательно, и размеров) клетки. Но рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению. • Для подавляющего большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток. При таком способе деления имеет место симметрия в отношении продольной и поперечной оси. У большинства грамположительных эубактерий и нитчатых цианобактерий деление происходит путем синтеза поперечной перегородки, идущего от периферии к центру. • Вариантом бинарного деления является почкование, которое можно рассматривать как неравновеликое бинарное деление. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная стенка почки полностью синтезируется заново.
 Клетка эвкариот Цитата из книги Евгения Кунина • • Из седьмой главы книги Евгения Кунина «Логика случая» . «Организмы с большими, сложными клетками известны как эукариоты, что означает «обладающие настоящим ядром» …Обычно эукариотические клетки крупнее прокариотических и обладают сложной внутриклеточной организацией с разнообразными ограниченными мембраной органеллами, включая ядро, давшее им имя, и митохондрии, произошедшие от эндосимбиотических бактерий. Таким образом, по любому разумному критерию, эукариотические клетки значительно более сложны, чем бактерии и археи. Возникновение такого уровня сложности – загадка эволюционной биологии, представляющаяся ещё более интригующей вследствие особого «личного» интереса. Ведь изучая происхождение эукариот, мы всматриваемся в исток нашего собственного происхождения. Фундаментальные различия в клеточной организации между эукариотами и прокариотами являют собой, можно сказать, парадоксальное противопоставление особенностям организации генома, …. В то время как эукариотические клетки обладают гораздо более замысловатой, упорядоченной и сложной организацией, чем прокариотические клетки, геномы эукариот намного менее оптимизированы и содержат гораздо больше «мусора» , чем геномы прокариот. Объяснение эволюционных основ этого явного парадокса представляет собой принципиальную проблему, и её решение, вероятно, прольёт свет также и на происхождение эукариот – или, точнее говоря, на происхождение клеточной организации эукариот. Эта проблема далека от решения и остаётся предметом жарких (ингда даже слишком жарких) споров» (Кунин, 2014).
Клетка эвкариот Цитата из книги Евгения Кунина • • Из седьмой главы книги Евгения Кунина «Логика случая» . «Организмы с большими, сложными клетками известны как эукариоты, что означает «обладающие настоящим ядром» …Обычно эукариотические клетки крупнее прокариотических и обладают сложной внутриклеточной организацией с разнообразными ограниченными мембраной органеллами, включая ядро, давшее им имя, и митохондрии, произошедшие от эндосимбиотических бактерий. Таким образом, по любому разумному критерию, эукариотические клетки значительно более сложны, чем бактерии и археи. Возникновение такого уровня сложности – загадка эволюционной биологии, представляющаяся ещё более интригующей вследствие особого «личного» интереса. Ведь изучая происхождение эукариот, мы всматриваемся в исток нашего собственного происхождения. Фундаментальные различия в клеточной организации между эукариотами и прокариотами являют собой, можно сказать, парадоксальное противопоставление особенностям организации генома, …. В то время как эукариотические клетки обладают гораздо более замысловатой, упорядоченной и сложной организацией, чем прокариотические клетки, геномы эукариот намного менее оптимизированы и содержат гораздо больше «мусора» , чем геномы прокариот. Объяснение эволюционных основ этого явного парадокса представляет собой принципиальную проблему, и её решение, вероятно, прольёт свет также и на происхождение эукариот – или, точнее говоря, на происхождение клеточной организации эукариот. Эта проблема далека от решения и остаётся предметом жарких (ингда даже слишком жарких) споров» (Кунин, 2014).
 • • • Под «мусором» Е. Кунин подразумевает повторения нуклеотидных последовательностей") Клетка эвкариот (продолжение) • • • Под «мусором» Е. Кунин подразумевает повторения нуклеотидных последовательностей и большую долю интронов в геноме эвкариот. Попытаемся словесно и с помощью графических изображений составить принципиальные схемы эвкариотических клеток. Их должно быть несколько, ибо единой для всех эвкариот схемы быть не может из-за принципиальных различий в строении клеток грибов, растений, животных, протистов и хлористов. Исходя из того, что нельзя объять необъятное, остановимся на двух – клетке животных и клетке растений. Клетка растений и животных обязательно включает: поверхностный аппарат, цитоплазму и ядро. Поверхностный аппарат имеет: мембрану, надмембранный и субмембранный ком-плексы. Клеточная мембрана, она же цитолемма, плазмалемма, или цитоплазматическая мембрана (ЦПМ) – эластическая молекулярная структура, состоящая из белков и липидов. ЦПМ отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой. Внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки – компартменты или органеллы, в которых поддерживаются определённые условия среды.
Клетка эвкариот (продолжение) • • • Под «мусором» Е. Кунин подразумевает повторения нуклеотидных последовательностей и большую долю интронов в геноме эвкариот. Попытаемся словесно и с помощью графических изображений составить принципиальные схемы эвкариотических клеток. Их должно быть несколько, ибо единой для всех эвкариот схемы быть не может из-за принципиальных различий в строении клеток грибов, растений, животных, протистов и хлористов. Исходя из того, что нельзя объять необъятное, остановимся на двух – клетке животных и клетке растений. Клетка растений и животных обязательно включает: поверхностный аппарат, цитоплазму и ядро. Поверхностный аппарат имеет: мембрану, надмембранный и субмембранный ком-плексы. Клеточная мембрана, она же цитолемма, плазмалемма, или цитоплазматическая мембрана (ЦПМ) – эластическая молекулярная структура, состоящая из белков и липидов. ЦПМ отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой. Внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки – компартменты или органеллы, в которых поддерживаются определённые условия среды.
") Строение клеток растений и животных • • Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляют собой так называемые сложные липиды – фосфолипиды. Молекулы липидов имеют гидрофильную ( «головка» ) и гидрофобную ( «хвост» ) части. При образовании мембран гидрофобные участки молекул обращены внутрь, а гидрофильные – наружу. Мембраны разных организмов весьма сходны по строению. Исключение составляют, пожалуй, только археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7 -8 нм. Биологическая мембрана включает также белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
Строение клеток растений и животных • • Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляют собой так называемые сложные липиды – фосфолипиды. Молекулы липидов имеют гидрофильную ( «головка» ) и гидрофобную ( «хвост» ) части. При образовании мембран гидрофобные участки молекул обращены внутрь, а гидрофильные – наружу. Мембраны разных организмов весьма сходны по строению. Исключение составляют, пожалуй, только археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7 -8 нм. Биологическая мембрана включает также белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
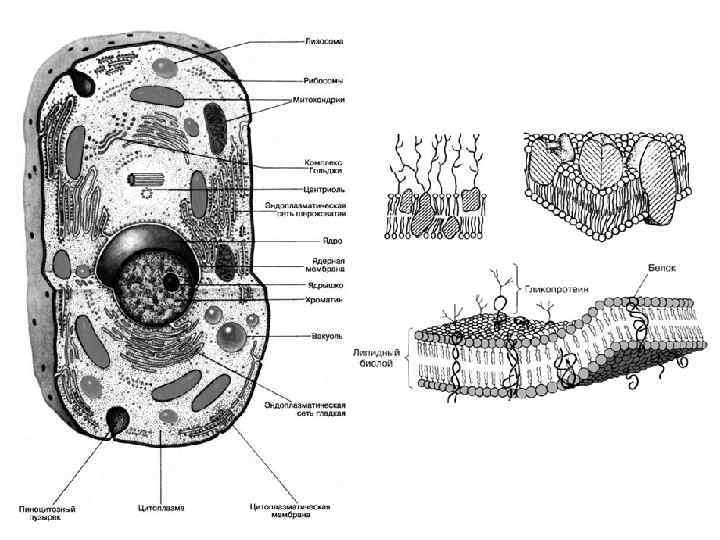
 Надмембранный комплекс • • Надмембранный комплекс – гликокаликс содержит углеводы и белки. У эвкариот универсальным компонентом надмембранного комплекса являются углеводы – компоненты гликолипидов и гликопротеинов плазмалеммы. Благодаря этому его исходно называли гликокаликсом (от греч. гликос – сладкий, углевод и лат. каллум – толстая кожа, оболочка). Кроме углеводов, в состав гликокаликса включают периферические белки над билипидным слоем. Более сложные варианты надмембранного комплекса встречаются у растений (клеточная стенка из целлюлозы), грибов и членистоногих (наружный покров из хитина). Клеточная стенка растений Клеточная стенка (нередко в качестве синонима термина "клеточная стенка" в учебной и научной литературе используется термин "клеточная оболочка". ) у растений – это структурное образование, располагающееся по периферии клетки, за пределами плазмалеммы, придающее клетке прочность, сохраняющее ее форму и защищающее протопласт. Клеточная стенка растений противостоит высокому осмотическому давлению большой центральной вакуоли и препятствует разрыву клетки. Кроме того, совокупность прочных клеточных стенок выполняет роль своеобразного внешнего скелета, поддерживающего форму растения и придающего ему механическую прочность. Клеточная стенка, обладая большой прочностью, в то же время способна к росту, и прежде всего к росту растяжением.
Надмембранный комплекс • • Надмембранный комплекс – гликокаликс содержит углеводы и белки. У эвкариот универсальным компонентом надмембранного комплекса являются углеводы – компоненты гликолипидов и гликопротеинов плазмалеммы. Благодаря этому его исходно называли гликокаликсом (от греч. гликос – сладкий, углевод и лат. каллум – толстая кожа, оболочка). Кроме углеводов, в состав гликокаликса включают периферические белки над билипидным слоем. Более сложные варианты надмембранного комплекса встречаются у растений (клеточная стенка из целлюлозы), грибов и членистоногих (наружный покров из хитина). Клеточная стенка растений Клеточная стенка (нередко в качестве синонима термина "клеточная стенка" в учебной и научной литературе используется термин "клеточная оболочка". ) у растений – это структурное образование, располагающееся по периферии клетки, за пределами плазмалеммы, придающее клетке прочность, сохраняющее ее форму и защищающее протопласт. Клеточная стенка растений противостоит высокому осмотическому давлению большой центральной вакуоли и препятствует разрыву клетки. Кроме того, совокупность прочных клеточных стенок выполняет роль своеобразного внешнего скелета, поддерживающего форму растения и придающего ему механическую прочность. Клеточная стенка, обладая большой прочностью, в то же время способна к росту, и прежде всего к росту растяжением.
 А – клетка под световым микроскопом; Б – часть клетки") Строение растительной клетки (схема) А – клетка под световым микроскопом; Б – часть клетки под электронным микроскопом; 1 – оболочка (а – первичная, б – вторичная, в – пора); 2 – цитоплазма; 3 – хлоропласт (г – оболочка, д – строма с ламеллами и гранами, е – первичное крахмальное зерно); 4 – ядро с ядрышком; 5 – вакуоль; 6 – митохондрии; 7 – каналы эндоплазматической сети с рибосомами на мембранах; 8 – плазмалемма; 9 – тонопласт; 10 – диктиосомы; 11 – цитоплазматический матрикс, или гиалоплазма
Строение растительной клетки (схема) А – клетка под световым микроскопом; Б – часть клетки под электронным микроскопом; 1 – оболочка (а – первичная, б – вторичная, в – пора); 2 – цитоплазма; 3 – хлоропласт (г – оболочка, д – строма с ламеллами и гранами, е – первичное крахмальное зерно); 4 – ядро с ядрышком; 5 – вакуоль; 6 – митохондрии; 7 – каналы эндоплазматической сети с рибосомами на мембранах; 8 – плазмалемма; 9 – тонопласт; 10 – диктиосомы; 11 – цитоплазматический матрикс, или гиалоплазма
 Электронная микрофотография клеток Nicotiana tabacum. Продольный срез молодых клеток. ЭР –эндоплазматический ретикулум; М – митохондрии; Я – ядро; ЯО – ядерная оболочка; ЯД – ядрышко; МАТ – масляное тельце; ПЛ – пластида; В – вакуоль; КС – клеточная стенка; (Эзау, 1997 из Эверт, 2015)
Электронная микрофотография клеток Nicotiana tabacum. Продольный срез молодых клеток. ЭР –эндоплазматический ретикулум; М – митохондрии; Я – ядро; ЯО – ядерная оболочка; ЯД – ядрышко; МАТ – масляное тельце; ПЛ – пластида; В – вакуоль; КС – клеточная стенка; (Эзау, 1997 из Эверт, 2015)
 Клеточная стенка • • Клеточная стенка живой функционирующей клетки растений, как правило, прозрачна и хорошо пропускает солнечный свет. Через нее легко проникают вода и низкомолекулярные вещества, но для высокомолекулярных веществ она полностью или частично непроницаема. У многоклеточных организмов стенки соседних клеток скреплены между собой пектиновыми веществами и лигнином, образующими срединную пластинку, например, клетки древесины. При специальной обработке растительных тканей некоторыми веществами (крепкие щелочи, азотная кислота) стенки соседних клеток разъединяются в результате разрушения срединной пластинки. Этот процесс называется мацерацией. Естественная мацерация происходит у перезрелых плодов груши, дыни, персика и др. Стенка клетки представляет собой продукт жизнедеятельности ее протопласта, поэтому стенка может расти, только находясь в контакте с протопластом. Однако при отмирании протопласта стенка сохраняется, и мертвая клетка может продолжать выполнять функции проведения воды или играть роль механической опоры, например в древесине стволов, ветвей и корней древесных растений. В клеточной стенке каркасом являются пучки молекул целлюлозы, а связующим веществом служат гемицеллюлоза и пектины, которые образуют матрикс клеточной стенки. Эти вещества транспортируются во время роста клеточной пластинки из комплекса Гольджи. Лигнин. Камбий.
Клеточная стенка • • Клеточная стенка живой функционирующей клетки растений, как правило, прозрачна и хорошо пропускает солнечный свет. Через нее легко проникают вода и низкомолекулярные вещества, но для высокомолекулярных веществ она полностью или частично непроницаема. У многоклеточных организмов стенки соседних клеток скреплены между собой пектиновыми веществами и лигнином, образующими срединную пластинку, например, клетки древесины. При специальной обработке растительных тканей некоторыми веществами (крепкие щелочи, азотная кислота) стенки соседних клеток разъединяются в результате разрушения срединной пластинки. Этот процесс называется мацерацией. Естественная мацерация происходит у перезрелых плодов груши, дыни, персика и др. Стенка клетки представляет собой продукт жизнедеятельности ее протопласта, поэтому стенка может расти, только находясь в контакте с протопластом. Однако при отмирании протопласта стенка сохраняется, и мертвая клетка может продолжать выполнять функции проведения воды или играть роль механической опоры, например в древесине стволов, ветвей и корней древесных растений. В клеточной стенке каркасом являются пучки молекул целлюлозы, а связующим веществом служат гемицеллюлоза и пектины, которые образуют матрикс клеточной стенки. Эти вещества транспортируются во время роста клеточной пластинки из комплекса Гольджи. Лигнин. Камбий.
 Строение клеток древесины Схема строения стенки клеток вторичной ксилемы: 1 – пектиновая прослойка между оболочками соседних клеток; 2 – так называемая серединная пластинка, объединяющая первичные оболочки прилегающих друг к другу соседних клеток; 3 – вторичная оболочка, состоящая из трёх слоёв, с разной ориентацией фибрилл целлюлозы Схемы строения пор в клетках древесины древесных растений. А – круглая окаймлённая пора хвойных. Б – щелевидные полуокаймлённая (а) и простая (б) поры лиственных
Строение клеток древесины Схема строения стенки клеток вторичной ксилемы: 1 – пектиновая прослойка между оболочками соседних клеток; 2 – так называемая серединная пластинка, объединяющая первичные оболочки прилегающих друг к другу соседних клеток; 3 – вторичная оболочка, состоящая из трёх слоёв, с разной ориентацией фибрилл целлюлозы Схемы строения пор в клетках древесины древесных растений. А – круглая окаймлённая пора хвойных. Б – щелевидные полуокаймлённая (а) и простая (б) поры лиственных
 Клеточная стенка, субмембранный комплекс • • Стенки некоторых типов клеток могут включать слои липидов: воска, кутина и суберина. Кутин и воск обычно покрывают наружные стенки клеток эпидермы. Слой кутина создает на поверхности растения водо- и воздухонепроницаемый слой кутикулы. Суберин пропитывает стенки, например, пробкового слоя коры деревьев. Он непроницаем для воды и газов, поэтому такая суберинизированная, или опробковевшая, клетка быстро отмирает. У большинства видов берёзы в клетках пробки откладывается бетулин – белое порошкообразное вещество – тритерпеновый спирт. Субмембранный комплекс или субмембранный опорно-сократительный аппарат располагается под плазмалеммой, с внутренней стороны поверхностного аппарата клетки. В составе субмембранного опорносократительного аппарата выделяют периферическую гиалоплазму и опорносократительную систему. Периферическая гиалоплазма это жидкое гетерогенное вещество. Она обеспечивает реализацию многих функций поверхностного аппарата клетки. В периферической гиалоплазме располагается второй компонент субмембранного опорно-сократительного аппарата – опорно -сократительная система, которая состоит из: микрофибрилл или микрофиламентов, скелетных фибрилл или промежуточных филаментов и микротрубочек. Микрофиблиллы – нитевидные структуры, состоят из: сократительного белка актина и миозина. В опорно-сократительной системе имеются миозиновые микрофибриллы. Особенностями их строения является наличие “головок”, способных расщеплять АТФ.
Клеточная стенка, субмембранный комплекс • • Стенки некоторых типов клеток могут включать слои липидов: воска, кутина и суберина. Кутин и воск обычно покрывают наружные стенки клеток эпидермы. Слой кутина создает на поверхности растения водо- и воздухонепроницаемый слой кутикулы. Суберин пропитывает стенки, например, пробкового слоя коры деревьев. Он непроницаем для воды и газов, поэтому такая суберинизированная, или опробковевшая, клетка быстро отмирает. У большинства видов берёзы в клетках пробки откладывается бетулин – белое порошкообразное вещество – тритерпеновый спирт. Субмембранный комплекс или субмембранный опорно-сократительный аппарат располагается под плазмалеммой, с внутренней стороны поверхностного аппарата клетки. В составе субмембранного опорносократительного аппарата выделяют периферическую гиалоплазму и опорносократительную систему. Периферическая гиалоплазма это жидкое гетерогенное вещество. Она обеспечивает реализацию многих функций поверхностного аппарата клетки. В периферической гиалоплазме располагается второй компонент субмембранного опорно-сократительного аппарата – опорно -сократительная система, которая состоит из: микрофибрилл или микрофиламентов, скелетных фибрилл или промежуточных филаментов и микротрубочек. Микрофиблиллы – нитевидные структуры, состоят из: сократительного белка актина и миозина. В опорно-сократительной системе имеются миозиновые микрофибриллы. Особенностями их строения является наличие “головок”, способных расщеплять АТФ.
 Цитоскелет, микротрубочки • • Цитоскелет. Скелетные фибриллы образуются путем полимеризации отдельных белковых молекул. Скелетные фибриллы разного типа клеток состоят из разных белков. В эпителиальных клетках скелетные фибриллы формируются белком прекератином и называются тонофибриллами. Все скелетные фибриллы устойчивы к физическим и химическим агентам. Они выполняют опорную функцию и являются элементами цитоскелета. Число и длина скелетных фибрилл регулируется клеточными механизмами, изменения которых может вызывать аномальное функционирование клеток. Микротрубочки занимают наиболее отдаленное от плазмалеммы положение. Стенки микротрубочек сформированы белками тубулинами. Микротрубочки включают и другие виды белков, которые называются МАР-белки. Эти белки обеспечивают эффективное функционирование микротрубочек. Формирование микротрубочек основано на процессе полимеризации тубулиновых димеров. Сначала образуются тубулиновые нити – протофиламенты, которые взаимодействуют между собой, образуя стенку микротрубочки. Наиболее мощным центром организации микротрубочек являются центриоли в животных клетках, в растительных клетках они не найдены. В цитоплазме выделяют: гиалоплазму, органеллы общего назначения, органеллы специального назначения и многочисленные разнородные включения.
Цитоскелет, микротрубочки • • Цитоскелет. Скелетные фибриллы образуются путем полимеризации отдельных белковых молекул. Скелетные фибриллы разного типа клеток состоят из разных белков. В эпителиальных клетках скелетные фибриллы формируются белком прекератином и называются тонофибриллами. Все скелетные фибриллы устойчивы к физическим и химическим агентам. Они выполняют опорную функцию и являются элементами цитоскелета. Число и длина скелетных фибрилл регулируется клеточными механизмами, изменения которых может вызывать аномальное функционирование клеток. Микротрубочки занимают наиболее отдаленное от плазмалеммы положение. Стенки микротрубочек сформированы белками тубулинами. Микротрубочки включают и другие виды белков, которые называются МАР-белки. Эти белки обеспечивают эффективное функционирование микротрубочек. Формирование микротрубочек основано на процессе полимеризации тубулиновых димеров. Сначала образуются тубулиновые нити – протофиламенты, которые взаимодействуют между собой, образуя стенку микротрубочки. Наиболее мощным центром организации микротрубочек являются центриоли в животных клетках, в растительных клетках они не найдены. В цитоплазме выделяют: гиалоплазму, органеллы общего назначения, органеллы специального назначения и многочисленные разнородные включения.
 • • Органеллы – жизненно важные части клетки. Они постоянно присутствуют в") Органеллы (органоиды) • • Органеллы – жизненно важные части клетки. Они постоянно присутствуют в клетке, имеют специфическое строение и выполняют свойственные каждой из них функции. Органеллы общего назначения: ядро, эндоплазматическая сеть, комплекс Гольджи, митохондрии, рибосомы, лизосомы (свойственны в основном животной клетке), клеточный центр, пластиды (хлоропласты, хромопласты, лейкопласты); . органеллы специального назначения: жгутики, реснички, миофибриллы, нейрофибриллы; включения (непостоянные компоненты клетки): запасные, секреторные, специфические. Многие органеллы ещё не выявлены. Ядерная оболочка. Внутреннее пространство клеточного ядра отделено от цито-плазмы ядерной оболочкой, состоящей из двух мембран. Мембраны это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который встроены молекулы белков. Пространство между внутренней и внешней ядерными мембранами называется перинуклеарным. На поверхности внешней ядерной мембраны обычно располагается большое число рибосом, и иногда удается наблюдать непосредственный переход этой мембраны в систему каналов гранулярной эндоплазматической сети клетки. Густая сеть фибрилл ядерной ламины способна обеспечить целостность ядра, даже после растворения липидных мембран оболочки ядра в эксперименте. С внутренней стороны к ламине крепятся петли хроматина, заполняющего ядро.
Органеллы (органоиды) • • Органеллы – жизненно важные части клетки. Они постоянно присутствуют в клетке, имеют специфическое строение и выполняют свойственные каждой из них функции. Органеллы общего назначения: ядро, эндоплазматическая сеть, комплекс Гольджи, митохондрии, рибосомы, лизосомы (свойственны в основном животной клетке), клеточный центр, пластиды (хлоропласты, хромопласты, лейкопласты); . органеллы специального назначения: жгутики, реснички, миофибриллы, нейрофибриллы; включения (непостоянные компоненты клетки): запасные, секреторные, специфические. Многие органеллы ещё не выявлены. Ядерная оболочка. Внутреннее пространство клеточного ядра отделено от цито-плазмы ядерной оболочкой, состоящей из двух мембран. Мембраны это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который встроены молекулы белков. Пространство между внутренней и внешней ядерными мембранами называется перинуклеарным. На поверхности внешней ядерной мембраны обычно располагается большое число рибосом, и иногда удается наблюдать непосредственный переход этой мембраны в систему каналов гранулярной эндоплазматической сети клетки. Густая сеть фибрилл ядерной ламины способна обеспечить целостность ядра, даже после растворения липидных мембран оболочки ядра в эксперименте. С внутренней стороны к ламине крепятся петли хроматина, заполняющего ядро.
 Интерфазное ядро в клетке корневой меристемы чернушки посевной Nigella sativa: В – вакуоли; Ядр – ядрышко; ФК – фибрилярный компонент ядрышка; Хн – хроматин; КО – оболочки двух соседних клеток. Ув. 25 000 раз. Электронная микрофотография.
Интерфазное ядро в клетке корневой меристемы чернушки посевной Nigella sativa: В – вакуоли; Ядр – ядрышко; ФК – фибрилярный компонент ядрышка; Хн – хроматин; КО – оболочки двух соседних клеток. Ув. 25 000 раз. Электронная микрофотография.
 • Я дерная лами на — фибриллярная сеть жесткой структуры, подстилает ядерную") Органеллы (продолжение) • Я дерная лами на — фибриллярная сеть жесткой структуры, подстилает ядерную мембрану (находится под ядерной мембраной), участвует в организации хроматина.
Органеллы (продолжение) • Я дерная лами на — фибриллярная сеть жесткой структуры, подстилает ядерную мембрану (находится под ядерной мембраной), участвует в организации хроматина.
 Хроматин ядра • • Клеточное ядро является вместилищем основной, генетической информации клетки, поэтому основное содержимое клеточного ядра – это хроматин: комплекс дезоксирибонуклеиновой кислоты (ДНК) и различных белков. Хроматин в ядре представлен эухроматином и гетерохроматином. Эухроматин, то есть активный хроматин, представляет собой участки хроматина, сохраняющие деспирализованное состояние в покоящемся ядре, т. е. в интерфазе. Он характеризуется способностью к интенсивному синтезу рибонуклеиновой кислоты и большим содержанием негистоновых белков. Эухроматин содержит большинство структурных генов организма. Гетерохроматин – это участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость, то есть перенос генетической информации с ДНК на РНК. Гетерохроматин разделяют на основной, факультативный и конститутивный. Основное функциональное отличие факультативного гетерохроматина от конститутивного – возможность перехода в эухроматиновое состояние, при котором ДНК становится транскрипционно активной и, соответственно, происходит экспрессия генов, локализованных на данном участке хромосомы. Гетерохроматин весьма сложен по структуре и его функциям.
Хроматин ядра • • Клеточное ядро является вместилищем основной, генетической информации клетки, поэтому основное содержимое клеточного ядра – это хроматин: комплекс дезоксирибонуклеиновой кислоты (ДНК) и различных белков. Хроматин в ядре представлен эухроматином и гетерохроматином. Эухроматин, то есть активный хроматин, представляет собой участки хроматина, сохраняющие деспирализованное состояние в покоящемся ядре, т. е. в интерфазе. Он характеризуется способностью к интенсивному синтезу рибонуклеиновой кислоты и большим содержанием негистоновых белков. Эухроматин содержит большинство структурных генов организма. Гетерохроматин – это участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость, то есть перенос генетической информации с ДНК на РНК. Гетерохроматин разделяют на основной, факультативный и конститутивный. Основное функциональное отличие факультативного гетерохроматина от конститутивного – возможность перехода в эухроматиновое состояние, при котором ДНК становится транскрипционно активной и, соответственно, происходит экспрессия генов, локализованных на данном участке хромосомы. Гетерохроматин весьма сложен по структуре и его функциям.
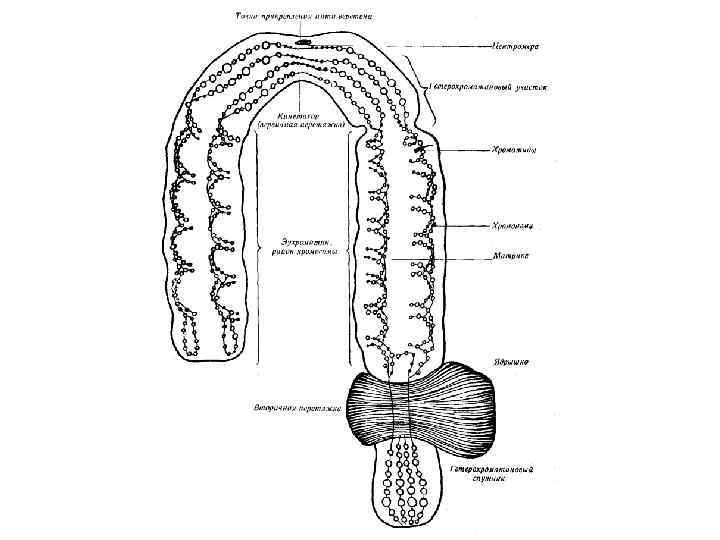
 • • • Гетерохроматин занимает значительные части геномов, например, у человека") Хроматин ядра (гетерохроматин) • • • Гетерохроматин занимает значительные части геномов, например, у человека и дрозофилы он составляет около 30%. Сначала считалось, что гетерохроматин является генетически инертным. В дальнейшем эта часть генома представлялась как «мусорная» из-за обилия некодирующих повторяющихся последовательностей ДНК и не большого количества структурных генов. Однако в настоящее время значимость гетерохроматина в процессах онто- и филогенеза очевидна: 1). Гетерохроматин присутствует в геномах подавляющего большинства видов, что косвенно может свидетельствовать о его необходимости. Для него характерна поздняя репликация и локальная недорепликация. Как правило, гетерохроматиновые участки реплицируются в последнюю очередь в конце S периода. 2). Последовательности ДНК, составляющие гетерохроматин, в ходе онтогенеза могут легко теряться из ядер соматических клеток, например: при политенизации хромосом у дрозофилы, диминуции хроматина аскарид или циклопов, при созревании макронуклеуса у инфузорий и т. д. Однако в клетках зародышевого пути эти последовательности ДНК всегда присутствуют.
Хроматин ядра (гетерохроматин) • • • Гетерохроматин занимает значительные части геномов, например, у человека и дрозофилы он составляет около 30%. Сначала считалось, что гетерохроматин является генетически инертным. В дальнейшем эта часть генома представлялась как «мусорная» из-за обилия некодирующих повторяющихся последовательностей ДНК и не большого количества структурных генов. Однако в настоящее время значимость гетерохроматина в процессах онто- и филогенеза очевидна: 1). Гетерохроматин присутствует в геномах подавляющего большинства видов, что косвенно может свидетельствовать о его необходимости. Для него характерна поздняя репликация и локальная недорепликация. Как правило, гетерохроматиновые участки реплицируются в последнюю очередь в конце S периода. 2). Последовательности ДНК, составляющие гетерохроматин, в ходе онтогенеза могут легко теряться из ядер соматических клеток, например: при политенизации хромосом у дрозофилы, диминуции хроматина аскарид или циклопов, при созревании макронуклеуса у инфузорий и т. д. Однако в клетках зародышевого пути эти последовательности ДНК всегда присутствуют.
. Те немногие гены, которые найдены в гетерохроматине, чаще") Свойства гетерохроматина • • • 3). Те немногие гены, которые найдены в гетерохроматине, чаще всего связаны с осуществлением каких-то функций в клетках зародышевого пути, например для созрева-ния гамет. Некоторые последовательности ДНК, расположенные в гетерохроматине, необходимы для осуществления функций в ходе мейоза, например для спаривания поло-вых хромосом в профазе. 4). Прицентромерный гетерохроматин необходим для правильного функциониро-вания центромер, для правильной сегрегации гомологичных хромосом в мейозе и поддер-жания связи сестринских хроматид в митозе. 5). Гетерохроматин влияет на экспрессию генов: инактивирует перенесенные к нему хромосомными перестройками соседние гены, обуславливая мозаичный эффект по-ложения. Эффект положения фенотипически проявляется как разнокачественность клеток одинаковых по генотипу. Приближение эухроматиновых районов к гетерохроматиновым приводит к переходу приближенных участков в гетерохроматизированное состояние. 6). Гетерохроматин способствует различным хромосомным перестройкам. Точки разрывов инверсий и робертсоновских слияний хромосом часто картируются в районах прицентромерного и интеркалярного гетерохроматина. 7). Гетерохроматин содержит некоторые жизненно важные гены, например гены рибосомальных РНК и гены фертильности, в частности Y-связанные гены фертильности у дрозофилы.
Свойства гетерохроматина • • • 3). Те немногие гены, которые найдены в гетерохроматине, чаще всего связаны с осуществлением каких-то функций в клетках зародышевого пути, например для созрева-ния гамет. Некоторые последовательности ДНК, расположенные в гетерохроматине, необходимы для осуществления функций в ходе мейоза, например для спаривания поло-вых хромосом в профазе. 4). Прицентромерный гетерохроматин необходим для правильного функциониро-вания центромер, для правильной сегрегации гомологичных хромосом в мейозе и поддер-жания связи сестринских хроматид в митозе. 5). Гетерохроматин влияет на экспрессию генов: инактивирует перенесенные к нему хромосомными перестройками соседние гены, обуславливая мозаичный эффект по-ложения. Эффект положения фенотипически проявляется как разнокачественность клеток одинаковых по генотипу. Приближение эухроматиновых районов к гетерохроматиновым приводит к переходу приближенных участков в гетерохроматизированное состояние. 6). Гетерохроматин способствует различным хромосомным перестройкам. Точки разрывов инверсий и робертсоновских слияний хромосом часто картируются в районах прицентромерного и интеркалярного гетерохроматина. 7). Гетерохроматин содержит некоторые жизненно важные гены, например гены рибосомальных РНК и гены фертильности, в частности Y-связанные гены фертильности у дрозофилы.
. Хромосомы • • • 8). Связи гетерохроматина с ядерной оболочкой являются основой") Гетерохроматин (продолжение). Хромосомы • • • 8). Связи гетерохроматина с ядерной оболочкой являются основой для пространственного упорядочения отдельных хромосом и интерфазного ядра в целом. 9). Изменения гетерохроматина в кариотипах связаны с видообразованием. Изменения касаются структуры, состава, количества и распределения гетерохроматина на хромосомах. Приведённые выше сведения о гетерохроматине ядерной ДНК не являются исчерпыващими. Изучение роли этих структур продолжаются. • Хромосомы • Клетки эвкариот содержат обычно несколько хромосом (от двух до нескольких сотен), которые в интерфазе (между митотическими делениями) теряют свою компактную форму, разрыхляются и заполняют объем ядра в виде хроматина. Несмотря на деконденсированное состояние хроматина в интерфазе, каждая хромосома занимает в ядре строго определенное положение и связана с ядерной оболочкой посредством ламины. Они строго закреплены на внутренней поверхности оболочки ядра такими структурами хромосом, как центромеры и теломеры. Число хромосом практически никак не связано со сложностью организации того или иного организма и его систематическим положением.
Гетерохроматин (продолжение). Хромосомы • • • 8). Связи гетерохроматина с ядерной оболочкой являются основой для пространственного упорядочения отдельных хромосом и интерфазного ядра в целом. 9). Изменения гетерохроматина в кариотипах связаны с видообразованием. Изменения касаются структуры, состава, количества и распределения гетерохроматина на хромосомах. Приведённые выше сведения о гетерохроматине ядерной ДНК не являются исчерпыващими. Изучение роли этих структур продолжаются. • Хромосомы • Клетки эвкариот содержат обычно несколько хромосом (от двух до нескольких сотен), которые в интерфазе (между митотическими делениями) теряют свою компактную форму, разрыхляются и заполняют объем ядра в виде хроматина. Несмотря на деконденсированное состояние хроматина в интерфазе, каждая хромосома занимает в ядре строго определенное положение и связана с ядерной оболочкой посредством ламины. Они строго закреплены на внутренней поверхности оболочки ядра такими структурами хромосом, как центромеры и теломеры. Число хромосом практически никак не связано со сложностью организации того или иного организма и его систематическим положением.
 Хромосомы Схематическое изображение метафазных хромосом. а – хромосома со спутником и двумя перетяжками: 1 – первичная перетяжка, 2 – меньшее плечо, 3 – большее плечо, 4 – вторичная перетяжка, 5 – спутник, 6 – центромера. б – то же, что и а, но хромосома без белков: 1 – центромера, 2 – спирализованная ДНК; В – разные типы хромосом: 1 – акроцентрическая, 2 – субметацентрическая, 3 – метацентрическая, А – плечо, Б – центромера
Хромосомы Схематическое изображение метафазных хромосом. а – хромосома со спутником и двумя перетяжками: 1 – первичная перетяжка, 2 – меньшее плечо, 3 – большее плечо, 4 – вторичная перетяжка, 5 – спутник, 6 – центромера. б – то же, что и а, но хромосома без белков: 1 – центромера, 2 – спирализованная ДНК; В – разные типы хромосом: 1 – акроцентрическая, 2 – субметацентрическая, 3 – метацентрическая, А – плечо, Б – центромера
 • • Изменение числа хромосом вследствие возникновения полиплоидии или иных хромосомных мутаций") Хромосомы (продолжение) • • Изменение числа хромосом вследствие возникновения полиплоидии или иных хромосомных мутаций приводит к весьма неоднозначным последствиям. В природе и экспериментах наблюдаются следующие виды хромосомных перестроек: делеции (утрата участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы с одной на другую), а также дицентрические и кольцевые хромосомы. Известны также изохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной (инверсии, делеции, дупликации, кольцевые хромосомы), если же двух разных, то межхромосомной (дупликации, транслокации, дицентрические хромосомы). Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки (инверсии, реципрокные транслокации) не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки (делеции и дупликации) меняют дозовое соотношение генов, и, как правило, их возникновение сопряжено с существенными отклонениями от нормы.
Хромосомы (продолжение) • • Изменение числа хромосом вследствие возникновения полиплоидии или иных хромосомных мутаций приводит к весьма неоднозначным последствиям. В природе и экспериментах наблюдаются следующие виды хромосомных перестроек: делеции (утрата участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы с одной на другую), а также дицентрические и кольцевые хромосомы. Известны также изохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной (инверсии, делеции, дупликации, кольцевые хромосомы), если же двух разных, то межхромосомной (дупликации, транслокации, дицентрические хромосомы). Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки (инверсии, реципрокные транслокации) не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки (делеции и дупликации) меняют дозовое соотношение генов, и, как правило, их возникновение сопряжено с существенными отклонениями от нормы.
 Хромосомы. Старение • • Генетическая нестабильность и укорочение теломер Согласно распространенной гипотезе, сигналом для начала клеточного старения может служить критическое укорочение теломерных концов хромосом вследствие последовательной потери повторяющихся ДНКпоследовательностей с каждым новым циклом деления клетки. Существует предположение, что укорочение теломер, приводя в итоге к формированию дицентрических хромосом, может запускать реакцию на повреждение ДНК. В результате клетка перестает делиться и стареет Введение в геном фибробластных клеток активного фермента теломеразы, достраивающего теломеры , позволило продлить жизнь клеточным популяциям in vitro. Ядрышко Кроме хроматина, составляющего хромосомы, в ядрах эвкариот обычно содержится одно или несколько ядрышек. Это плотные структуры, не имеющие собственной оболочки и представляющие собой скопления молекул рибосомной РНК (р-РНК) в комплексе с белками. Такие комплексы называют рибонуклеопротеидами (РНП). Гены рибосомной РНК, в отличие от большинства других генов, кодирующих белки, содержатся в геноме в виде многочисленных копий. Эти копии, расположенные в молекуле ДНК тандемно, т. е. друг за другом, располагаются в определенных районах нескольких хромосом генома. Такие районы хромосом называют ядрышковыми организаторами.
Хромосомы. Старение • • Генетическая нестабильность и укорочение теломер Согласно распространенной гипотезе, сигналом для начала клеточного старения может служить критическое укорочение теломерных концов хромосом вследствие последовательной потери повторяющихся ДНКпоследовательностей с каждым новым циклом деления клетки. Существует предположение, что укорочение теломер, приводя в итоге к формированию дицентрических хромосом, может запускать реакцию на повреждение ДНК. В результате клетка перестает делиться и стареет Введение в геном фибробластных клеток активного фермента теломеразы, достраивающего теломеры , позволило продлить жизнь клеточным популяциям in vitro. Ядрышко Кроме хроматина, составляющего хромосомы, в ядрах эвкариот обычно содержится одно или несколько ядрышек. Это плотные структуры, не имеющие собственной оболочки и представляющие собой скопления молекул рибосомной РНК (р-РНК) в комплексе с белками. Такие комплексы называют рибонуклеопротеидами (РНП). Гены рибосомной РНК, в отличие от большинства других генов, кодирующих белки, содержатся в геноме в виде многочисленных копий. Эти копии, расположенные в молекуле ДНК тандемно, т. е. друг за другом, располагаются в определенных районах нескольких хромосом генома. Такие районы хромосом называют ядрышковыми организаторами.
 Хромосомы, ядрышковые организаторы • Морфологически в ядрышке с помощью электронного микроскопа можно выделить следующие 3 зоны: гомогенные компактные фибриллярные центры, содержащие ДНК ядрышковых организаторов; плотный фибриллярный компонент вокруг них, где идет транскрипция генов рибосомной РНК и массивный гранулярный компонент ядрышка, состоящий из частиц РНП – будущих рибосом. Эти гранулы РНП, образующиеся в ядрышке, транспортируются в цитоплазму и образуют рибосомы, осуществляющие синтез всех белков клетки. Третий основной тип клеточных РНК – мелкие транспортные РНК – транскрибируются в различных участках ядра и выходят в цитоплазму через ядерные поры. Там они обеспечивают транспортировку аминокислот к рибосомам в процессе синтеза белков.
Хромосомы, ядрышковые организаторы • Морфологически в ядрышке с помощью электронного микроскопа можно выделить следующие 3 зоны: гомогенные компактные фибриллярные центры, содержащие ДНК ядрышковых организаторов; плотный фибриллярный компонент вокруг них, где идет транскрипция генов рибосомной РНК и массивный гранулярный компонент ядрышка, состоящий из частиц РНП – будущих рибосом. Эти гранулы РНП, образующиеся в ядрышке, транспортируются в цитоплазму и образуют рибосомы, осуществляющие синтез всех белков клетки. Третий основной тип клеточных РНК – мелкие транспортные РНК – транскрибируются в различных участках ядра и выходят в цитоплазму через ядерные поры. Там они обеспечивают транспортировку аминокислот к рибосомам в процессе синтеза белков.