

Балашов Е.В. ГНГ-11-1.03.ppt
- Количество слайдов: 81
 КЛАСС REPTILIA. ПРЕСМЫКАЮЩИЕСЯ
КЛАСС REPTILIA. ПРЕСМЫКАЮЩИЕСЯ
 — класс (по традиционной классификации), или парафилетическая") Пресмыка ющиеся (рептилии — от лат. Reptilia) — класс (по традиционной классификации), или парафилетическая группа (по кладистической классификации) преимущественно наземных позвоночных животных, включающий современных черепах, крокодилов, клювоголовых, амфисбен, ящериц и змей. В XVIII—XIX веках вместе с амфибиями объединялись в группу гады — холоднокровные наземные позвоночные. В мире известно около 9400 видов пресмыкающихся, на территории России обитает 72 вида. Крупнейшие наземные животные принадлежали к динозаврам — представителям древних пресмыкающихся. Пресмыкающиеся пережили расцвет в эпоху мезозоя, когда они доминировали на земле, в море и в воздухе. В конце мелового периода большая часть пресмыкающихся вымерла. Современные рептилии — лишь разрозненные остатки того мира.
Пресмыка ющиеся (рептилии — от лат. Reptilia) — класс (по традиционной классификации), или парафилетическая группа (по кладистической классификации) преимущественно наземных позвоночных животных, включающий современных черепах, крокодилов, клювоголовых, амфисбен, ящериц и змей. В XVIII—XIX веках вместе с амфибиями объединялись в группу гады — холоднокровные наземные позвоночные. В мире известно около 9400 видов пресмыкающихся, на территории России обитает 72 вида. Крупнейшие наземные животные принадлежали к динозаврам — представителям древних пресмыкающихся. Пресмыкающиеся пережили расцвет в эпоху мезозоя, когда они доминировали на земле, в море и в воздухе. В конце мелового периода большая часть пресмыкающихся вымерла. Современные рептилии — лишь разрозненные остатки того мира.
 Современные представители класса Пресмыкающиеся
Современные представители класса Пресмыкающиеся
 Разнообразие современных рептилий Когда-то было 16 отрядов рептилий, известных по ископаемым остаткам, только четыре дожили до настоящего времени. В настоящее время класс Пресмыкающихся немногочислен, включает около 6000 видов и подразделяется на 4 отряда: Чешуйчатые (ящерицы, змеи) Черепахи Крокодилы Клювоголовые (один вид - гаттерия, которая обитает в Новой Зеландии)
Разнообразие современных рептилий Когда-то было 16 отрядов рептилий, известных по ископаемым остаткам, только четыре дожили до настоящего времени. В настоящее время класс Пресмыкающихся немногочислен, включает около 6000 видов и подразделяется на 4 отряда: Чешуйчатые (ящерицы, змеи) Черепахи Крокодилы Клювоголовые (один вид - гаттерия, которая обитает в Новой Зеландии)




 Ящерицы являются хищниками. Имеют 4 конечности с коготками зубы роговой покров сухую кожу подвижные веки им свойственно явление регенерации Бывают безногие (веретеница) Бывают двуногие (айолот) Некоторые меняют окраску в зависимости от времени суток, под влиянием беспокойства или испуга (хамелеоны)
Ящерицы являются хищниками. Имеют 4 конечности с коготками зубы роговой покров сухую кожу подвижные веки им свойственно явление регенерации Бывают безногие (веретеница) Бывают двуногие (айолот) Некоторые меняют окраску в зависимости от времени суток, под влиянием беспокойства или испуга (хамелеоны)


 Змеи Отсутствуют конечности Передвигаются за счет изгибания туловища Не имеют век Нет внутреннего уха и звуки воспринимаются брюшной поверхностью Имеют термолокатор Свойственна линька
Змеи Отсутствуют конечности Передвигаются за счет изгибания туловища Не имеют век Нет внутреннего уха и звуки воспринимаются брюшной поверхностью Имеют термолокатор Свойственна линька

 Удавы (питон, анаконда) Ужеподобные (полоз, уж)") По способу питания змеи делятся: Ядовитые (гадюка, кобра) Удавы (питон, анаконда) Ужеподобные (полоз, уж)
По способу питания змеи делятся: Ядовитые (гадюка, кобра) Удавы (питон, анаконда) Ужеподобные (полоз, уж)
 Неуязвимые черепахи Как ни тяжеловесен панцирь, но именно он помог черепахам успешно просуществовать долгих 175 мл. лет. Форма панциря зависит от способа существования вида. У морских панцирь тоньше, поэтому они самые быстроходные рептилии.
Неуязвимые черепахи Как ни тяжеловесен панцирь, но именно он помог черепахам успешно просуществовать долгих 175 мл. лет. Форма панциря зависит от способа существования вида. У морских панцирь тоньше, поэтому они самые быстроходные рептилии.
 У наземных черепах панцирь легкий и тонкий, он хорошо защищает от врагов и потери влаги. Конечности приспособлены для хождения.
У наземных черепах панцирь легкий и тонкий, он хорошо защищает от врагов и потери влаги. Конечности приспособлены для хождения.
 Гигантские крокодилы Крокодилы – вторичноводные животные предки, которых обитали на суше. Это самые крупные и высокоорганизован ные рептилии.
Гигантские крокодилы Крокодилы – вторичноводные животные предки, которых обитали на суше. Это самые крупные и высокоорганизован ные рептилии.
 Среди крокодилов встречаются: Нильский крокодил – длина более 5 -ти метров. Индийский гавиал – имеет длину до 6, 5 метров и самую узкую и длинную среди всех крокодилов морду. Южно-американский кайман - длина 2 -4 метра. Способен издавать разнообразные звуки – от рычания и карканья до храпа и фырканья.
Среди крокодилов встречаются: Нильский крокодил – длина более 5 -ти метров. Индийский гавиал – имеет длину до 6, 5 метров и самую узкую и длинную среди всех крокодилов морду. Южно-американский кайман - длина 2 -4 метра. Способен издавать разнообразные звуки – от рычания и карканья до храпа и фырканья.
 Аллигатор или крокодил ? У аллигатора более широкая и округлая морда, а зубы верхней челюсти при закрытой пасти располагаются снаружи. У крокодила морда более заостренная, а зубы верхней и нижней челюстей находятся на одной линии.
Аллигатор или крокодил ? У аллигатора более широкая и округлая морда, а зубы верхней челюсти при закрытой пасти располагаются снаружи. У крокодила морда более заостренная, а зубы верхней и нижней челюстей находятся на одной линии.
- единственный вид, оставшийся от отряда клювоголовых. Обитают они в") Избранница судьбы Гаттерия (или туатара)- единственный вид, оставшийся от отряда клювоголовых. Обитают они в Новой Зеландии. Эти животные способны жить в прохладном климате. Обладают настоящим голосом, их печальное пение можно услышать туманными ночами.
Избранница судьбы Гаттерия (или туатара)- единственный вид, оставшийся от отряда клювоголовых. Обитают они в Новой Зеландии. Эти животные способны жить в прохладном климате. Обладают настоящим голосом, их печальное пение можно услышать туманными ночами.
 ДРЕВНИЕ ПРЕДСТАВИТЕЛИ ПРЕСМЫКАЮЩИХСЯ
ДРЕВНИЕ ПРЕДСТАВИТЕЛИ ПРЕСМЫКАЮЩИХСЯ
 Крупнейшие наземные животные принадлежали к динозаврам — представителям древних пресмыкающихся. Пресмыкающиеся пережили расцвет в эпоху мезозоя, когда они доминировали на земле, в море и в воздухе. В конце мелового периода большая часть пресмыкающихся вымерла.
Крупнейшие наземные животные принадлежали к динозаврам — представителям древних пресмыкающихся. Пресмыкающиеся пережили расцвет в эпоху мезозоя, когда они доминировали на земле, в море и в воздухе. В конце мелового периода большая часть пресмыкающихся вымерла.
 Ископаемые рептилии. Рептилии известны с каменноугольного периода. В течение пермского и триасового периодов они достигли крупных размеров и размножились в огромных количествах, приспособившись к различным местообитаниям. В мезозое они господствовали среди животных в воздухе, на суше и на море, так что вся эта эра получила название века рептилий. Плезиозавры и ихтиозавры отлично плавали, птерозавры летали, а самые крупные, динозавры, были сухопутными формами.
Ископаемые рептилии. Рептилии известны с каменноугольного периода. В течение пермского и триасового периодов они достигли крупных размеров и размножились в огромных количествах, приспособившись к различным местообитаниям. В мезозое они господствовали среди животных в воздухе, на суше и на море, так что вся эта эра получила название века рептилий. Плезиозавры и ихтиозавры отлично плавали, птерозавры летали, а самые крупные, динозавры, были сухопутными формами.
 В России динозавры найдены в Якутии.
В России динозавры найдены в Якутии.
 Местонахождения динозавров в Якутии находится в глухой тайге.
Местонахождения динозавров в Якутии находится в глухой тайге.
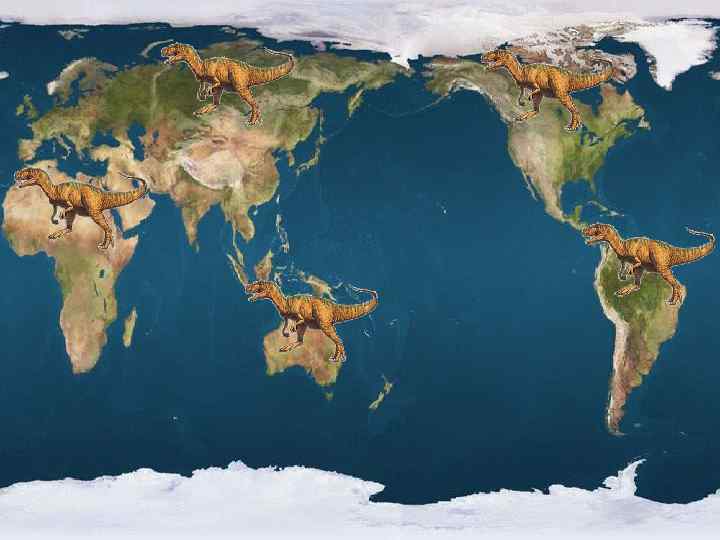
 Отдельные кости и целые скелеты динозавров обнаружены учёными на всех континентах, кроме Антарктиды.
Отдельные кости и целые скелеты динозавров обнаружены учёными на всех континентах, кроме Антарктиды.
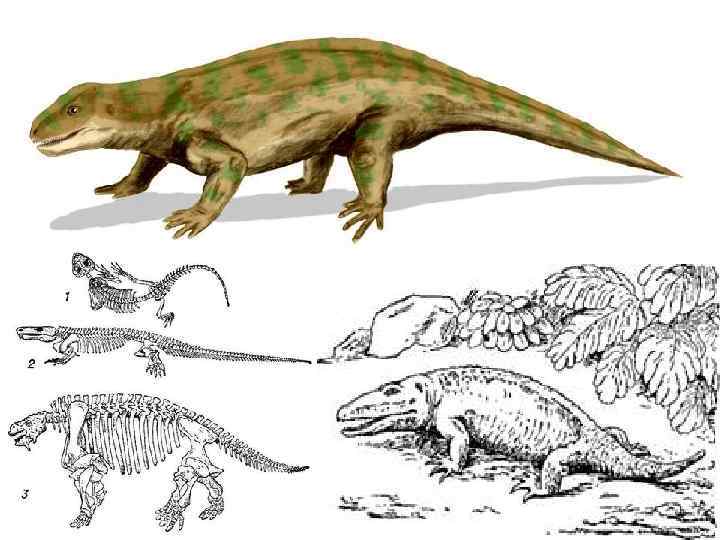
 C 3 -T 1 Котилозавры (Cotylosauria), подкласс вымерших наиболее") Подкласс Cotylosauria ( «котeлкоголовые» ) C 3 -T 1 Котилозавры (Cotylosauria), подкласс вымерших наиболее древних и примитивных пресмыкающихся; известны с середины каменноугольного по триасовый период. Размеры варьируют от мелких насекомоядных проколофонов и средней величины хищников (в т. ч. водных) до крупных растительноядных парейазавров. Большинство котилозавров сохранило многие примитивные признаки, унаследованные от земноводных: череп без височных впадин с хорошо развитым теменным отверстием; небные зубы; двояковогнутые позвонки; очень короткий шейный отдел; короткие и массивные кости конечностей и их поясов и др. Остатки котилозавров многочисленны в Северной Америке и Южной Африке, а также в Европейской части СССР; используются в качестве руководящих форм в геологии.
Подкласс Cotylosauria ( «котeлкоголовые» ) C 3 -T 1 Котилозавры (Cotylosauria), подкласс вымерших наиболее древних и примитивных пресмыкающихся; известны с середины каменноугольного по триасовый период. Размеры варьируют от мелких насекомоядных проколофонов и средней величины хищников (в т. ч. водных) до крупных растительноядных парейазавров. Большинство котилозавров сохранило многие примитивные признаки, унаследованные от земноводных: череп без височных впадин с хорошо развитым теменным отверстием; небные зубы; двояковогнутые позвонки; очень короткий шейный отдел; короткие и массивные кости конечностей и их поясов и др. Остатки котилозавров многочисленны в Северной Америке и Южной Африке, а также в Европейской части СССР; используются в качестве руководящих форм в геологии.
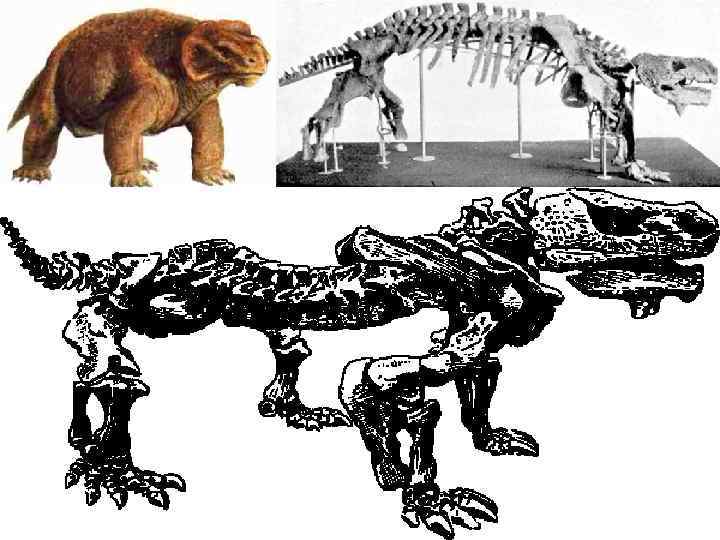
 P 2 Парейазавры (Pareiasauria), подотряд вымерших пресмыкающихся из подкласса") Род Pareiasaurus ( «щекастый» ) P 2 Парейазавры (Pareiasauria), подотряд вымерших пресмыкающихся из подкласса котилозавров; жили в позднепермскую эпоху. Для Парейазавров характерны: короткий и широкий череп со скульптурой в виде крупных ячей; боковые выросты на скуловых костях; зубы с зазубренной по краям коронкой, приспособленные к перетиранию растительной пищи; массивные конечности. Некоторые имели спинной панцирь из костных пластин. Размеры от средних (около 1 м) до крупных (до 3 м). Типичный представитель — скутозавр, несколько скелетов которого были найдены в начале 20 в. на Северной Двине Парейазавры были распространены к Западу от Урала и в Южной Африке.
Род Pareiasaurus ( «щекастый» ) P 2 Парейазавры (Pareiasauria), подотряд вымерших пресмыкающихся из подкласса котилозавров; жили в позднепермскую эпоху. Для Парейазавров характерны: короткий и широкий череп со скульптурой в виде крупных ячей; боковые выросты на скуловых костях; зубы с зазубренной по краям коронкой, приспособленные к перетиранию растительной пищи; массивные конечности. Некоторые имели спинной панцирь из костных пластин. Размеры от средних (около 1 м) до крупных (до 3 м). Типичный представитель — скутозавр, несколько скелетов которого были найдены в начале 20 в. на Северной Двине Парейазавры были распространены к Западу от Урала и в Южной Африке.
, подкласс вымерших морских пресмыкающихся. Известны") Подкласс Synaptosauria. Синаптозавры T 3 -K (Synaptosauria, или Euryapsida), подкласс вымерших морских пресмыкающихся. Известны с перми до позднего мела всех материков (кроме Антарктиды). В СССР многочисленные находки Синаптозавров в Поволжье. Длинной от 20— 30 см до 15 м. Экологически и морфологически Синаптозавры разнообразны. У большинства череп удлинённый, глазницы расположены почти посредине, теменное отверстие обычно сохраняется; конечности ластовидные, характерно одно височное окно. Ихтио- и малакофаги. 2 отр. — завроптеригии и плакодонты.
Подкласс Synaptosauria. Синаптозавры T 3 -K (Synaptosauria, или Euryapsida), подкласс вымерших морских пресмыкающихся. Известны с перми до позднего мела всех материков (кроме Антарктиды). В СССР многочисленные находки Синаптозавров в Поволжье. Длинной от 20— 30 см до 15 м. Экологически и морфологически Синаптозавры разнообразны. У большинства череп удлинённый, глазницы расположены почти посредине, теменное отверстие обычно сохраняется; конечности ластовидные, характерно одно височное окно. Ихтио- и малакофаги. 2 отр. — завроптеригии и плакодонты.

, подкласс вымерших рептилий.") Подкласс Jchthyopterygia. Ихтиоптеригии T-K 1 (ихтиоптеригии, иначе – рыбоящеры, или рыбоплавниковые), подкласс вымерших рептилий. Существовали в морях с раннего триаса до раннего мела. Наибольшего разнообразия достигли в ранней юре. Происхождение неясно. Судя по строению черепа, предками ихтиозавров могли быть ранние котилозавры. Внешне конвергентно сходны с дельфинами и быстроплавающими океаническими рыбами. Имели веретеновидное тело дл. от 50 см до 15 м, вертикальный двухлопастный хвостовой плавник и большинство – удлинённую заострённую морду. На челюстях в один ряд многочисленные конические или реже уплощённые (у моллюскоядов) зубы. В глазах склеротические кольца из мелких косточек. Имелось отверстие третьего (теменного) глаза. Характерны крупные парные передние и задние ласты (с большим числом пальцев и фаланг), непарный спинной плавник. Хвостовой отдел позвоночник
Подкласс Jchthyopterygia. Ихтиоптеригии T-K 1 (ихтиоптеригии, иначе – рыбоящеры, или рыбоплавниковые), подкласс вымерших рептилий. Существовали в морях с раннего триаса до раннего мела. Наибольшего разнообразия достигли в ранней юре. Происхождение неясно. Судя по строению черепа, предками ихтиозавров могли быть ранние котилозавры. Внешне конвергентно сходны с дельфинами и быстроплавающими океаническими рыбами. Имели веретеновидное тело дл. от 50 см до 15 м, вертикальный двухлопастный хвостовой плавник и большинство – удлинённую заострённую морду. На челюстях в один ряд многочисленные конические или реже уплощённые (у моллюскоядов) зубы. В глазах склеротические кольца из мелких косточек. Имелось отверстие третьего (теменного) глаза. Характерны крупные парные передние и задние ласты (с большим числом пальцев и фаланг), непарный спинной плавник. Хвостовой отдел позвоночник

 Род Ichthyosaurus J 1 Ихтиоза вры (лат. Ichthyosauria, «рыбоящеры» от греч. — рыба и — ящер) — вымерший отряд крупных (до 24 метров в длину, в среднем 2— 4 метра) морских рептилий, имевших форму, конвергентную рыбам и дельфинам. Предки ихтиозавров, вероятно, спустились в море по руслу одной из доисторических рек, где останки практически не сохраняются, — все известные науке виды уже жили в морской воде и не выбирались на сушу. Были живородящими. Имели эвриапсидное строение черепа, произошли от диапсидных предков. Зубы сменялись неоднократно в течение жизни. Конечности использовались для поддержания равновесия и контроля направления. У хвоста было две лопасти, нижняя из которых поддерживалась позвоночным столбом. У типичного ихтиозавра были очень большие глаза, защищённые костяным кольцом, говорящим о том, что охотились они ночью. В связи с этим, некоторые виды обладали огромными глазами (до 20 см в диаметре). Дополнительно, по-видимому, имелись какие-то кожные рецепторы, типа боковой линии, на что указывают следы нервов и сосудов на костях черепа. Кожа, лишённая чешуи, для лучшего скольжения в воде была, возможно, покрыта слизью. Созданные для скорости, как современный тунец, некоторые ихтиозавры также наводят на предположения, что они были хорошими глубоководными ныряльщиками, как современные киты. Наиболее вероятная окраска — противотень (тёмный верх и светлый низ) с синеватым оттенком. Самый крупный описанный вид по обнаруженным окаменелостям ихтиозавр — шонизавр из позднего триаса полярной Канады (в 2003 году описан скелет длиной около 23 метров). У некоторых ранних ихтиозавров были зубы, с помощью которых они питались моллюсками — аммонитами, наутилоидеми и кальмарами. Очень похоже на то, что они также питались рыбой, а у части более крупных видов были тяжёлые челюсти и зубы, которые показывают, что они питались более мелкими рептилиями. Типовой вид этой группы был описан в 1821 году — Ichthyosaurus communis de la Beche & Conybeare, 1821 typus в составе нового семейства Ichthyosauridae De la Beche & Conybeare, 1821. Ихтиозавры существовали в течение почти всего мезозоя 250— 90 млн лет назад, наибольшего расцвета достигли в юрский период, пока в меловой период их не сменили плезиозавры. В мелу количество видов ихтиозавров резко снизилось, лишь один род — Платиптеригий (Platypterygius) — дожил до начала позднего мела (единственный описанный представитель меловой группы).
Род Ichthyosaurus J 1 Ихтиоза вры (лат. Ichthyosauria, «рыбоящеры» от греч. — рыба и — ящер) — вымерший отряд крупных (до 24 метров в длину, в среднем 2— 4 метра) морских рептилий, имевших форму, конвергентную рыбам и дельфинам. Предки ихтиозавров, вероятно, спустились в море по руслу одной из доисторических рек, где останки практически не сохраняются, — все известные науке виды уже жили в морской воде и не выбирались на сушу. Были живородящими. Имели эвриапсидное строение черепа, произошли от диапсидных предков. Зубы сменялись неоднократно в течение жизни. Конечности использовались для поддержания равновесия и контроля направления. У хвоста было две лопасти, нижняя из которых поддерживалась позвоночным столбом. У типичного ихтиозавра были очень большие глаза, защищённые костяным кольцом, говорящим о том, что охотились они ночью. В связи с этим, некоторые виды обладали огромными глазами (до 20 см в диаметре). Дополнительно, по-видимому, имелись какие-то кожные рецепторы, типа боковой линии, на что указывают следы нервов и сосудов на костях черепа. Кожа, лишённая чешуи, для лучшего скольжения в воде была, возможно, покрыта слизью. Созданные для скорости, как современный тунец, некоторые ихтиозавры также наводят на предположения, что они были хорошими глубоководными ныряльщиками, как современные киты. Наиболее вероятная окраска — противотень (тёмный верх и светлый низ) с синеватым оттенком. Самый крупный описанный вид по обнаруженным окаменелостям ихтиозавр — шонизавр из позднего триаса полярной Канады (в 2003 году описан скелет длиной около 23 метров). У некоторых ранних ихтиозавров были зубы, с помощью которых они питались моллюсками — аммонитами, наутилоидеми и кальмарами. Очень похоже на то, что они также питались рыбой, а у части более крупных видов были тяжёлые челюсти и зубы, которые показывают, что они питались более мелкими рептилиями. Типовой вид этой группы был описан в 1821 году — Ichthyosaurus communis de la Beche & Conybeare, 1821 typus в составе нового семейства Ichthyosauridae De la Beche & Conybeare, 1821. Ихтиозавры существовали в течение почти всего мезозоя 250— 90 млн лет назад, наибольшего расцвета достигли в юрский период, пока в меловой период их не сменили плезиозавры. В мелу количество видов ихтиозавров резко снизилось, лишь один род — Платиптеригий (Platypterygius) — дожил до начала позднего мела (единственный описанный представитель меловой группы).

 Подкласс Archosauria. Архозавры T 1 - ныне Архоза вры (лат. Archosaurs, др. -греч. «правящие ящеры» ) — группа диапсидных пресмыкающихся, представленная в настоящее время крокодилами, а из вымерших организмов также — динозаврами, птерозаврами и другими. В кладистике птиц включают в число архозавров. Проблемы терминологии Жак Готье определил архозавров как кроновую группу: все потомки позднейшего общего предка птиц и крокодилов. Оказалось, что под это определение не подпадают многие формы, традиционно относимые к архозаврам (такие, как Archosaurus rossicus и (или) Protorosaurus speneri). Для объединения их с архозаврами в узком смысле Готье ввёл имя Archosauriformes. Другие исследователи предпочитают использование имени Archosauria в широком смысле, а для обозначения кроновой группы (архозавров в узком смысле) использует имя Avesuchia. Оба варианта достаточно распространены, что создаёт путаницу.
Подкласс Archosauria. Архозавры T 1 - ныне Архоза вры (лат. Archosaurs, др. -греч. «правящие ящеры» ) — группа диапсидных пресмыкающихся, представленная в настоящее время крокодилами, а из вымерших организмов также — динозаврами, птерозаврами и другими. В кладистике птиц включают в число архозавров. Проблемы терминологии Жак Готье определил архозавров как кроновую группу: все потомки позднейшего общего предка птиц и крокодилов. Оказалось, что под это определение не подпадают многие формы, традиционно относимые к архозаврам (такие, как Archosaurus rossicus и (или) Protorosaurus speneri). Для объединения их с архозаврами в узком смысле Готье ввёл имя Archosauriformes. Другие исследователи предпочитают использование имени Archosauria в широком смысле, а для обозначения кроновой группы (архозавров в узком смысле) использует имя Avesuchia. Оба варианта достаточно распространены, что создаёт путаницу.
 Отличительные особенности Простейшими и наиболее согласованными синапоморфиями группы архозавров являются следующие характеристики: Зубы находятся в ячейках — теках (так называемые текодонтные зубы), которые снижают риск потерять их в процессе питания. Данная характеристика послужила основой для выделяемой ранее палеонтологами группы текодонтов, включавшей всех или почти всех архозавров. Между глазницей и ноздрёй в крыше черепа имеется ещё одно, так называемое суборбитальное или предглазничное, окно. Большинство ранних архозавров имели относительно длинные тяжёлые черепа, отверстие позволило снизить общий вес черепа. Размер суборбитальных отверстий часто превышает размер глазниц. Нижняя челюсть имеет небольшие нижнечелюстные отверстия, которые незначительно облегчают её вес. Бедренная кость имеет четвёртый вертел — бугорок для крепления мышц. Достаточно незначительная деталь, которая, тем не менее, могла сделать возможной эволюцию динозавров (все ранние динозавры и много поздних были двуногими), а также может быть связана со способностью архозавров или их непосредственных потомков пережить пермско-триасовое вымирание. Для защиты от обезвоживания кожу покрывают роговые щитки, первоначально прямоугольные и расположенные рядами вдоль тела, более крупные в верхней части тела для большей защиты и более мелкие снизу для лучшей гибкости. Щитки постепенно нарастают с внутренней стороны и медленно истираются с наружной. У крокодилов (кроме некоторых вымерших), некоторых динозавров и ранних архозавров щитки укреплены костными остеодермами. Некоторые щитки и/или остеодермы у многих видов видоизменялись или срастались. У гомойотермных видов, тетануров, часть роговых щитков превратилась в пух; каждое перо снабдилось мышцей, рефлекторно поднимающей или опускающей перо в зависимости от окружающей температуры. Известковая оболочка яиц, удерживающая воду внутри и не препятствующая газообмену (у гомойотермных видов — ещё и выдерживающая вес «наседки» ). Благодаря ей рептилии могут не прятать яйца от солнца. Такие же яйца откладывают птицы. Чтобы разбить скорлупу, у эмбриона перед вылуплением образуется специальное приспособление для разбивания скорлупы (у крокодилов — роговые выступы над бровями, у птиц — яйцевой зуб).
Отличительные особенности Простейшими и наиболее согласованными синапоморфиями группы архозавров являются следующие характеристики: Зубы находятся в ячейках — теках (так называемые текодонтные зубы), которые снижают риск потерять их в процессе питания. Данная характеристика послужила основой для выделяемой ранее палеонтологами группы текодонтов, включавшей всех или почти всех архозавров. Между глазницей и ноздрёй в крыше черепа имеется ещё одно, так называемое суборбитальное или предглазничное, окно. Большинство ранних архозавров имели относительно длинные тяжёлые черепа, отверстие позволило снизить общий вес черепа. Размер суборбитальных отверстий часто превышает размер глазниц. Нижняя челюсть имеет небольшие нижнечелюстные отверстия, которые незначительно облегчают её вес. Бедренная кость имеет четвёртый вертел — бугорок для крепления мышц. Достаточно незначительная деталь, которая, тем не менее, могла сделать возможной эволюцию динозавров (все ранние динозавры и много поздних были двуногими), а также может быть связана со способностью архозавров или их непосредственных потомков пережить пермско-триасовое вымирание. Для защиты от обезвоживания кожу покрывают роговые щитки, первоначально прямоугольные и расположенные рядами вдоль тела, более крупные в верхней части тела для большей защиты и более мелкие снизу для лучшей гибкости. Щитки постепенно нарастают с внутренней стороны и медленно истираются с наружной. У крокодилов (кроме некоторых вымерших), некоторых динозавров и ранних архозавров щитки укреплены костными остеодермами. Некоторые щитки и/или остеодермы у многих видов видоизменялись или срастались. У гомойотермных видов, тетануров, часть роговых щитков превратилась в пух; каждое перо снабдилось мышцей, рефлекторно поднимающей или опускающей перо в зависимости от окружающей температуры. Известковая оболочка яиц, удерживающая воду внутри и не препятствующая газообмену (у гомойотермных видов — ещё и выдерживающая вес «наседки» ). Благодаря ей рептилии могут не прятать яйца от солнца. Такие же яйца откладывают птицы. Чтобы разбить скорлупу, у эмбриона перед вылуплением образуется специальное приспособление для разбивания скорлупы (у крокодилов — роговые выступы над бровями, у птиц — яйцевой зуб).
 Расцвет архозавров Большинство преобладавших среди наземных позвоночных пермского периода синапсид исчезло в ходе пермско-триасового вымирания. Единственными крупными наземными животными, пережившими вымирание, оказались травоядные листрозавры. Какое-то время эти звероподобные рептилии были самыми многочисленными наземными животными на Земле, однако уже в раннем триасе преобладающими наземными животными стали архозавры.
Расцвет архозавров Большинство преобладавших среди наземных позвоночных пермского периода синапсид исчезло в ходе пермско-триасового вымирания. Единственными крупными наземными животными, пережившими вымирание, оказались травоядные листрозавры. Какое-то время эти звероподобные рептилии были самыми многочисленными наземными животными на Земле, однако уже в раннем триасе преобладающими наземными животными стали архозавры.
 Основные типы архозавров С 1970 -х годов учёные классифицируют архозавров в основном по строению их пояса конечностей. У самых ранних архозавров были примитивные мезотарзальные кости: таранная (talus) и пяточная (calcaneus) кости прикреплялись костным швом к большой берцовой и малоберцовой костям, и это соединение изгибалось в месте контакта со ступнёй. Строение бедра и локомоция Тазовый пояс и положение задних конечностей, справа налево: примитивных рептилий, динозавров и млекопитающих, равизухий Как и у других ранних тетраподов, конечности примитивных архозавров были широко расставлены по бокам тела. С середины триаса у некоторых групп архозавров появляется изменение бедренного сустава: в отличие от латерального положения задних конечностей (по бокам от туловища), у архозавров развивается парасагиттальное положение, при котором тело приподнято над землёй и опирается на вертикально направленную бедренную кость. Достигается это в разных группах двумя путями: у равизухий за счёт поворота вниз вертлюжной впадины, а у линии, ведущей к динозаврам, — за счёт поворота внутрь головки бедренной кости, как и у млекопитающих[6]. Такое изменение позволило архозаврам (или же потребовало от них) перейти к новому типу локомоции — двуногой ходьбе (бипедализму). Подобное парасигательное строение появилось ещё в перми у горгонопсид, представителей другой ветви амниотов — синапсид.
Основные типы архозавров С 1970 -х годов учёные классифицируют архозавров в основном по строению их пояса конечностей. У самых ранних архозавров были примитивные мезотарзальные кости: таранная (talus) и пяточная (calcaneus) кости прикреплялись костным швом к большой берцовой и малоберцовой костям, и это соединение изгибалось в месте контакта со ступнёй. Строение бедра и локомоция Тазовый пояс и положение задних конечностей, справа налево: примитивных рептилий, динозавров и млекопитающих, равизухий Как и у других ранних тетраподов, конечности примитивных архозавров были широко расставлены по бокам тела. С середины триаса у некоторых групп архозавров появляется изменение бедренного сустава: в отличие от латерального положения задних конечностей (по бокам от туловища), у архозавров развивается парасагиттальное положение, при котором тело приподнято над землёй и опирается на вертикально направленную бедренную кость. Достигается это в разных группах двумя путями: у равизухий за счёт поворота вниз вертлюжной впадины, а у линии, ведущей к динозаврам, — за счёт поворота внутрь головки бедренной кости, как и у млекопитающих[6]. Такое изменение позволило архозаврам (или же потребовало от них) перейти к новому типу локомоции — двуногой ходьбе (бипедализму). Подобное парасигательное строение появилось ещё в перми у горгонопсид, представителей другой ветви амниотов — синапсид.
 Питание Большинство архозавров — хищники, однако в отдельных группах есть исключения: травоядные некоторые ящеротазовые динозавры (зауроподы, прозавроподы, теризинозавры) травоядные птицетазовые динозавры травоядные, возможно, некоторые птерозавры (тапейяра) этозавры (Aetosauria) — травоядные известно несколько травоядных крокодилов, например, симозух (Simosuchus) большие крокодилы Stomatosuchus могли питаться путём фильтрации.
Питание Большинство архозавров — хищники, однако в отдельных группах есть исключения: травоядные некоторые ящеротазовые динозавры (зауроподы, прозавроподы, теризинозавры) травоядные птицетазовые динозавры травоядные, возможно, некоторые птерозавры (тапейяра) этозавры (Aetosauria) — травоядные известно несколько травоядных крокодилов, например, симозух (Simosuchus) большие крокодилы Stomatosuchus могли питаться путём фильтрации.

 Надотряд Dinosauria. Динозавры T 2 -K Диноза вры (лат. Dinosauria, от др. -греч. δεινός — страшный, ужасный, опасный и σαῦρος — ящер, ящерица) — надотряд наземных позвоночных животных, доминировавших на Земле в мезозойскую эру — в течение более 160 миллионов лет, начиная с позднего триасового периода (приблизительно 225 млн лет назад) до конца мелового периода (около 65 млн лет назад), когда большинство из них стали вымирать на стыке мелового и третичного периодов во время крупномасштабного исчезновения животных и многих разновидностей растений в относительно короткий геологический период истории. Ископаемые останки динозавров обнаружены на всех континентах планеты. Ныне палеонтологами описано более 500 различных родов и более чем 1000 различных видов, которые чётко делятся на две группы — птицетазовых и ящеротазовых
Надотряд Dinosauria. Динозавры T 2 -K Диноза вры (лат. Dinosauria, от др. -греч. δεινός — страшный, ужасный, опасный и σαῦρος — ящер, ящерица) — надотряд наземных позвоночных животных, доминировавших на Земле в мезозойскую эру — в течение более 160 миллионов лет, начиная с позднего триасового периода (приблизительно 225 млн лет назад) до конца мелового периода (около 65 млн лет назад), когда большинство из них стали вымирать на стыке мелового и третичного периодов во время крупномасштабного исчезновения животных и многих разновидностей растений в относительно короткий геологический период истории. Ископаемые останки динозавров обнаружены на всех континентах планеты. Ныне палеонтологами описано более 500 различных родов и более чем 1000 различных видов, которые чётко делятся на две группы — птицетазовых и ящеротазовых

![Ящеротазовые динозавры Ящерота зовые (лат. Saurischia) — один из двух главных отрядов динозавров[1][2]. Отличаются](https://present5.com/presentation/3/26512404_64559167.pdf-img/26512404_64559167.pdf-46.jpg "Ящеротазовые динозавры Ящерота зовые (лат. Saurischia) — один из двух главных отрядов динозавров[1][2]. Отличаются") Ящеротазовые динозавры Ящерота зовые (лат. Saurischia) — один из двух главных отрядов динозавров[1][2]. Отличаются тем, что лобковые кости первично наклонены вперёд (позднее у некоторых представителей — сегнозавры, дромеозавры и другие, лобковые кости отклоняются назад). К ящеротазовым относятся: Крупные растительноядные зауроподы, ходившие на четырех ногах. В среднем более мелкие плотоядные тероподы, передвигавшиеся на двух задних конечностях (ранняя юра — поздний мел). Этот отряд в свою очередь разделяется на две основные группы: Тероподы, или звероногие (Theropoda) К тероподам относятся двуногие, плотоядные динозавры. завроподоморфы (Sauropodomorpha).
Ящеротазовые динозавры Ящерота зовые (лат. Saurischia) — один из двух главных отрядов динозавров[1][2]. Отличаются тем, что лобковые кости первично наклонены вперёд (позднее у некоторых представителей — сегнозавры, дромеозавры и другие, лобковые кости отклоняются назад). К ящеротазовым относятся: Крупные растительноядные зауроподы, ходившие на четырех ногах. В среднем более мелкие плотоядные тероподы, передвигавшиеся на двух задних конечностях (ранняя юра — поздний мел). Этот отряд в свою очередь разделяется на две основные группы: Тероподы, или звероногие (Theropoda) К тероподам относятся двуногие, плотоядные динозавры. завроподоморфы (Sauropodomorpha).

 Род Tarbosaurus T 2 -K 2 Тарбозавр (лат. Tarbosaurus, от др. -греч. τάρβος — ужас, почтение и σαῦρος — ящерица) — род гигантских (до 12 метров длиной) хищных ящеротазовых динозавров семейства тираннозаврид, живших в позднем меловом периоде (около 70— 65 млн лет назад) на территории нынешних Монголии и Китая. Обнаруженные с 1946 года останки нескольких десятков особей тарбозавра, включая целые черепа и скелеты, позволяют воссоздать его внешний облик и сделать выводы об образе жизни, а также проследить его эволюцию. С 1955 года, когда название Tarbosaurus было впервые использовано Е. А. Малеевым, исследователи дали обнаруженным останкам несколько видовых названий. Однако к началу XXI века в научном сообществе сомнений не вызывает существование только одного из этих видов, Tarbosaurus bataar (реже, из-за значительного сходства с североамериканскими тираннозаврами, их классифицируют как Tyrannosaurus bataar). Близкими родичами тарбозавра также являются обнаруженные в Монголии алиорамы. Тарбозавры были крупными двуногими хищниками весом до 4— 6 тонн, с непропорционально маленькими относительно остального тела двупалыми передними конечностями. В пасти тарбозавра располагались около шести десятков зубов до 85 мм длиной. Хотя ряд исследователей считает тарбозавров падальщиками, более распространена точка зрения, согласно которой они были высшими хищниками своей эпохи и региона, охотившимися на крупных травоядных динозавров во влажных речных поймах.
Род Tarbosaurus T 2 -K 2 Тарбозавр (лат. Tarbosaurus, от др. -греч. τάρβος — ужас, почтение и σαῦρος — ящерица) — род гигантских (до 12 метров длиной) хищных ящеротазовых динозавров семейства тираннозаврид, живших в позднем меловом периоде (около 70— 65 млн лет назад) на территории нынешних Монголии и Китая. Обнаруженные с 1946 года останки нескольких десятков особей тарбозавра, включая целые черепа и скелеты, позволяют воссоздать его внешний облик и сделать выводы об образе жизни, а также проследить его эволюцию. С 1955 года, когда название Tarbosaurus было впервые использовано Е. А. Малеевым, исследователи дали обнаруженным останкам несколько видовых названий. Однако к началу XXI века в научном сообществе сомнений не вызывает существование только одного из этих видов, Tarbosaurus bataar (реже, из-за значительного сходства с североамериканскими тираннозаврами, их классифицируют как Tyrannosaurus bataar). Близкими родичами тарбозавра также являются обнаруженные в Монголии алиорамы. Тарбозавры были крупными двуногими хищниками весом до 4— 6 тонн, с непропорционально маленькими относительно остального тела двупалыми передними конечностями. В пасти тарбозавра располагались около шести десятков зубов до 85 мм длиной. Хотя ряд исследователей считает тарбозавров падальщиками, более распространена точка зрения, согласно которой они были высшими хищниками своей эпохи и региона, охотившимися на крупных травоядных динозавров во влажных речных поймах.

 — род ящеротазовых динозавров из группы") Род Diplodocus J 3 Диплодо к (лат. Diplodocus) — род ящеротазовых динозавров из группы зауропод. Первый окаменелый скелет был найден в 1877 году в Скалистых горах (Колорадо) палеонтологом С. У. Уилистоном (Samuel Wendell Williston). Позже были обнаружены и другие останки в отложениях юрского периода (формация Моррисон). Все они датируются возрастом 150— 147 млн лет назад. Является крупнейшим из динозавров, известных по полным скелетам. Это был один из настоящих гигантов позднеюрского периода. Диплодок достигал в длину 27 метров, но по мнению ученых размеры самых крупных особей могли и вовсе достигать 35 метров. [1] Из них большая часть приходилась на шею и хвост. Кости шеи и хвоста у диплодока были полыми. Вес диплодока по одним оценкам составлял 10— 20 тонн[2], а по другим достигал 20— 80 тонн[3]. Диплодок имел длинную шею, состоящую из 15 позвонков, возможно, заполненных сообщающимися воздушными мешками. Череп диплодока имел непарное носовое отверстие, расположенное не на кончике морды, а в верхней части головы впереди глаз. Зубы в форме узких лопаточек имелись только в передней части рта. Конечности диплодока были пятипалыми, с короткими массивными когтями на внутренних пальцах. Длинный хвост диплодока, заканчивавшийся тонким «хлыстом» , служил прекрасным орудием защиты. Вероятно, диплодоки вели стадный образ жизни, питаясь листьями невысоких деревьев. Не умея жевать, они заглатывали камни, которые помогали им перетирать пищу. Подобно брахиозавру, диплодок передвигался на четырех ногах, причём задние были длиннее передних.
Род Diplodocus J 3 Диплодо к (лат. Diplodocus) — род ящеротазовых динозавров из группы зауропод. Первый окаменелый скелет был найден в 1877 году в Скалистых горах (Колорадо) палеонтологом С. У. Уилистоном (Samuel Wendell Williston). Позже были обнаружены и другие останки в отложениях юрского периода (формация Моррисон). Все они датируются возрастом 150— 147 млн лет назад. Является крупнейшим из динозавров, известных по полным скелетам. Это был один из настоящих гигантов позднеюрского периода. Диплодок достигал в длину 27 метров, но по мнению ученых размеры самых крупных особей могли и вовсе достигать 35 метров. [1] Из них большая часть приходилась на шею и хвост. Кости шеи и хвоста у диплодока были полыми. Вес диплодока по одним оценкам составлял 10— 20 тонн[2], а по другим достигал 20— 80 тонн[3]. Диплодок имел длинную шею, состоящую из 15 позвонков, возможно, заполненных сообщающимися воздушными мешками. Череп диплодока имел непарное носовое отверстие, расположенное не на кончике морды, а в верхней части головы впереди глаз. Зубы в форме узких лопаточек имелись только в передней части рта. Конечности диплодока были пятипалыми, с короткими массивными когтями на внутренних пальцах. Длинный хвост диплодока, заканчивавшийся тонким «хлыстом» , служил прекрасным орудием защиты. Вероятно, диплодоки вели стадный образ жизни, питаясь листьями невысоких деревьев. Не умея жевать, они заглатывали камни, которые помогали им перетирать пищу. Подобно брахиозавру, диплодок передвигался на четырех ногах, причём задние были длиннее передних.

 — один из двух отрядов динозавров. Несмотря на своё") Птицетазовые динозавры Птицетазовые (лат. Ornithischia) — один из двух отрядов динозавров. Несмотря на своё название, не они, а ящеротазовые динозавры по одной из версий были предками птиц. Были травоядными (за исключением ранних всеядных видов), меньших размеров, чем ящеротазовые зауроподы. Кроме самых примитивных, имеют приспособления к питанию растительной пищей: сложный кишечник и щёки. Ранние представители были двуногими, позже птицетазовые повторно освоили хождение на четырёх конечностях. В ходе эволюции у поздних представителей отряда концевые фаланги пальцев стали более плоскими и приобрели вид копыт. У многих развились приспособления для защиты от хищников: броня, рога, шипы. Строение таза Таз имел четырёхлучевое строение. Лобковая кость имела вырост, направленный назад вдоль седалищной кости (у ящеротазовых кости были практически перпендикулярны). Лобковая кость часто образует новый вырост, направленный вперёд параллельно позвоночнику. Подобное строение таза развивалась у динозавров независимо трижды: у птицетазовых, у надсемейства Therizinosauroidea и семейства Dromaeosauridae. Последнее и является наиболее вероятным предком птиц. У некоторых видов (Hypsilophodon, Talenkauen и Thescelosaurus) имеются межреберные костные пластины в передней части ребер. Некоторые исследователи предлагают, что эти пластины выполняли биомеханические функции, помогали в дыхании либо для обеспечения повышенной жесткости передней части грудной клетки, в то время как другие полагают, они служили для оборонительных функций.
Птицетазовые динозавры Птицетазовые (лат. Ornithischia) — один из двух отрядов динозавров. Несмотря на своё название, не они, а ящеротазовые динозавры по одной из версий были предками птиц. Были травоядными (за исключением ранних всеядных видов), меньших размеров, чем ящеротазовые зауроподы. Кроме самых примитивных, имеют приспособления к питанию растительной пищей: сложный кишечник и щёки. Ранние представители были двуногими, позже птицетазовые повторно освоили хождение на четырёх конечностях. В ходе эволюции у поздних представителей отряда концевые фаланги пальцев стали более плоскими и приобрели вид копыт. У многих развились приспособления для защиты от хищников: броня, рога, шипы. Строение таза Таз имел четырёхлучевое строение. Лобковая кость имела вырост, направленный назад вдоль седалищной кости (у ящеротазовых кости были практически перпендикулярны). Лобковая кость часто образует новый вырост, направленный вперёд параллельно позвоночнику. Подобное строение таза развивалась у динозавров независимо трижды: у птицетазовых, у надсемейства Therizinosauroidea и семейства Dromaeosauridae. Последнее и является наиболее вероятным предком птиц. У некоторых видов (Hypsilophodon, Talenkauen и Thescelosaurus) имеются межреберные костные пластины в передней части ребер. Некоторые исследователи предлагают, что эти пластины выполняли биомеханические функции, помогали в дыхании либо для обеспечения повышенной жесткости передней части грудной клетки, в то время как другие полагают, они служили для оборонительных функций.
 — род позднеюрских травоядных") Род Stegosaurus J-K 1 Стегозавры (лат. Stegosaurus — «крышеящер» ) — род позднеюрских травоядных динозавров, существовавший 155— 145 млн лет назад. В его составе выделено три вида. Благодаря шипам на хвосте и костяным пластинам на спине являются одними из самых узнаваемых динозавров.
Род Stegosaurus J-K 1 Стегозавры (лат. Stegosaurus — «крышеящер» ) — род позднеюрских травоядных динозавров, существовавший 155— 145 млн лет назад. В его составе выделено три вида. Благодаря шипам на хвосте и костяным пластинам на спине являются одними из самых узнаваемых динозавров.
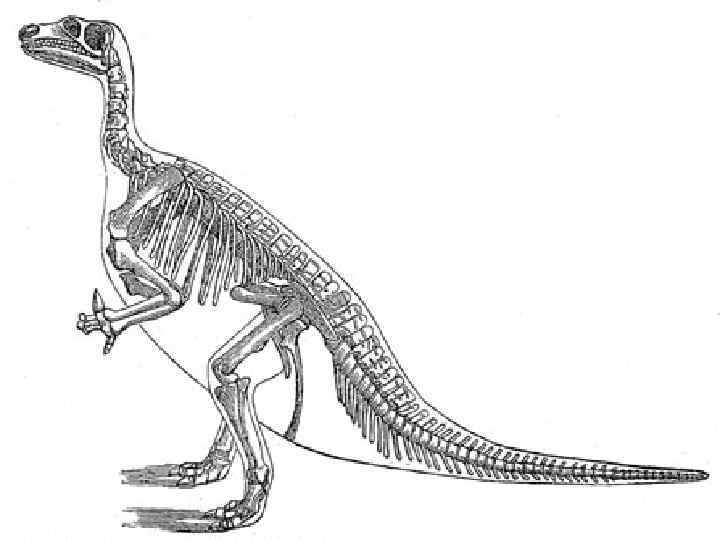
 — род растительноядных птицетазовых динозавров,") Род Iguanodon K 1 Игуанодонт, также игуанодон (лат. Iguanodon) — род растительноядных птицетазовых динозавров, живших в первой половине мелового периода 140 -120 млн. лет назад на территории современных Европы, Северной Америки, Азии, Африки. Был открыт в 1822 Гидеоном Мантеллом и был вторым получившим название - динозавр, после мегалозавра. Игуанодоны были большими растительноядными динозаврами, переходившими от передвижения на двух ногах к передвижению на четырёх. В передней части челюсти игуанодоны имели клюв, состоящий из кератина, за ним следовали зубы, подобные зубам игуаны, но больших размеров и более частые. Передние конечности были примерно на четверть короче задних, три центральные пальца на них были приспособлены для опоры. На больших пальцах располагались шипы, использовавшиеся предположительно для защиты. В начале XIX века эти шипы считались рогами и помещались палеонтологами на нос животного, истинное их положение было показано позже. «Мизинцы» в противоположность всем остальным пальцам были длинными и гибкими. На задних ногах, приспособленных для ходьбы, но не для бега, было лишь три пальца. Позвоночник и хвост поддерживались окостеневшими сухожилиями.
Род Iguanodon K 1 Игуанодонт, также игуанодон (лат. Iguanodon) — род растительноядных птицетазовых динозавров, живших в первой половине мелового периода 140 -120 млн. лет назад на территории современных Европы, Северной Америки, Азии, Африки. Был открыт в 1822 Гидеоном Мантеллом и был вторым получившим название - динозавр, после мегалозавра. Игуанодоны были большими растительноядными динозаврами, переходившими от передвижения на двух ногах к передвижению на четырёх. В передней части челюсти игуанодоны имели клюв, состоящий из кератина, за ним следовали зубы, подобные зубам игуаны, но больших размеров и более частые. Передние конечности были примерно на четверть короче задних, три центральные пальца на них были приспособлены для опоры. На больших пальцах располагались шипы, использовавшиеся предположительно для защиты. В начале XIX века эти шипы считались рогами и помещались палеонтологами на нос животного, истинное их положение было показано позже. «Мизинцы» в противоположность всем остальным пальцам были длинными и гибкими. На задних ногах, приспособленных для ходьбы, но не для бега, было лишь три пальца. Позвоночник и хвост поддерживались окостеневшими сухожилиями.

 K 2 Трицера топсы (лат. Triceratops, от др. -греч.") Род Triceratops ( «рогатые» динозавры) K 2 Трицера топсы (лат. Triceratops, от др. -греч. τρι, tri — «три» , κέρας, keras — «рог» и ωψ, ops — «морда» , «лицо» ) — вымерший род растительноядных динозавров из семейства цератопсид, существовали в конце маастрихтского яруса мелового периода, примерно от 68 до 65 миллионов лет назад на территории современной Северной Америки[4]. Имея большой костяной воротник, три рога на морде, крупные толстые конечности и сходное с носорогом телосложение, являются одними из наиболее узнаваемых динозавров. Хотя, возможно, самый крупный хищник того времени тираннозавр, мог охотиться на трицератопсов, неясно могли ли они сражаться друг с другом, как это часто изображается. Полный скелет трицератопса до сих пор не найден, было обнаружено множество частичных скелетов, с момента открытия в 1887 году. О функциях наростов и рогов давно идут дискуссии. Традиционно они рассматриваются в качестве оборонительного оружия против хищников, но самые новые теории утверждают, что более вероятно, что они, были использованы в ухаживании и для демонстрации, так же, как и рога современных оленей, горных козлов, или жуков носорогов. Трицератопс является наиболее известным из цератопсид, хотя точное расположение этого рода в семействе было предметом спора между палеонтологами. В настоящее время валидными считаются два вида Трицератопс хорридус и Трицератопс прорсус, хотя ошибочно были описаны и другие. Последние исследования показали, что трицератопс являлся молодой особью торозавра, который раньше выделялся в отдельный вид родственных
Род Triceratops ( «рогатые» динозавры) K 2 Трицера топсы (лат. Triceratops, от др. -греч. τρι, tri — «три» , κέρας, keras — «рог» и ωψ, ops — «морда» , «лицо» ) — вымерший род растительноядных динозавров из семейства цератопсид, существовали в конце маастрихтского яруса мелового периода, примерно от 68 до 65 миллионов лет назад на территории современной Северной Америки[4]. Имея большой костяной воротник, три рога на морде, крупные толстые конечности и сходное с носорогом телосложение, являются одними из наиболее узнаваемых динозавров. Хотя, возможно, самый крупный хищник того времени тираннозавр, мог охотиться на трицератопсов, неясно могли ли они сражаться друг с другом, как это часто изображается. Полный скелет трицератопса до сих пор не найден, было обнаружено множество частичных скелетов, с момента открытия в 1887 году. О функциях наростов и рогов давно идут дискуссии. Традиционно они рассматриваются в качестве оборонительного оружия против хищников, но самые новые теории утверждают, что более вероятно, что они, были использованы в ухаживании и для демонстрации, так же, как и рога современных оленей, горных козлов, или жуков носорогов. Трицератопс является наиболее известным из цератопсид, хотя точное расположение этого рода в семействе было предметом спора между палеонтологами. В настоящее время валидными считаются два вида Трицератопс хорридус и Трицератопс прорсус, хотя ошибочно были описаны и другие. Последние исследования показали, что трицератопс являлся молодой особью торозавра, который раньше выделялся в отдельный вид родственных


 — отряд") Надотряд Pterosauria. Птерозавры J-K Птероза вры (лат. Pterosauria — «летающие ящеры» ) — отряд вымерших летающих архозавров. Жили в мезозое на протяжении более 200 млн лет. Последние представители относятся ко времени вымирания динозавров, около 65 млн лет назад. История открытия В середине XVIII в. учёные извлекли из юрских зольнхофенских известняков (Бавария, Германия) несколько полных скелетов небольшого животного. Это существо настолько отличалось от всех современных животных, что долгие годы учёные терялись в догадках, что это за животное. Одни сочли его неведомой уткой, другие — летучей мышью и даже миниатюрным драконом. И только в 1809 году великий французский зоолог и палеонтолог Жорж Кювье окончательно установил, что найденные скелеты принадлежали вымершей летающей рептилии. Кювье назвал её птеродактилем ( «пальцекрыл» ). Позднее, в XIX веке вокруг птерозавров развернулась ещё одна дискуссия. Ведущие британские учёные того времени Ричард Оуэн и Гарри Сили спорили о том, были ли птерозавры холоднокровными рептилиями или теплокровными «примитивными птицами» . Картину прояснили последующие ископаемые находки. Без сомнения, птерозавры являлись рептилиями (как и полагал Оуэн), но теплокровными.
Надотряд Pterosauria. Птерозавры J-K Птероза вры (лат. Pterosauria — «летающие ящеры» ) — отряд вымерших летающих архозавров. Жили в мезозое на протяжении более 200 млн лет. Последние представители относятся ко времени вымирания динозавров, около 65 млн лет назад. История открытия В середине XVIII в. учёные извлекли из юрских зольнхофенских известняков (Бавария, Германия) несколько полных скелетов небольшого животного. Это существо настолько отличалось от всех современных животных, что долгие годы учёные терялись в догадках, что это за животное. Одни сочли его неведомой уткой, другие — летучей мышью и даже миниатюрным драконом. И только в 1809 году великий французский зоолог и палеонтолог Жорж Кювье окончательно установил, что найденные скелеты принадлежали вымершей летающей рептилии. Кювье назвал её птеродактилем ( «пальцекрыл» ). Позднее, в XIX веке вокруг птерозавров развернулась ещё одна дискуссия. Ведущие британские учёные того времени Ричард Оуэн и Гарри Сили спорили о том, были ли птерозавры холоднокровными рептилиями или теплокровными «примитивными птицами» . Картину прояснили последующие ископаемые находки. Без сомнения, птерозавры являлись рептилиями (как и полагал Оуэн), но теплокровными.
 Крылья Их крылья представляли собой складки кожи, натянутые между боками тела и очень длинным четвёртым пальцем передних конечностей. Скелет обладал облегченной конструкцией: тонкие полые кости, лёгкий череп почти что сетчатой структуры, грудина имела киль, как у птиц. Строение скелета крыльев и плечевого пояса указывает на наличие мощной махательной мускулатуры. Необходимость в полёте возникла в связи с питанием рыбой и трупами, плавающими в воде, но питались также насекомыми. В настоящее время споры о способности птерозавров летать не утихают. Одни специалисты полагают, что гигантские рептилии могли преодолевать огромные расстояния по воздуху, а другие утверждают, что если и могли, то лишь в тихую погоду. На полуводный образ жизни указывают перепончатые лапы. Вытянутые в клюв челюсти могли нести зубы. У некоторых поздних форм, например, птеранодонов, были беззубые челюсти. Зубы некоторых видов свидетельствуют о питании планктоном. Птерозавры имели достаточно хорошо развитый мозг (преимущественно за счет мозжечка, отвечающего за координацию движений) и острое зрение. У них были редуцированны (уменьшены) обонятельные доли и развиты зрительные доли, что говорит о том, что летучих ящеров больше интересовало, как что-то выглядело, нежели как оно пахло. Тело было покрыто своеобразной «шерстью» , что позволяет делать предположения о теплокровности птерозавров. На теплокровность указывает и обильное кровоснабжение костей, подобное тому, что наблюдается у птиц и млекопитающих. Возможно, что «шерсть» — это «пуховое оперение» молодых животных, так как у многих видов (например, Eudimorphodon) следы власоподобных структур обнаружены только на останках молодых особей. Птерозаврам для целей минимизации проблем с терморегуляцией при полноценном бодрствовании в светлое время суток просто необходимо было иметь многослойную, сложно устроенную мембрану крыла с развитой сосудистой сетью, обеспечивающей кондиционирование, а также наружные покровы, способные защитить от попадания избыточного количества прямых и отражённых солнечных лучей (например, во время полётов над водой), и гарантировать доставку крови к голове одной и той же температуры.
Крылья Их крылья представляли собой складки кожи, натянутые между боками тела и очень длинным четвёртым пальцем передних конечностей. Скелет обладал облегченной конструкцией: тонкие полые кости, лёгкий череп почти что сетчатой структуры, грудина имела киль, как у птиц. Строение скелета крыльев и плечевого пояса указывает на наличие мощной махательной мускулатуры. Необходимость в полёте возникла в связи с питанием рыбой и трупами, плавающими в воде, но питались также насекомыми. В настоящее время споры о способности птерозавров летать не утихают. Одни специалисты полагают, что гигантские рептилии могли преодолевать огромные расстояния по воздуху, а другие утверждают, что если и могли, то лишь в тихую погоду. На полуводный образ жизни указывают перепончатые лапы. Вытянутые в клюв челюсти могли нести зубы. У некоторых поздних форм, например, птеранодонов, были беззубые челюсти. Зубы некоторых видов свидетельствуют о питании планктоном. Птерозавры имели достаточно хорошо развитый мозг (преимущественно за счет мозжечка, отвечающего за координацию движений) и острое зрение. У них были редуцированны (уменьшены) обонятельные доли и развиты зрительные доли, что говорит о том, что летучих ящеров больше интересовало, как что-то выглядело, нежели как оно пахло. Тело было покрыто своеобразной «шерстью» , что позволяет делать предположения о теплокровности птерозавров. На теплокровность указывает и обильное кровоснабжение костей, подобное тому, что наблюдается у птиц и млекопитающих. Возможно, что «шерсть» — это «пуховое оперение» молодых животных, так как у многих видов (например, Eudimorphodon) следы власоподобных структур обнаружены только на останках молодых особей. Птерозаврам для целей минимизации проблем с терморегуляцией при полноценном бодрствовании в светлое время суток просто необходимо было иметь многослойную, сложно устроенную мембрану крыла с развитой сосудистой сетью, обеспечивающей кондиционирование, а также наружные покровы, способные защитить от попадания избыточного количества прямых и отражённых солнечных лучей (например, во время полётов над водой), и гарантировать доставку крови к голове одной и той же температуры.
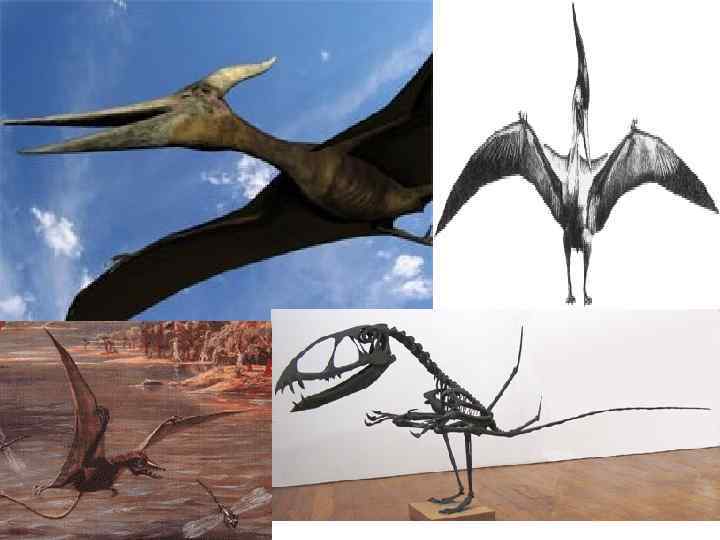
 — род вымерших рептилий отряда летающих ящеров") Род Rhamphorhynchus J 3 Рамфоринхи (лат. Rhamphorhynchus) — род вымерших рептилий отряда летающих ящеров (птерозавров), живших в юрском периоде (около 170— 140 млн лет назад) на территории Европы (Великобритания, Испания и Германия) и Африки (Ангола и Танзания). Впервые описан палеонтологом Мейером (Meyer) в 1847 году.
Род Rhamphorhynchus J 3 Рамфоринхи (лат. Rhamphorhynchus) — род вымерших рептилий отряда летающих ящеров (птерозавров), живших в юрском периоде (около 170— 140 млн лет назад) на территории Европы (Великобритания, Испания и Германия) и Африки (Ангола и Танзания). Впервые описан палеонтологом Мейером (Meyer) в 1847 году.
 Рoд Pteranodon K 2 Птеранодон (от греч. πτερόν — «крыло» и αν-οδων — «беззубый» ) — род птерозавров. Известен с верхнего мела Северной Америки. Отличался крупным выростом на голове и беззубым клювом.
Рoд Pteranodon K 2 Птеранодон (от греч. πτερόν — «крыло» и αν-οδων — «беззубый» ) — род птерозавров. Известен с верхнего мела Северной Америки. Отличался крупным выростом на голове и беззубым клювом.
 или тероморфы (лат.") Подкласс Synapsida. Зверообразные C 3 -J 2 Синапси ды (лат. Synapsida) или тероморфы (лат. Theromorpha, от др. греч. θήρ, θηρίον — «зверь» и μορφή — «форма» — звероподобные) — звероящеры — таксономическая группа в составе амниот (наряду с завропсидами), возникшая около 320 млн лет назад. Одна из групп синапсид — терапсиды — была предком млекопитающих. В начале XX века синапсиды считались одним из подклассов класса рептилий. Однако по мере развития филогенетики рептилии были признаны парафилетической группой, была в корне пересмотрена их классификация. Новая классификация на основе кладистики появилась в конце 1980 -х годов. Характерными для синапсид были отверстия в щёчных костях (боковая височная яма с двух сторон черепа), служащие для выхода челюстных мышц и позволявшие более эффективно кусать.
Подкласс Synapsida. Зверообразные C 3 -J 2 Синапси ды (лат. Synapsida) или тероморфы (лат. Theromorpha, от др. греч. θήρ, θηρίον — «зверь» и μορφή — «форма» — звероподобные) — звероящеры — таксономическая группа в составе амниот (наряду с завропсидами), возникшая около 320 млн лет назад. Одна из групп синапсид — терапсиды — была предком млекопитающих. В начале XX века синапсиды считались одним из подклассов класса рептилий. Однако по мере развития филогенетики рептилии были признаны парафилетической группой, была в корне пересмотрена их классификация. Новая классификация на основе кладистики появилась в конце 1980 -х годов. Характерными для синапсид были отверстия в щёчных костях (боковая височная яма с двух сторон черепа), служащие для выхода челюстных мышц и позволявшие более эффективно кусать.

 — хищный дейноцефал группы антеозавров. Известен из") Род Titanophoneus P 2 Титанофонеус (лат. Titanophoneus) — хищный дейноцефал группы антеозавров. Известен из «средней» перми Приуралья и Поволжья (описан из Ишеево в Татарии — нижнетатарский ярус). Во многом сходен с южноафриканским антеозавром. Череп высокий и узкий, у взрослых особей развиты утолщения заглазничной и лобной области (пахиостоз). Мощные резцы и клыки, 8— 9 пар мелких заклыковых зубов. Ноги мощные, не очень длинные, с широкими кистями и стопами (возможно, имелась плавательная перепонка). Хвост длинный. В целом скелет довольно легкий. Молодые особи (первые из описанных экземпляров) достигали в длину 3 метров, черепа взрослых могли достигать 60 см в длину.
Род Titanophoneus P 2 Титанофонеус (лат. Titanophoneus) — хищный дейноцефал группы антеозавров. Известен из «средней» перми Приуралья и Поволжья (описан из Ишеево в Татарии — нижнетатарский ярус). Во многом сходен с южноафриканским антеозавром. Череп высокий и узкий, у взрослых особей развиты утолщения заглазничной и лобной области (пахиостоз). Мощные резцы и клыки, 8— 9 пар мелких заклыковых зубов. Ноги мощные, не очень длинные, с широкими кистями и стопами (возможно, имелась плавательная перепонка). Хвост длинный. В целом скелет довольно легкий. Молодые особи (первые из описанных экземпляров) достигали в длину 3 метров, черепа взрослых могли достигать 60 см в длину.


 Тревожные перспективы Одна из сложнейших задач современности – проблема замедления процесса уничтожения живой природы. Рептилии – неотъемлемая часть животного населения Земли. Допустить их исчезновение – значит признаться, что мы готовы расстаться со всей живой природой.
Тревожные перспективы Одна из сложнейших задач современности – проблема замедления процесса уничтожения живой природы. Рептилии – неотъемлемая часть животного населения Земли. Допустить их исчезновение – значит признаться, что мы готовы расстаться со всей живой природой.
 C 3") Систематика использовавшаяся в работе: Класс Reptilia. Пресмыкающиеся Подкласс Cotylosauria ( «котeлкоголовые» ) C 3 -T 1 Род Pareiasaurus ( «щекастый» ) P 2 Подкласс Synaptosauria. Синаптозавры T 3 -K Род Plesiosaurus J 1 Подкласс Jchthyopterygia. Ихтиоптеригии T-K 1 Род Ichthyosaurus J 1 Подкласс Archosauria. Архозавры T 1 - ныне Надотряд Dinosauria. Динозавры T 2 -K Ящеротазовые динозавры Род Tarbosaurus T 2 -K 2 Род Diplodocus J 3 Птицетазовые динозавры Род Stegosaurus J-K 1 Род Iguanodon K 1 Род Triceratops ( «рогатые» динозавры) K 2 Надотряд Pterosauria. Птерозавры J-K Род Rhamphorhynchus J 3 Рoд Pteranodon K 2 Подкласс Synapsida. Зверообразные C 3 -J 2 Род Titanophoneus P 2 Род Jnostrancevia P 2
Систематика использовавшаяся в работе: Класс Reptilia. Пресмыкающиеся Подкласс Cotylosauria ( «котeлкоголовые» ) C 3 -T 1 Род Pareiasaurus ( «щекастый» ) P 2 Подкласс Synaptosauria. Синаптозавры T 3 -K Род Plesiosaurus J 1 Подкласс Jchthyopterygia. Ихтиоптеригии T-K 1 Род Ichthyosaurus J 1 Подкласс Archosauria. Архозавры T 1 - ныне Надотряд Dinosauria. Динозавры T 2 -K Ящеротазовые динозавры Род Tarbosaurus T 2 -K 2 Род Diplodocus J 3 Птицетазовые динозавры Род Stegosaurus J-K 1 Род Iguanodon K 1 Род Triceratops ( «рогатые» динозавры) K 2 Надотряд Pterosauria. Птерозавры J-K Род Rhamphorhynchus J 3 Рoд Pteranodon K 2 Подкласс Synapsida. Зверообразные C 3 -J 2 Род Titanophoneus P 2 Род Jnostrancevia P 2
 В Москве есть Палеонтологический музей, в котором очень интересно побывать.
В Москве есть Палеонтологический музей, в котором очень интересно побывать.








 Спасибо за внимание Автор презентации Студент группы ГНГ-11 -1 Балашов Е. В. Преподаватель: Бакиева Л. Б.
Спасибо за внимание Автор презентации Студент группы ГНГ-11 -1 Балашов Е. В. Преподаватель: Бакиева Л. Б.