

fungi6.ppt
- Количество слайдов: 22
 КЛАСС PEZIZOMYCETES 1. Плодовые тела – апотеции, обычно сложного строения 2. Сумки оперкулятные (вскрываются крышечкой) 3. Наблюдается редукция конидиального спороношения
КЛАСС PEZIZOMYCETES 1. Плодовые тела – апотеции, обычно сложного строения 2. Сумки оперкулятные (вскрываются крышечкой) 3. Наблюдается редукция конидиального спороношения
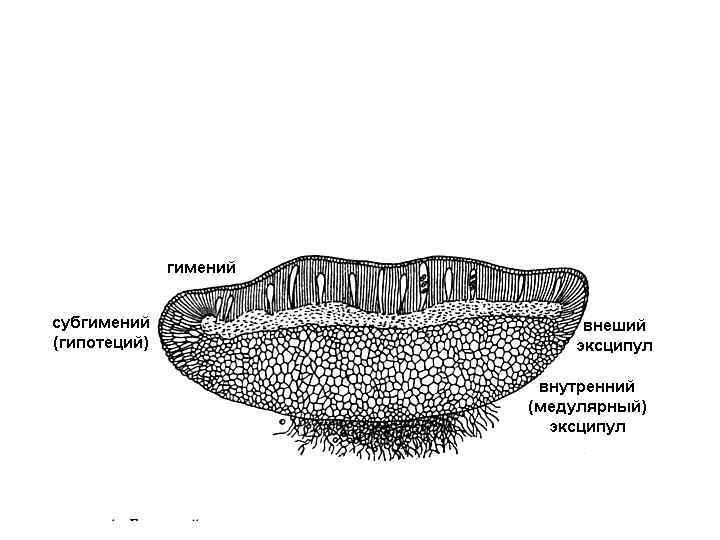
 КЛАСС LEOTIOMYCETES 1. У большинства видов – мелкие апотеции 2. Сумки иноперкулятные, вскрываются щелью Порядок Leotiales 1. Апотеции не имеют внутреннего эксципула 2. Выбрасывание спор радиосенситивное или тактиосенситивное
КЛАСС LEOTIOMYCETES 1. У большинства видов – мелкие апотеции 2. Сумки иноперкулятные, вскрываются щелью Порядок Leotiales 1. Апотеции не имеют внутреннего эксципула 2. Выбрасывание спор радиосенситивное или тактиосенситивное

 Порядок Phacidiales Апотеции формируются в темно-окрашенной строме и вскрываются продольной щелью или радиально-расходящимися трещинами (гистеротеции)
Порядок Phacidiales Апотеции формируются в темно-окрашенной строме и вскрываются продольной щелью или радиально-расходящимися трещинами (гистеротеции)

 Порядок Erysiphales 1. Плодовые тела замкнутые – клейстотеции, с упорядоченным расположением сумок на дне клейстотеция 2. Конидии развиваются цепочками на конце неразветвленного конидиеносца вследствие нарастания его кончика и отделения перегородкой (меристем- артроконидии). 3. Все представители – облигатные паразиты высших растений
Порядок Erysiphales 1. Плодовые тела замкнутые – клейстотеции, с упорядоченным расположением сумок на дне клейстотеция 2. Конидии развиваются цепочками на конце неразветвленного конидиеносца вследствие нарастания его кончика и отделения перегородкой (меристем- артроконидии). 3. Все представители – облигатные паразиты высших растений
 1. Сумки битуникатные, с двумя оболочками, наружной, плотной, разрывающейся при") КЛАСС DOTHIDEOMYCETES (Loculoascomycetes) 1. Сумки битуникатные, с двумя оболочками, наружной, плотной, разрывающейся при созревании на вершине, и внутренней, эластичной, вытягивающейся в трубку. 2. Сумки формируются в полостях сплетения гиф – аскостромах. Настоящих плодовых тел с собственными оболочками нет. Полости разрываются на вершине (перитециевидные), вследствие растворения гиф аскостромы
КЛАСС DOTHIDEOMYCETES (Loculoascomycetes) 1. Сумки битуникатные, с двумя оболочками, наружной, плотной, разрывающейся при созревании на вершине, и внутренней, эластичной, вытягивающейся в трубку. 2. Сумки формируются в полостях сплетения гиф – аскостромах. Настоящих плодовых тел с собственными оболочками нет. Полости разрываются на вершине (перитециевидные), вследствие растворения гиф аскостромы
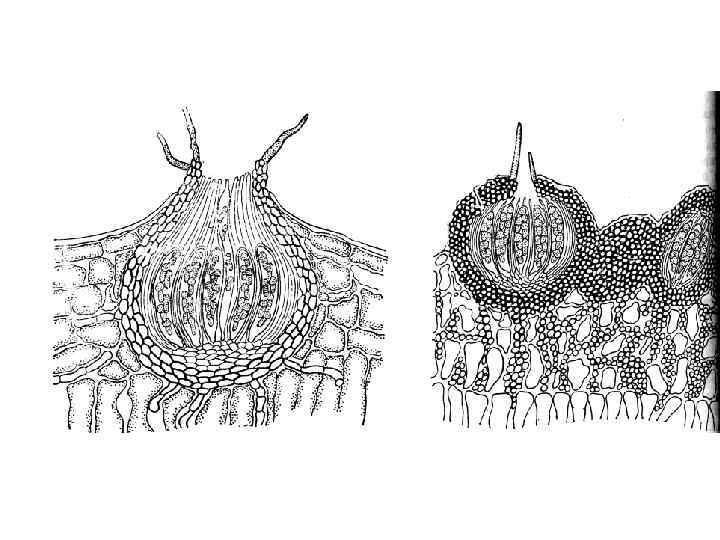
 КЛАСС LECANOROMYCETES 1. Большинство видов – лихенизированные или лихенофильные грибы 2. Плодовые тела – апотеции, хотя многие виды утратили половой процесс (несовершенные лишайники)
КЛАСС LECANOROMYCETES 1. Большинство видов – лихенизированные или лихенофильные грибы 2. Плодовые тела – апотеции, хотя многие виды утратили половой процесс (несовершенные лишайники)
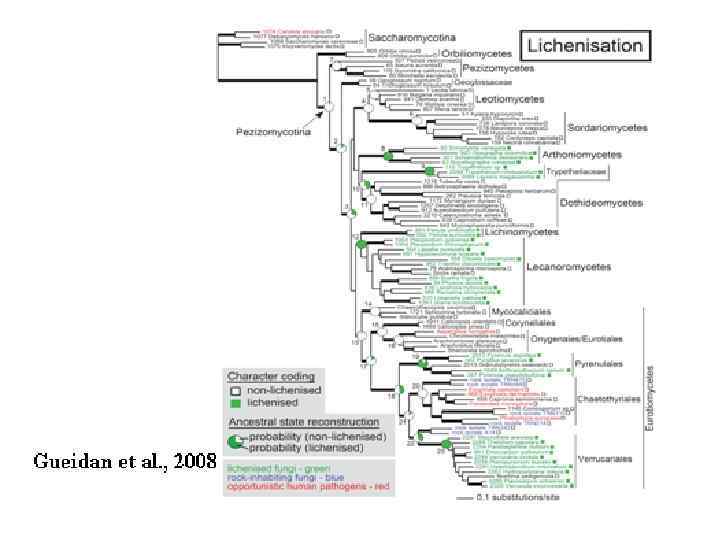
 ЛИШАЙНИКОВЫЕ МИКОБИОНТЫ около 21% от всех грибов Лишайники – около 15 000 видов. Большинство – дискомицеты, но есть – пиреномицеты и даже базидиомицеты. Аскомицеты – 98% всех лишайников. 40% всех аскомицетов лихенизированы Базидиомицеты – 0, 3% всех базидиомицетов (ок. 50 видов) лихенизированы Дейтеромицеты – 1, 2% (ок. 200 видов) несовершенных грибов лихенизированы
ЛИШАЙНИКОВЫЕ МИКОБИОНТЫ около 21% от всех грибов Лишайники – около 15 000 видов. Большинство – дискомицеты, но есть – пиреномицеты и даже базидиомицеты. Аскомицеты – 98% всех лишайников. 40% всех аскомицетов лихенизированы Базидиомицеты – 0, 3% всех базидиомицетов (ок. 50 видов) лихенизированы Дейтеромицеты – 1, 2% (ок. 200 видов) несовершенных грибов лихенизированы
 ЛИШАЙНИКОВЫЕ ФОТОБИОНТЫ Водоросли – фотобионт – менее 200 видов, т. е. 1 фотобионт может формировать ассоциации с многими микобионтами, но более 40% лишайниковых микобионтов ассоциированы с водорослями рода Trebouxia. Отдел Charophyta – 1 род; Отдел Chlorophyta (главным образом, класс Trbouxiophycea) – 10 родов; Класс Xanthophycea – 1 вид; Класс Phaeophycea – 1 вид; Цианобактерии – 13 родов, 4 порядка. Возможны двойные симбиозы. У Peltigera первичый симбионт – зеленая водоросль Coccomyxa, а вторичный (в цефалодиях) – цианобактерия Nostoc.
ЛИШАЙНИКОВЫЕ ФОТОБИОНТЫ Водоросли – фотобионт – менее 200 видов, т. е. 1 фотобионт может формировать ассоциации с многими микобионтами, но более 40% лишайниковых микобионтов ассоциированы с водорослями рода Trebouxia. Отдел Charophyta – 1 род; Отдел Chlorophyta (главным образом, класс Trbouxiophycea) – 10 родов; Класс Xanthophycea – 1 вид; Класс Phaeophycea – 1 вид; Цианобактерии – 13 родов, 4 порядка. Возможны двойные симбиозы. У Peltigera первичый симбионт – зеленая водоросль Coccomyxa, а вторичный (в цефалодиях) – цианобактерия Nostoc.
 Анатомия лишайников Гомеомерные, гетеромерные Гетеромерные лишайники: Наружный слой – кортекс, фотобионтный слой – гонидии, сердцевины – медула, ризоиды – ризины. Специализированные гифы: ищущие, охватывающие, двигающие. Морфология лишайников: Накипные – крустозные, Ареолярные – сквамулезные, Листоватые – фолиозные, Кустистые – фрутикозные
Анатомия лишайников Гомеомерные, гетеромерные Гетеромерные лишайники: Наружный слой – кортекс, фотобионтный слой – гонидии, сердцевины – медула, ризоиды – ризины. Специализированные гифы: ищущие, охватывающие, двигающие. Морфология лишайников: Накипные – крустозные, Ареолярные – сквамулезные, Листоватые – фолиозные, Кустистые – фрутикозные
 ФИЗИОЛОГИЯ ЛИШАЙНИКОВ Фотобионт теряет 80 -90% углерода из соединений, созданных в результате фотосинтеза. Они поглощаются грибными гифами и откладываются в виде сахароспиртов, гликогена и трегалозы. Фотобионт теряет жгутиковые стадии и размножается только делением клеток. Содержание хлорофилла на единицу массы лишайника в 4 - 6 раз меньше, чем у высших растений, соответственно менее активен процесс фотосинтеза и очень медленный рост. Лишайниковые вещества – жирные кислоты, лактоны, поликетиды, антрахиноны, фенилпропаноиды.
ФИЗИОЛОГИЯ ЛИШАЙНИКОВ Фотобионт теряет 80 -90% углерода из соединений, созданных в результате фотосинтеза. Они поглощаются грибными гифами и откладываются в виде сахароспиртов, гликогена и трегалозы. Фотобионт теряет жгутиковые стадии и размножается только делением клеток. Содержание хлорофилла на единицу массы лишайника в 4 - 6 раз меньше, чем у высших растений, соответственно менее активен процесс фотосинтеза и очень медленный рост. Лишайниковые вещества – жирные кислоты, лактоны, поликетиды, антрахиноны, фенилпропаноиды.
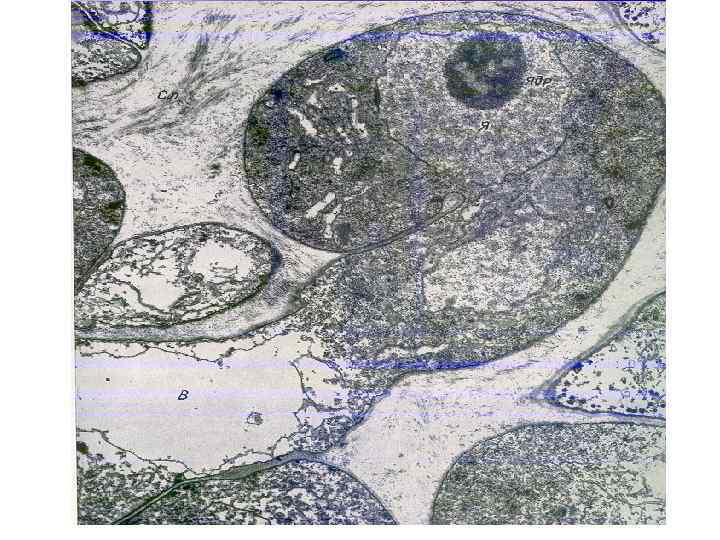
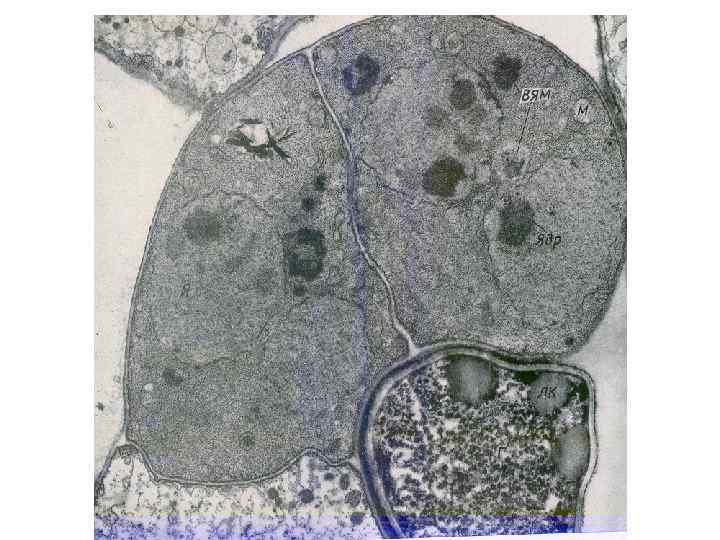
 Взаимодействие мико- и фотобионта 1. Гаустории не образу- ются, поглощение эк- страклеточных выделе- ний фотобионта. 2. Гаустории интракле- точные (стенка водоро- сли с месте контакта разрушена (контакт с плазмалеммой) 3. Гаустории интрапа- риетальные. Стенка во- доросли вдавливается, но не разрушается
Взаимодействие мико- и фотобионта 1. Гаустории не образу- ются, поглощение эк- страклеточных выделе- ний фотобионта. 2. Гаустории интракле- точные (стенка водоро- сли с месте контакта разрушена (контакт с плазмалеммой) 3. Гаустории интрапа- риетальные. Стенка во- доросли вдавливается, но не разрушается
 группы лишайников Эпигейные – на почве Эпифитные – на коре деревьев Эпиксильные") Экологические (топические) группы лишайников Эпигейные – на почве Эпифитные – на коре деревьев Эпиксильные – на обнаженной древесине Эпилитные – на камнях
Экологические (топические) группы лишайников Эпигейные – на почве Эпифитные – на коре деревьев Эпиксильные – на обнаженной древесине Эпилитные – на камнях
 Причины высокой чувствительности лишайников к ксенобиотикам 1. Лишайники – долгожители (индивидуальный таллом может вегетировать 1, 5 -2 тыс. лет). Они подвергаются вредным воздействиям круглый год в течение длительного времени. 2. Медленное отмирание таллома не позволяет освобождаться от токсических соединений в нем. 3. У лишайников отсутствуют кутикула и устьица, они впитывают поллютанты всем телом. 4. При подсыхании таллома концентрация токсических веществ, попавших в него, увеличивается до летальной
Причины высокой чувствительности лишайников к ксенобиотикам 1. Лишайники – долгожители (индивидуальный таллом может вегетировать 1, 5 -2 тыс. лет). Они подвергаются вредным воздействиям круглый год в течение длительного времени. 2. Медленное отмирание таллома не позволяет освобождаться от токсических соединений в нем. 3. У лишайников отсутствуют кутикула и устьица, они впитывают поллютанты всем телом. 4. При подсыхании таллома концентрация токсических веществ, попавших в него, увеличивается до летальной