

Macrophylogeny_lections.ppt
- Количество слайдов: 112
") Класичний період розвитку макро- та мегатаксономії (античні часи – середина ХХ ст. )
Класичний період розвитку макро- та мегатаксономії (античні часи – середина ХХ ст. )
") IV ст. до н. е. : Арістотель – поняття про рослини (ростуть та живляться) та тварини (ростуть, живляться та відчувають)
IV ст. до н. е. : Арістотель – поняття про рослини (ростуть та живляться) та тварини (ростуть, живляться та відчувають)
 -") Теофраст (370 -285 рр. до н. е. ) - "батько" ботаніки Автор двох перших фундаментальних ботанічних праць – "Причини рослин" (шість томів) та "Дослідження рослин" (дев'ять томів). Ввів у наукову літературу термін "mycos" – гриб, та “phycos” – водорість, від яких далі пішли назви двох розділів ботаніки – мікології та фікології. Пліній Старший (23 -70 рр. ) Автор 37 -и томної праці "Природня історія". Запропонував назву "algae" - водорості, за якою розділ ботаніки, що вивчає водорості, отримав іншу синонімічну назву - альгологія (від латинського algae - водорость та грецького logos - наука).
Теофраст (370 -285 рр. до н. е. ) - "батько" ботаніки Автор двох перших фундаментальних ботанічних праць – "Причини рослин" (шість томів) та "Дослідження рослин" (дев'ять томів). Ввів у наукову літературу термін "mycos" – гриб, та “phycos” – водорість, від яких далі пішли назви двох розділів ботаніки – мікології та фікології. Пліній Старший (23 -70 рр. ) Автор 37 -и томної праці "Природня історія". Запропонував назву "algae" - водорості, за якою розділ ботаніки, що вивчає водорості, отримав іншу синонімічну назву - альгологія (від латинського algae - водорость та грецького logos - наука).
 1665: Р. Гук описує клітину (R. Hooke. Micrographia or, Some physiological descriptions of minute bodies made by magnifying glasses. – London, J. Martin & J. Allestry, 1665).
1665: Р. Гук описує клітину (R. Hooke. Micrographia or, Some physiological descriptions of minute bodies made by magnifying glasses. – London, J. Martin & J. Allestry, 1665).
 1676: А. ван Левенгук описує бактерій, перших найпростіших, дріжжі та ін. - т. зв. "анімалькуль" (відкрив він їх de facto 24 квітня 1673 р. , у тритижневому водному настої кореню хрону).
1676: А. ван Левенгук описує бактерій, перших найпростіших, дріжжі та ін. - т. зв. "анімалькуль" (відкрив він їх de facto 24 квітня 1673 р. , у тритижневому водному настої кореню хрону).
 1753: К. Лінней, "Species plantarum". Бінарна номенклатура. 1767: К. Лінней (Linne, 1767) – поняття про царства. Три царства природи – Regnum Vegetabilia (тіла організовані, живі, але не відчувають), Reg. Animalia (тіла організовані, живі, відчувають і довільно рухаються), Reg. Lapides (камені).
1753: К. Лінней, "Species plantarum". Бінарна номенклатура. 1767: К. Лінней (Linne, 1767) – поняття про царства. Три царства природи – Regnum Vegetabilia (тіла організовані, живі, але не відчувають), Reg. Animalia (тіла організовані, живі, відчувають і довільно рухаються), Reg. Lapides (камені).
 1768: Едвард Дженнер проводить першу вакцинацію 1771: Д. Прістлі відкриває природу фотосинтезу 1802: Ламарк (1802) – термін "біологія". Ідея еволюції живої природи.
1768: Едвард Дженнер проводить першу вакцинацію 1771: Д. Прістлі відкриває природу фотосинтезу 1802: Ламарк (1802) – термін "біологія". Ідея еволюції живої природи.
 – Regnum Mycetoideum") 1821: Еліас Фріз (Fries, 1821) – Regnum Mycetoideum
1821: Еліас Фріз (Fries, 1821) – Regnum Mycetoideum
 Дві нотатки щодо внеску Михайла Олександровича Максимовича у ботаніку
Дві нотатки щодо внеску Михайла Олександровича Максимовича у ботаніку
 Період активної діяльності М. О. Максимовича як ботаніка - 1821 -1834 рр. (“Московський” період – студент, викладач, професор кафедри ботаніки Московського університету, завідувач ботанічним садом при Московськом університеті) Перша робота – студентський реферативний огляд "К чему служат листья растений“ (1821) Останні власне ботанічні роботи - "Историческое изложение системы растительного царства“ (1933), "Обозрение органологии растений“ (1933), "Явноцветные и тайноцветные растения (1934)"
Період активної діяльності М. О. Максимовича як ботаніка - 1821 -1834 рр. (“Московський” період – студент, викладач, професор кафедри ботаніки Московського університету, завідувач ботанічним садом при Московськом університеті) Перша робота – студентський реферативний огляд "К чему служат листья растений“ (1821) Останні власне ботанічні роботи - "Историческое изложение системы растительного царства“ (1933), "Обозрение органологии растений“ (1933), "Явноцветные и тайноцветные растения (1934)"
 Розробка національної ботанічної термінології Національні еквіваленти латинських назв рослин Розробка вітчизняної ботанічної термінології У галузі флористики М. О. Максимович виконав біля десяти робіт, з яких найголовнішими були дослідження, присвячені інвентаризації флори Курської та Московської губерній, інвентаризації флори рослин закритого грунту Московського університетського ботанічного саду та флористичне зведення, присвячене видам роду Galium (Підмаренник) Центральної Росії. Флористичні дослідження, виконані Максимовичем, припадають на початковий період інвентаризації флори Російської імперії. Майже всі нечисленні російські флористичні роботи цього часу видруковані латиною, і не містять ані національних назв рослин, ані російських ботанічних термінів.
Розробка національної ботанічної термінології Національні еквіваленти латинських назв рослин Розробка вітчизняної ботанічної термінології У галузі флористики М. О. Максимович виконав біля десяти робіт, з яких найголовнішими були дослідження, присвячені інвентаризації флори Курської та Московської губерній, інвентаризації флори рослин закритого грунту Московського університетського ботанічного саду та флористичне зведення, присвячене видам роду Galium (Підмаренник) Центральної Росії. Флористичні дослідження, виконані Максимовичем, припадають на початковий період інвентаризації флори Російської імперії. Майже всі нечисленні російські флористичні роботи цього часу видруковані латиною, і не містять ані національних назв рослин, ані російських ботанічних термінів.
 Національні еквіваленти латинських назв рослин "Список растений московской флоры" (1826) "Каталог диких растений в Дмитриевском уезде Курской губернии" (1826) “Словарь родовых названий растений”, составленный И. Мартыновым (1826) “Разсуждение о семействе крестовидных растений, Декандоля, переведенное с Французскаго П. Троцким, с фигурами” (1826) "Список многолетних оранжерейных и тепличных растений ботанического сада при Московском университете (1828)"
Національні еквіваленти латинських назв рослин "Список растений московской флоры" (1826) "Каталог диких растений в Дмитриевском уезде Курской губернии" (1826) “Словарь родовых названий растений”, составленный И. Мартыновым (1826) “Разсуждение о семействе крестовидных растений, Декандоля, переведенное с Французскаго П. Троцким, с фигурами” (1826) "Список многолетних оранжерейных и тепличных растений ботанического сада при Московском университете (1828)"
 "Некоторые родовые имена заимствованы мною из народных, мною собранных на месте в губерниях Московской и Черниговской“ (Максимович, 1926) Iris – Косатик, Phleum – Аржанец, Asperugo – Червенница, Vinca – Барвинок, Selinum – Гирча, Conioselinum – Гирчовник, Ligusticum – Зоря, Luzula – Ожика, Veratrum – Чемерица, Alisma – Частуха, Dianthus – Травянец, Sedum – Очиток, Sempervivium – Молодило, Prunella – Горлянка, Draba – Сухоребрица, Bunias – Свербига, Scorzonera – Козелец, Serratula – Серпуха, Bidens – Череда, Artemisisa – Нехворощ, Conyza – Богатинка
"Некоторые родовые имена заимствованы мною из народных, мною собранных на месте в губерниях Московской и Черниговской“ (Максимович, 1926) Iris – Косатик, Phleum – Аржанец, Asperugo – Червенница, Vinca – Барвинок, Selinum – Гирча, Conioselinum – Гирчовник, Ligusticum – Зоря, Luzula – Ожика, Veratrum – Чемерица, Alisma – Частуха, Dianthus – Травянец, Sedum – Очиток, Sempervivium – Молодило, Prunella – Горлянка, Draba – Сухоребрица, Bunias – Свербига, Scorzonera – Козелец, Serratula – Серпуха, Bidens – Череда, Artemisisa – Нехворощ, Conyza – Богатинка
 Рецензія на: “Словарь родовых имен растений, с переводом на российский язык. . . , составленный Иваном Мартыновым” (1826)
Рецензія на: “Словарь родовых имен растений, с переводом на российский язык. . . , составленный Иваном Мартыновым” (1826)

 Розробка вітчизняної ботанічної термінології "О русских подмаренниках" (1829) “Систематика растений” (1831) "Обозрение органологии растений“ (1933) "Очерк растительного царства“ (1933, рукопис)
Розробка вітчизняної ботанічної термінології "О русских подмаренниках" (1829) “Систематика растений” (1831) "Обозрение органологии растений“ (1933) "Очерк растительного царства“ (1933, рукопис)

 Оформлення та визначення ботанічних термінів здійснено в монографіях наукового та навчальнометодичного спрямування “Систематика растений” (1831) "Обозрение органологии растений“ (1933) Деякі приклади ботанічних термінів, що закріпилися в російській мові: “венчик”, “лепестки”, “пестик”, “тычинки”, “завязь”, “зародыш”, “столбик”, “рыльце”, “однодольные”, “двудольные” Деякі терміни, запропоновані М. О. Максимовичем, пізніше були дещо видозмінені (наприклад, “колоцвет” – на “околоцвет” та далі – на “околоцветник”) Частина термінів була надалі перекладена українською мовою зі збереженням коренів, запропонованих М. О. Максимовичем (наприклад, зав’язь, зародок, стовпчик, оцвітина)
Оформлення та визначення ботанічних термінів здійснено в монографіях наукового та навчальнометодичного спрямування “Систематика растений” (1831) "Обозрение органологии растений“ (1933) Деякі приклади ботанічних термінів, що закріпилися в російській мові: “венчик”, “лепестки”, “пестик”, “тычинки”, “завязь”, “зародыш”, “столбик”, “рыльце”, “однодольные”, “двудольные” Деякі терміни, запропоновані М. О. Максимовичем, пізніше були дещо видозмінені (наприклад, “колоцвет” – на “околоцвет” та далі – на “околоцветник”) Частина термінів була надалі перекладена українською мовою зі збереженням коренів, запропонованих М. О. Максимовичем (наприклад, зав’язь, зародок, стовпчик, оцвітина)
 У витоків вітчизняної науки про водорості "На території Російської імперії до кінця ХVIII ст. водорості називали поростами. На початку ХІХ ст. їх стали називати водораслями. У 30 -х роках ХІХ ст. Михайло Максимович запропонував назву "водоросли", яка закріпилась у російській мові, і була надалі перекладена українською як "водорості" (з конспекту вступної лекції до спецкурсу “Альгологія”, прочитаної завдувачем кафедри нижчих рослин Київського державного університету ім. Т. Г. Шевченка, професором Н. П. Масюк, 1980 р. )
У витоків вітчизняної науки про водорості "На території Російської імперії до кінця ХVIII ст. водорості називали поростами. На початку ХІХ ст. їх стали називати водораслями. У 30 -х роках ХІХ ст. Михайло Максимович запропонував назву "водоросли", яка закріпилась у російській мові, і була надалі перекладена українською як "водорості" (з конспекту вступної лекції до спецкурсу “Альгологія”, прочитаної завдувачем кафедри нижчих рослин Київського державного університету ім. Т. Г. Шевченка, професором Н. П. Масюк, 1980 р. )
 До кінця XVIII ст. водорості лишаються найбільш незрозумілою, найбільш невивченою групою рослин - Ruda et Confusa (грубі та незрозумілі) 1703 р. – Британським Королівським товариством опубліковано перше зображення мікроскопічних водоростей (робота анонімного містера С. , в якій наведено рисунки одноклітинних діатомових водоростей з поверхні кореня вищої водної рослини ряски). Деякі діатомові водорості (латинська назва – Diatomeae – запропонована шведським ботаніком Фрізом у 1821 р. )
До кінця XVIII ст. водорості лишаються найбільш незрозумілою, найбільш невивченою групою рослин - Ruda et Confusa (грубі та незрозумілі) 1703 р. – Британським Королівським товариством опубліковано перше зображення мікроскопічних водоростей (робота анонімного містера С. , в якій наведено рисунки одноклітинних діатомових водоростей з поверхні кореня вищої водної рослини ряски). Деякі діатомові водорості (латинська назва – Diatomeae – запропонована шведським ботаніком Фрізом у 1821 р. )
 Карл Лінней у XVIII ст. об'єднав водорості та гриби разом з лишайниками, мохоподібними та папоротеподібними в окремий 24 -й клас системи царства рослин – Cryptogamen (Таємношлюбні). Проте природа водоростей для Ліннея залишилась незрозумілою. Наприклад, Лінней стверджує, що "Всі види рослин мають квітку і плід, навіть коли око їх не вловлює. . . Квітки фукусів (Fuci) спостерігав Реомюр. " У "Видах рослин" (Species plantarum, 1753) він наводить лише чотири роди водоростей - Fucus, Ulva, Conferva та Bussus (два останніх роди є збірними та у ХІХ ст. закриті).
Карл Лінней у XVIII ст. об'єднав водорості та гриби разом з лишайниками, мохоподібними та папоротеподібними в окремий 24 -й клас системи царства рослин – Cryptogamen (Таємношлюбні). Проте природа водоростей для Ліннея залишилась незрозумілою. Наприклад, Лінней стверджує, що "Всі види рослин мають квітку і плід, навіть коли око їх не вловлює. . . Квітки фукусів (Fuci) спостерігав Реомюр. " У "Видах рослин" (Species plantarum, 1753) він наводить лише чотири роди водоростей - Fucus, Ulva, Conferva та Bussus (два останніх роди є збірними та у ХІХ ст. закриті).
 Датою народження альгології вважають 1801 р. , коли Дж. Стакхаус описав процес запліднення та проростання зиготи у бурої морської водорості Fucus. Дослідження Стакхауса спростували твердження К. Ліннея про наявіність у водоростей квіток та плодів і показали необхідність побудови системи водоростей на інших, не ліннеївських, засадах. Але якими мають бути ці засади, на початку ХІХ ст. було зовсім незрозуміло. Морська бура водорість Fucus, на прикладі якої Дж. Стакхаусом було вперше доведено, що водорості не утворюють квіток
Датою народження альгології вважають 1801 р. , коли Дж. Стакхаус описав процес запліднення та проростання зиготи у бурої морської водорості Fucus. Дослідження Стакхауса спростували твердження К. Ліннея про наявіність у водоростей квіток та плодів і показали необхідність побудови системи водоростей на інших, не ліннеївських, засадах. Але якими мають бути ці засади, на початку ХІХ ст. було зовсім незрозуміло. Морська бура водорість Fucus, на прикладі якої Дж. Стакхаусом було вперше доведено, що водорості не утворюють квіток
: “Жизнь Московского") Роботи М. О. Максимовича, присвячені нижчим рослинам (водоростям та грибам, включаючи лишайники): “Жизнь Московского Профессора Ботаники Гофмана (1826); "Обозрение поростов, мхов и папоротников, ч. 1" (1827); "Обозрение поростов, мхов и папоротников, ч. 2" (1828); "Общие сравнения грибов и поростов" (1828); “О разделении Поростов на порядки” (1928); "О диатомеях" (1828); "Обозрение водорослей" (1828); “Письмо шведскому ученому Фрису относительно системы Линнея” (1833)
Роботи М. О. Максимовича, присвячені нижчим рослинам (водоростям та грибам, включаючи лишайники): “Жизнь Московского Профессора Ботаники Гофмана (1826); "Обозрение поростов, мхов и папоротников, ч. 1" (1827); "Обозрение поростов, мхов и папоротников, ч. 2" (1828); "Общие сравнения грибов и поростов" (1828); “О разделении Поростов на порядки” (1928); "О диатомеях" (1828); "Обозрение водорослей" (1828); “Письмо шведскому ученому Фрису относительно системы Линнея” (1833)
 Фрагмент неопублікованої системи Г. Ф. Гофмана 1826 р. у частині, що стосується водоростей (за Максимович, 1826)
Фрагмент неопублікованої системи Г. Ф. Гофмана 1826 р. у частині, що стосується водоростей (за Максимович, 1826)
 «Царство растительное начинается Поростами и оканчивается Грибами. . . Поросты и Грибы составляют два класса, ближайшие друг к другу по своему несовершенству, но отдаленнейшие по времени: это так сказать альфа и омега всего царства»
«Царство растительное начинается Поростами и оканчивается Грибами. . . Поросты и Грибы составляют два класса, ближайшие друг к другу по своему несовершенству, но отдаленнейшие по времени: это так сказать альфа и омега всего царства»
 Визначення принципів вибору національних назв водоростей: “Я старался Латинския названия, если только могли оне оставаться без перевода на Русском языке. Если не льзя было оставить Латинскаго имени, то я старался перевод оного заимствовать от слов Русских, иногда Малороссийских, наблюдая, чтобы имя было сколько можно: 1) короче и благозвучнее, 2) что бы оно выражало качество им означаемого предмета, 3) что бы от него можно было произвести удобно имя прилагательное, ибо сие необходимо в Ботанике”.
Визначення принципів вибору національних назв водоростей: “Я старался Латинския названия, если только могли оне оставаться без перевода на Русском языке. Если не льзя было оставить Латинскаго имени, то я старался перевод оного заимствовать от слов Русских, иногда Малороссийских, наблюдая, чтобы имя было сколько можно: 1) короче и благозвучнее, 2) что бы оно выражало качество им означаемого предмета, 3) что бы от него можно было произвести удобно имя прилагательное, ибо сие необходимо в Ботанике”.
 Корковые водоросли") Система водоростей М. О. Максимовича I. Поросты водяные: 1. Водоросли (Hydrophyceae Fries) Корковые водоросли Фукусовые (Fucoideae) (10 родов) Цветистые (Florideae) (16 родов) Серпянковые (Ulvea) (4 рода) Нитчатки (Conferveae) (3 рода) Тинничные (Zygnemeae) (4 рода) Водоросли жаборинные 2. Діатомеї (Diatomeae Fries) Ундины (напр. Nostoc) Маяльные (напр. Oscillatoria) Фрагилярийные (Fragilarinae) Ехинеллевые (Echinelleae) II. Поросты воздушные: 3. Лишаи (Lichenes) 4. Жилочниковые (Byssaceae)
Система водоростей М. О. Максимовича I. Поросты водяные: 1. Водоросли (Hydrophyceae Fries) Корковые водоросли Фукусовые (Fucoideae) (10 родов) Цветистые (Florideae) (16 родов) Серпянковые (Ulvea) (4 рода) Нитчатки (Conferveae) (3 рода) Тинничные (Zygnemeae) (4 рода) Водоросли жаборинные 2. Діатомеї (Diatomeae Fries) Ундины (напр. Nostoc) Маяльные (напр. Oscillatoria) Фрагилярийные (Fragilarinae) Ехинеллевые (Echinelleae) II. Поросты воздушные: 3. Лишаи (Lichenes) 4. Жилочниковые (Byssaceae)
 «Истинное принадлежит природе, и, будучи открыто умом, остается всегда достоянием Системы. Ошибочное принадлежит своему автору и времени, и остается только, как поучительный памятник исторический» (М. А. Максимович. «Очерк растительного царства» (1833))
«Истинное принадлежит природе, и, будучи открыто умом, остается всегда достоянием Системы. Ошибочное принадлежит своему автору и времени, и остается только, как поучительный памятник исторический» (М. А. Максимович. «Очерк растительного царства» (1833))
 1828: Х. Еренберг пропонує термін "бактерії“ 1831: Р. Б. Браун описує ядро
1828: Х. Еренберг пропонує термін "бактерії“ 1831: Р. Б. Браун описує ядро
 1838: ботанік Маттіас Шлейден та зоолог Теодор Шванн формулюють основні положення клітинної теорії
1838: ботанік Маттіас Шлейден та зоолог Теодор Шванн формулюють основні положення клітинної теорії
 демонструє відсутність морфологічно оформленого ядра у бактерій та синьозелених водоростей,") 1853: Кон (Cohn, 1853) демонструє відсутність морфологічно оформленого ядра у бактерій та синьозелених водоростей, і у 1875 р. (Cohn, 1875) об’єднує бактерії та синьозелені водорості в єдиний відділ – дроб’янки (Schizophyta).
1853: Кон (Cohn, 1853) демонструє відсутність морфологічно оформленого ядра у бактерій та синьозелених водоростей, і у 1875 р. (Cohn, 1875) об’єднує бактерії та синьозелені водорості в єдиний відділ – дроб’янки (Schizophyta).
 1859: Дж. Гукер представляє Лінеївському товариству у Лондоні роботи Ч. Дарвіна та А. Р. Уоллеса, що містять виклад теорії природнього добору. У цьому ж році виходить друком "Походження видів. . . " Ч. Дарвіна.
1859: Дж. Гукер представляє Лінеївському товариству у Лондоні роботи Ч. Дарвіна та А. Р. Уоллеса, що містять виклад теорії природнього добору. У цьому ж році виходить друком "Походження видів. . . " Ч. Дарвіна.
. 1865:") 1862: Л. Пастер доводить неможливість самозародження життя, а також встановлює природу бродіння (1861). 1865: Г. Мендель формулює три закони спадковості.
1862: Л. Пастер доводить неможливість самозародження життя, а також встановлює природу бродіння (1861). 1865: Г. Мендель формулює три закони спадковості.
 – Reg. Protista (всі одноклітинні,") Ернст Геккель – “батько” протозоології 1866: Геккель (Haeckel, 1866) – Reg. Protista (всі одноклітинні, міксоміцети, губки), Reg. Plantae, Reg. Animalia.
Ернст Геккель – “батько” протозоології 1866: Геккель (Haeckel, 1866) – Reg. Protista (всі одноклітинні, міксоміцети, губки), Reg. Plantae, Reg. Animalia.
 Виділяються три царства Plantae, Animalia, Protista (проміжне між") Система трьох царств “раннього” Геккеля (1866) Виділяються три царства Plantae, Animalia, Protista (проміжне між рослинами та тваринами), а також “вихідний стовбур” – Radix Moneres. Перша сліпа гілка в Protista названа Moneres. Plantae Fungi Animalia Protista Radix Monera 1878: У системы "середнього" Геккеля (Haeckel, 1878) окремою гілкою від протистів виведено гриби
Система трьох царств “раннього” Геккеля (1866) Виділяються три царства Plantae, Animalia, Protista (проміжне між рослинами та тваринами), а також “вихідний стовбур” – Radix Moneres. Перша сліпа гілка в Protista названа Moneres. Plantae Fungi Animalia Protista Radix Monera 1878: У системы "середнього" Геккеля (Haeckel, 1878) окремою гілкою від протистів виведено гриби
 та її модифікація (система Уіттекера-Маргеліс, 1981) ілюстують прислів’я «нове") Найпопулярніша “західна” мегасистема Уіттекера (1969) та її модифікація (система Уіттекера-Маргеліс, 1981) ілюстують прислів’я «нове – це добре забуте старе» Plantae Fungi Animalia Protista Radix Monera Система "середнього" Геккеля (Haeckel, 1878) Система Уіттекера (1969)
Найпопулярніша “західна” мегасистема Уіттекера (1969) та її модифікація (система Уіттекера-Маргеліс, 1981) ілюстують прислів’я «нове – це добре забуте старе» Plantae Fungi Animalia Protista Radix Monera Система "середнього" Геккеля (Haeckel, 1878) Система Уіттекера (1969)
 1892: Дмитро Йосипович Івановський відкриває віруси Мартін Бейерінк підтверджує відкриття Д. Й. Івановського та пропонує термін “вірус”
1892: Дмитро Йосипович Івановський відкриває віруси Мартін Бейерінк підтверджує відкриття Д. Й. Івановського та пропонує термін “вірус”
 Ідеї Адольфа Пашера 1914: Ідеї Пашера: поліфілія, первинність монадної організації, теорія морфологічного паралелізму та типів структур тіла водоростей, розділення філ рослин за забарвленням. А. Пашер (Pascher, 1914) висуває ідею первинності монадних організмів та розвиває концепцію відмін основних філ водоростей за забавленням. На основі постулату про первинність монадної організації розробляє концепцію типів морфологічних структур тіла водоростей. Застосовує цю концепцію для розробки ідеї морфологічного паралелізму з метою розробки принципів класифікації водоростей на рівні середніх таксонів (класів та порядків).
Ідеї Адольфа Пашера 1914: Ідеї Пашера: поліфілія, первинність монадної організації, теорія морфологічного паралелізму та типів структур тіла водоростей, розділення філ рослин за забарвленням. А. Пашер (Pascher, 1914) висуває ідею первинності монадних організмів та розвиває концепцію відмін основних філ водоростей за забавленням. На основі постулату про первинність монадної організації розробляє концепцію типів морфологічних структур тіла водоростей. Застосовує цю концепцію для розробки ідеї морфологічного паралелізму з метою розробки принципів класифікації водоростей на рівні середніх таксонів (класів та порядків).
 Вперше органічний світ поділяється на царства на основі цитологічних") Схема класифікації найпростіших Шаттона (1925) Вперше органічний світ поділяється на царства на основі цитологічних критеріїв. Вперше пропонуються терміни “прокаріоти” та “еукаріоти”
Схема класифікації найпростіших Шаттона (1925) Вперше органічний світ поділяється на царства на основі цитологічних критеріїв. Вперше пропонуються терміни “прокаріоти” та “еукаріоти”
 поглиблює концепцію прокаріот та евкаріот, і на її") 1938: Коупленд (Copeland, 1938, 1947, 1956) поглиблює концепцію прокаріот та евкаріот, і на її основі пропонує систему органічного світу з чотирьох царств: Monera (всі прокаріоти), Protoctista (всі одноклітинні, гриби, слизовики, червоні, бурі, харові водорості), Plantae (зелені водорості та всі вищі рослини), Animalia (всі Metazoa, крім губок)
1938: Коупленд (Copeland, 1938, 1947, 1956) поглиблює концепцію прокаріот та евкаріот, і на її основі пропонує систему органічного світу з чотирьох царств: Monera (всі прокаріоти), Protoctista (всі одноклітинні, гриби, слизовики, червоні, бурі, харові водорості), Plantae (зелені водорості та всі вищі рослини), Animalia (всі Metazoa, крім губок)
") Ендосимбіогенез 1870: Андрій Сергійович Фамінцин – вперше висловлює здогадку про ендосимбіотичну природу клітини (1870)
Ендосимбіогенез 1870: Андрій Сергійович Фамінцин – вперше висловлює здогадку про ендосимбіотичну природу клітини (1870)
: можливо, зелені рослини є наслідком об’єднання безбарвних") 1883: Шимпер Андреас Франс Вільгельм (Schimper, 1883): можливо, зелені рослини є наслідком об’єднання безбарвних організмів з організмами, забарвленими у зелений колір. Розмежував хлоропласти (у рослин) та хроматофори (симбіотичні утвори у морських тварин)
1883: Шимпер Андреас Франс Вільгельм (Schimper, 1883): можливо, зелені рослини є наслідком об’єднання безбарвних організмів з організмами, забарвленими у зелений колір. Розмежував хлоропласти (у рослин) та хроматофори (симбіотичні утвори у морських тварин)
: походження хлоропластів та ядра внаслідок симбіозів різних організмів.") 1896: Михайло Семенович Цвєт (Tswett, 1896): походження хлоропластів та ядра внаслідок симбіозів різних організмів. Спостереження поділу хлоропласту та ядерця у Spirogyra.
1896: Михайло Семенович Цвєт (Tswett, 1896): походження хлоропластів та ядра внаслідок симбіозів різних організмів. Спостереження поділу хлоропласту та ядерця у Spirogyra.
 Костянтин Сергійович Мережковський (1825 -1921)") Теорія двох плазм (1905) Костянтин Сергійович Мережковський (1825 -1921)
Теорія двох плазм (1905) Костянтин Сергійович Мережковський (1825 -1921)
 Царство Мікоїдов (без симбіогенеза) бактерії,") Теорія двох плазм К. С. Мережковського (1905, 1909, 1910) Царство Мікоїдов (без симбіогенеза) бактерії, гриби, синьозелені водорості, а також “хроматофори” та “хроміоли ядра” Царство Тварин (перший симбіоз – утворення ядра) рослини всі тварини та слизовики тварини гриби “хроматофори” “хроміоли ядра” бактерії Мікоплазма Амебоплазма Царство Рослин (перший та другий симбіози – утворення ядра та хлоропластів) всі водорості (за винятком синьозелених), вищі рослини та фікоміцети (зокрема, сучасні Oomycota)
Теорія двох плазм К. С. Мережковського (1905, 1909, 1910) Царство Мікоїдов (без симбіогенеза) бактерії, гриби, синьозелені водорості, а також “хроматофори” та “хроміоли ядра” Царство Тварин (перший симбіоз – утворення ядра) рослини всі тварини та слизовики тварини гриби “хроматофори” “хроміоли ядра” бактерії Мікоплазма Амебоплазма Царство Рослин (перший та другий симбіози – утворення ядра та хлоропластів) всі водорості (за винятком синьозелених), вищі рослини та фікоміцети (зокрема, сучасні Oomycota)
 І. Царство Мікоїдів (симбіоз відсутній) Вільноіснуючі 1. Бактерії (Bacteria)") Система К. С. Мережковського (1910) І. Царство Мікоїдів (симбіоз відсутній) Вільноіснуючі 1. Бактерії (Bacteria) 2. Гриби (Fungi) 3. Ціанофіцеї (Cyanophyceae) Симбіонти 1. Хроматофори 2. Хроміоли ядра 2. Царство Рослин (подвійний симбіоз) 1. Algophyta Algae Leucophyta (Phycomycetes) 2. Bryophyta 3. Pteridophyta 4. Spermatophyta Angiospermae Gymnospermae 3. Царство Тварин (простий симбіоз)
Система К. С. Мережковського (1910) І. Царство Мікоїдів (симбіоз відсутній) Вільноіснуючі 1. Бактерії (Bacteria) 2. Гриби (Fungi) 3. Ціанофіцеї (Cyanophyceae) Симбіонти 1. Хроматофори 2. Хроміоли ядра 2. Царство Рослин (подвійний симбіоз) 1. Algophyta Algae Leucophyta (Phycomycetes) 2. Bryophyta 3. Pteridophyta 4. Spermatophyta Angiospermae Gymnospermae 3. Царство Тварин (простий симбіоз)
 1970: теорія ендосимбіогенезу Л. Маргеліс. Виходить друком перша книга Л. Маргеліс, де викладено основи сучасної ендосимбіотичної теорії (L. Margulis. Origin of Eukaryotic Cells. – Yale Univ. Press, 1970)
1970: теорія ендосимбіогенезу Л. Маргеліс. Виходить друком перша книга Л. Маргеліс, де викладено основи сучасної ендосимбіотичної теорії (L. Margulis. Origin of Eukaryotic Cells. – Yale Univ. Press, 1970)
 Виникнення евкаріот та їх дивергенція згідно з синтетичною гіпотезою. Суцільними стрілками позначені напрямки еволюції клітини, штриховими стрілками – ендосимбіози, штрих-пунктирною стрілкою – горизонатальний перенос гену, що кодує тубулін, внаслідок чого виникли мікротрубочкові клітинні системи. І – автогенетичне утворення ядра та одномембранних евкаріотичних органел; ІІ – виникнення мікротрубочкових систем, джгутикового апарату, мітозу, мейозу, статевого процесу; ІІІ – ендосимбіоз, вналідок якого виникли мітохондрії з дископодібними кристами; IV – виникнення платикристат та тубулокристат; V – ендосимбіоз, внаслідок якого виникла первинна пластида; VI – велика радіація евкаріот та виникнення в межах платикристат рослин, грибів та тварин, а в межах тубулокристат – амебо-флагелят, альвеолят та страменопілів.
Виникнення евкаріот та їх дивергенція згідно з синтетичною гіпотезою. Суцільними стрілками позначені напрямки еволюції клітини, штриховими стрілками – ендосимбіози, штрих-пунктирною стрілкою – горизонатальний перенос гену, що кодує тубулін, внаслідок чого виникли мікротрубочкові клітинні системи. І – автогенетичне утворення ядра та одномембранних евкаріотичних органел; ІІ – виникнення мікротрубочкових систем, джгутикового апарату, мітозу, мейозу, статевого процесу; ІІІ – ендосимбіоз, вналідок якого виникли мітохондрії з дископодібними кристами; IV – виникнення платикристат та тубулокристат; V – ендосимбіоз, внаслідок якого виникла первинна пластида; VI – велика радіація евкаріот та виникнення в межах платикристат рослин, грибів та тварин, а в межах тубулокристат – амебо-флагелят, альвеолят та страменопілів.
") Цитологічний період у макро- та мегатаксономії (50 – 90 -і роки ХХ ст. )
Цитологічний період у макро- та мегатаксономії (50 – 90 -і роки ХХ ст. )
 1931: Е. Руска з колегами конструює перший прототип електронного мікроскопу 1939: корпорація Siemens створює перший комерційний електронний мікроскоп.
1931: Е. Руска з колегами конструює перший прототип електронного мікроскопу 1939: корпорація Siemens створює перший комерційний електронний мікроскоп.
 1951: І. Мантон починає перші дослідження біологічних об’єктів під електронним мікроскопом
1951: І. Мантон починає перші дослідження біологічних об’єктів під електронним мікроскопом
, спираючись на дані ультраструктурних досліджень, формулює гіпотезу автогенетичного походження евкаріотичної клітини.") 1969: Алсопп (Allsopp), спираючись на дані ультраструктурних досліджень, формулює гіпотезу автогенетичного походження евкаріотичної клітини. Надалі цю гіпотезу розробляє Шадефо (Chadefaud, 1974)
1969: Алсопп (Allsopp), спираючись на дані ультраструктурних досліджень, формулює гіпотезу автогенетичного походження евкаріотичної клітини. Надалі цю гіпотезу розробляє Шадефо (Chadefaud, 1974)
") А. Массальський та Г. Лідаль відкривають ретронеми у золотистих та жовтозелених водоростей (1969)
А. Массальський та Г. Лідаль відкривають ретронеми у золотистих та жовтозелених водоростей (1969)
 Ретронеми ультратонки волоски, що розташовані на поверхні джгутиків. Мають складну будову і утворюються внаслідок взаємодії структур, похідних від ядерної мембраною, CER, комплексу Гольдж та плазмалеми термінальні волоски мікротубулярний сегмент базальна частина аксонема джгутика Схема будови ретронеми Ретронеми утворюються у перинуклеарному просторі - між зовнішньою та внутрішньою мембранами ядерної оболонки
Ретронеми ультратонки волоски, що розташовані на поверхні джгутиків. Мають складну будову і утворюються внаслідок взаємодії структур, похідних від ядерної мембраною, CER, комплексу Гольдж та плазмалеми термінальні волоски мікротубулярний сегмент базальна частина аксонема джгутика Схема будови ретронеми Ретронеми утворюються у перинуклеарному просторі - між зовнішньою та внутрішньою мембранами ядерної оболонки
") Eucaryota Procaryota Acytobionta Схема Д. К. Зерова (1972)
Eucaryota Procaryota Acytobionta Схема Д. К. Зерова (1972)

") Система 18 -и евкаріотичних царств Гордона Лідаля (Leedale, 1974)
Система 18 -и евкаріотичних царств Гордона Лідаля (Leedale, 1974)
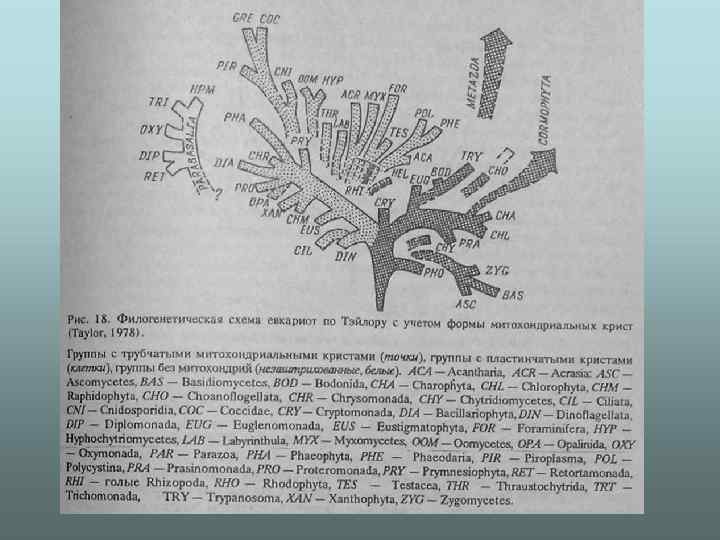
") Система Уіттекера-Маргеліс (по: Маргелис, 1981)
Система Уіттекера-Маргеліс (по: Маргелис, 1981)
 The kingdom Chromista: origin and systematics. In: Round FE, Chapman DJ") Cavalier-Smith T (1986) The kingdom Chromista: origin and systematics. In: Round FE, Chapman DJ (eds) Progress in phycological research. Biopress, Bristol, pp 309– 347 Кавальє-Сміс (1986) та його царство Chromista
Cavalier-Smith T (1986) The kingdom Chromista: origin and systematics. In: Round FE, Chapman DJ (eds) Progress in phycological research. Biopress, Bristol, pp 309– 347 Кавальє-Сміс (1986) та його царство Chromista
 До традиційних царств системи Геккеля-Уіттекера (Protista,") Система п’яти царств евкаріот Т. Кавальє-Сміса (Cavalier-Smith, 1986) До традиційних царств системи Геккеля-Уіттекера (Protista, Plantae, Fungi, Animalia) додано п’яте царство – Chromista. Тварини та гриби виводяться від спільного пращура і утворюють єдину кладу, яку Кавальє-Сміс пізніше назве царством Opistokontae.
Система п’яти царств евкаріот Т. Кавальє-Сміса (Cavalier-Smith, 1986) До традиційних царств системи Геккеля-Уіттекера (Protista, Plantae, Fungi, Animalia) додано п’яте царство – Chromista. Тварини та гриби виводяться від спільного пращура і утворюють єдину кладу, яку Кавальє-Сміс пізніше назве царством Opistokontae.
") Альвеоляти Тубулокристати Страменопіли Платикристати Дискокристати Домітохондріальні евкаріоти Cтруктурне дерево евкаріот (Patterson, 1994)
Альвеоляти Тубулокристати Страменопіли Платикристати Дискокристати Домітохондріальні евкаріоти Cтруктурне дерево евкаріот (Patterson, 1994)
 Багатоклітинні тварини Деякі найпростіші Тварини Rhodophyta Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Glaucocystophyta Eustigmatophyta Вищі рослини Chlorophyta Basidiomycota Haptophyta Стаменопіли Chytridiomycota Справжні гриби Апікомплекси Альвеоляти Oomycota Деякі найпростіші Рослини Zygomycota Dinophyta Chrysophyta Raphidophyta Cryptophyta Ascomycota Інфузорії Амебоїдні найпростіші Chlorarachniopyta Plasmodiophoromycota Myxomycota Амебо-флагеляти Платикристати Евкаріоти Euglenophyta Тубулокристати Кінетопластиди Дискокристати Acrasiomycota Деякі найпростіші Гіпохондріати Прокаріоти Cyanophyta Архебактерії Евбактерії
Багатоклітинні тварини Деякі найпростіші Тварини Rhodophyta Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Glaucocystophyta Eustigmatophyta Вищі рослини Chlorophyta Basidiomycota Haptophyta Стаменопіли Chytridiomycota Справжні гриби Апікомплекси Альвеоляти Oomycota Деякі найпростіші Рослини Zygomycota Dinophyta Chrysophyta Raphidophyta Cryptophyta Ascomycota Інфузорії Амебоїдні найпростіші Chlorarachniopyta Plasmodiophoromycota Myxomycota Амебо-флагеляти Платикристати Евкаріоти Euglenophyta Тубулокристати Кінетопластиди Дискокристати Acrasiomycota Деякі найпростіші Гіпохондріати Прокаріоти Cyanophyta Архебактерії Евбактерії
 Макро- та мегатаксономія на сучасному етапі (90 -і роки ХХ ст. – початок ХХІ ст. )
Макро- та мегатаксономія на сучасному етапі (90 -і роки ХХ ст. – початок ХХІ ст. )
 Eubacteria Firmacutes Cyanophyta пластиди Eukaryota мітохондрії бактероїди хламідії протеобактерії еукаріоти зелені сірчані бактерії спірохети пурпурні бактерії Archaebacteria зелені несірчані бактерії Gracillicutes Mendosicutes Tenericutes прокаріоти гідролітичні екзоферменти наявні утилізують складні органічні сполуки мешкають в біотопах з широкою екол. амплітудою здатні до азотфіксації Progenota ліпіди гліцерол-фосфатні прокаріоти відсутні гідролітичні екзоферменти не засвоюють складні органічні сполуки здатні до азотфіксації мешкають в екстремальних біотопах ліпіди ізопреноїдно-гліцерол-фосфатні Система трьох доменів органічного світу (Woese, 1981)
Eubacteria Firmacutes Cyanophyta пластиди Eukaryota мітохондрії бактероїди хламідії протеобактерії еукаріоти зелені сірчані бактерії спірохети пурпурні бактерії Archaebacteria зелені несірчані бактерії Gracillicutes Mendosicutes Tenericutes прокаріоти гідролітичні екзоферменти наявні утилізують складні органічні сполуки мешкають в біотопах з широкою екол. амплітудою здатні до азотфіксації Progenota ліпіди гліцерол-фосфатні прокаріоти відсутні гідролітичні екзоферменти не засвоюють складні органічні сполуки здатні до азотфіксації мешкають в екстремальних біотопах ліпіди ізопреноїдно-гліцерол-фосфатні Система трьох доменів органічного світу (Woese, 1981)
") Архебактерії 1977 – Woese (Карл Воз)
Архебактерії 1977 – Woese (Карл Воз)
 Справжні бактерії
Справжні бактерії
 (Sogin M. L Early evolution and origin") Походження евкаріот – гіпотеза М. Согайна (1991) (Sogin M. L Early evolution and origin of eukaryotes. - , Current Opinion in Genetics and Development // 1991, 1: 457 -463)
Походження евкаріот – гіпотеза М. Согайна (1991) (Sogin M. L Early evolution and origin of eukaryotes. - , Current Opinion in Genetics and Development // 1991, 1: 457 -463)
") Походження евкаріот – гіпотеза М. Согайна (1991)
Походження евкаріот – гіпотеза М. Согайна (1991)
 Гипотетическая схема этапов эволюции эндомембранных систем") Походження евкаріот: ядро, ЕПР, Гольджі (Becker, Melkonian, 1996) Гипотетическая схема этапов эволюции эндомембранных систем (по: Becker, Melkonian, 1996). А — плазматическая мембрана (PM) прокариот состоит из фосфолипидов, кольцевая хромосома прикреплена к ней. Б—участок PM, примыкающий к хромосоме, погружается в цитоплазму, окружая ядро (N) и формируя эндоплазматический ретикулум (ER). В — происходит разобщение между PM и ER: рециклирование белковых компонентов осуществляется в везикулах, возможно с клатриновым окаймлением; PM состоит из фосфолипидов и незначительного количества стеролов и гликолипидов. Г—дополнительная система мембран встраивается между ER и PM; PM состоит из фосфолипидов и значительного количества стиролов и гликолипидов.
Походження евкаріот: ядро, ЕПР, Гольджі (Becker, Melkonian, 1996) Гипотетическая схема этапов эволюции эндомембранных систем (по: Becker, Melkonian, 1996). А — плазматическая мембрана (PM) прокариот состоит из фосфолипидов, кольцевая хромосома прикреплена к ней. Б—участок PM, примыкающий к хромосоме, погружается в цитоплазму, окружая ядро (N) и формируя эндоплазматический ретикулум (ER). В — происходит разобщение между PM и ER: рециклирование белковых компонентов осуществляется в везикулах, возможно с клатриновым окаймлением; PM состоит из фосфолипидов и незначительного количества стеролов и гликолипидов. Г—дополнительная система мембран встраивается между ER и PM; PM состоит из фосфолипидов и значительного количества стиролов и гликолипидов.
 Походження мітохондрій • Гідрогеносоми
Походження мітохондрій • Гідрогеносоми
 Походження мікротрубочок Білок Fts. Z
Походження мікротрубочок Білок Fts. Z
 Fts. Z – попередник тубуліну евкаріот? Fts. Z – цитоскелетний білок прокаріот, представлений GTPазою, що фомує кільце поділу (Z-кільце) прокаріотичних клітин. Виявлений також у пластидах зелених, червоних, криптофітових та золотистих водорстей та у мітохондріях золотистих водоростей та слизовиків. В мітохондріях опістоконтів (тварин та грибів) функції Fts. Z виконують динаміни. а) зв’язує та гідролізує GTP; Fts. Z, подібно до тубуліну: б) має іденичний до тубуліну GTP-зв’язуючий мотив - GGGTG(T/S)G, саме у сайті зв’язування GTP (N-термінальний домен) Fts. Z та тубуліни гомологічні; в) хоча за амінокислотними послідовностями С-термінальні домени не гомологічні, проте вони мають подібну структуру; третинна структура Fts. Z схожа зі структурою альфа/бета-тубулінових гетеродимерів г) здатний збиратися у профіламенти, утворює двовимірні складки та профіламентні кільця; д) механізми збірки протофіламентів у Fts. Z та тубулінів подібні (гідролізує GTP тільки у полімерній формі завдяки проникненню петлі Т 7 сусідньої протофіламента в активний сайт зв’язування нуклеотидів) Але схожіть амінокислотних послідовностей Fts. Z та тубуліну невисока (менша від 20%). Профламенти Fts. Z, на відміну від тубуліну, не утворюють мікротрубочок, проте можуть з’єднувтись латеральними сторонами у платівки Протофіламенти Fts. Z та динаміну, що утворюють спіралі (динамін) та спіралі й кільця (Fts. Z) (Erikson, 2000)
Fts. Z – попередник тубуліну евкаріот? Fts. Z – цитоскелетний білок прокаріот, представлений GTPазою, що фомує кільце поділу (Z-кільце) прокаріотичних клітин. Виявлений також у пластидах зелених, червоних, криптофітових та золотистих водорстей та у мітохондріях золотистих водоростей та слизовиків. В мітохондріях опістоконтів (тварин та грибів) функції Fts. Z виконують динаміни. а) зв’язує та гідролізує GTP; Fts. Z, подібно до тубуліну: б) має іденичний до тубуліну GTP-зв’язуючий мотив - GGGTG(T/S)G, саме у сайті зв’язування GTP (N-термінальний домен) Fts. Z та тубуліни гомологічні; в) хоча за амінокислотними послідовностями С-термінальні домени не гомологічні, проте вони мають подібну структуру; третинна структура Fts. Z схожа зі структурою альфа/бета-тубулінових гетеродимерів г) здатний збиратися у профіламенти, утворює двовимірні складки та профіламентні кільця; д) механізми збірки протофіламентів у Fts. Z та тубулінів подібні (гідролізує GTP тільки у полімерній формі завдяки проникненню петлі Т 7 сусідньої протофіламента в активний сайт зв’язування нуклеотидів) Але схожіть амінокислотних послідовностей Fts. Z та тубуліну невисока (менша від 20%). Профламенти Fts. Z, на відміну від тубуліну, не утворюють мікротрубочок, проте можуть з’єднувтись латеральними сторонами у платівки Протофіламенти Fts. Z та динаміну, що утворюють спіралі (динамін) та спіралі й кільця (Fts. Z) (Erikson, 2000)
") Філогенетичні дерева, побудовані на основі амінокислотних послідовностей Fts. Z (Beech et al. , 2000) та Fts. Z та тубулінів (Демчук, Блюм, 2005) підтверджують як гіпотезу походження тубулінів від Fts. Z, так і ціанофітну гіпотезу походження пластидних Fts. Z та альфа-протеобактеріальну гіпотезу походження мітохондріальних Fts. Z страменопілів та Amoebozoa типові GTP-ази зі збіркою за Rossman (типу p 21 Ras, GAPDH) пращурна GTP-аза з GTP-зв’язуючим мотивом та збіркою за Rossman “еубактеріальні” Fts. Z атипові GTP-ази: Fts. Z та тубуліни “архебактеріальні” Fts. Z Cyanophyta та пластид Fts. Z α-протеобактерій, мітохондрій Chrysophyta та Myxomycota Fts. Z архебактерій та фірмакутів тубуліни (схематизовано у відповідності до реконструкції Erickson, 1998, та дендритів Beech et al. , 2000; Демчук, Блюм, 2005)
Філогенетичні дерева, побудовані на основі амінокислотних послідовностей Fts. Z (Beech et al. , 2000) та Fts. Z та тубулінів (Демчук, Блюм, 2005) підтверджують як гіпотезу походження тубулінів від Fts. Z, так і ціанофітну гіпотезу походження пластидних Fts. Z та альфа-протеобактеріальну гіпотезу походження мітохондріальних Fts. Z страменопілів та Amoebozoa типові GTP-ази зі збіркою за Rossman (типу p 21 Ras, GAPDH) пращурна GTP-аза з GTP-зв’язуючим мотивом та збіркою за Rossman “еубактеріальні” Fts. Z атипові GTP-ази: Fts. Z та тубуліни “архебактеріальні” Fts. Z Cyanophyta та пластид Fts. Z α-протеобактерій, мітохондрій Chrysophyta та Myxomycota Fts. Z архебактерій та фірмакутів тубуліни (схематизовано у відповідності до реконструкції Erickson, 1998, та дендритів Beech et al. , 2000; Демчук, Блюм, 2005)
 1. За пластидним Fts. Z монофілія родопластів Chromalveolata не підтверджується 2. Відсутність Fts. Z у мітохондріях опістоконтів та його наявність в мітохондріях страменопілів та слизовиків не підтверждують гіпотезу T. Cavalier-Smith щодо первинності одноджгутикових евкаріот Ms. Fts. Z-cp – хлоропластний Fts. Z Mallomonas splendens (Chrysophyta) Філогенетичне дерево Fts. Z бактерій, пластид та мітохондрій в різних філах евкаріот (Beech et al. , 2000)
1. За пластидним Fts. Z монофілія родопластів Chromalveolata не підтверджується 2. Відсутність Fts. Z у мітохондріях опістоконтів та його наявність в мітохондріях страменопілів та слизовиків не підтверждують гіпотезу T. Cavalier-Smith щодо первинності одноджгутикових евкаріот Ms. Fts. Z-cp – хлоропластний Fts. Z Mallomonas splendens (Chrysophyta) Філогенетичне дерево Fts. Z бактерій, пластид та мітохондрій в різних філах евкаріот (Beech et al. , 2000)
 Походження та еволюція пластид
Походження та еволюція пластид
 ФСІІ b 6 -f НАДФ-Н 2 ФСІ CO 2 АТФ-синтетаза АТФ Цикл Кальвіна Ru. Bis. Co C 5 H 10 O 5 світлова фаза фотосинтезу темнова фаза фотосинтезу
ФСІІ b 6 -f НАДФ-Н 2 ФСІ CO 2 АТФ-синтетаза АТФ Цикл Кальвіна Ru. Bis. Co C 5 H 10 O 5 світлова фаза фотосинтезу темнова фаза фотосинтезу
 та вторинно-симбіотичних пластид (Bhattacharya and Medlin, 1998)") Походження первинно-симбіотичних (архепластид) та вторинно-симбіотичних пластид (Bhattacharya and Medlin, 1998)
Походження первинно-симбіотичних (архепластид) та вторинно-симбіотичних пластид (Bhattacharya and Medlin, 1998)
 Поширення пластид через вторинні симбіози Первинний симбіоз: гетеротрофний джгутиконосець + синьозелена водорість = архепластидні водорості (зелені, червоні, глаукоцистофітові) Вторинний симбіоз: гетеротрофний джгутиконосець + архепластидна водорість = евгленофіти, хлорарахніофіти, страменопіли, альвеоляти Третинний симбіоз (? ) Palmer, 2003
Поширення пластид через вторинні симбіози Первинний симбіоз: гетеротрофний джгутиконосець + синьозелена водорість = архепластидні водорості (зелені, червоні, глаукоцистофітові) Вторинний симбіоз: гетеротрофний джгутиконосець + архепластидна водорість = евгленофіти, хлорарахніофіти, страменопіли, альвеоляти Третинний симбіоз (? ) Palmer, 2003

 виник внаслідок симбіозу") Типи фотосинтетичного апарату 1. Прокаріотичний 2. Первинно-симбіотичний (пластида з двомембранною оболонкою) виник внаслідок симбіозу синьозеленої водорості з гетеротрофною платикристатою пластида вкрита лише двома мембранами; представлений трьома типами – хлоропластом, родопластом, цианопластом 3. Вторинно-симбіотичний (пластида з чотирьохмембранною оболонкою) виникав багаторазово, внаслідок симбіозів різних гетеротрофних клітин з різними первинно-пластидними водоростями Представлений чотирма основними типами: вторинно-симбіотичний хлоропласт з нуклеоморфом вторинно-симбіотичний хлоропласт без нуклеоморфу вторинно-симбіотичний родопласт з нуклеоморфом вторинно-симбіотичний родопласт без нуклеоморфу
Типи фотосинтетичного апарату 1. Прокаріотичний 2. Первинно-симбіотичний (пластида з двомембранною оболонкою) виник внаслідок симбіозу синьозеленої водорості з гетеротрофною платикристатою пластида вкрита лише двома мембранами; представлений трьома типами – хлоропластом, родопластом, цианопластом 3. Вторинно-симбіотичний (пластида з чотирьохмембранною оболонкою) виникав багаторазово, внаслідок симбіозів різних гетеротрофних клітин з різними первинно-пластидними водоростями Представлений чотирма основними типами: вторинно-симбіотичний хлоропласт з нуклеоморфом вторинно-симбіотичний хлоропласт без нуклеоморфу вторинно-симбіотичний родопласт з нуклеоморфом вторинно-симбіотичний родопласт без нуклеоморфу
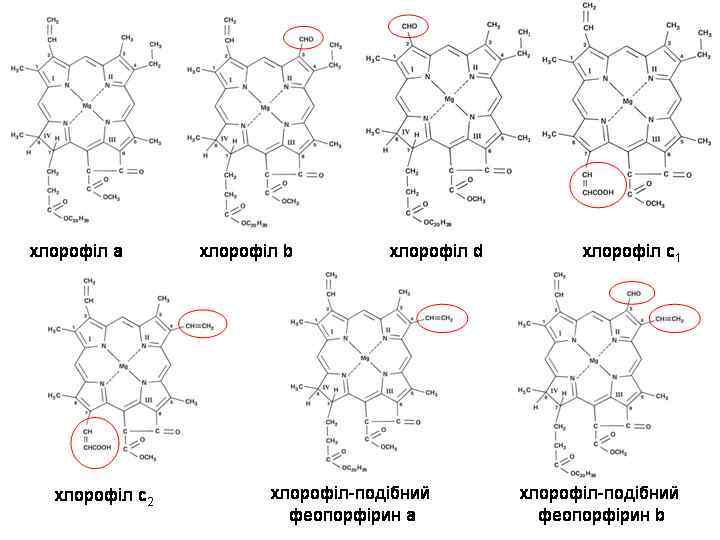
 максимуми поглинання: хл. а: 430, 660 нм") Спектри поглинання різних хлорофілів (у ацетоновому екстракті) максимуми поглинання: хл. а: 430, 660 нм хл. b: 435, 643 нм хл. c 1: 440, 583, 634 нм хл. c 2: 452, 586, 635 нм хл. d: 400, 456, 696 нм
Спектри поглинання різних хлорофілів (у ацетоновому екстракті) максимуми поглинання: хл. а: 430, 660 нм хл. b: 435, 643 нм хл. c 1: 440, 583, 634 нм хл. c 2: 452, 586, 635 нм хл. d: 400, 456, 696 нм
 окислює дивініловий радикал до формілового (наявна у Acaryochloris: хл. d) хлорофілід-а-оксигеназа") феофорбід-подібна оксигеназа (CAO-like) окислює дивініловий радикал до формілового (наявна у Acaryochloris: хл. d) хлорофілід-а-оксигеназа (CAO) конвертує метиловий радікал у форміловий (наявна у Prochloron та Prochlorothrix: хл. а+b) вініл-редуктаза перетворює дивінілпротохлорофілід у моновінілпротохлорофілід (відсутня у Prochlorococcus: хлорофіл-подібні феопорфірини a та b) (більшість Cyanophyta: хл. а) (за Green, 2005)
феофорбід-подібна оксигеназа (CAO-like) окислює дивініловий радикал до формілового (наявна у Acaryochloris: хл. d) хлорофілід-а-оксигеназа (CAO) конвертує метиловий радікал у форміловий (наявна у Prochloron та Prochlorothrix: хл. а+b) вініл-редуктаза перетворює дивінілпротохлорофілід у моновінілпротохлорофілід (відсутня у Prochlorococcus: хлорофіл-подібні феопорфірини a та b) (більшість Cyanophyta: хл. а) (за Green, 2005)
: прохлорофітові ЛП") LHC, cao => хл. a+b LHC => хл. a ЛП cao (хлорофілід-а-оксигеназа): прохлорофітові ЛП rbc. L: протеобактерії, ЛП cao-like: Acaryochloris (? ) ЛП LHC (донор не з’ясований) та подальше заміщення pcb (Prochlorophyte-like Chlorophyll-binding proteins) на LHC pcb (cao відсутній) => хл. a кластери генів atp/rps 2/rpo та psb. B/NH pcb. A+pcb. C (прохлорофіти) або isi. A (більшість ціанофітів) (відповідно до: Green, 2005; Chen et al. , 2005; Gogarten et al. , 2002; Kühl et al. , 2005)
LHC, cao => хл. a+b LHC => хл. a ЛП cao (хлорофілід-а-оксигеназа): прохлорофітові ЛП rbc. L: протеобактерії, ЛП cao-like: Acaryochloris (? ) ЛП LHC (донор не з’ясований) та подальше заміщення pcb (Prochlorophyte-like Chlorophyll-binding proteins) на LHC pcb (cao відсутній) => хл. a кластери генів atp/rps 2/rpo та psb. B/NH pcb. A+pcb. C (прохлорофіти) або isi. A (більшість ціанофітів) (відповідно до: Green, 2005; Chen et al. , 2005; Gogarten et al. , 2002; Kühl et al. , 2005)
 Схема будови PDF-системи: 1 – динамінове кільце, 2 – зовнішне PD-кільце, 3 – зовнішня (цитозольна) та внутрішня (строматична) мембрани оболонки пластиди, 4 – внутрішне PD-кільце, 5 – Z-кільце (за Yoshida et al. , 2006; Kuroiwa et al. , 2008).
Схема будови PDF-системи: 1 – динамінове кільце, 2 – зовнішне PD-кільце, 3 – зовнішня (цитозольна) та внутрішня (строматична) мембрани оболонки пластиди, 4 – внутрішне PD-кільце, 5 – Z-кільце (за Yoshida et al. , 2006; Kuroiwa et al. , 2008).
 PDV 2 PDV 1 2") DRP 5 B MCD 1 ARC 6 (Ftn 2) PDV 2 PDV 1 2 Fts. Z 2 ARC 3 1 Fts. Z 1 1 Min. E Min. D MSLs MCD 1 GC 1 (Yfc. H) 2 GC 1 (Yfc. H) ARC 3 (Fts. Z+PIP 5 K)
DRP 5 B MCD 1 ARC 6 (Ftn 2) PDV 2 PDV 1 2 Fts. Z 2 ARC 3 1 Fts. Z 1 1 Min. E Min. D MSLs MCD 1 GC 1 (Yfc. H) 2 GC 1 (Yfc. H) ARC 3 (Fts. Z+PIP 5 K)
 DRP 5 B outer PD-ring PDV 1 ARC 6 inner PD-ring PDV 2 MCD 1 Min. D 2 Fts. Z 2 ARC 3 1 Fts. Z 1 1 Min. E Min. D MSLs MCD 1 GC 1 2 GC 1
DRP 5 B outer PD-ring PDV 1 ARC 6 inner PD-ring PDV 2 MCD 1 Min. D 2 Fts. Z 2 ARC 3 1 Fts. Z 1 1 Min. E Min. D MSLs MCD 1 GC 1 2 GC 1
 DRP 5 B PDV 2 DRP 5 B PDV 1 ARC 3 PDV 2 ARC 6 2 Fts. Z 2 MCD 1 1 Fts. Z 1 Min. D ARC 3 Min. E Min. D MSLs MCD 1 GC 1 Min. D MCD 1 GC 1 PDV 1 Fts. Z 1 1 Fts. Z 2 2 ARC 6 ARC 3 PDV 1 PDV 2 DRP 5 B
DRP 5 B PDV 2 DRP 5 B PDV 1 ARC 3 PDV 2 ARC 6 2 Fts. Z 2 MCD 1 1 Fts. Z 1 Min. D ARC 3 Min. E Min. D MSLs MCD 1 GC 1 Min. D MCD 1 GC 1 PDV 1 Fts. Z 1 1 Fts. Z 2 2 ARC 6 ARC 3 PDV 1 PDV 2 DRP 5 B
 Схема eволюції апарату поділу пластиди у архепластидних евкаріот
Схема eволюції апарату поділу пластиди у архепластидних евкаріот
 Багатоклітинні тварини Деякі найпростіші Тварини Rhodophyta Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Glaucocystophyta Eustigmatophyta Вищі рослини Chlorophyta Basidiomycota Haptophyta Стаменопіли Chytridiomycota Справжні гриби Апікомплекси Альвеоляти Oomycota Деякі найпростіші Рослини Zygomycota Dinophyta Chrysophyta Raphidophyta Cryptophyta Ascomycota Інфузорії Амебоїдні найпростіші Chlorarachniopyta Plasmodiophoromycota Myxomycota Амебо-флагеляти Платикристати Евкаріоти Euglenophyta Тубулокристати Кінетопластиди Дискокристати Acrasiomycota Деякі найпростіші Гіпохондріати Прокаріоти Cyanophyta Архебактерії Евбактерії
Багатоклітинні тварини Деякі найпростіші Тварини Rhodophyta Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Glaucocystophyta Eustigmatophyta Вищі рослини Chlorophyta Basidiomycota Haptophyta Стаменопіли Chytridiomycota Справжні гриби Апікомплекси Альвеоляти Oomycota Деякі найпростіші Рослини Zygomycota Dinophyta Chrysophyta Raphidophyta Cryptophyta Ascomycota Інфузорії Амебоїдні найпростіші Chlorarachniopyta Plasmodiophoromycota Myxomycota Амебо-флагеляти Платикристати Евкаріоти Euglenophyta Тубулокристати Кінетопластиди Дискокристати Acrasiomycota Деякі найпростіші Гіпохондріати Прокаріоти Cyanophyta Архебактерії Евбактерії
 Нижчі рослини та новітня мегасистема: Adl et al. , 2005 "While this revised classification of protists is proposed by the International Society of Protistologists, it should be noted that it is the work of a committee that worked in collaboration with specialists from many societies (phycologists, mycologists, parasitologists, and other protistologists), and that many experts were consulted on issues as needed. However, it should not be assumed that all contributors agreed on all points". Встановлює 6 кластерів, "that may represent the basic groupings similar to traditional ‘‘kingdoms. " "We have included in our descriptions in Table 2 the apomorphies for each group"
Нижчі рослини та новітня мегасистема: Adl et al. , 2005 "While this revised classification of protists is proposed by the International Society of Protistologists, it should be noted that it is the work of a committee that worked in collaboration with specialists from many societies (phycologists, mycologists, parasitologists, and other protistologists), and that many experts were consulted on issues as needed. However, it should not be assumed that all contributors agreed on all points". Встановлює 6 кластерів, "that may represent the basic groupings similar to traditional ‘‘kingdoms. " "We have included in our descriptions in Table 2 the apomorphies for each group"
 Chromalveolata Glaucocystophyta Тварини Dictyochophyta Bacillariophyta") Багатоклітинні тварини Вищі рослини Chlorophyta Oomycota, ін. псевдогриби (2) Chromalveolata Glaucocystophyta Тварини Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Archaeplastida Деякі найпростіші (Mesomycetozoa, Choanomonada) Eustigmatophyta Апікомплекси (*) Chrysophyta Raphidophyta Альвеоляти Rhodophyta Microsporidia (+) Basidiomycota Ascomycota Glomeromycota Zygomycota Chytridiomycota Cryptophyta (!) Haptophyta (!) Cercozoa (incl. Plasmodiophoromycota) Chlorarachniophyta Тубулокр. Платикр. Amoebozoa Heterolobosea (incl. Acrasiomycota) Jakobida (Reclinomonas) Дискокр. Euglenophyta Гіпохондр. Деякі монадні найпростіші (Fornicata, Parabasalia, Preaxostyla) (+) криптон (*) апікопласт (1) базальна група (Nozaki, 2005) Деякі амебоїдні найпростіші (Haplosporidia, Foraminifera, Radiolaria) Rhizaria Opistokonta Кінетопластиди Dinophyta Опаліни, бікосоїциди, актінофрідії Стаменопіли Гриби Інфузорії Myxomycota (1) Деякі амебоїдні найпростіші (Tubulinea, Flabellinea, Acanthamoebidae, Entamoebida (+) Mastigamoebidae, Pelomyxa) Excavata Архебактерії (2) ціанофітна gnd (6 -фосфоглюконат дегідрогеназа) (Yoon et al. , 2002) Cyanophyta Евбактерії
Багатоклітинні тварини Вищі рослини Chlorophyta Oomycota, ін. псевдогриби (2) Chromalveolata Glaucocystophyta Тварини Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Archaeplastida Деякі найпростіші (Mesomycetozoa, Choanomonada) Eustigmatophyta Апікомплекси (*) Chrysophyta Raphidophyta Альвеоляти Rhodophyta Microsporidia (+) Basidiomycota Ascomycota Glomeromycota Zygomycota Chytridiomycota Cryptophyta (!) Haptophyta (!) Cercozoa (incl. Plasmodiophoromycota) Chlorarachniophyta Тубулокр. Платикр. Amoebozoa Heterolobosea (incl. Acrasiomycota) Jakobida (Reclinomonas) Дискокр. Euglenophyta Гіпохондр. Деякі монадні найпростіші (Fornicata, Parabasalia, Preaxostyla) (+) криптон (*) апікопласт (1) базальна група (Nozaki, 2005) Деякі амебоїдні найпростіші (Haplosporidia, Foraminifera, Radiolaria) Rhizaria Opistokonta Кінетопластиди Dinophyta Опаліни, бікосоїциди, актінофрідії Стаменопіли Гриби Інфузорії Myxomycota (1) Деякі амебоїдні найпростіші (Tubulinea, Flabellinea, Acanthamoebidae, Entamoebida (+) Mastigamoebidae, Pelomyxa) Excavata Архебактерії (2) ціанофітна gnd (6 -фосфоглюконат дегідрогеназа) (Yoon et al. , 2002) Cyanophyta Евбактерії
, яка покладена в основу системи Adl et.") ? ? Узагальнена мегасистема Т. Cavalier-Smith (2002), яка покладена в основу системи Adl et. al. , (2005) Cavalier-Smith, 2007
? ? Узагальнена мегасистема Т. Cavalier-Smith (2002), яка покладена в основу системи Adl et. al. , (2005) Cavalier-Smith, 2007
 та Адл зі співавторами (2005) За Паттерсоном (1994),") Основна розбіжність між системами Паттерсона (1994) та Адл зі співавторами (2005) За Паттерсоном (1994), Альвеоляти, Страменопіли, Криптофітові та Гаптофітові водорості НЕ мають спільного пращура. Вторинно симбіотичні пластиди родофітного типу (родопласти) виникали багаторазово. За Адл зі співавт. (2005), ці таксони мають спільне походження (т. зв. Хромальвеоляти) і виникли внаслідок однократного вторинного симбіозу гетеротрофного господаря з червоною водорістю.
Основна розбіжність між системами Паттерсона (1994) та Адл зі співавторами (2005) За Паттерсоном (1994), Альвеоляти, Страменопіли, Криптофітові та Гаптофітові водорості НЕ мають спільного пращура. Вторинно симбіотичні пластиди родофітного типу (родопласти) виникали багаторазово. За Адл зі співавт. (2005), ці таксони мають спільне походження (т. зв. Хромальвеоляти) і виникли внаслідок однократного вторинного симбіозу гетеротрофного господаря з червоною водорістю.
 вищі рослини Streptophyta Chlorophyta s. s. Cyanophyta Rhodophyta Glaucocystophyta редукція п. пл. ? Cryptophyta* Платикристати Haptophyta* Opistokonta Dinophyta Apicomplexa Ciliophora Chromophytes Stramenopiles Тубулокристати Chlorarachniophyta Rhizaria Amoebozoa Euglenophyta Kinetoplastida Excavata Дискокристати
вищі рослини Streptophyta Chlorophyta s. s. Cyanophyta Rhodophyta Glaucocystophyta редукція п. пл. ? Cryptophyta* Платикристати Haptophyta* Opistokonta Dinophyta Apicomplexa Ciliophora Chromophytes Stramenopiles Тубулокристати Chlorarachniophyta Rhizaria Amoebozoa Euglenophyta Kinetoplastida Excavata Дискокристати
 є дискусійним: Гіпотеза 1:") Положення Cryptophyta та Haptophyta у системі Adl et al. (2005) є дискусійним: Гіпотеза 1: поліфілія вторинно симбіотичних родопластів 1. Cryptophyta та Glaucocystophyta не мають апоморфії страменопілів - ретронем 2. Cryptophyta та Glaucocystophyta на 18 S r. DNA деревах виступають сестринськими групами (Bhattacharya & al. , 1995; Delwiche, Palmer, 1997). 3. Належність Cryptophyta та Haptophyta не лише до клади страменопілів, але й до хромальвеолят в цілому за ядерними генами (18 S r. DNA (Bhattacharya & al. , 1992; Bhattacharya, Schmidt, 1997; Mc. Fadden & al. , 1997, та ін. ), група ядерних генів, що кодують шість цитоплазматичних протеінів – актин, альфа- та бета-тубулін, EF-1, HSP 70, HSP 90 - (Harper et al. , 2005)) не підтверджується. 4. Cучасні Cryptophyta ймовірно походять від автотрофних Cryptophyta з первинно-симбіотичними пластидами, які втратили ці пластиди, захопивши як другого ендосимбіонта червону водорість. Ця гіпотеза підтверджується результатами дослідження генів, що кодують гліцеральдегід-3 -фосфат дегідрогеназу (GAPDH) (Liaud et al. , 1997). 5. За пластидними генами rbc. L, tuf. A та 16 S r. DNA родопласти Cryptophyta, Haptophyta та хромофітових водорстей потраляють у три різні клади (Bhattaharya, Medlin, 1995; Medlin et al. , 1997; Turner et al. , 1999) Гіпотеза 1 не узгоджується з системою Adl et al. (2005) у частині, що стосується мегаклад Archaeplastida та Chromalveolata, але в цілому узгоджується с системою Patterson (1995)
Положення Cryptophyta та Haptophyta у системі Adl et al. (2005) є дискусійним: Гіпотеза 1: поліфілія вторинно симбіотичних родопластів 1. Cryptophyta та Glaucocystophyta не мають апоморфії страменопілів - ретронем 2. Cryptophyta та Glaucocystophyta на 18 S r. DNA деревах виступають сестринськими групами (Bhattacharya & al. , 1995; Delwiche, Palmer, 1997). 3. Належність Cryptophyta та Haptophyta не лише до клади страменопілів, але й до хромальвеолят в цілому за ядерними генами (18 S r. DNA (Bhattacharya & al. , 1992; Bhattacharya, Schmidt, 1997; Mc. Fadden & al. , 1997, та ін. ), група ядерних генів, що кодують шість цитоплазматичних протеінів – актин, альфа- та бета-тубулін, EF-1, HSP 70, HSP 90 - (Harper et al. , 2005)) не підтверджується. 4. Cучасні Cryptophyta ймовірно походять від автотрофних Cryptophyta з первинно-симбіотичними пластидами, які втратили ці пластиди, захопивши як другого ендосимбіонта червону водорість. Ця гіпотеза підтверджується результатами дослідження генів, що кодують гліцеральдегід-3 -фосфат дегідрогеназу (GAPDH) (Liaud et al. , 1997). 5. За пластидними генами rbc. L, tuf. A та 16 S r. DNA родопласти Cryptophyta, Haptophyta та хромофітових водорстей потраляють у три різні клади (Bhattaharya, Medlin, 1995; Medlin et al. , 1997; Turner et al. , 1999) Гіпотеза 1 не узгоджується з системою Adl et al. (2005) у частині, що стосується мегаклад Archaeplastida та Chromalveolata, але в цілому узгоджується с системою Patterson (1995)
 Opistokonta (гриби) 1. 57 1. 5 І Chlorophyta 1. 3 Rhodophyta (кронові)") Opistokonta (тварини) Opistokonta (гриби) 1. 57 1. 5 І Chlorophyta 1. 3 Rhodophyta (кронові) Rhodophyta (Cyanidiales) ІІ Glaucocystophyta первинний ендосимбіоз (пластида) 1. 2 1. 1 Cryptophyta 0. 98 Haptophyta 0. 65*** вторинний ендосимбіоз (пластида) 1. 6 1. 5 1. 4 1. 3 1. 2 1. 1 1. 0 0. 9 0. 8 0. 7 0. 6 Страменопіли* ІІІ Альвеоляти** млрд. років Топологія Opstokonta, Archaeplastida та Chromalveolata згідно до гіпотези монофілії вторинносимбіотичних родопластів та час дивергенції основних груп за результатами множинного аналізу 5 -и пластидних генів та розрахунків за технікою “молекулярного годинника” (за Yoon et al. , 2002) І – Opistokonta ІІ – Archaeplastida ІІІ - Chromalveolata * включаючи хромофітові, псевдогриби, найпростіші (опаліни, бікосоециди, актинофрідії) ** включаючи Dinophyta, Ciliophora, Apicomplexa *** за розрахунками Medlin et al. (1997)
Opistokonta (тварини) Opistokonta (гриби) 1. 57 1. 5 І Chlorophyta 1. 3 Rhodophyta (кронові) Rhodophyta (Cyanidiales) ІІ Glaucocystophyta первинний ендосимбіоз (пластида) 1. 2 1. 1 Cryptophyta 0. 98 Haptophyta 0. 65*** вторинний ендосимбіоз (пластида) 1. 6 1. 5 1. 4 1. 3 1. 2 1. 1 1. 0 0. 9 0. 8 0. 7 0. 6 Страменопіли* ІІІ Альвеоляти** млрд. років Топологія Opstokonta, Archaeplastida та Chromalveolata згідно до гіпотези монофілії вторинносимбіотичних родопластів та час дивергенції основних груп за результатами множинного аналізу 5 -и пластидних генів та розрахунків за технікою “молекулярного годинника” (за Yoon et al. , 2002) І – Opistokonta ІІ – Archaeplastida ІІІ - Chromalveolata * включаючи хромофітові, псевдогриби, найпростіші (опаліни, бікосоециди, актинофрідії) ** включаючи Dinophyta, Ciliophora, Apicomplexa *** за розрахунками Medlin et al. (1997)
 Гіпотеза 2: монофілітичність вторинно симбіотичних родопластів 1. У Cryptophyta, Haptophyta, Страменопілів та Альвеолят GAPDH (гліцеральдегід-3 -фосфат дегідрогеназа) пластид гомологічні (дупліковані цитозольні GAPDH), і не є гомологом перенесеної до ядра пластидної GADPH Rhodophyta (Fast et al. , 2001) 2. Монофілітичне походження вторинно симбіотичних родопластів підтверджується множинним аналізом 5 и пластидних генів: 16 S r. RNA, psa. A, psb. A, rbc. L, tuf. A (Yoon et al. , 2002 a) 3. Cryptophyta та Haptophyta включаються у базальну частину хромальвеолят за множинним аналізом ядерних генів, хоча й без високої бутстреп-підтримки (Harper et al. , 2005) 4. Розрахунки дивергенції опітоконтів та архепластид, радіації родофітів, часу проходження вторинного ендосимбіозу, дивергенції криптофітів, гаптофітів та хромальвеолят за технікою молекулярного годинника при різних способах його калібровки дозволяють запропонувати несуперечливу топологію для цих груп (Yoon et al. , 2002)
Гіпотеза 2: монофілітичність вторинно симбіотичних родопластів 1. У Cryptophyta, Haptophyta, Страменопілів та Альвеолят GAPDH (гліцеральдегід-3 -фосфат дегідрогеназа) пластид гомологічні (дупліковані цитозольні GAPDH), і не є гомологом перенесеної до ядра пластидної GADPH Rhodophyta (Fast et al. , 2001) 2. Монофілітичне походження вторинно симбіотичних родопластів підтверджується множинним аналізом 5 и пластидних генів: 16 S r. RNA, psa. A, psb. A, rbc. L, tuf. A (Yoon et al. , 2002 a) 3. Cryptophyta та Haptophyta включаються у базальну частину хромальвеолят за множинним аналізом ядерних генів, хоча й без високої бутстреп-підтримки (Harper et al. , 2005) 4. Розрахунки дивергенції опітоконтів та архепластид, радіації родофітів, часу проходження вторинного ендосимбіозу, дивергенції криптофітів, гаптофітів та хромальвеолят за технікою молекулярного годинника при різних способах його калібровки дозволяють запропонувати несуперечливу топологію для цих груп (Yoon et al. , 2002)
 є дискусійним: Гіпотеза 1:") Місце Cryptophyta та Haptophyta у системі Adl et al. (2005) є дискусійним: Гіпотеза 1: поліфілія вторинно симбіотичних родопластів 1. Cryptophyta та Glaucocystophyta не мають апоморфії страменопілів - ретронем 2. Cryptophyta та Glaucocystophyta на 18 S r. DNA деревах виступають сестринськими групами (Bhattacharya & al. , 1995; Delwiche, Palmer, 1997). 3. Належність Cryptophyta та Haptophyta не лише до клади страменопілів, але й до хромальвеолят в цілому за ядерними генами (18 S r. DNA (Bhattacharya & al. , 1992; Bhattacharya, Schmidt, 1997; Mc. Fadden & al. , 1997, та ін. ), група ядерних генів, що кодують шість цитоплазматичних протеінів – актин, альфа- та бета-тубулін, EF-1, HSP 70, HSP 90 - (Harper et al. , 2005)) не підтверджується. 4. Cучасні Cryptophyta ймовірно походять від автотрофних Cryptophyta з первинно-симбіотичними пластидами, які втратили ці пластиди, захопивши як другого ендосимбіонта червону водорість. Ця гіпотеза підтверджується результатами дослідження генів, що кодують гліцеральдегід-3 -фосфат дегідрогеназу (GAPDH) (Liaud et al. , 1997). 5. За пластидними генами rbc. L, tuf. A та 16 S r. DNA родопласти Cryptophyta, Haptophyta та хромофітових водорстей потраляють у три різні клади (Bhattaharya, Medlin, 1995; Medlin et al. , 1997; Turner et al. , 1999) Гіпотеза 1 не узгоджується з системою Adl et al. (2005) у частині, що стосується мегаклад Archaeplastida та Chromalveolata, але узгоджується с системою Patterson (1995)
Місце Cryptophyta та Haptophyta у системі Adl et al. (2005) є дискусійним: Гіпотеза 1: поліфілія вторинно симбіотичних родопластів 1. Cryptophyta та Glaucocystophyta не мають апоморфії страменопілів - ретронем 2. Cryptophyta та Glaucocystophyta на 18 S r. DNA деревах виступають сестринськими групами (Bhattacharya & al. , 1995; Delwiche, Palmer, 1997). 3. Належність Cryptophyta та Haptophyta не лише до клади страменопілів, але й до хромальвеолят в цілому за ядерними генами (18 S r. DNA (Bhattacharya & al. , 1992; Bhattacharya, Schmidt, 1997; Mc. Fadden & al. , 1997, та ін. ), група ядерних генів, що кодують шість цитоплазматичних протеінів – актин, альфа- та бета-тубулін, EF-1, HSP 70, HSP 90 - (Harper et al. , 2005)) не підтверджується. 4. Cучасні Cryptophyta ймовірно походять від автотрофних Cryptophyta з первинно-симбіотичними пластидами, які втратили ці пластиди, захопивши як другого ендосимбіонта червону водорість. Ця гіпотеза підтверджується результатами дослідження генів, що кодують гліцеральдегід-3 -фосфат дегідрогеназу (GAPDH) (Liaud et al. , 1997). 5. За пластидними генами rbc. L, tuf. A та 16 S r. DNA родопласти Cryptophyta, Haptophyta та хромофітових водорстей потраляють у три різні клади (Bhattaharya, Medlin, 1995; Medlin et al. , 1997; Turner et al. , 1999) Гіпотеза 1 не узгоджується з системою Adl et al. (2005) у частині, що стосується мегаклад Archaeplastida та Chromalveolata, але узгоджується с системою Patterson (1995)
 ЛПГ rbc. L: протеобактерії Cyanophyta вищі рослини Streptophyta Chlorophyta s. s. Rhodophyta Glaucocystophyta редукція п. пл. ? Cryptophyta* Haptophyta* Opistokonta Dinophyta Apicomplexa Ciliophora Chromophytes Stramenopiles Chlorarachniophyta Rhizaria Amoebozoa Euglenophyta Kinetoplastida Excavata
ЛПГ rbc. L: протеобактерії Cyanophyta вищі рослини Streptophyta Chlorophyta s. s. Rhodophyta Glaucocystophyta редукція п. пл. ? Cryptophyta* Haptophyta* Opistokonta Dinophyta Apicomplexa Ciliophora Chromophytes Stramenopiles Chlorarachniophyta Rhizaria Amoebozoa Euglenophyta Kinetoplastida Excavata
 Гіпотеза 2: монофілітичність вторинно симбіотичних родопластів 1. У Cryptophyta, Haptophyta, Страменопілів та Альвеолят GAPDH (гліцеральдегід-3 -фосфат дегідрогеназа) пластид гомологічні (дупліковані цитозольні GAPDH), і не є гомологом перенесеної до ядра пластидної GADPH Rhodophyta (Fast et al. , 2001) 2. Монофілітичне походження вторинно симбіотичних родопластів підтверджується множинним аналізом 5 и пластидних генів: 16 S r. RNA, psa. A, psb. A, rbc. L, tuf. A (Yoon et al. , 2002 a) 3. Cryptophyta та Haptophyta включаються у базальну частину хромальвеолят за множинним аналізом ядерних генів, хоча й без високої бутстреп-підтримки (Harper et al. , 2005) 4. Розрахунки дивергенції опітоконтів та архепластид, радіації родофітів, часу проходження вторинного ендосимбіозу, дивергенції криптофітів, гаптофітів та хромальвеолят за технікою молекулярного годинника при різних способах його калібровки дозволяють запропонувати несуперечливу топологію для цих груп (Yoon et al. , 2002) Топологія Bhattaharya на часовый шкалі – дати схему
Гіпотеза 2: монофілітичність вторинно симбіотичних родопластів 1. У Cryptophyta, Haptophyta, Страменопілів та Альвеолят GAPDH (гліцеральдегід-3 -фосфат дегідрогеназа) пластид гомологічні (дупліковані цитозольні GAPDH), і не є гомологом перенесеної до ядра пластидної GADPH Rhodophyta (Fast et al. , 2001) 2. Монофілітичне походження вторинно симбіотичних родопластів підтверджується множинним аналізом 5 и пластидних генів: 16 S r. RNA, psa. A, psb. A, rbc. L, tuf. A (Yoon et al. , 2002 a) 3. Cryptophyta та Haptophyta включаються у базальну частину хромальвеолят за множинним аналізом ядерних генів, хоча й без високої бутстреп-підтримки (Harper et al. , 2005) 4. Розрахунки дивергенції опітоконтів та архепластид, радіації родофітів, часу проходження вторинного ендосимбіозу, дивергенції криптофітів, гаптофітів та хромальвеолят за технікою молекулярного годинника при різних способах його калібровки дозволяють запропонувати несуперечливу топологію для цих груп (Yoon et al. , 2002) Топологія Bhattaharya на часовый шкалі – дати схему
 Opistokonta (гриби) 1. 57 1. 5 І Chlorophyta 1. 3 Rhodophyta (кронові)") Opistokonta (тварини) Opistokonta (гриби) 1. 57 1. 5 І Chlorophyta 1. 3 Rhodophyta (кронові) Rhodophyta (Cyanidiales) ІІ Glaucocystophyta 1. 1 1. 2 Cryptophyta 0. 98 вторинний ендосимбіоз Haptophyta 0. 65*** Страменопіли* ІІІ Альвеоляти** 1. 6 1. 5 1. 4 1. 3 1. 2 1. 1 1. 0 0. 9 0. 8 0. 7 0. 6 млрд. років Топологія Opstokonta, Archaeplastida та Chromalveolata згідно до гіпотези монофілії вторинносимбіотичних родопластів та час дивергенції основних груп за результатами множинного аналізу 5 -и пластидних генів та розрахунків за технікою “молекулярного годинника” (за Yoon et al. , 2002) І – Opistokonta ІІ – Archaeplastida ІІІ - Chromalveolata * включаючи хромофітові, псевдогриби, найпростіші (опаліни, бікосоециди, актинофрідії) ** включаючи Dinophyta, Ciliophora, Apicomplexa *** за розрахунками Medlin et al. (1997)
Opistokonta (тварини) Opistokonta (гриби) 1. 57 1. 5 І Chlorophyta 1. 3 Rhodophyta (кронові) Rhodophyta (Cyanidiales) ІІ Glaucocystophyta 1. 1 1. 2 Cryptophyta 0. 98 вторинний ендосимбіоз Haptophyta 0. 65*** Страменопіли* ІІІ Альвеоляти** 1. 6 1. 5 1. 4 1. 3 1. 2 1. 1 1. 0 0. 9 0. 8 0. 7 0. 6 млрд. років Топологія Opstokonta, Archaeplastida та Chromalveolata згідно до гіпотези монофілії вторинносимбіотичних родопластів та час дивергенції основних груп за результатами множинного аналізу 5 -и пластидних генів та розрахунків за технікою “молекулярного годинника” (за Yoon et al. , 2002) І – Opistokonta ІІ – Archaeplastida ІІІ - Chromalveolata * включаючи хромофітові, псевдогриби, найпростіші (опаліни, бікосоециди, актинофрідії) ** включаючи Dinophyta, Ciliophora, Apicomplexa *** за розрахунками Medlin et al. (1997)
 (Bhattacharya, Medlin, 1995;") 2. Хлоропласти Euglenophyta – від Prasinophyceae, Chlorarachniophyta – від Chlorophyceae (Chlamydomonas) (Bhattacharya, Medlin, 1995; Delwiche, Palmer, 1997; Mc. Fadden et al. , 1997)
2. Хлоропласти Euglenophyta – від Prasinophyceae, Chlorarachniophyta – від Chlorophyceae (Chlamydomonas) (Bhattacharya, Medlin, 1995; Delwiche, Palmer, 1997; Mc. Fadden et al. , 1997)
 вищі рослини Streptophyta Chlorophyta s. s. Rhodophyta Glaucocystophyta Cryptophyta* Haptophyta Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Eustigmatophyta Chrysophyta Raphydophyta Pelagophyceae Bolidophyceae Oomycota, Hyphochytriomycota Опаліни, Бікосоециди, Актинофрідії Labyrynthulomycota
вищі рослини Streptophyta Chlorophyta s. s. Rhodophyta Glaucocystophyta Cryptophyta* Haptophyta Dictyochophyta Bacillariophyta Phaeophyta Xanthophyta Eustigmatophyta Chrysophyta Raphydophyta Pelagophyceae Bolidophyceae Oomycota, Hyphochytriomycota Опаліни, Бікосоециди, Актинофрідії Labyrynthulomycota
 Фікобіліни (фікоцианін,") Біохімічні ознаки відділів водоростей 1. Склад пігментів: Хлорофіли (а, b, c, d) Фікобіліни (фікоцианін, аллофікоцианін, фікоеритрин) Каротиноїди: Каротини (α-, β-, γ-, ε-каротин) Ксантофіли: лютеінового ряду діатоксантинового ряду специфічні ксантофіли 2. Продукти асиміляції: α - 1, 4 – глюкани (глікоген, крохмаль синьозелених водоростей, багрянковий крохмаль β- 1, 3 – глюкани (хризоламінарин, парамілон)
Біохімічні ознаки відділів водоростей 1. Склад пігментів: Хлорофіли (а, b, c, d) Фікобіліни (фікоцианін, аллофікоцианін, фікоеритрин) Каротиноїди: Каротини (α-, β-, γ-, ε-каротин) Ксантофіли: лютеінового ряду діатоксантинового ряду специфічні ксантофіли 2. Продукти асиміляції: α - 1, 4 – глюкани (глікоген, крохмаль синьозелених водоростей, багрянковий крохмаль β- 1, 3 – глюкани (хризоламінарин, парамілон)
 Postulated roles of Fts. Z and dynamin in organelle constriction. In the cyanobacterial and a-proteobacterial ancestors of chloroplasts and mitochondria, Fts. Z ring formation and cell wall synthesis at midcell were presumably both essential for cell division. During organelle evolution, Fts. Z was recruited to function in constriction in the lineages leading to plants and algae, but not fungi or animals. In higher plant chloroplasts and algal mitochondria, Fts. Z 1 and Fts. Z-mt are localized inside their respective organelles and may be functionally analogous to bacterial Fts. Z 2 in chloroplasts, and dynamin in yeast and animal mitochondria, are proposed to function on the cytosolic surface in organelle restriction, each perhaps having replaced the requirement for cell wall synthesis in division of the endosymbionts. The potential participation of cytosolic and organellar partners, respectively, of Fts. Z-mt in algae, and dynamin in yeast and animals, is indicated by question marks (Osteryoung, 2000)
Postulated roles of Fts. Z and dynamin in organelle constriction. In the cyanobacterial and a-proteobacterial ancestors of chloroplasts and mitochondria, Fts. Z ring formation and cell wall synthesis at midcell were presumably both essential for cell division. During organelle evolution, Fts. Z was recruited to function in constriction in the lineages leading to plants and algae, but not fungi or animals. In higher plant chloroplasts and algal mitochondria, Fts. Z 1 and Fts. Z-mt are localized inside their respective organelles and may be functionally analogous to bacterial Fts. Z 2 in chloroplasts, and dynamin in yeast and animal mitochondria, are proposed to function on the cytosolic surface in organelle restriction, each perhaps having replaced the requirement for cell wall synthesis in division of the endosymbionts. The potential participation of cytosolic and organellar partners, respectively, of Fts. Z-mt in algae, and dynamin in yeast and animals, is indicated by question marks (Osteryoung, 2000)
 ch mb m nu
ch mb m nu
 Аналіз складу генів, що входять до пластидних геномів представників різних філ (портативна пластидна гіпотези втрати генів, Grzebyk et al. , 2003), не дозволяє однозначно прийняти ані гіпотезу одноразового, ані гіпотезу багаторазових вторинних симбіозів Гіпотетична схема втрати генів внаслідок радіації фотоавтотрофних евкаріот, побудована за аналізом наявності 190 ідентифікованих білок-кодуючих генів (Grzebyk et al. , 2003)
Аналіз складу генів, що входять до пластидних геномів представників різних філ (портативна пластидна гіпотези втрати генів, Grzebyk et al. , 2003), не дозволяє однозначно прийняти ані гіпотезу одноразового, ані гіпотезу багаторазових вторинних симбіозів Гіпотетична схема втрати генів внаслідок радіації фотоавтотрофних евкаріот, побудована за аналізом наявності 190 ідентифікованих білок-кодуючих генів (Grzebyk et al. , 2003)
, яка демонструє перенос генів та їх редукцію у первиннота вторинно-симбіотичних пластидах") Схема Delwiche (2007), яка демонструє перенос генів та їх редукцію у первиннота вторинно-симбіотичних пластидах
Схема Delwiche (2007), яка демонструє перенос генів та їх редукцію у первиннота вторинно-симбіотичних пластидах