
Кембрий В кембрийском периоде основная жизнь была



















































































paleozoy.pptx
- Размер: 60.2 Мб
- Автор:
- Количество слайдов: 93
Описание презентации Кембрий В кембрийском периоде основная жизнь была по слайдам
Кембрий
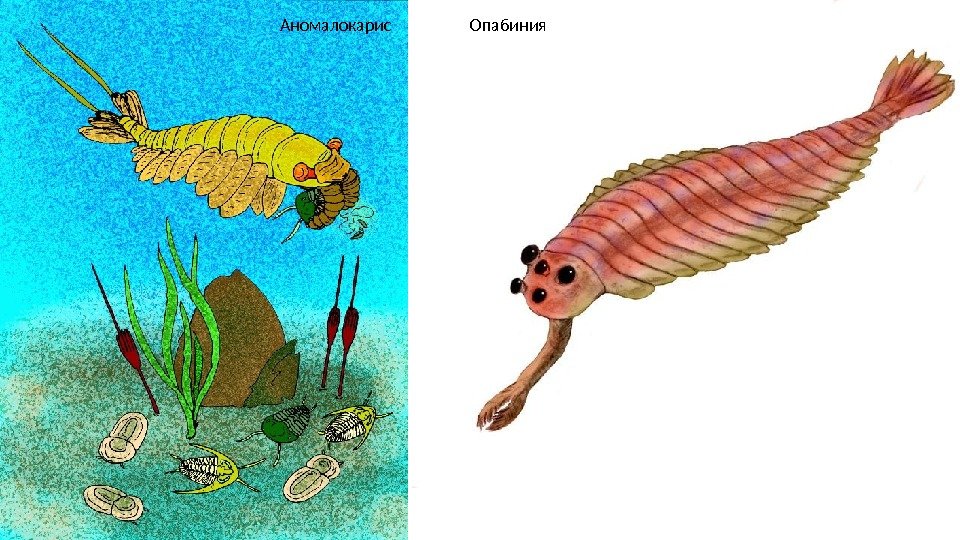
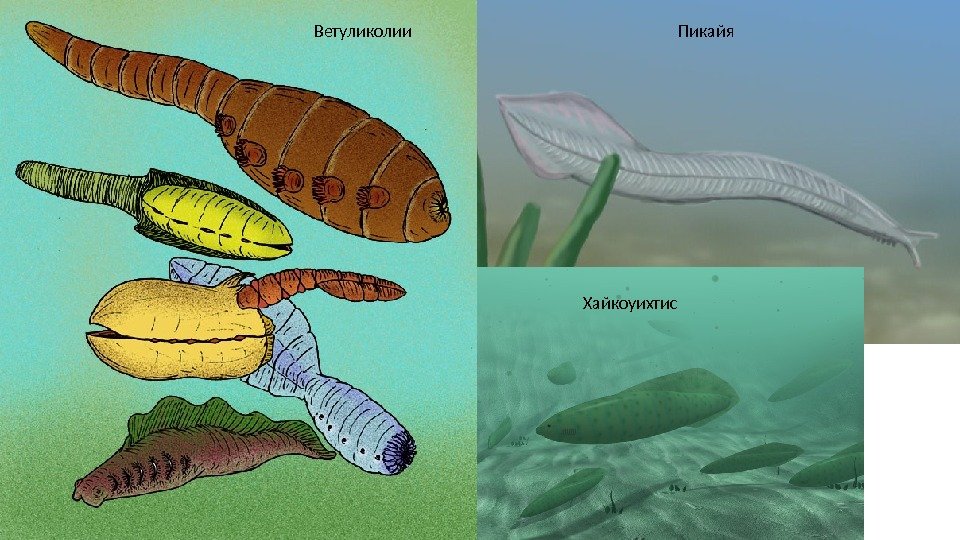
В кембрийском периоде основная жизнь была сосредоточена в морях. Самые известные обитатели кембрийских морей это донные хищные членистоногие трилобиты , которые обнаруживаются в отложениях, датируемых возрастом 530 млн лет. Длина их тела доходила до 1 метра. К этому типу также относят марреллу. Самым большим морским членистоногим (до 2 м в длину) считается охотник на трилобитов Аномалокарис , которого относят к классу Динокарид ( «ужасных креветок» ). Также в кембрийских морях обитали гусеницеобразная Галлюцигения и Опабиния. Наиболее странными организмами были Pambdelurion и Kerygmachela. Их длинное тело, состоящее их мягких сегментов, с парой широких «плавников» на большинстве сегментов и парой сегментированных придатков сзади делают их схожими с аномалокаридами. При этом наружные части верхней поверхности «плавников» имели рифлёные поверхности, которые могут быть жабрами. Под каждым «плавником» имеется короткая бескостная нога. Такое строение позволяет связать их с членистоногими. Не менее загадочны рыбообразные Ветуликолии , которых также по некоторым особенностям строения сближают с членистоногими. В результате кембрийского взрыва появились первые ракообразные (Ercaia) Ещё одним знаменательным представителем кембрийских морей были Бесчелюстные Хайкоуихтис и Пикайя (предки миног), принадлежащие к «нашему» типу хордовых
Аномалокарис Опабиния
Ветуликолии Пикайя Хайкоуихтис
Археоциаты
Хардия – новый представитель аномалокарид. См. http: // wiki. ru/sites/paleontologiya_stratigrafiya_i_geokhronologiya/id-news-21759. html Hurdia — это родственник аномалокариса. Подобно аномалокарису, его тело подразделяется на головной и туловищный отдел. Головной отдел не сегментирован, покрыт сетчатой кутикулой и вытянут в виде необычно длинного рострума. По бокам рострума на коротких стебельках сидят глаза , а по центру на нижней стороне головы располагается круглый рот. Рот очень даже зубастый — пять кругов из 32 зубных пластин венчают край рта и уходят внутрь глотки. По обеим сторонам рта имеются длинные членистые конечности с рядами шипов — именно они вместе с зубастым ртом придают животному облик завзятого хищника. Туловище поделено на 7– 9 сегментов, каждый из которых покрыт плотной кутикулой. Сегменты по бокам несут пластинки с жабрами в виде овальных щитков, внахлест налегающих друг на друга. Хвост оформлен в виде двух выростов. Авторы исследования представили наглядную и весьма художественную реконструкцию Hurdia.
Древнейшие иглокожие обнаруживаются примерно в это же время, хотя и имеются предположения о существовании иглокожих, либо их предшественников, уже в докембрии. Раннекембрийские Helicoplacus были сигароподобными вертикально стоящими созданиями длиной до 7 см. В отличие от современных иглокожих (радиально симметричных и имеющих рот в середине тела), они имели наружный спиральный пищевой желоб, вдоль которого пища двигалась ко рту, находящемуся сбоку. Другими обитателями кембрийских морей были полипы, медузы (стрекающие), гребневики и черви (нематоды, приапулиды, форониды, щетинкочелюстные). Весьма загадочна небольшая виваксия , которая имела хитиновый панцирь, состоящий из длинных вертикальных и коротких перекрывающих горизонтальных игл. Также она обладала чем-то, схожим с радулой (хитиновый зубастый «язык» , как у моллюсков. Некоторые исследователи полагают, что такие особенности приближают Wiwaxia к кольчатым червям. Кембрийская фауна широко представлена ракушками, создателями которых были как моллюски, так и плеченогие. Среди них особенно выделяются хиолиты. Они получили известность как «мелкая ракушковая фауна» (англ. small shelly fossils). Не менее широко были представлены губки, в том числе археоциаты.
Виваксия
т. Chaetognatha — Морские стрелки
Личинка хаоборуса (коретра) конвергентно похожа на взрослых щетинкочелюстных
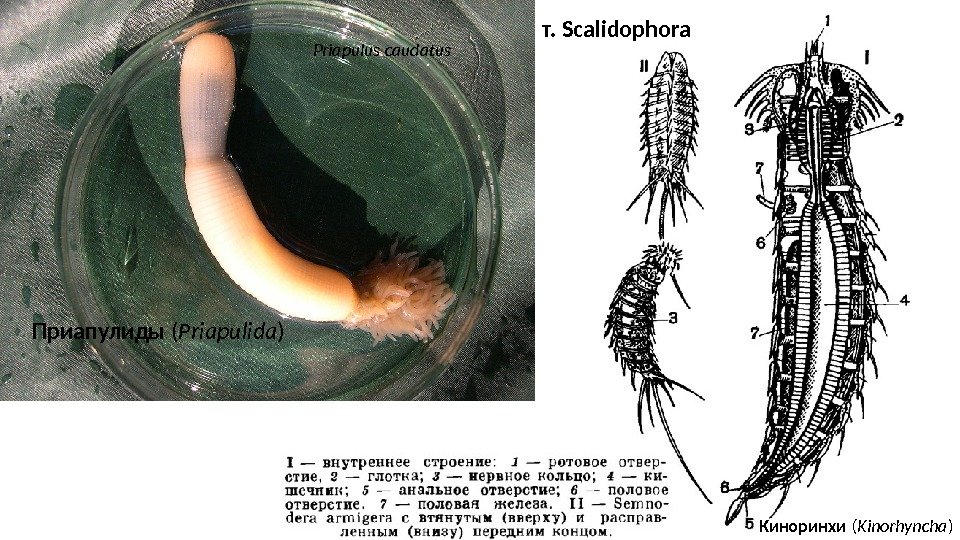
Priapulus caudatus т. Scalidophora Приапулиды ( Priapulida ) Киноринхи ( Kinorhyncha )
Схема строения тела Scalidophora : ca — хвостовые придатки; cg — хвостовой «ганглий» ; gn — гонада; gt — кишка; in — центральная часть интроверта; ir — ретракторы интроверта; mc — ротовой конус; ng — шейный «ганглий» ; nk — шейный отдел; os — вооружение ротового конуса; pn — скопления протонефридиев (правая часть: вариант киноринх; левая: Vinctiplicata); rb — кольцевой мозг; sc — скалиды; tk — туловище; vn — брюшной нервный ствол
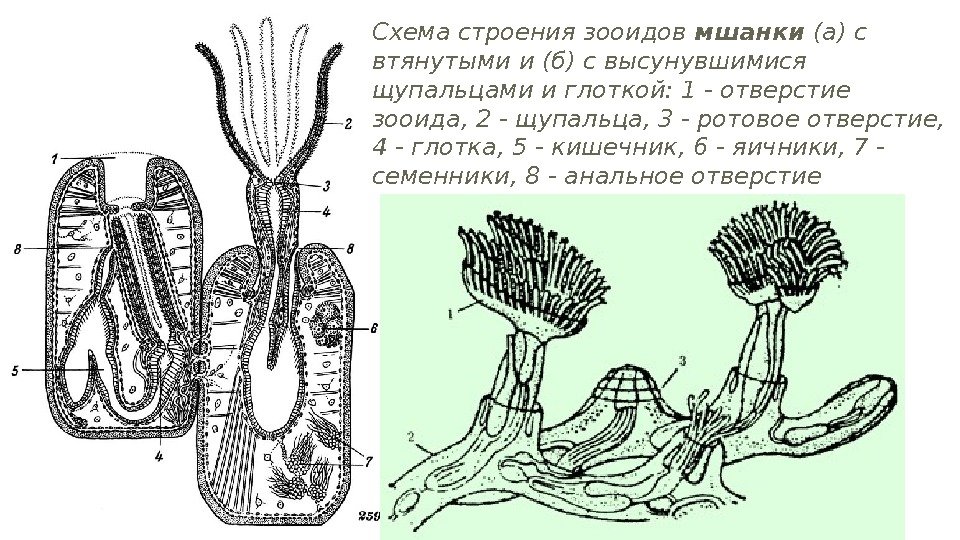
Схема строения зооидов мшанки (а) с втянутыми и (б) с высунувшимися щупальцами и глоткой: 1 — отверстие зооида, 2 — щупальца, 3 — ротовое отверстие, 4 — глотка, 5 — кишечник, 6 — яичники, 7 — семенники, 8 — анальное отверстие
т. Брахиоподы (плеченогие) Lingula anatina
начался 485, 4 ± 1, 9 млн лет назад, кончился 443, 4 ± 1, 5 млн лет назад, продолжался, таким образом, около 42 млн лет В ордовикском периоде , как и в кембрии, господствовали бактерии , в т. ч. цианобактерии. Пышного развития достигают известковые зелёные и красные водоросли , обитавшие в тёплых морях на глубине до 50 м. О существовании в ордовикском периоде наземной растительности свидетельствуют остатки спор и редкие находки отпечатков стеблей, вероятно, принадлежавших сосудистым растениям. Из животных ордовикского периода хорошо известны только обитатели морей , океанов , а также некоторые представители пресных и солоноватых вод. Существовали представители почти всех типов и большинства классов морских беспозвоночных. Тогда же жили первые хорошо изученные бесчелюстные позвоночные ( арандаспиды ). В толще вод океанов и морей обитали планктонные радиолярии и фораминиферы. В тепловодных морях обитали кораллы и другие кишечнополостные. Из иглокожих достигли пышного расцвета морские бутоны , морские пузыри , морские лилии , морские звёзды , эдриоастероидеи. Были широко распространены моллюски — двустворчатые , брюхоногие и головоногие , которые включали, среди прочих, такие роды, как ортоцерас , эндоцерас , онкоцерас и аскоцерас. Среди улиток было, в частности, много крылораковинных и беллерофонов. Кроме того, в ордовике были распространены ракоскорпионы , трилобиты , брахиоподы , мшанки , губки , граптолиты , м ечехвосты и многие другие животные. Ордовиком заканчивается крупный этап развития древне-палеозойского органического мира. К началу силура вымирают многие семейства среди граптолитов, брахиопод, кораллов, головоногих моллюсков и трилобитов, а также ряд своеобразных групп иглокожих, характерных только для ордовикского период
• Инфратип Gnathostomata ( челюстноротые ) • Класс Placodermi ( плакодермы ) † • Класс Acanthodii ( акантоды ) † • Класс Chondrichthyes ( хрящевые рыбы ) • Подкласс Elasmobranchii ( пластиножаберные ): акулы , скаты • Подкласс Holocephali • Класс Osteichthyes ( костные рыбы ) • Подкласс Actinopterygii ( лучепёрые рыбы ) • Подкласс Sarcopterygii ( лопастеперые рыбы ) • Надкласс Tetrapoda ( четвероногие ) • Класс Amphibia ( земноводные или амфибии), • Класс Reptilia ( пресмыкающиеся или рептилии) • Подкласс Anapsida ( анапсиды ) • Подкласс Diapsida ( диапсиды ) • Класс Aves ( птицы ) • Класс Mammalia ( млекопитающие ) • Подтип Vertebrata или Craniata ( позвоночные ) • Класс Agnatha ( бесчелюстные ) • Подкласс Myxinoidea ( миксины ) • Подкласс Petromyzontida ( миноги ) • Подкласс Conodonta † • Подкласс Pteraspidomorphi † и несколько других вымерших групп неясного положения. Тип Хордовые, Chordata
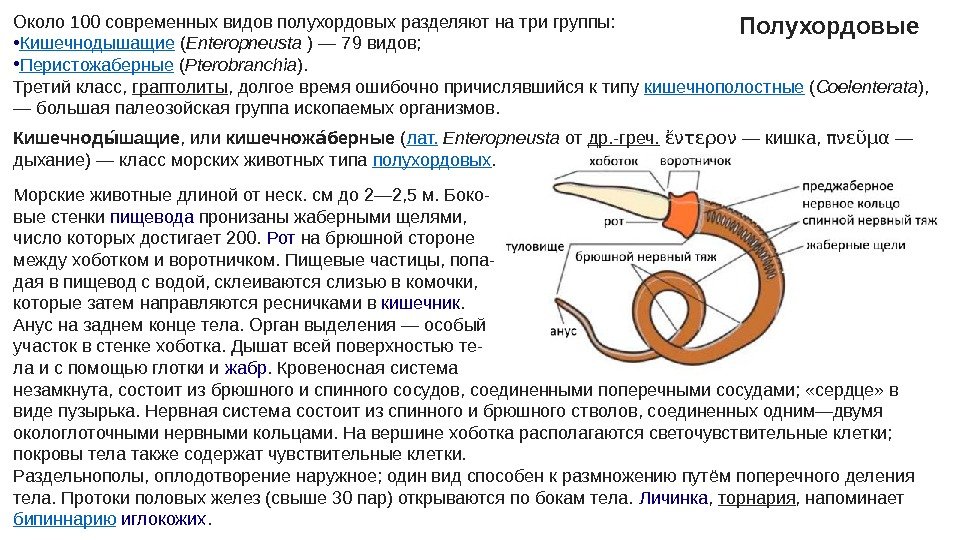
Около 100 современных видов полухордовых разделяют на три группы: • Кишечнодышащие ( Enteropneusta ) — 79 видов; • Перистожаберные ( Pterobranchia ). Третий класс, граптолиты , долгое время ошибочно причислявшийся к типу кишечнополостные ( Coelenterata ), — большая палеозойская группа ископаемых организмов. Полухордовые Кишечнод шащиеыы , или кишечнож берныеаы ( лат. Enteropneusta от др. -греч. ἔντερον — кишка, πνεῦμα — дыхание) — класс морских животных типа полухордовых. Морские животные длиной от неск. см до 2— 2, 5 м. Боко- вые стенки пищевода пронизаны жаберными щелями, число которых достигает 200. Рот на брюшной стороне между хоботком и воротничком. Пищевые частицы, попа- дая в пищевод с водой, склеиваются слизью в комочки, которые затем направляются ресничками в кишечник. Анус на заднем конце тела. Орган выделения — особый участок в стенке хоботка. Дышат всей поверхностью те- ла и с помощью глотки и жабр. Кровеносная система незамкнута, состоит из брюшного и спинного сосудов, соединенными поперечными сосудами; «сердце» в виде пузырька. Нервная система состоит из спинного и брюшного стволов, соединенных одним—двумя окологлоточными нервными кольцами. На вершине хоботка располагаются светочувствительные клетки; покровы тела также содержат чувствительные клетки. Раздельнополы, оплодотворение наружное; один вид способен к размножению путём поперечного деления тела. Протоки половых желез (свыше 30 пар) открываются по бокам тела. Личинка , торнария , напоминает бипиннарию иглокожих.
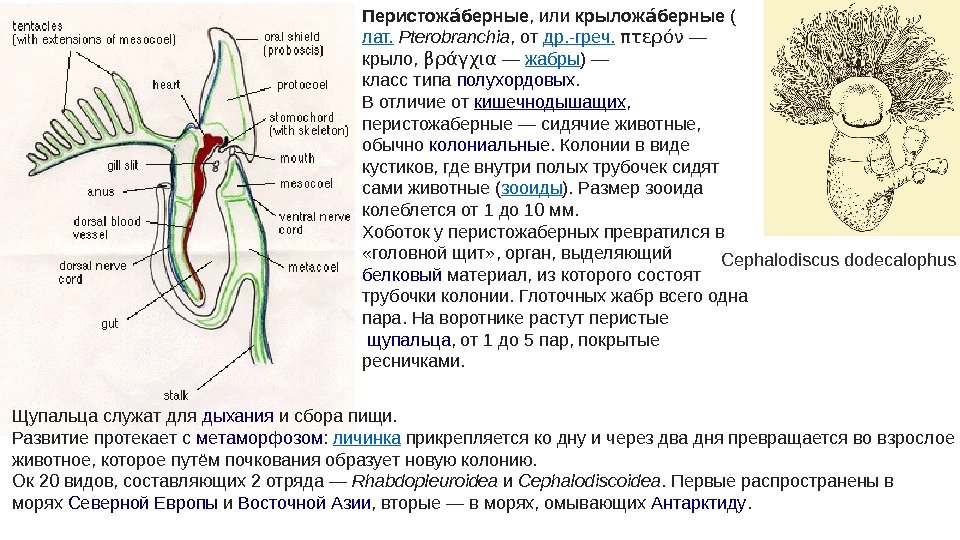
Перистож берныеаы , или крылож берныеаы ( лат. Pterobranchia , от др. -греч. πτερόν — крыло, βράγχια — жабры ) — класс типа полухордовых. В отличие от кишечнодышащих , перистожаберные — сидячие животные, обычно колониальные. Колонии в виде кустиков, где внутри полых трубочек сидят сами животные ( зооиды ). Размер зооида колеблется от 1 до 10 мм. Хоботок у перистожаберных превратился в «головной щит» , орган, выделяющий белковый материал, из которого состоят трубочки колонии. Глоточных жабр всего одна пара. На воротнике растут перистые щупальца , от 1 до 5 пар, покрытые ресничками. Cephalodiscus dodecalophus Щупальца служат для дыхания и сбора пищи. Развитие протекает с метаморфозом : личинка прикрепляется ко дну и через два дня превращается во взрослое животное, которое путём почкования образует новую колонию. Ок 20 видов, составляющих 2 отряда — Rhabdopleuroidea и Cephalodiscoidea. Первые распространены в морях Северной Европы и Восточной Азии , вторые — в морях, омывающих Антарктиду.
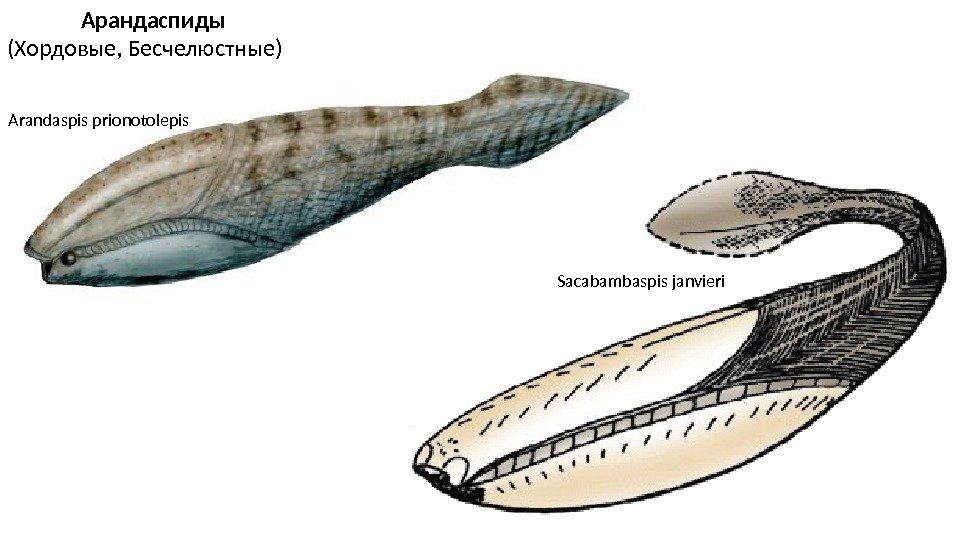
Арандаспиды (Хордовые, Бесчелюстные) Arandaspis prionotolepis Sacabambaspis janvieri
Арандаспиды имели вытянутое каплевидное тело длиной до 35 см, покрытое экзоскелетом. Брюшная сторона была более выпуклой, спинная — более плоской, рот – на нижней стороне головы. Единственный плавник — хвостовой, в него заходила хорда. Глаза находились на самом конце головы. В них были окостенения — склеротические кольца. Между глазами расположены ноздри, а позади — пара отверстий, которые обычно интерпретируются как пинеальные. У них было до 10 пар жаберных мешков, которые открывались наружу отдельными отверстиями (по другим данным — в общие проходы, открывавшиеся одним отверстием с каждой стороны). Скелет Эндоскелета у арандаспид не было или почти не было, зато был хорошо развит экзоскелет. Переднюю часть тела покрывали две большие костные пластины (спинная и брюшная), а заднюю — сильно вытянутые вертикальные чешуйки. Длина передних пластин составляет около половины длины животного, а толщина очень маленькая (у Arandaspis < 0, 1 мм). Они цельные (без следов слияния отдельных элементов) и не имеют признаков роста. Это означает, что они появлялись уже у взрослого животного. Между спинной и брюшной пластиной с каждой стороны был ряд из 15— 20 небольших многоугольных пластинок, между которыми находились жаберные отверстия. Кроме того, много рядов мелких костных пластинок было с нижней стороны рта. На спинном и брюшном щитке, а также на мелких чешуйках видны маленькие, но хорошо развитые каналы боковой линии Все элементы экзоскелета образованы аспидином — бесклеточной костью. В них различаются 3 слоя: нижний (пластинчатый), средний (сотоподобный или сетчатый) и верхний (бугорчатый).
Бластоидеи ( морские бутоны , лат. Blastoidea ) — класс вымерших беспозвоночных животных из группы иглокожие. Длина до 35 см. Жили с ордовика по пермь. Насчитывалось 300 видов. Характеризуются почковидной чашечкой пятилучевого строения, отсутствием рук. Снабжены были коротким стебельком, служившим для прикрепления к подводным предметам. Чашечка состоит из 13 соединенных между собой известковых пластинок, расположенных в три пояса с пятью амбулакральными полями. Наибольшим распространением бластоидеи пользуются в каменноугольном известняке (область реки Миссисипи ), реже встречаются в девонских отложениях ( Эйфель , Испания и пр. ) и в виде редких представителей в верхнесилурийских отложениях.
Шаровики ( морские пузыри , цистоидеи , лат. Cystoidea ) — вымерший класс иглокожих. Известны с ордовика. Вымерли, по разным данным, в каменноугольном либо в пермском периоде. От других иглокожих отличаются наличием в панцире поровых отверстий треугольной формы. Внешне напоминали морских лилий , с тем различием, что тело было овальным, а не в форме чаши. Ротовое отверстие располагалось на верхнем конце тела. Нижним концом тела шаровики крепились к морскому дну, часто с помощью стебельков. Анальное отверстие открывалось вбок. Поверхность тела делилась на пять, реже на три амбулакральные области, которые сходились ко рту. Некоторое количество небольших щупалец либо окружали рот, либо располагались рядами по телу, в зависимости от вида. Поровые отверстия служили, скорее всего, для циркуляции воды, чтобы обеспечить дыхание. У одних видов они сосредоточены в определённой области, тогда как у других распределены по всей поверхности тела.
Класс Морские лилии
Донные животные с телом в виде чашечки, в центре которой находится рот, а вверх отходит венчик из ветвящихся лучей (рук). Вниз от чашечки у стебельчатых морских лилий отходит прикрепительный стебелёк длиной до 1 м, прирастающий к грунту и несущий боковые придатки ( цирры ); у бесстебельчатых — только подвижные цирры. На концах цирр могут иметься зубчики, или «коготки» , которыми бесстебельные лилии прикрепляются к грунту. Морские лилии — единственные иглокожие , сохранившие характерную для предков иглокожих ориентацию тела: рот у них обращён вверх, а к поверхности грунта повёрнута спинная сторона. Как и у всех иглокожих, строение тела морских лилий подчинено пятилучевой радиальной симметрии. Рук 5, однако они могут неоднократно разделяться, давая от 10 до 200 «ложных рук» , снабжённых многочисленными боковыми веточками ( пиннулами ). Распущенный венчик морской лилии образует сеть для улавливания планктона и детрита. Руки на своей внутренней (ротовой) стороне имеют слизисто-ресничные амбулакральные желобки, ведущие ко рту; по ним захваченные из воды пищевые частички переносятся к ротовому отверстию. На краю чашечки, на коническом возвышении ( папилле ) находится анальное отверстие. Имеется внешний скелет; эндоскелет рук и стебелька состоит из известковых члеников. Внутрь рук и стебелька заходят ответвления нервной, амбулакральной и половой систем. Помимо внешней формы и ориентации спинно-брюшной оси тела, морские лилии отличаются от других иглокожих упрощённой амбулакральной системой — нет ампул, управляющих ножками, и мадрепоровой пластинки.
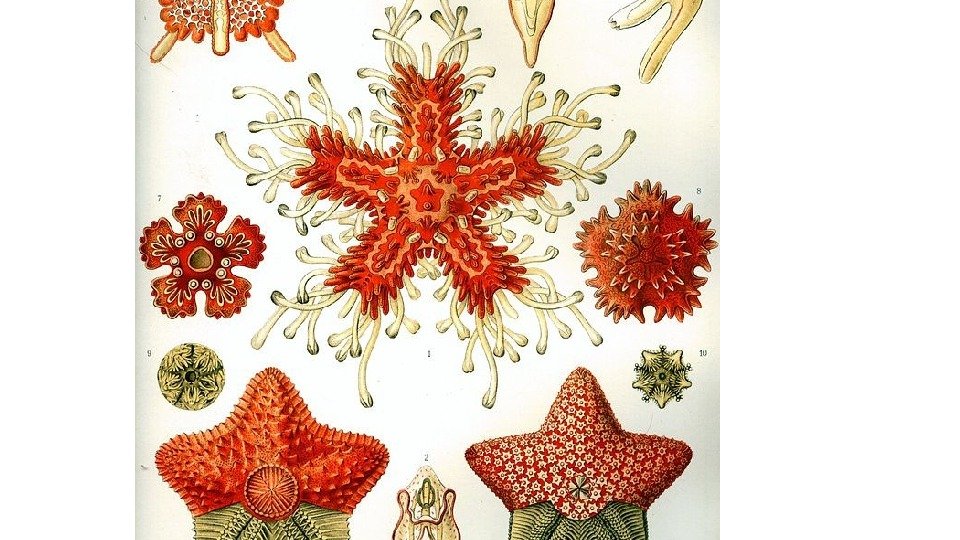
Звездообразные или пятиугольные иглокожие. Величина звёзд колеблется от 2 см до 1 м, хотя большинство — 12— 25 см. Многие виды ярко окрашены. Это малоподвижные животные, наиболее часто имеющие пять лучей. Лучи или руки — орган, отходящий от центрального плоского либо слабо выпуклого диска морской звезды. При укорочении лучей форма тела приближается к пятиугольной. Лучи заключают в себе пищеварительные выросты желудка и отростки половых органов, внутри луча находится продольный ряд позвонков. Кожа снабжена правильно расположенными скелетными пластинками, которые вооружены шипами, иглами, иногда педицелляриями. Амбулакральные ножки сидят в открытых бороздках, тянущихся по нижней стороне лучей до рта. Ножки представляют собой гибкие трубчатые выросты, как правило, с присосками на конце, и приводятся в действие давлением воды во внутренних каналах и ампулах амбулакральной системы. Рот в центре брюшной стороны, заднепроходное отверстие на спинной стороне; мадрепоровая пластинка на спинной стороне. Пищеварительная система отличается своеобразием. Над ртом внутри диска расположены два желудочных мешка; от верхнего (пилорического) отходит десять пищеварительных (печёночных) выростов, по два в каждую руку. Нижний желудок способен выворачиваться, обволакивая и частично переваривая пищу. Органы чувств: красные глазные пятна на концах лучей и осязательные окончания кожи. У морских звёзд сильно развита способность к регенерации : восстановлению оторванных лучей. Кроме того, часть морской звезды, включающая в себя достаточно крупный кусок центрального диска, способна вырасти в полноценное животное. Продолжительность жизни морской звезды — 20 и более лет.
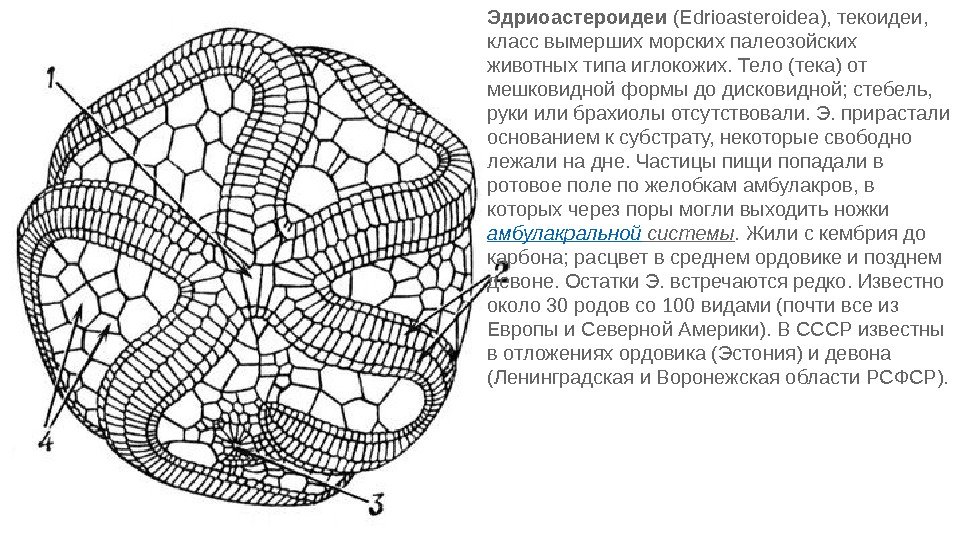
Эдриоастероидеи (Edrioasteroidea), текоидеи, класс вымерших морских палеозойских животных типа иглокожих. Тело (тека) от мешковидной формы до дисковидной; стебель, руки или брахиолы отсутствовали. Э. прирастали основанием к субстрату, некоторые свободно лежали на дне. Частицы пищи попадали в ротовое поле по желобкам амбулакров, в которых через поры могли выходить ножки амбулакральной системы. Жили с кембрия до карбона; расцвет в среднем ордовике и позднем девоне. Остатки Э. встречаются редко. Известно около 30 родов со 100 видами (почти все из Европы и Северной Америки). В СССР известны в отложениях ордовика (Эстония) и девона (Ленинградская и Воронежская области РСФСР).
Ракоскорпионы , или морские скорпионы , или эвриптериды (лат. Eurypterida) — ископаемый отряд членистоногих из класса Меростомовые подтипа (по другой сист. — надкласса) хелицеровых (Chelicerata). Отдельные представители достигали 2 метров в длину, однако характерные размеры большинства видов не превышали 20 см. Существовали в течение всего палеозоя 510— 248 млн лет назад. Ранние формы обитали на мелководье в морях. Около 325— 299 млн лет назад большая часть перешла к жизни в пресной воде. На примере эволюции ракоскорпионов хорошо прослеживается переход от водного образа жизни к сухопутному. Отряд включает в себя семейство Pterygotidae, живших около 470— 370 млн лет назад. Они были членами надсемейства Pterygotioidea. В эту группу входили виды, которые считаются крупнейшими членистоногими в истории планеты. Длина тела таких существ достигала двух с половиной метров. Включает в себя ряд родов: Acutiramus — которые жили с позднего силура до раннего девона. Исследования 2011 года показали, что данное животное скорее всего было падальщиком или даже вегетарианцем. К ракоскорпионам относятся крупнейшее известное членистоногое — Jaekelopterus rhenaniae , жившее примерно 460— 255 миллионов лет назад, и имевшее длину в 2, 5 метра.
Модель представителя ракоскорпионов (Национальный музей естественной истории в Вашингтоне)
Граптолиты — колониальные планктонные организмы, составлявшие в раннем палеозое (с кембрия до карбона) значительную часть планктона. К пневматофору — «плавательному пузырю», который редко сохраняется в ископаемом состоянии, прикреплялись длинные нити, на которых сидели тесно расположенные «домики» с зооидами. Домики состояли из хитиноподобного вещества. По своей микроструктуре аналогичны домикам современных полухордовых — птеробранхий . Каждый зооид состоял из ножки (стебелька), туловища, пары рук со щупальцами (такой пищесборный аппарат — лофофор — есть еще у мшанок и брахиопод, дальних родственников граптолитов). И еще была маленькая «головная» лопасть с коротким упругим тяжом, напоминающим зачаток хорды. http: //www. evolbiol. ru/_pgrapt. htm См. http: //www. evolbiol. ru/_pgrapt. htm
Крыложаберное Rhabdopleura — ближайший современный родственник вымерших граптолитов
Акантоды , или колючкоз быеуы (лат. Acanthodii, раньше — Acanthodei) — класс вымерших рыб. Существовали с позднего силура до ранней перми. Также появляются челюстноротые рыбы — костнопанцирные и беспанцирные. Расцвет граптолитов и прямораковинных наутилоидей. Заметно возросло разнообразие брахиопод. В позднем силуре появляются хрящекостные лучепёрые рыбы из отряда палеонискообразных (Palaeonisciformes). Megamastax amblyodus из позднего силура, костная рыба длиной до метра, на 2014 год считается первым позвоночным хищником, специализирующимся на поедании других позвоночных. Силур йский пер од (сил р иы иы уы ) — геологический период, третий период палеозоя: после ордовика, перед девоном. Начался 443, 4 ± 1, 5 млн лет назад, кончился 419, 2 ± 3, 2 млн лет назад. Продолжался, таким образом, около 24 млн лет. Комплекс отложений (горных пород), соответствующих данному возрасту, называется силур йской сист мой. ии еи Нижняя граница силура определяется по крупному вымиранию, в результате которого исчезло около 60 % видов существовавших в ордовике морских организмов, — так называемому ордовикско-силурийскому вымиранию. Назван в честь кельтского племени силуров, на землях обитания которого проводились геологические исcледования, способствовавшие его выделению.
Плакод рмыеы или пластинокожие рыбы (лат. Placodermi) — класс челюстноротых (Gnathostomata), живших в палеозое, начиная с раннего силура. Расцвет пластинокожих рыб пришёлся на девон , однако в его конце группа полностью вымерла. Ранние плакодермы жили исключительно в морской воде, но позже переселились и в солоноватоводные водоемы. Термин «панцирные рыбы», часто применяемый для класса в старой литературе, является некорректным и не отражает особенностей анатомии плакодерм: их так называемый «панцирь» на самом деле представлял собой набор пластин, сверху покрытых кожей, а не роговыми щитками. Titanichthys agassizi Характерным признаком плакодерм была защищённость головы и туловища кожными (дермальными) пластинками. Самых крупных размеров достигли хищник дунклеостей (до 10 м в длину), планктонофаги Homostius и Titanichthys (до 8 -9 м). Плакодермы были одними из первых позвоночных, у которых развились челюсти (из жаберных дуг бесчелюстных рыбообразных). При этом строение челюстей у плакодерм сильно отличается от строения челюстей у современных рыб. Так, у современных рыб эндохондральный нёбноквадратный элемент верхней челюсти расположен медиальнее основных мышц, смыкающих челюсти; у примитивных плакодерм он тесно связан со щекой, находясь латеральнее пространства, занимаемого челюстной мускулатурой. У большинства плакодерм вместо зубов к краям челюстей прикреплены крупные покровные костные пластины, причём ни у одного из видов не известно регулярной смены зубов, обычной для других рыб. Эти особенности позволяют предполагать, что плакодермы произошли от линии примитивных челюстноротых позвоночных, отличных от форм, которые дали начало современным хрящевым и костным рыбам. Все роды плакодерм обладали грудными и брюшными плавниками, но ни у одного не известен анальный. Хвост преимущественно гетероцеркальный (верхняя его лопасть, в которую заходит позвоночник, больше нижней).
Homosteus milleri
Ан спидыаы , беспанцирные , или бесщитковые ( лат. Anaspida от др. -греч. ἀν — — частица отрицания и ἀσπίς — «щит» ) — группа вымерших бесчелюстных позвоночных , традиционно относимая к остракодермам. Название получили за то, что их экзоскелет был развит намного слабее, чем у других остракодерм. Это были небольшие рыбоподобные животные, обитавшие в основном в пресных водоёмах или в прибрежных зонах морей. Жили в силуре и девоне. По некоторым предположениям, более популярным в прошлом — предки миног. Анаспиды имели веретенообразное и сплюснутое с боков тело, из-за чего напоминали рыб сильнее всех остальных остракодерм. У разных видов форма тела отличалась мало. В длину они достигали 15 см (Lasanius, возможно, 20 см). Иногда к анаспидам относят и некоторых животных (Jamoytius), достигавших 25 см. Хвостовой плавник анаспид, в отличие от большинства других позвоночных, был гипоцеркальный (хорда заходит в нижнюю лопасть). Из-за этого первые исследователи считали их спинную сторону брюшной и наоборот. Кроме хвостового плавника, анаспиды имели парные плавниковые складки, которые тянулись от грудных шипов на большую или меньшую длину вдоль нижней стороны тела. Гомологичны ли они грудным плавникам челюстноротых, неизвестно. Обычно был и анальный плавник. На каждом боку позади жабр был расположен трёхлучевой шип (у Lasanius — несколько), от которого и начиналась плавниковая складка. Шипом начинался и анальный плавник. Вдоль спины проходил гребень из особых шиповидных чешуй, часто загнутых крючком. Голова анаспид была защищена небольшими пластинками и чешуйками. Чешуя, уложенная косыми рядами, обычно (за исключением Lasanius) покрывала и остальную часть тела. На верхней части каждой стороны тела эти ряды чешуй были направлены передним концом вниз, а на нижней — вверх, образуя V-образные фигуры. Вероятно, они соответствовали миомерам, и в таком случае конфигурация миомеров у анаспид была такая же, как у ланцетника, но не как у круглоротых и рыб (у которых она W-образная). Все элементы экзоскелета состоят из пластинчатого аспидина (бесклеточной костной ткани); дентина (в отличие от многих других остракодерм) и эмали нет. Внутренний скелет не был минерализован и почти неизвестен, но у некоторых анаспид обнаружены дуги позвонков, крепившиеся к хорде. Рот анаспид был расположен на конце морды. С каждой стороны передней части тела у них был наклонный ряд из 6– 15 пар небольших жаберных отверстий (у некоторых форм, не всегда относимых к анаспидам, — более 30), переднее из которых было относительно далеко от глаза. Глаза большие, по бокам головы. Склеротических колец (окостенений в глазах) нет. В верхней части головы есть пинеальное отверстие (для теменного глаза), расположенное на специальной относительно крупной пластинке, а перед ним — ещё одно отверстие, которое обычно интерпретируется как назогипофизарное (объединявшее ноздрю и гипофизарный проход, как и у большинства других непарноноздрёвых бесчелюстных), хотя точно это не известно. Как и у многих других остракодерм, у анаспид была сейсмосенсорная система, каналы которой у них видны только на головных пластинках. Главные отличительные признаки анаспид — трёхлучевые шипы за жабрами, ряд своеобразных чешуй вдоль спины и ярко выраженная гипоцеркальность хвостового плавника.
Класс Cephalopoda – Головоногие моллюски Современные представители класса головоногих делятся на два подкласса: Четырёхжаберные ( Tetrabranchia ) и Двужаберные ( Dibranchia ). Если принимать во внимание ископаемые виды, то более точным будет деление на три других подкласса: • Подкласс † Аммоноидеи ( Ammonoidea ), • Подкласс Nautiloidea (в т. ч. современные Tetrabranchia ) • Подкласс Belemnitida (в т. ч. современные Dibranchia ) Четырёхжаберные ( Tetrabranchia ) содержит единственный род Nautilus Двужаберные ( Dibranchia ) Включают подавляющее большинство современных представителей класса, в том числе кальмаров ( Teuthida ), каракатиц ( Sepiida ) и осьминогов ( Octopoda ). У представителей этой группы раковина располагается внутри тела и подвергается сильной редукции, лишь у немногих сохраняя спиральную закрученность (Spirula spirula). Глаза обладают сложным строением и способны к аккомодации. В мантийной полости залегает пара жабр (ктенидиев) , которым соответствует пара предсердий.
• Род † Nectocaris • Род † Petalilium • Род † Vetustovermis † Nectocaris pteryx Впервые животное было открыто палеонтологами в 1910 году среди первых находок из знаменитого теперь местонахождения кембрийской фауны в сланцах Бёрджес) в Канаде. Достигало 2— 5 см длины, имело 2 щупальца, стебельчатые глаза, «плавники» по бокам тела, и вело хищный образ жизни. Жило в кембрийском периоде. В 2010 году канадские палеонтологи Мартин Смит (Martin Smith) и Жан-Бернард Кэрон (Jean-Bernard Caron) из Университета Торонто и Королевского музея Онтарио опубликовали в журнале Nature статью, в которой доказывают, что Nectocaridae — предки головоногих. Они считают, что у нектокариса был сифон, через который животное могло с силой выбрасывать воду, как это делают современные цефалоподы. Другие учёные считают нектокариса членистоногим из класса Dinocarida. Против причисления нектокариса к головоногим указывают некоторые факты, например, отсутствие у нектокариса раковины, характерной для ископаемых палеозойских головоногих моллюсков, данные эмбриологии не подтверждают родство нектокарисов с Cephalopoda. В целом, положение Nectocarididae в системе животного мира остаётся предметом дискуссий. Семейство † Nectocarididae , возможно, также относится к Cephalopoda)
Наутило деииы ( лат. Nautiloidea ) — надотряд головоногих моллюсков. Имеют наружную раковину , подразделённую на камеры (многие — спиральную). Многочисленные щупальца без присосок, несросшаяся воронка, проявление метамерии ( четыре ктенидия, четыре почки, четыре предсердия ). Раковины наутилоидей разделены перегородками на камеры, через которые проходит специальная трубка — сифон. Через сифон камеры могут быстро наполняться газом или водой, обеспечивая всплытие или погружение моллюска. Основные отличия раковин наутилоидей от раковин аммонитов — простая форма перегородок (не образуют сложных изгибов в месте крепления к стенкам раковины) и расположение сифона недалеко от центра перегородок. Наутилоидеи — хищники, в толще воды они захватывают щупальцами мелких беспозвоночных, нападая иногда на червей, других моллюсков и мелкую рыбу, могут есть и падаль. Раковины современных наутилусов очень красивы, иногда их используют при создании украшений. жк – жилая камера; ф – фрагмокон; вк – воздушные камеры; с – сифон; пвк – перегородки воздушных камер; п — первичная камера – протоконх; гл – глаза; щ – щупальца
Наутилоидеи известны из нижнекембрийских отложений (роды Volbortella, Salterella). В среднем кембрии известен род Vologdinella. Некоторые палеонтологи считают, что указанные моллюски не относятся к наутилоидеям. Типичные наутилоидеи определены из позднекембрийских отложений Сибири, Китая, Северной Америки. Эти наутилоидеи очень мелкие, со сжатым поперечным сечением и широким сифоном. В ордовике наблюдался расцвет наутилоидей. В этот период появились все известные отряды и семейства, увеличились размеры раковин, более разнообразными стали их форма, скульптура и строение сифона. В девоне, карбоне и перми количество наутилоидей несколько сокращается. В мезозое и кайнозое наутилоидеи представлены только одним отрядом. Среди современных наутилоидей известен только род Nautilus. Аммониты (лат. Ammonoidea) — вымерший подкласс головоногих моллюсков, существовавших с девона по мел. Своё название аммониты получили в честь древнеегипетского божества Амона с закрученными рогами. Вымерли в ходе мел-палеогенового вымирания. Раковина аммонитов была разделена на много камер, ближайшая к устью (от 0, 5 до 2 оборотов) была жилой. Большинство камер, если судить по современным наутилусам, было заполнено газом (воздушные камеры), несколько — жидкостью (гидростатические камеры). Перегородка между камерами аммонитов имеет гофрированный край, который образует сложную линию прикрепления к раковине — лопастную линию. Её строение — один из основных систематических признаков аммонитов. Выделяют четыре типа лопастной линии. Различна также скульптура раковины: различают гладкие и разнообразные скульптурированные раковины с различным типом ветвления ребер, местоположением бугорков и т. д. Размеры аммонитов от 1— 2 см до 2 м в диаметре (Parapuzosia seppenradensis).
Аммониты
Анатомия Гигантского кальмара рода Architeuthis Белемниты (лат. Belemnitida, от др. -греч. Βελεμνoν (белемнон) — стрела) — отряд вымерших головоногих моллюсков из подкласса двужаберных. Устаревшее название подкласса — внутрираковинные , так как все части их раковины располагались внутри тела. Эти хищные животные, как правило, хорошо плавали, имели плавники, крупные глаза, роговые челюсти, чернильный мешок, а на щупальцах были крючки. Белемниты обитали в морях с кембрийского по меловой период (516 — 66 млн лет назад), наиболее широко распространились с триаса , вымерли в конце мезозоя. В качестве ископаемых остатков часто находят ростр белемнитов, похожий на наконечник стрелы — отсюда происходит их греческое название.
Хрящевые ганоиды или хрящекостные рыбы (лат. Chondrostei) — подкласс лучепёрых рыб. Ископаемые формы известны с конца силурийского периода. Останки хищной рыбы Andreolepis hedei из отряда палеонискообразных (Palaeonisciformes), найденные в России, Швеции и Эстонии, имеют возраст 420 млн лет. Возникли хрящевые ганоиды в реках Лавразии, впадавших в океан Тетис. Современные группы хрящекостных рыб произошли от палеонисков. Современные хрящекостные рыбы достигают длины до 9 метров (белуга). Тело веретенообразной формы. Может быть покрыто костной, ганоидной чешуёй, пятью рядами костных жучек или голое. Иногда чешуйки сливаются, образуя пластинки. Имеется рострум, рот нижний. Хвост гетероцеркальный или укороченно гетероцеркальный, на верхней лопасти имеются ганоидная чешуя. Парные плавники расположены горизонтально по отношению к телу, анальное отверстие находится вблизи брюшных плавников. Брюшные плавники занимают абдоминальное положение, грудные сидят низко. Скелет состоит в основном из хрящевой ткани. Эндокраниум хрящевой, с хорошо развитыми покровными костями. Хорда нерасчленённая, имеются только хрящевые дуги, тела позвонков отсутствуют. В сердце есть артериальный конус, в кишечнике — спиральный клапан. Плавательный пузырь соединяется каналом с пищеводом (пузырь открытого типа).
Дев ноы (девонский период, девонская система) — четвёртый геологический период палеозойской эры. Начался 419, 2 ± 3, 2 млн лет назад, закончился 358, 9 ± 0, 4 млн лет назад. Продолжался, таким образом, около 60 млн лет. Этот период богат биотическими событиями. Жизнь бурно развивалась и осваивала новые экологические ниши. Название дано по имени графства Девонш р, или Девон в юго-западной Англии, на территории которого распространены ии геологические породы этого периода. На суше от риниофитов произошли плауновидные , хвощевидные , папоротникообразные и голосеменные растения, многие из них были представлены древесными формами (например археоптерисы ). Растения по всей Земле были одинаковыми, не было географической дифференциации. Появились первые наземные позвоночные. Палеонтологи предполагают, что лёгкие, которыми дышат наземные существа, первоначально возникли у обитающих в болотах рыб. От таких — кистеперых — рыб возникли земноводные. Одни из первых земноводных — ихтиостеги, акантостеги — обладали множеством рыбьих признаков, но имели вполне сформированные конечности. Они были тесно связаны с водой, может быть даже теснее, чем современные лягушки. Возникли пауки, клещи, насекомые — жизнь осваивала сушу. В морях в девонском периоде тоже происходили перемены. Появились первые аммониты — головоногие моллюски со спирально закрученными раковинами, которых ещё ждал расцвет в мезозое. Донные хищники ракоскорпионы — эвриптероидеи достигают 1, 5— 2 метров в длину. Трилобиты начинают вымирать, судя по всему, им стало сложно жить при таком изобилии хищников. Девон часто называют веком рыб. Действительно, бесчелюстные и челюстноротые заселяют практически все морские и пресноводные бассейны и достигают большого разнообразия. В конце периода происходит массовое вымирание животных. В середине периода существенно повышается разнообразие растений. Среди распространённых во второй половине девона групп — древовидные плаунообразные, первые хвощёвые, прапапоротники, прогимноспермы и первые голосеменные. Появился почвенный покров
http: //elementy. ru/news/431004 В отложениях раннего девона (около 400– 410 млн лет назад) на западе Германии обнаружено десятисантиметровое существо с двумя парами крупных конечностей и 10 парами мелких, представляющее собой переходную форму между среднекембрийскими хищниками аномалокарисами и настоящими членистоногими. Находка позволила существенно уточнить расположение базальных ветвей на эволюционном древе членистоногих. Schinderhannes bartelsi сочетает в себе признаки настоящих членистоногих и аномалокарисов. Реконструкция из обсуждаемой статьи в Science
Кстати, там же обнаружились формы, близкие к маррелле Reconstruction of Marrella Шиндерханнес жил в раннедевонскую эпоху, когда моря уже кишели гораздо более эволюционно «продвинутыми» членистоногими (трилобитами, эвриптеридами, ракообразными), а также крупными рыбами и головоногими моллюсками. Тем не менее он сохранил множество архаичных черт, характерных для кембрийских хищников. Авторы отмечают, что в этом раннедевонском лагерштатте присутствуют и другие архаичные родственники членистоногих, которые ранее были известны только из кембрия (в том числе формы, близкие к марелле). Таким образом, удивительная среднекембрийская фауна древних членистоногих и их родственников на самом деле не вымерла в кембрии, а только исчезла из поля зрения палеонтологов. Спустя 100 миллионов лет она вдруг снова появляется в уникальном «окне в прошлое» — раннедевонском лагерштатте в Бунденбахе.
Orthrozanclus reburrus — морское животное, жившее 505 млн лет назад, — видимо, является близким родственником общего предка моллюсков и кольчатых червей (рис. из обсуждаемой статьи в Science). Животное сочетает в себе признаки двух известных ранее ископаемых групп — халькиериид и виваксиид. Находка позволила объединить все эти формы в единую группу хальваксиид. Эта группа, по-видимому, близка к общим предкам моллюсков, кольчатых червей и брахиопод. Halkieria — загадочное животное, встречающееся в отложениях раннего и среднего кембрия (500– 540 млн лет назад), — попеременно трактовалось разными исследователями как близкое к предкам кольчатых червей, моллюсков или брахиопод. Масштаб 5 мм. (Изображения с сайта macroevolution. narod. ru )
Шипы и щетинки у Orthrozanclus почти такие же, как у виваксиид, а раковинка — как у халькиериид. Несмотря на некоторые существенные различия (например, у халькиериид, в отличие от Orthrozanclus, кроме передней раковинки имеется еще и задняя, а склериты минерализованы), авторы полагают, что их находка доказывает близкое родство и единое происхождение халькиериид, виваксиид и Orthrozanclus. Kimberella — более древнее животное, жившее в вендском периоде (свыше 542 млн лет назад), — возможно, была близка к общему предку всех Lophotrochozoa (из книги: Я. Е. Малаховская, А. Ю. Иванцов. Вендские жители земли) Simon Conway Morris, Jean-Bernard Caron. Halwaxiids and the Early Evolution of the Lophotrochozoans // Science. 2007. V. 315. P. 1255–
Одной из наиболее «продвинутых» кистепёрых рыб являлся тиктаалик ( Tiktaalik ), имевший ряд переходных признаков, сближающих его с земноводными. К таким признакам относятся укороченный череп, отделенная от пояса передних конечностей и относительно подвижная голова, наличие локтевого и плечевого суставов. Плавник тиктаалика мог занимать несколько фиксированных положений, одно из которых предназначалось для того, чтобы животное могло находиться в приподнятом положении над грунтом (вероятно, чтобы «ходить» на мелководье). Тиктаалик дышал через отверстия, расположенные на конце плоской «крокодильей» морды. Воду, а возможно и атмосферный воздух, в лёгкие нагнетали уже не жаберные крышки, а щёчные помпы. Некоторые из указанных при- способлений характерны так- же для кистепёрой рыбы Пандерихтис ( Panderichthys ).
Pederpes
Первые амфибии, появившиеся в пресных водоемах в конце девона , — ихтиостегиды (Ichthyostegidae). Они были настоящими переходными формами между кистепёрыми рыбами и земноводными. Так, у них были рудименты жаберной крышки, настоящий рыбий хвост, сохранялся клейтрум. Кожа была покрыта мелкой рыбьей чешуёй. Однако наряду с этим они имели парные пятипалые конечности наземных позвоночных (см. схему конечностей кистепёрых и древнейших земноводных). Ихтиостегиды жили не только в воде, но и на суше. Видимо, они не только размножались, но и кормились в воде, систематически выползая на сушу
Грудной плавник кистеперой рыбы и передняя лапа древней амфибии (по Кэрролл, 1992). 1 — клейтрум, 2 — лопатка, 3 — базалия, соответствующая плечевой кости, 4 — базалия, соответствующая локтевой кости, 5 — базалия, соответствующая лучевой кости, 6 — радиалии, 7 — ключица. Сравнение поясов передних конечностей кистепёрых рыб (слева) и амфибий (справа) (по Квашенко, 2014). 1 — клейтрум, 2 — лопатка, 3 — ключица, 4 — грудина, 5 — коракоид, 6 — предгрудина, 7 — загрудина.
Каменно гольный пер одуы иы , или карб ноы (С) — предпоследний (пятый) геологический период палеозойской эры. Начался 358, 9 ± 0, 4 млн лет назад, кончился 298, 9 ± 0, 15 млн лет назад. Продолжался, таким образом, около 60 млн лет. Название получил из-за сильного углеобразования в это время. Впервые появляются очертания величайшего суперконтинента в истории Земли — Пангеи. Пангея образовалась при столкновении Лавразии (включала Северную Америку и Евразию) с древним южным суперконтинентом Гондваной. Незадолго до столкновения Гондвана повернулась по часовой стрелке, так что её восточная часть (Индия, Австралия, Антарктида) переместилась к югу, а западная (Южная Америка и Африка) оказалась на севере. В результате поворота на востоке появился новый океан — Тетис , а на западе закрылся старый — океан Рея. В то же время океан между Балтикой и Сибирью становился все меньше; вскоре эти континенты тоже столкнулись В течение карбона появились среди беспозвоночных: новые отряды фораминифер, переднежаберные и легочные гастроподы, среди позвоночных — рептилии (котилозавры и звероподобные), среди высших растений — различные голосеменные: хвойные, кордаитовые и цикадовые. Массовых вымираний не наблюдалось. Вымерли только некоторые головоногие моллюски, иглокожие (текоидеи) и граптолиты (стереостолонаты). В целом карбон характеризуется процветанием многих групп организмов, таких как, фораминиферы — фузулиниды, кораллы — табулятоидеи, хететиды и ругозы (четырёхлучевые), среди моллюсков — гастроподы, наутилоидеи со спиральной раковиной, аммоноидеи (гониатиты), мшанки, замковые брахиоподы — продуктиды и спирефириды, морские лилии и древние морские ежи, также расцвет испытывают земноводные (стегоцефалы), членистоногие (в особенности насекомые) и высшие растения (плауны и хвощи). В морях царили разнообразные хрящевые рыбы (акулы и брадиодонты). Некоторые акулы (например, камподусы) могли достигать длины до 13 метров. Сохранялись пресноводные кистеперые, в том числе и рипидистии. Обитавшие в болотах рипидистии-ризодонты были высшими хищниками. Ризод, например, достигал 8 метров в длину и был самым страшным хищником пресных вод. Отмечается большое разнообразие амфибий. Эогиринус — крупное, до 4, 5 м длиной, земноводное, — возможно, охотился на манер аллигатора. А 15 -сантиметровый микробрахий питался мельчайшим животным планктоном. У бранхиозавра, похожего на головастика, были жабры. Зауроплевра и сцинкозавр больше напоминали тритонов. Помимо основных групп «амфибий» — батрахоморфов и рептилиеморфов — существовали разнообразные немногочисленные группы (лепоспондилы, локсомматиды, микрозавры, крассигириниды). В нижнем карбоне возникают примитивные формы рептилий, которые, избегая конкуренции и хищников, заселяли более сухие пространства. Сначала первые рептилии ещё обитали вблизи воды, как например вестлотиана, но позже они удалялись от неё всё дальше и дальше, пока не стали сухопутными животными. В карбоне дальнейшее распространение получили споровые растения: сигиллярии, лепидодендрон (плауновидные), каламиты (хвощевидные), ставроптерисы, различные ужовниковые (папоротниковидные), семенные хвощи, кордаиты (голосеменные). Возникшие семенные растения могли поселяться в более сухих местах обитания, так как особенности их размножения не связаны с наличием воды. Тёплые болота изобиловали насекомыми и земноводными. Среди деревьев порхали гигантские летучие тараканы, стрекозы (меганевры) и подёнки. В гниющей растительности пировали гигантские артроплевры, дальние родичи многоножек. В подлеске встречались также многочисленные арахниды: метровый пульмоноскорпий, различные пауки и далёкие предки клещей.
Педерпес ( Pederpes finneyae ) — примитивный лабиринтодонт ( «земноводное» ) начала карбона. Единственное четвероногое этой эпохи, известное по достаточно полному скелету. Педерпес заполняет так называемый «провал Ромера» — промежуток примерно в 20 миллионов лет между поздним девоном (временем существования примитивных водных тетрапод вроде ихтиостеги и акантостеги ) и появлением настоящих четвероногих в раннем карбоне. Педерпес достигал в длину около 1 метра (длина сохранившегося скелета — 65 см, хвост утрачен). Он обладал массивным высоким и узким черепом с мощными зубами. Борозды боковой линии погружены в кость в виде каналов (как у кистеперых), стремя сходно со стременем акантостеги. «Ушная вырезка» (в действительности, вероятно, вмещавшая брызгальце) неглубокая. Присутствуют «клыки» на сошниках, нёбо покрыто мелкими зубчиками, закрытое. Ребра расширены, напоминают ребра ихтиостеги. Конечности очень массивные, относительно короткие. Не исключается наличие добавочных пальцев на кисти, стопа пятипалая.
В девоне возник надотряд лабиринтодонтов (Labyrinthodontia). Ранние формы имели сравнительно небольшие размеры и рыбообразное тело. Более поздние достигали весьма крупных размеров (1 м и более) в длину, тело их было уплощено и заканчивалось коротким толстым хвостом. Лабиринтодонты существовали до конца юры и занимали наземные, околоводные и водные местообитания. К некоторым лабиринтодонтам ( Temnospondyli ) относительно близки предки бесхвостых — отряды Proanura, Eoanura, известные с конца карбона и из пермских отложений Seymouria baylorensis • Сильно изогнутая поверхность зубов , искривлённые дентин и эмаль, так, что на срезе виден лабиринт, от которого и произошло название группы. Впрочем, такая же структура зубов характерна для кистепёрых рыб — рипидистий. • Массивная крыша черепа , с отверстиями только для ноздрей, глаз и теменных глаз , подобных тем, что встречаются у Анапсид. За исключением поздних рептилообразных форм, череп лабиринтодонтов был довольно плоским со значительной площадью кожного панциря, который и дал группе прежнее имя Стегоцефалы • Ушные выемки за каждым глазом в задней части черепа. Для примитивных форм это может означать наличие брызгалец , а для более прогрессивных — барабанных перепонок • Сложные позвонки состоящие из четырёх элементов, intercentrum, двух pleurocentra, и дужки позвонка. Размеры элементов существенно разнятся в зависимости от видовой принадлежности того или иного лабиринтодонта. Поперечный срез зуба лабиринтодонта
Ранними представителями лабиринтодонтов были Рептилиоморфы , известные как Anthracosauria или Reptiliomorpha. Tulerpeton является наиболее ранним из известных сегодня видов, возможно, разделение групп произошло ещё до перехода от Девона к Каменноугольному периоду. Их черепа были относительно глубокими и узкими по сравнению с другими лабиринтодонтами. Передние и задние лапы большинства видов имели по пять пальцев. Некоторые ранние группы, обитавшие в солоноватой или морской водной средах, вернулись затем к более или менее полностью водной жизни. За исключением Diadectomorpha , наземные формы имели средние размеры и появились в начале Каменноугольного периода. Наиболее известен род Seymouria. Некоторые члены наиболее эволюционно развитой группы, Diadectomorpha , были травоядными и вырастали до нескольких метров в длину. Они имели крупные, бочкообразные тела. Более мелкие родственники Diadectomorpha стали предками первых рептилий. Это произошло в конце Каменноугольного периода Tulerpeton curtum известен по практически полностью сохранившимся плечевому поясу, передним и задним конечностям, фрагментам черепа и другим мелким частям. Тулерпетон достигал 60 см в длину, имел по 6 пальцев на каждой конечности и полностью утратил внутренние жабры. Строение скелета говорит о том, что он был более приспособленным к жизни на суше, чем акантостега , однако его ноги больше подходили для проталкивания туловища по мягкому влажному грунту, чем для ползания по земле. Скорее всего тулерпетон был обитал на морском мелководье, т. к. находки остатков сделаны в морских отложениях Tulerpeton curtum
Диадектоморфы ( лат. Diadectomorpha ) — клада (в ранге отряда или подотряда) крупных рептилиеподобных амфибий из карбона и нижней перми Еврамерики и верхней перми ( вучапинский ярус ) Азии , очень близкая к родоначальникам Amniota. Включают крупные (более 2 метров длиной) плотоядные и еще более крупные (свыше 3 метров) травоядные формы Limnoscelis Diadectes Размножение диадектоморф является важным вопросом. От его разрешения напрямую зависит, к какому классу нужно отнести данную группу — к амфибиям или рептилиям. Если бы эта группа принадлежала к амниотам, то они должны были откладывать амниотические яйца, пусть и примитивные. Окаменелостей яиц диадектоморфов до сих пор не обнаружено. Вероятно, диадектоморфы откладывали анамнийные яйца ( икру ) в воду. Однако, однозначно принадлежащие диадектоморфам личиночные формы также не известны науке. Таким образом, диадектоморфы выглядят переходным звеном между амфибиями и настоящими амниотами. Являются они амфибиями или боковой ветвью эволюции амниот, уверенно сказать нельзя. Некоторые исследователи сближают их с ранними синапсидами. Пока не будут найдены личинки или яйца диадектоморф, данная группа животных может быть с равным успехом отнесена как к амниотам, так и анамниям
Гефиростег ( лат. Gephyrostegus ) — вымерший род рептилиоморфных амфибий семейства гефиростегид из верхнего карбона (311, 45— 306, 95 млн лет) Чехии. Небольшое животное общей длиной 22 см, ящерицеподобное по плану строения и, предположительно, ведущее сходный с ящерицами образ жизни. Большие глаза и большое количество мелких заостренных зубов, указывают, что гефиростег был активным охотником на насекомых. Лабиринтодонты ( Ichthyostegalia, Reptiliomorpha, Temnospondyli, Lepospondyli ) Соленодонзавр Solenodonsaurus janenschi Вестлотиана. Вероятные родственники общего предка амниот (из группы Diadectomorpha) Насекомоядные, либо хищники, имели не-лабиринтодонтные зубы, некоторые откладывали яйца на суше
Темноспондильные (Temnospondyli) Считаются наиболее разнообразной группой лабиринтодонтов. Появились в позднем Девоне и различались по размеру от саламандроподобных Stereospondyli, которые сновали вдоль кромки воды и обитали в прибрежных зарослях, до гигантского бронированного Archegosauroidea, больше напоминавшего крокодила. Девятиметровый Prionosuchus был особенно крупным представителем этой группы, он известен как крупнейшая из когда-либо живших амфибий. Были также и полностью водные формы, как, например, Dvinosauria, и даже морские разновидности, например, Trematosauridae. Темноспондильные могли стать предтечами современных лягушек и саламандр в конце Пермского или начале Триасового периода Platyoposaurus (Пермь) Eryops megacephalus (дл. черепа до 50 см, общ длина до 2 м) Zygosaurus (ср. Пермь) Архегозавр Peltobatrachus pustulatus Грирерпетон
В карбоне возникли лепоспондилы ( Lepospondyli, тонкопозвонковые ). Они имели мелкие размеры и были хорошо приспособлены к жизни в воде. Некоторые из них вторично утратили конечности. Они просуществовали до середины пермского периода. Есть предположение, что они дали начало отрядам современных амфибий — хвостатых (Caudata) и безногих (Apoda). Hyloplesion Положение Тонкопозвонковых по отношению к другим группам лабиринтодонтов неясно, иногда их даже выделяют в отдельный подкласс. Зубы представителей этой группы не такие, как у других лабиринтодонтов. Существуют и некоторые сомнения по поводу того, образуют ли Тонкопозвонковые нечто филогенетически единое в принципе, или же речь идёт о таксоне, просто вобравшем в себя педоморфные формы и головастиков представителей других групп. Diplocaulus magnicornis
Группа темноспондильных Amphibamidae считается возможным предком современных амфибий. Один из входящих в неё родов, Gerobatrachus (Пермский период) , описанный в 2008 году, может быть транзитной формой между темноспондильными и Anura (то есть, лягушками и жабами) и саламандрами. Он похож и на лягушек, и на саламандр, включая широкий череп, короткий хвост и некрупные зубы. Подробнее происхождение амфибий и рептилий – см. htps: //ru. wikipedia. org/wiki/% D 0%9 B%D 0%B 0%D 0%B 1%D 0%B 8%D 1%80%D 0%B 8%D 0%BD%D 1%82%D 0%BE%D 0%B 4%D 0%BE%D 0%BD%D 1%82%D 1%
Происхождение рептилий Diplovertebron Diadectes, a permian reptiliomorph. Карбон. Остатки наиболее древних пресмыкающихся известны с верхнего карбона (около 300 млн. лет назад). Видимо, в среднем карбоне (320 млн. лет) от Антракозавров (лабиринтодонтов, подобных Diplovertebron) , обособлялись формы, лучше приспособленные к наземному образу жизни. От подобных форм возникает новая ветвь амфибий — сеймуриоморфы ( Seymouriomorpha ), остатки которых обнаружены в верхнем карбоне — средней перми . Пермь. Из верхней перми известны остатки котилозавров ( Cotylosauria ). По ряду признаков они очень близки к лабиринтодонтам. Их череп был в виде сплошной костной коробки с отверстиями только для глаз, ноздрей и теменного органа , шейный отдел позвоночника был слабо оформлен (хотя имеется характерное для современных пресмыкающихся строение первых двух позвонков — атланта и эпистрофея ), крестец имел от 2 до 5 позвонков; в плечевом поясе сохранялся клейтрум — кожная кость, свойственная рыбам; конечности были коротки и широко расставлены.
Cotylosauria , от др. -греч. κοτύλη — «чашка» и σαῦρος — «ящерица» . Известны с середины каменноугольного по триасовый период. Размеры — от мелких насекомоядных проколофонов и средней величины хищников (в том числе водных) до крупных растительноядных парейазавров. сеймуриаморфы , капториноморфы , парейазавры и проколофоны. У большинства котилозавров наблюдаются признаки, характерные для земноводных животных. Наиболее существенные: 1. череп без височных впадин с хорошо развитым теменным отверстием; 2. нёбные зубы; 3. двояковогнутые позвонки; 4. очень короткий шейный отдел; 5. короткие и массивные кости конечностей и их поясов. Проколофоны , лат. Procolophonidae — название вымершего семейства небольших рептилий. Известны по черепам размером около 5 см в диаметре. Вплоть до раннего триасового периода зубы проколофонов были острыми, что указывает на их насекомоядность. Позднее в триасовом периоде зубы стали более широкими, что говорит о переходе на растительную пищу. К концу триасового периода проколофоны вымерли. Недавние находки указывают, что эти животные, возможно, обитали в Антарктике в раннем триасовом периоде, то есть были первыми четвероногими в Антарктике.
Парейаз врыаы ( лат. Pareiasauridae ) — группа тяжёловесных растительноядных пресмыкающихся (или парарептилий ) крупных размеров (от 1, 5 до 4 метров), с коротким толстым туловищем и хвостом. В коже спины и головы этих ящеров формировались окостенения, которые придавали кожной поверхности неровный или бугристый характер. Короткие и очень массивные конечности делали парейазавров весьма неповоротливыми существами. Вероятно, прибрежные животные, которые много времени проводили в воде подобно бегемотам. Время расцвета — конец пермского периода. Scutosaurus karpinskii, 3, 5 м Arganaceras vacanti
Сначала обособились анапсиды ( Anapsida, совр представители — черепахи ) и синапсиды ( Synapsida, совр — крокодилы , клювоголовые , чешуйчатые , птицы ). В структуре черепа анапсид отсутствовали какие-либо впадины. Предком черепах обычно считают пермского эунотозавра ( Eunotosaurus ) — небольшое ящерообразное животное с короткими и очень широкими рёбрами, образующими подобие спинного щита. Синапсиды — рептилии с нижними височными впадинами, ограниченными скуловой, чешуйчатой и заглазничной костями. Уже в позднем карбоне эта группа стала самой многочисленной. В палеонтологической летописи они представлены двумя последовательно существовавшими отрядами: пеликозавров ( Pelicosauria ) и терапсид ( Therapsida ). Период своего расцвета терапсиды пережили задолго до того, как появились первые динозавры , потомками терапсид стали более высокоорганизованные зверозубые ( Theriodontia ). К ним относится, в частности, хищная иностранцевия , напоминающая саблезубого тигра. Зверозубые были многочисленны ещё в раннем триасе , но после появления хищных динозавров они исчезли. Звероподобные представляют большой интерес как группа, давшая начало млекопитающим Эунотозавр — предполагаемый предок черепах, до 20 см
Пеликозавры внешне походили на ящериц; они имели двояковогнутые позвонки и хорошо сохранившиеся брюшные рёбра. Однако зубы их сидели в альвеолах (как у терапсид и млекопитающих ), а у некоторых пеликозавров (например, у сфенакодона ) намечалась слабовыраженная дифференциация зубов. Среди пеликозавров встречались как крупные (длиной 3— 6, 5 м), так и существенно более мелкие (1— 2 м) животные — хищные и растительноядные. В конце карбона и начале перми (300— 270 млн лет тому назад) пеликозавры доминировали в наземных экосистемах. У некоторых сфенакодонтов (например, у диметродона ) и всех эдафозавров на спине имелся гребень ( «парус» ) из кожи, образующей перепонку, которая была натянута на верхние остистые отростки позвонков. Предположительно служил для терморегуляции. Диметродон. Два вида сфенакодонов
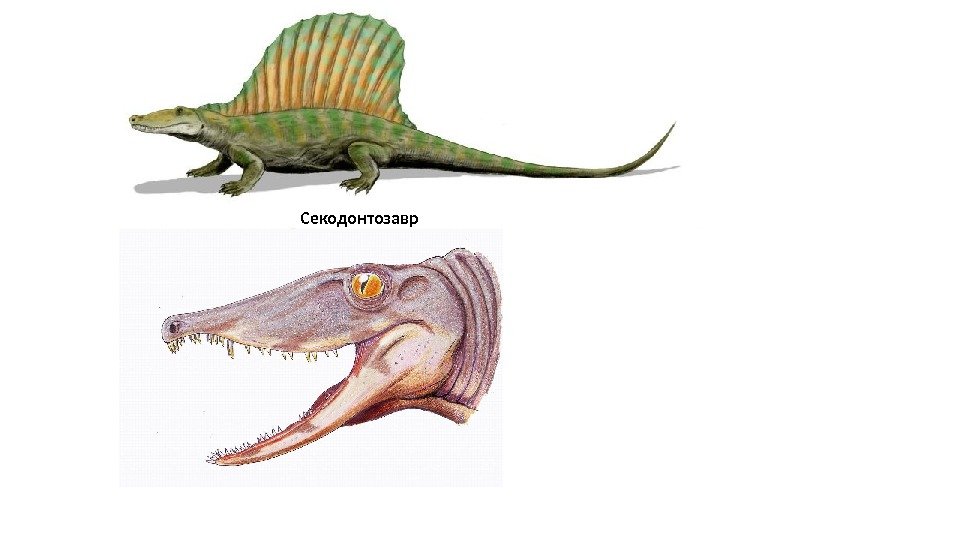
Секодонтозавр
Три совый пер од аы иы (три с) — первый геологический период мезозойской эры. Следует за пермским периодом и аи предшествует юрскому. Начался 250 млн лет назад, кончился 200 млн лет назад. Продолжался, таким образом, около 51 млн лет. Вымирание семенных папоротников. В наземной растительности начали преобладать голосеменные, цикадовые, гинкго и хвойные. Растительный мир суши наследовал черты верхнепермской эпохи. В Триасе исчезли древовидные плауновые и каламитовые, кордаитовые, прапапоротники и большая часть древних хвойных. Были распространены диптериевые папоротники, саговники, беннеттиты, гинкговые, мезофитные хвойные, хвощовые растения. Рептилии: нотозавры, ихтиозавры, плакодонты, склерозавры, талаттозавры (аскептозавр). Появляются новые группы моллюсков (улитки, устрицы). Крупнейшими хищниками становятся водные. В это же время, происходит значительное падение разнообразия позвоночных. На суше развиваются рептилии. Появляются первые теплокровные — яйцекладущие млекопитающие, а также протоавис, по предположению некоторой части палеонтологов, предки птиц. В триасе широкое распространение получают текодонты — быстропередвигающиеся сухопутные рептилии, предки динозавров. Наряду с лабиринтодонтами, котилозаврами, существовавшими ещё в пермское время, появились характерные для мезозоя архозавры, ихтиозавры, черепахи, в конце Триасового периода — костистые рыбы. Среди морских беспозвоночных преобладали цератиты (головоногие моллюски), появившиеся в конце перми и вымершие в конце Триасового периода; характерны пелециподы, гастроподы, наутилоидеи.
Иностранцевия. Терапс дыиы ( лат. Therapsida), ранее известные под названием «звероподобные рептилии» , — отряд класса синапсид (по другим классификациям — инфракласс или даже подкласс). Возникли как потомки пеликозавров в раннем пермском периоде. Конкретные переходные формы между сфенакодонтными пеликозаврами и терапсидами неизвестны. Возможной переходной формой долгое время считался тетрацератопс из ранней перми Техаса. Не исключено, что реликтами таких переходных форм могут быть мелкие малоизвестные никказавры из средней перми Приуралья. Большинство терапсид вымерло во время пермской катастрофы , немногочисленные представители дожили до триасового периода , после чего почти все они вымерли окончательно. Исключение составили цинодонты , входившие в группу териодонтов , — от них произошли млекопитающие. Цинодонты дожили до раннего мелового периода. Не исключено также выживание дицинодонтов в Австралии вплоть до начала меловой эпохи. Тетрацератопс. Inostrancevia (Theriodontia)
Терапс дыиы (лат. Therapsida)
Склероторакс — раннетриасовый лабиринтодонт Мастодонзавр — гигантский среднетриасовый лабиринтодонт Плакод нты (лат. Placodontia) — отряд ископаемых морских ои пресмыкающихся надотряда Sauropterygia, живших в триасовом периоде на территории Европы, Азии и Африки. Название означает «плоские зубы» . По некоторым представлениям плакодонты являются первыми пресмыкающимися, перебравшимися в воду около 215 млн. лет назад. Их рацион составляли разнообразные моллюски и ракообразные, которых они отрывали от морского дна (в этом им помогали ковшевидные челюсти, снабжённые мощными мышцами, и шесть конических острых передних зубов). А 14 тупых, плоских коренных зубов позволяли перемалывать раковины, превращая их в кашицу.
Nothosaurus mirabilis, a Nothosaur from the Triassic of Europe, North Africa and Russia, pencil drawing
Ящерот зовыеаы (лат. Saurischia) — один из двух главных отрядов динозавров. Отличаются тем, что лобковые кости первично наклонены вперёд (позднее у некоторых представителей — сегнозавры, дромеозавры и другие — лобковые кости отклоняются назад). Считается, что от этой группы произошли птицы. К ящеротазовым относятся: Крупные растительноядные зауроподы , ходившие на четырёх ногах; В среднем более мелкие плотоядные тероподы , передвигавшиеся на двух задних конечностях (ранняя юра — поздний мел). Этот отряд в свою очередь разделяется на две основные группы: Тероподы, или звероногие (Theropoda), к которым относятся двуногие, плотоядные динозавры. Завроподоморфы (Sauropodomorpha).
Птицетазовые (лат. Ornithischia) — один из двух отрядов динозавров. Несмотря на своё название, не они, а ящеротазовые динозавры по одной из версий были предками птиц. Были травоядными (за исключением ранних всеядных видов), меньших размеров, чем ящеротазовые зауроподы. Кроме самых примитивных, имеют приспособления к питанию растительной пищей: сложный кишечник и щёки. Ранние представители были двуногими, позже птицетазовые повторно освоили хождение на четырёх конечностях. В ходе эволюции у поздних представителей отряда концевые фаланги пальцев стали более плоскими и приобрели вид копыт. У многих развились приспособления для защиты от хищников: броня, рога, шипы. Таз имел четырёхлучевое строение. Лобковая кость имела вырост, направленный назад вдоль седалищной кости (у ящеротазовых кости были практически перпендикулярны). Лобковая кость часто образует новый вырост, направленный вперёд параллельно позвоночнику. Подобное строение таза развивалось у динозавров независимо трижды: у птицетазовых, у надсемейства Therizinosauroidea и семейства Dromaeosauridae. Последнее и является наиболее вероятным предком птиц.
Согласно реконструкции Чаттерджи длина протоависа составляла около 35 см. Хотя он жил за 60— 75 млн лет до археоптерикса, строение его скелета ближе к современным птицам. Из-за наличия зубов на клюве в нём предполагают хищника, размещение глаз на передней части черепа свидетельствует о ночном или сумеречном образе жизни. По мнению Чаттерджи, Протоавис был адаптирован к полету, на его костях присутствуют особые отметины — точки прикрепления перьев. Однако, по мнению других палеонтологов, небольшое количество окаменелостей не позволяет определить степень способности протоависа к полёту, более того, детальный их анализ не даёт никаких свидетельств наличия у него перьев. Протоавис (лат. Protoavis) — название рода, предложенное для окаменелостей позднего триаса (225— 210 млн лет), обнаруженных на северо-западе Техаса (США). В 1991 году протоавис был описан палеонтологом Шанкаром Чаттерджи (Sankar Chatterjee) как примитивная птица, однако эта интерпретация была подвергнута сомнению. Если интерпретация Чаттерджи верна, то протоавис является древнейшей из известных птиц, на 60— 75 млн лет старше археоптерикса. Выделяется единственный вид Protoavis texensis ( «первоптица штата Техас» ). Принятие птичьего статуса протоависа подразумевает, что становление класса птиц произошло не позднее конца триасового периода. Следовательно, предков птиц следует искать среди триасовых архозавров. Это не согласуется с наиболее распространённой точкой зрения на филогению ранних птиц, по которой самыми близкими родственниками птиц являются троодонтиды и дромеозавриды — два семейства хищных динозавров. Если данная филогения верна, то эти семейства должны были возникнуть в триасовом периоде, однако наиболее ранние окаменелости представителей этих семейств относятся к раннему мелу.
Тероподная гипотеза Гипотеза первоначально была выдвинута Томасом Генри Хаксли в XIX веке. В 70 -х годах XX века её возродил Джон Остром. Основываясь на сравнении пересмотренной им остеологии археоптерикса и тероподных динозавров (в особенности описанного им дейнониха), Остром заключил, что ближайшими родственниками археоптерикса являются целурозавры из семейства дромеозаврид. Впоследствии выводы Острома были подтверждены кладистическими исследованиями, обнаружением экземпляров целурозавров с отпечатками перьев, сравнением микроструктуры яичной скорлупы и репродуктивной биологии целурозавров и птиц. Сегодня тероподная гипотеза принимается большинством палеонтологов. Современная филогения птиц и наиболее близких к ним групп тероподов выглядит следующим образом: Tetanurae Coelurosauria — Целурозавры Maniraptoriformes Maniraptora Eumaniraptora Deinonychosauria (Dromaeosauridae + Troodontidae) Aves — Птицы Archaeopterygiformes Pygostylia Neornithes Однако после открытия четырёхкрылого оперённого динозавра в Китае основная идея гипотезы — «с земли вверх» — стала всё чаще подвергаться острой критике. При этом надо заметить, что остальные находки оперённых динозавров в Китае довольно хорошо вписываются в концепцию «с земли вверх» . Целурозавры (лат. Coelurosauria — полохвостые) — самая многочисленная группа динозавров-тероподов, живших около 167— 65, 5 миллионов лет назад. Целур (ср. Юра)
Тероп дыоы (лат. Theropoda, от др. -греч. θήρ «зверь» , и πούς «нога» , — звероногие), или хищные динозавры — один из подотрядов ящеротазовых динозавров. Все тероподы перемещались на двух ногах, большинство — хищники, реже всеядные или растительноядные ( теризинозавры , орнитомимиды ). Большинство учёных с недавних пор стали относить к тероподам и современных птиц, таким образом птицы из потомков динозавров стали относиться к одному из подотрядов. Кисть обычно трёхпалая, стопа — обычно четырёхпалая с опорой на 2— 4 -й пальцы. Крупнейшим среди теропод был спинозавр (более 17 метров). Тероподы составляли около 37 % всех известных родов динозавров, среди последних находок Nemegtomaia (2004), Гуанлонг (2006), Limusaurus (2009) и др. Эволюционное происхождение перьев можно проследить начиная с карнотавра , покрытого многочисленными бугорками, похожими на фолликулы. Следующий этап виден у синозавроптерикса и Dilong paradoxus , покрытых волокнистым пухом. И наконец у каудиптерикса, Protarchaeopteryx, синорнитозавра и микрораптора можно наблюдать настоящие перья.
Манирапторы (лат. Maniraptora, от греческих слов «рука» и «хватать» ) — клада архозавров, включающая целурозавров , по своим характеристикам примыкающих к птицам тесней, чем к орнитомимозаврам. Оригинальное определение клады дано в 1986 году Жаком Готье. Наиболее древние известные останки манирапторов относятся к раннему юрскому периоду , а, поскольку кладистический анализ включает в эту группу птиц, это единственная группа динозавров, дожившая до современности. Protarchaeopteryx
Предками млекопитающих были древние палеозойские рептилии, еще не утратившие некоторых черт строения амфибий: кожные железы, расположение сочленений в конечностях (между голенью и предплюсной и между предплечьем и запястьем) и, быть может, двойной затылочный мыщелок. Такой предковой группой считают подкласс звероподобных (Theromorpha). Особенно близок к млекопитающим один из отрядов этого подкласса — отряд зверозубых (Theriodontia), у представителей которого зубы были дифференцированы на те же категории, что и у млекопитающих, т. е. на резцы, клыки и коренные, и сидели они в альвеолах. Имелось вторичное костное нёбо — признак весьма характерный для млекопитающих. Квадратная кость и сочленовная кость нижней челюсти были сильно уменьшены. При этом сочленовная кость не была приращена к нижней челюсти. Зубная кость, наоборот, была очень крупной. Здесь следует учесть, что у млекопитающих нижняя челюсть состоит только из зубной кости, а сочленовная кость превращена в одну из слуховых косточек — молоточек. Такие известные высшие зверозубые, как иностранцевия (из пермских отложений на Северной Двине) и циногнатус (из триасовых отложений Южной Африки), не могли быть предками зверей, так как были высокоспециализированными и при этом крупными (2— 3 м длиной) животными. Малые размеры древнейших млекопитающих дают основание искать предков зверей среди более примитивных и мелких зверозубых. В этом отношении представляет интерес иктидозавр (из верхнего триаса Южной Африки). Это было животное размером с крысу, скелет которого удивительно сходен с таковым млекопитающих; только нижняя челюсть у него состояла из нескольких костей. Очевидно, от таких мелких зверозубых и произошли млекопитающие. Зверозубые просуществовали недолгое время, лишь до верхнего триаса.
Териодонты или зверозубые ящеры — вымершая группа (подотряд или отряд) зверообразных, одна из трёх главных групп терапсид. К териодонтам принадлежал отряд цинодонтов — предки современных млекопитающих. Сюда относят также горгонопсов и тероцефалов. Зверозубые ящеры имеют сходство с млекопитающими в строении черепа, позвоночника и конечностей, а также в делении зубов на клыки, резцы и коренные. В отличие от других ящеров, их ноги располагались под телом, приподнимая его над землёй. Циногнатус Эоарктопс Гофмейерия — тероцефал позднепермской эпохи
Находки древнейших млекопитающих приурочены к юрским отложениям. Это были разнообразные и в ряде случаев высокоспециализированные животные; следовательно , возникновение млекопитающих надо отнести к более раннему времени — к триасу. С конца этого периода и до первой половины третичного времени существовали так называемые многобугорчатые. Это были мелкие, размером с крысу, зверьки, и только немногие достигали размеров сурка. Свое название они получили в связи с наличием на коренных зубах многочисленных бугорков. Клыков у них не было, а резцы развивались, наоборот, очень сильно. Многобугорчатые представляли специализированных растительноядных зверей, и их нельзя считать прямыми предками последующих групп млекопитающих. Лишь предположительно допускают, что наиболее ранние формы могли дать начало однопроходным. Основанием для такого предположения служит сходство строения их зубов с зубами зародыша утконоса. В отложениях с середины юры и до мела обнаруживаются многообразные трехбугорчатые , открывшие подлинную историю класса млекопитающих. Зубная система у них менее специализированная, чем у многобугорчатых; зубной ряд сплошной. Это были мелкие зверьки, близкие к насекомоядным; питались они частью животной, частью растительной пищей. Трехбугорчатые , в частности пантотерии , — наиболее вероятные предки сумчатых и плацентарных.
Многобуг рчатыеоы (лат. Multituberculata) — вымерший отряд древних млекопитающих (инфракласс аллотерии (лат. Allotheria), отряд Multituberculata). Название происходит от формы коренных зубов, на которых были расположены многочисленные бугорки. Внешне похожие на грызунов, они возникли в середине юрского периода и вымерли в раннем олигоцене. Были распространены только в Северном полушарии (праматерик Лавразия). Skull of Ptilodus mediaevus, a paleocene multituberculate
Сумчатые появились, видимо, в начале мелового периода. Однако первые ископаемые остатки их известны из верхнемеловых отложений Северной Америки. Это были мелкие зверьки типа опоссумов с бугорчато-режущими коренными и крупными клыками. В отложениях нижнетретичного времени они известны и в Евразии. Таким образом, родиной сумчатых следует считать северное полушарие. Однако еще до конца третичного времени они были вытеснены более высокоорганизованными плацентарными млекопитающими и в настоящее время сохранились только в Австралии, Новой Гвинее, Тасмании, Южной Америке, т. е. в южном полушарии, и лишь отчасти в Северной Америке (1 вид) и на острове Сулавеси (1 вид). Только в Австралии, куда из плацентарных проникли лишь рукокрылые и мышевидные грызуны , сумчатые дали большое разнообразие форм зверей, приспособленных к существованию в весьма различной жизненной обстановке. Высшие, плацентарные млекопитающие , как и сумчатые, произошли от трехбугорчатых в начале мелового периода и развивались самостоятельно, параллельно с сумчатыми. Первые ископаемые остатки, относимые к насекомоядным , известны из верхнемеловых отложений Монголии. Плацентарные образовали 31 отряд, из которых 17 распространены в настоящее время, а 14 полностью вымерли.