Рыбы-гаструла-16.ppt
- Количество слайдов: 39



Карта презумптивных зачатков для глубоких клеток стадии поздней бластулы


Распределение предшественников по анимальновегетативной оси поздней бластулы - ранней гаструлы Эктодерма – самое анимальное положение Мезодерма – по краю бластодермы Энтодерма – смешанное с мезодермой по краю бластодермы в дорсальной и латеральной областях Распределение эктодермы по дорсовентральной оси Эпидермис – анимальновентральное положение Нервная система: Передний и средний мозг – анимальное и дорсальное положение Задний мозг – ближе к краю бластодермы и латерально Спинной мозг – ближе к краю бластодермы и вентрально Распределение мезодермы по дорсовентральной оси Осевая мезодермы (прехордальная пластинка, хорда) - самое дорсальное положение Параксиальная мезодерма (сомиты) - дорсолатеральное положение Промежуточная мезодерма (пронефрос) – латеральное положение Боковая пластинка мезодермы (кровь) вентральное положение Распределение энтодермы по дорсовентральной оси Дорсально и латерально.

Типы морфогенетических движений, составляющих гаструляцию: • радиальная интеркаляция • инволюция • ингрессия • конвергенция/растяжение • медиолатеральная интеркаляция




Вьюн, начало гаструляции. Слева – вид с латеральной стороны, справа – вид с анимального полюса
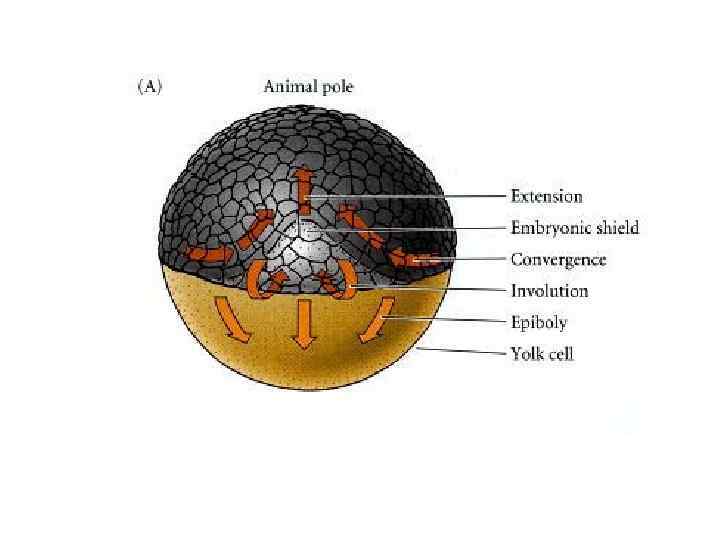

Гаструляция у данио
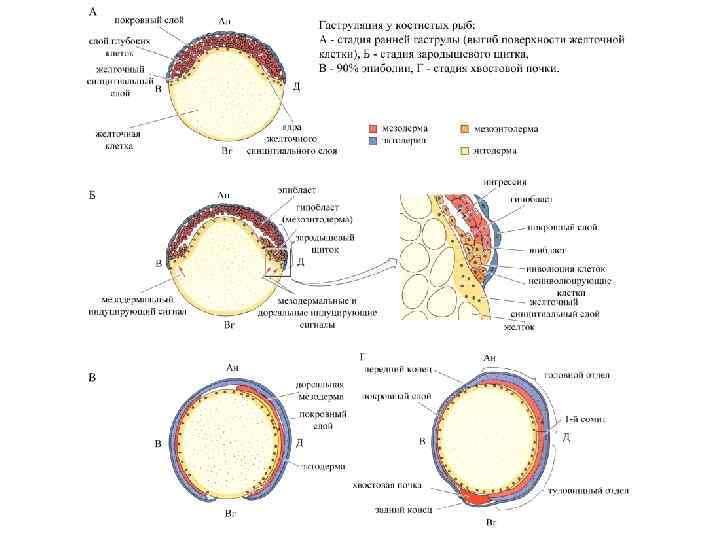

Гаструляция и нейруляция у данио

Гаструляция и нейруляция у карпа

Глубокие клетки на поверхности внутреннего желточного синцитиального слоя

Органы локомоции глубоких клеток

Глубокая клетка на внутренней поверхности эпибласта
 клетки на поздней бластуле – ранней гаструле Зародыши на стадии")
NEM (noninvoluting endocytic marginal) клетки на поздней бластуле – ранней гаструле Зародыши на стадии высокой эпителиальной бластулы метили SYTO-11. Верхний ряд – оптика Номарского, нижний ряд – конфокальная микроскопия. (А, В) 30% эпиболии, начало выгиба желточной клетки. (C, D) 55% эпиболии, зародышевое кольцо и начало образования зародышевого щитка. (E, F) 65% эпиболии, удлинение зародышевого язычка. Меченые клетки на стадии поздней бластулы локализуются на будущей дорсальной стороне в месте формирования зародышевого щитка. В ходе гаструляции кластер NEM клеток не инволюирует и остается на дорсальной стороне зародыша спереди от края глубоких клеток бластодермы.

Экспрессия некоторых мезодермальных маркеров

50% Экспрессия goosecoid на разных стадиях эпиболии 75% вид с дорсальной стороны вид сбоку 100%

Экспрессия ntl в ходе развития
ectodermal, mesodermal and endodermal progenitor cells")
Fig. 1. Expression patterns of marker genes for (neuro)ectodermal, mesodermal and endodermal progenitor cells within the shield region at the onset of gastrulation (60% epiboly). Sagittal (A-D) and frontal (E-H) sections of the shield region demarcating the expression domains of goosecoid (gsc; A, E), gata 5 (B, F), notail (ntl; C, G) and forkhead 3 (fkd 3; D, H). The horizontal lines in A-D indicate the level where the frontal sections in E-H were taken. (I-L) Orientation of the embryos (I, K) and schematic representations (J, L) of the shield region (boxed areas in I, K), representing sagittal (I, J) and frontal (K, L) views of the shield as shown in panels A-D and E-H, respectively. Please note that number of cell layers depicted in the schematic diagrams is lower than in the real embryo (for details about cell layer numbers, see Results). epi, epiblast; hyp, hypoblast; evl, enveloping cell layer; ysl, yolk syncytial layer. Scale bar in A: 100 µm for A-H.

Детерминация дорсовентральной оси у золотой рыбки

Перемещение дорсальных детерминант с помощью микротрубочек в ходе ооплазматической сегрегации


Желточный синцитиальный слой – Ньюкуповский центр у костистых рыб. Трансплантация желточной клетки. a. Схема опыта. Меченную биотином желточную клетку зародыша-донора без бластодермы пересаживали на анимальную поверхность бластодермы немеченного зародыша на той же стадии развития (средняя бластула). Анализировали экспрессию мезодермальных маркеров в рекомбинантных зародышах методом in situ гибридизации. b. Зародыш-хозяин на стадии средней бластулы. Желточный синцитиальный слой и клетки бластодермы плотно прилежат друг к другу. c. Желточная клетка зародыша-донора. Часть краевых бластомеров не удалена, но установлено, что при трансплантации эти клетки не индуцируют эктопическую экспрессию ntl.

Трансплантированная желточная клетка индуцирует экспрессию ntl и gsc в бластодерме хозяина a. Экспрессия ntl в зародыше с трансплантированной желточной клеткой. Трансплантация на стадии перехода от 512 клеток к высокой бластуле, фиксация на стадии 50 % эпиболии. Эктопическая экспрессия (стрелки) видна в виде кольца вокруг трансплантированной желточной клетки. b. Гистологический срез зародыша с трансплантированной желточной клеткой. Эктопическая экспрессия ntl (стрелка) видна в клетках зародыша-хозяина (неокрашенные) около пересаженной желточной клетки (окрашенная). c. Нет эктопической экспрессии ntl при трансплантации неоплодотворенной яйцеклетки. d. Экспрессия ntl в нормальном зародыше. e. Экспрессия gsc в зародыше с трансплантированной желточной клеткой. Локальная экспрессия обнаруживается около пересаженной желточной клетки (стрелка). f. Экспрессия gsc в нормальном зародыше на стадии 50 % эпиболии.

squint – ген nodal-related белка bozozok – аналог гена siamois chordino – аналог chordin

Формирование сигнальных центров гаструлы у костистых рыб

Зародышевый щиток – Шпемановский организатор у костистых рыб

 Трансплантация дорсального (красный) и вентрального (зеленый) фрагментов")
Организационные свойства краевой зоны ранней гаструлы (A) Трансплантация дорсального (красный) и вентрального (зеленый) фрагментов края бластодермы зародыша на стадии ранней гаструлы (зародышевый щиток) на анимальный полюс и на вентральную сторону зародыша на стадии поздней бластулы. (B) Трансплантация дорсального фрагмента на вентральную сторону вызывает формирование дополнительного зародыша с головными и туловищными структурами. (C) Трансплантация дорсального фрагмента на анимальный полюс вызывает образование дополнительной короткой хорды (маркер – Shh). (D) Трансплантация вентрального фрагмента на анимальный полюс индуцирует образование дополнительного хвоста. (E-H) Трансплантация латерального фрагмента (оранжевый) индуцирует образование частичной вторичной оси с переднетуловищными и заднеголовными структурами (кончик стрелки на F – слуховые пузырьки в первичной и вторичной оси). Вторичная ось растет от первичной головы (G) и имеет материал донора (GFP) и немеченные клетки хозяина (H, кончик стрелки – вторичный слуховой пузырек). (I, J) in situ гибридизация маркеров krox 20 и cdh 17. Стрелки – вторичные оси; кончик стрелки – проток пронефроса. J. -D. Fauny et al. , Development 136, 3811 -3819 (2009)
 Пересадки разных фрагментов на анимальный полюс. Зародыш-донор")
Организационные свойства краевой зоны ранней гаструлы (А) Пересадки разных фрагментов на анимальный полюс. Зародыш-донор слева, вид сверху; зародыш-хозяин справа, вид сбоку. 0° - DM, 45° - DLM, 90° - LM, 180° - VM. (J) Частота встречаемости различных структур после пересадки фрагментов различных участков края бластодермы. Показана колинеарность между дорсовентральным положением пересаженных фрагментов края бластодермы гаструлы и частями зародыша вдоль переднезадней оси, которые они могут индуцировать. J. -D. Fauny et al. , Development 136, 3811 -3819 (2009)


Ст. 8 Ст. 9 Ст. 10 Ст. 11 Ст. 12 Д В Л А


Solnica-Krezel, 2005

Градиент BMP регулирует конвергенциюрастяжение в ходе гаструляции

Фенотипический класс Дорсальный организатор, класс 1 а; уменьшение дорсальных и передних структур Локус Генный продукт Молекулярная роль Фенотип bozozok(boz) гомеодомен транскрипционный репрессор отсутствие или редукция хорды и прехордальной пластинки; редукция передних и вентральных нейральных структур оne-eye pinhead (oep) EGF-CFC Nodal-корецептор Отсутствие производных прехордальной пластинки; циклопия, недостаточность вентральных структур ЦНС cyclops (cyc) Nodal-сигнал Редукция прехордальной пластинки; циклопия и недостаточность вентральных структур ЦНС Schmalspur (sur) FAST 1/Fox H 1 Транскрипционный фактор Редукция прехордальной пластинки; циклопия и недостаточность вентральных структур ЦНС Floating head (flh) Транскрипционный фактор Отсутствие хорды, уменьшение дифференцировки эпифиза No tail (ntl) Транскрипционный фактор Аномалии дифференцировки в туловище, отсутствие хорды в хвосте; редукция хвоста chordin BMP-ингибитор Разная степень умножения хвостового плавника, увеличение вентральных и задних структур (кровь, пронефрос); уменьшение нейроэктодермы и сомитной мезодермы, укорочение тела Дорсальный организатор, класс Iб; осевая мезодерма присутствует, уменьшение вентральных и задних структур chordino (din) Вентральный организатор, класс II; уменьшение вентральных и задних структур swirl (swr) BMP 2 b BMP-сигнал somitabun (smb) Smad 5 транскрипционный фактор Различная степень редукции вентральных и задних структур, отсутствие хвостового плавника, прогрессивное уменьшение хвоста и туловища; редукция вентральных мезодермальных структур (кровь, пронефрос); распространение сомитов и нейроэктодермы snailhouse (snh) BMP 7 BMP-сигнал удлиненная форма гаструлы lost-a-fin (laf) ALK 8 Type I BMP-рецептор mini fin (mif) tolloid металлопротеаза для chordin ogon (ogo) mercedes (mes) piggytail (pgy)

Рыбы-гаструла-16.ppt