Презентация БПИ-3 Для доклада.ppt
- Количество слайдов: 36



Как возникла жизнь? § Интерес к проблеме существует с момента появления человека на Земле, однако реально этой проблемой стали заниматься после публикации теории А. И. Опарина и Дж. Холдейна о происхождении жизни в 1920 30 годы. Автор не считает целесообразным рассматривать эти теории как и тысячи других работ пытающихся ее подтвердить, так как прошедшие 85 лет показали их полную несостоятельность. § Нет смысла рассматривать и любую теорию из серии гипотез «панспермии» о заносе жизни на Землю из космоса метеоритами, пришельцами, «спорами жизни» и т. д. , так как проблема возникновения жизни остается независимо от места ее возникновения. § Нет смысла рассматривать любую теорию происхождения жизни не учитывающую современные знания о структуре, функции и механизмах воспроизведения клеток.

Клетки это единый жидкокристаллический, гомеостатический комплекс, состоящий из десятков тысяч разнообразных молекул: нуклеиновых кислот, белков, углеводов и липидов, работающих как единый механизм, управляемый едиными биохимическими самовоспроизведению, генетический информации, код, процессами, имеющий сходные независимо способный универсальный механизмы от к реализации уровня организации прокариотического или эукариотического типа и т. д. Поэтому все попытки объяснить происхождение жизни через первоначальное формирование фрагментов ДНК, РНК, белков или других структур с последующей эволюцией изначально обречены на провал. Времени на формирование клеток таким путем просто не существует ни на Земле, ни во Вселенной. Всегда надо помнить, что в основе появления жизни, в любой форме, лежат половые клетки, изначально являющиеся жидкими кристаллами с набором всех необходимых молекул для создания будущего организма, независимо от уровня организации прокариотического или эукариотического типа.
 Проблема возникновения жизни волнует человеческий разум не")
§ § КАК ВОЗНИКЛА ЖИЗНЬ (СОКРАЩЕННЫЙ ВАРИАНТ) Проблема возникновения жизни волнует человеческий разум не одно тысячелетие. Вопрос о происхождении жизни из неорганических веществ в серьезных научных работах до 20– 30 х гг. прошлого столетия не рассматривался. В 20– 30 е гг. ХХ в. А. И. Опарин и Дж. Холдейн предложили принципиально новый подход к решению этой проблемы. Они выдвинули гипотезу постепенной эволюции простых органических соединений, которые образовались на первобытной Земле из газов, составляющих атмосферу, под воздействием различных источников энергии. § Теория Опарина–Холдейна стимулировала проведение боль шого числа экспериментов, иллюстрирующих возможные пути обра зования и эволюции органических соединений в условиях примитив ной Земли. Появившиеся органические соединения, по мнению авторов самоорганизовываться таких в исследований, начали коацерват ные капли, пузырьки, микросферы, протоклетки, липосомы, люминосферы, коацерваты в коацервате, каталитические каскады и т. д. Это была смесь асимметричных мономерных или коротких полимерных органических молекул, из которых предстояло возникнуть будущей протоклетке путем самопроизвольной организации.

По поводу выше названных теорий лауреат Нобелевской премии Даниел Карлтон Гайдузек так оценил этот этап исследований: «Наиболее серьезные ученые, занимающиеся теорией происхождения жизни, уже давно отказались от всей этой чепухи насчет того, что в океане мол, был бульон из пуринов, пиримидинов и аминокислот и его активировало ультрафиолетовое излучение» . Автор работы давно придерживается аналогичной точки зрения. Нет смысла рассматривать и любую теорию из серии гипотез «панспермии» о заносе жизни на Землю из космоса метеоритами, пришельцами, «спорами жизни» и т. д. , так как проблема возникновения жизни остается независимо от места ее возникновения. На сегодняшний день мы имеем около 4000 работ, посвященных этой проблеме, среди них десятки монографий. Однако ни одна из них не приближает к ответу на интересующий нас вопрос. Их абсолютное большинство не учитывает современные знания о том, что клетки это жидкокристаллический, гомеостатический, апериодический комплекс, способный к самовоспроизведению, состоящий из десятков тысяч разнообразных углеводов и липидов, молекул нуклеиновых работающих как кислот, единый белков, механизм, управляемый едиными биохимическими процессами, имеющие универсальный генетический код, близкие механизмы реализации информации, сходные параметры основных структур ДНК, РНК, белков, углеводов, липидов независимо от уровня их организации, и

§ Ключевые вопросы проблемы происхождения, такие как возникновение матричного механизма и универсального гене тическогокода, остаются без ответа. В лучшем случае их автоматически переводят в плоскость эволюции, в худшем божественного – рассматривают происхождения. Но все в аспекте прекрасно понимают, что ни первое, ни второе нельзя расценивать как ответ на вопрос, по скольку для привлечения эволюции необходим исходный, самовос производящийсяматериал, проблема получения которого и является основополагающей в теории происхождения жизни. Получаемые вероятностные оценки возникновения системы репликации, транскрипции или трансляции нельзя рассматривать даже несерьезно ( 10– 225– 10– 800. ) § Попытка решить проблему через синтез асимметричных фрагментов органических молекул и их последующую эволюцию не представляется возможной, учитывая структуру, организацию и принципы функционирования клеток. Поэтому все попытки объяснить происхождение жизни через первоначальное формирование фрагментов ДНК, РНК, белков или других структур с последующей эволюцией изначально обречены на провал. Времени на формирование клеток таким путем просто не существует ни на Земле, ни во Вселенной.

Никто из авторов даже не пытается объяс нить, почему вездесущая эволюция снабдила эукариотические клетки разным числом хромосом, информативная часть которых составляет в лучшем случае несколько процентов, и тем не менее не избавилась от остальной части, как это имеет место у прокариот. Откуда взялись десятки уникальных белков для рибосом, функция которых синтези роватьэти и другие белки. Обычный ответ: появились в ходе эволю ции. Жаль, что сама эволюция этого не знает. Авторы работ 30– 50 х гг. прошлого века ответить на эти вопросы даже не пытались, поскольку такой информации о клетках тогда не существовало. Однако мало что изменилось и в последующие десятилетия, вплоть до сегодняшнего дня, хотя знания о строении, функционировании и воспроизведении клеток и их систем выросли многократно. Необходимо помнить, что в основе появления жизни, в любой форме, лежат половые клетки, изначально являющиеся жидкими кристаллами с набором всех необходимых молекул для создания будущего организма, независимо от уровня организации прокариотического или эукариотического типа. Поэтому фрагментарное рассмотрение возникновения протоклеток исключено. Также, как исключено усложнение живых систем при отсутствии изначально необходимой генетической информации.

§ Исходя из сегодняшних знаний о структуре клетки и механизмах ее воспроизведения, проблему возникновения жизни надо рассматривать через проблему возникновения клетки, а не ее фрагментов. Клетка – это сотни тысяч и миллионы молекул, сформировавших единый жидкокристаллический комплекс. Отдельные фрагменты данного кмплекса, в том числе вирусы или фаги, не являются жизнеспособными. Поэтому, ответив на вопрос, каким образом и почему клетка является апериодическим, гомеостатическим, самовоспроизводя щимся жидким кристаллом, сможем понять, как возникла жизнь. Пока на поставленные вопросы ответа нет.

Теоретически возникновение клетки может быть следствием различной последовательности событий. Мы же остановимся на событиях, которые реально, одномоментно могли привести к образованию протоклетки составляющими структурами со всеми ее органо минеральными в результате их специфической агрегации в соответствии с физико химическими законами , путем диффузии свободных радикалов и ионов газовой фазы в кристаллическую решетку. Полученная протоклетка, как и ее прародительница (матрица минералов), самовоспроизведению оказалась за трансляционного аппарата. счет способной к транскрипционно


Посвящается памяти В. В. Чернобровкина ПРОИСХОЖДЕНИЕ ПРОТОКЛЕТОК И ИХ СТРУКТУРНЫХ ЭЛЕМЕНТОВ НА ОСНОВЕ АПАТИТА И СОКРИСТАЛИЗУЮЩИХСЯ С НИМ МИНЕРАЛОВ § § Костецкий Э. Я. Отдел Биохимии и Биотехнологии, Дальневосточного Государственного университета. Владивосток, 690950, Октябрьская 27. Факс: 7 4232 429510. Е mail: kostetsky@nt. pin. dvgu. ru § «Теория оказывается тем более впечатляющей, чем проще ее предпосылки, чем значительнее разнообразие охватываемых ею явлений и чем шире ее применимость» § А. Эйнштейн § «Мир слишком совершенен, чтобы случайно возникнуть из хаоса»

§ Прежде чем мы перейдем к рассмотрению теории возникновения протоклеток автор предлагает в форме вопросов остановиться касающихся структуры, на ключевых свойств и обобщениях особенностей, присущих всем живым клеткам про или эукариотического типа. Эти обобщения помогут более осмысленно воспринимать изложенную ниже теорию. § 1. Почему все клеточные элементы и целые организмы являются жидкокристаллическими гомеостатическими структурами? § 2. Почему клетки каждого таксона являются целостной (неде лимой) системой со своими индивидуальными особенностями? Пере несение отдельных элементов клетки в другие, особенно филогенети чески далекие таксоны, как правило, невозможно. 3. Почему в ДНК эукариот имеются уникальные, умеренно повторяющиеся и сателлитные (балластные) последовательности, не несущие никакой информации? Нетранслируемые последовательности достигать 85 % и, вероятно, более. ДНК могут

4. Почему генетический код универсален? 5. Почему за универсальностью генетического кода, сходством принципов организации клеточных структур, метаболизма, набора клеточных элементов у разных таксонов не следует универсальность элементов, дешифрующих синте тазы, р. РНК, осуществляющих код (т. РНК, ДНК полимеразы метаболизм, структурных и ААт. РНК т. участвующий д. ), в формировании клеточных структур, и т. д. ? 6. Почему в структуре белков, выполняющих различные функ ции, имеются сходные блоки аминокислотной последовательности? 7. Почему в клетках присутствуют специфические белки для ио новкальция, фосфора, магния, стронция, кремния и др. ? 8. Почему ионы Са 2+ и РО 43 являются ключевыми элементами во всех живых системах?

§ 9. Почему в состав органической составляющей клеток входят преимущественно углерод, водород, азот, кислород? § 10. Почему ДНК имеет строго определенные параметры (диа метр – 18 Å, расстояние между парами оснований – 3, 4 Å), опреде ленную комплементарность (А=Т и Г Ц), хотя запрета на комплемен тарность между любыми другими парами нет; почему фосфодиэфир ные связи в ДНК и РНК только между 3' и 5' положениями рибозы, хотя запрета на другие положения нет? § Суть или причины возникновения перечисленных особенностей попытаемся понять в ходе дальнейшего изложения работы. § Обращение автора к созданию теории происхождения жизни произошло в начале 70 х гг. прошлого века. Наше внимание было об ращено на следующие свойства апатита, позволившие предположить его особую роль биомолекул. в процессе абиогенного синтеза

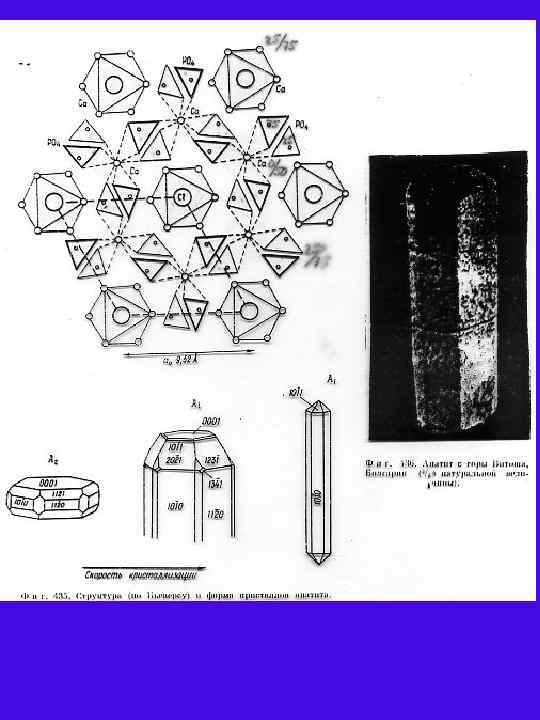

Д Б В Г Рис. 1. Б, В, Г - Cтруктура апатита по Н. В. Белову [Gilinskaya, Scherbacova 1975]: Б - призмы с катионами Са II; В - призмы с катионами Са I; Г анионы РО 4. Д - Апатит с горы Витоша, Болгария (4/5 натуральной величины). Числа указывают доли от высоты элементарной ячейки.
 построена по данным рентгеноструктурного анализа")
А Рис. 4. А – Структура апатита (вид сбоку) построена по данным рентгеноструктурного анализа [Kay et al. 1964; Sudarsanan, Young 1969]; А, Б – компьютерная графика.
 с указанием положения двух пар оснований ДНК и")
Рис. 2 Структура апатита (вид сверху) с указанием положения двух пар оснований ДНК и фрагментом пептидной цепи (в центре) [Kostetsky 1981; 1999].
 с указанием положения четырёх пар оснований ДНК и")
Рис. 3 Структура апатита (вид сверху) с указанием положения четырёх пар оснований ДНК и фрагментом пептидной цепи (в центре) [Kostetsky 1981; 1999].
 построена по данным рентгеноструктурного")
А Б Рис. 4. А – Структура апатита (вид сбоку) построена по данным рентгеноструктурного анализа [Kay et al. 1964; Sudarsanan, Young 1969]; Б – Структура апатита (вид сбоку) с цепями ДНК введёнными в его состав. А, Б – компьютерная графика.
. Для наглядности")
Рис. 5. Структура связей в комплементарных цепях Ап ДНК (Рис. 2 Б). Для наглядности основания даны под небольшим углом. син-, анти- угол вращения вокруг гликозидной связи С 1` N ; g торсионный угол вращения вокруг С 4` С 5` связи имеет an, +ск, -ск конформацию; С 2` эндо , или когда С 2` более чем С 3` удалён от плоскости С 1` О 1` С 4` и находится по одну сторону с С 5`; О 1` у d рибоз обеих цепей, имеющих син - конформацию гликозидной связи, всегда направлен в д. Г и д. Ц вниз или вверх относительно оси спирали, а у d рибоз имеющих анти- конформацию О 1` направлен в сторону от оси спирали
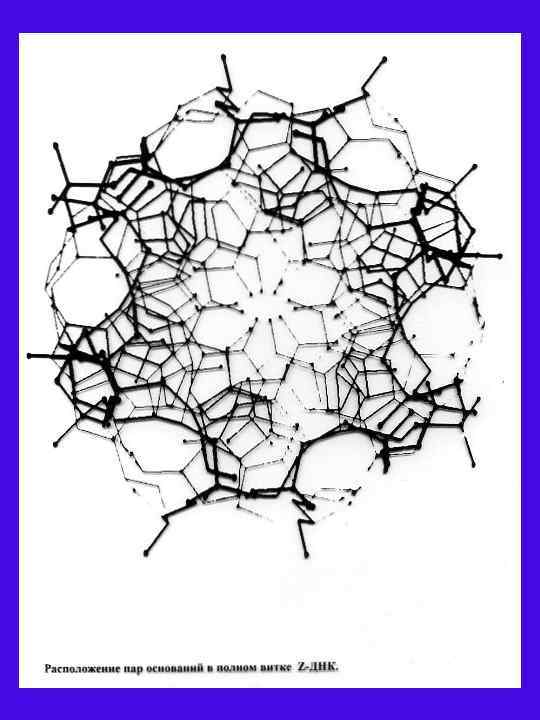
 Z-")
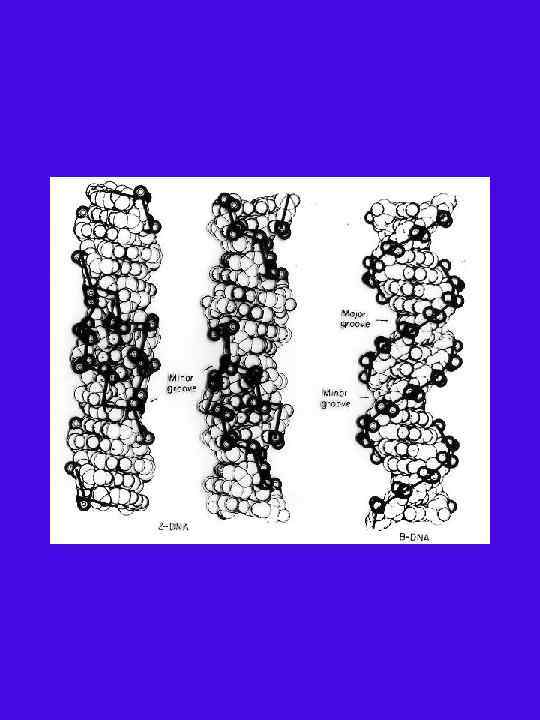
Сравнение спиралей B , Z ДНК и Aп ДНК Параметры B- ДНК*(14, 15) Z- ДНК*(14, 15) Aп-ДНК Правое Левое 10 12 (6 димеров) Диаметр спирали, 20 18 Трансляция на нуклеотид, 3, 4 3, 7 3, 44 Шаг спирали, 34 45 41, 3 Наклон пары оснований 6 7 0 Поворот на нуклеотид, град 36 36 -60 димер) (на -60 d-гуанозин Anti Syn Anti, Syn d-цитидин Anti Syn Anti, Syn C 2 endo C endo 2 C 3 endo C endo 2 C 2 endo C endo 2 an +ck an, +ck, -ck Направление спирали Число остатков на виток 12(3 тетраме р) -120 (per тетраме р) Угол вращения вокруг гликозидной связи Sugar pucker d-гуанозин d-цитидин Торсионный угол вращения вокруг C 4 -C 5 связи d-гуанозин d-цитидин Расстояние фосфора от оси, d Гф Ц d Цф. Г d Гф. Г d Цф. Ц 9 9 8 6, 9 - 8 8, 5 5, 8 3, 6 Расстояние между фосфорами комплементарных пар, d (Гф. Ц) d (Цф. Г) d (Гф. Г) d (Цф. Ц) 17, 5 15 12, 5 15, 3 13, 1 8, 9 7, 9
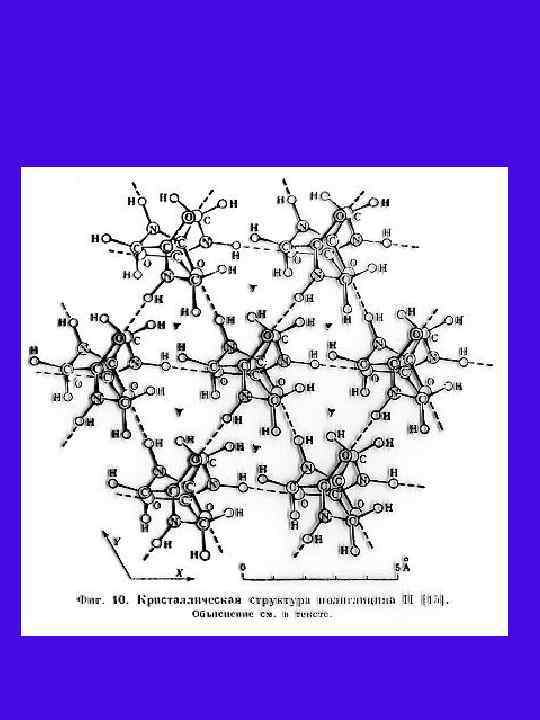

А Б В Рис. 6. А Структура апатита с расположенными в ней цепями полиглицина II(полипролина II, коллагена). В зоне формирования белковых цепей в ячейке апатита анионы РО 43 замещены на CО 32. Б, В Электронная микрофотография гексагональной упаковки молекулы белка в кристалле каталазы из печени быка (увеличение Х 5. 105) (Б) и кристаллической лейцинаминопептидазы (В) [Вайнштейн 1986, с. 156].

Рис. 8. Предполагаемая схема появления разного числа хромосом в будущих протоклетках.

§ Роль минералов в возникновении протоклеток § Вести речь о реальных механизмах возникновения жизни можно и нужно основываясь на твердофазных эффектах в минеральных и жидких кристаллах. Участие минералов в процессе возникновения жизни делает его закономерным, а время процесса весьма кратким. В клетках, какими мы их представляем сегодня, все элементы системы так тесно связаны друг с другом, что отсутствие даже одного из них нарушает работу всей системы. Первичные пробионты, по-видимому, должны были представлять собой хорошо упорядоченные образования. Возникает вопрос, а не были ли предшественники первичных протоклеток, органическими кристаллами? Подобное предположение было высказано и А. Кэрнс-Смитом [17, c. 68]. Ведь кристаллы - это образования, способные к самосборке. У кристаллов есть основной тип строения, для которого характерна высокая периодичность, но в каждом реальном кристалле эта структура имеет дефекты. Присутствие дефектов, малых или больших, в структуре кристаллов делает их потенциально высокоинформативными. Дефекты кристаллов могут приводить к образованию множества стабильных альтернативных конфигураций, что является необходимым условием для хранения информации.
, состоящая из стопки")
Рис. 9. Схема образования биогенных кристаллов. Вначале образуется органическая матрица (I), состоящая из стопки полисахаридных пластин гексагональной или эллиптической формы и «прошита» фибриллами белка. Затем внутри фибрилл и между белковыми фибриллами в самой полисахаридной матрице возникают зародыши минеральных кристаллов; которые растут и формируют элементарный минеральный кристаллит (2). Конечная стадия минерализации образование минерализованного поликристалла (3). Совокупность таких кристаллов и составляет скелетную ткань кость, раковину, панцирь и т. п. Схема по С. Н. Голубеву [13].

Рис. 10. Схема действия факторов ослабляющих кристаллическую решетку апатита, облегчающих и ускоряющих процесс диффузии элементов восстановительной газовой фазы внутрь кристалла при формировании органоминерального комплекса. Процесс идет однонаправленное (от центра Земли к поверхности) по свободным полостям или каналам в структуре апатита либо по границе кристаллических зерен, замещая атомы или вакансии в кристаллической решетке в безводной фазе при температуре » 200°С.
 в формировании ключевых")
Рис. 11. Общая схема участия комплекса сокристаллизующихся минералов (апатит карбонатапатит кальцит) в формировании ключевых структурных образований будущих протоклеток.

Рис. 12. Схема возникновения транскрипционно трансляционного механизма в протоклетках. В области накопления деффектов и сокристаллизации: 1 – образуется тройной комплекс — ДНК, РНК, белки; 2 – запуск матричного механизма может происходить при изменении концентрации ионов, путём раскручивания спирализованного комплекса; 3 – главный фактор перевода р. РНК и т. РНК из линейной структуры во 2 3 ю наличие обращённых повтлоров и комплексных белков; 4 донором аминокислот для ААт. РНК синтетазы является зона кальцита; 5 спаривание м. РНК с Ап ДНК нарушается подходом в результате конформационных изменений 30 SRb и 50 SRb частиц.

§ § Предложенный механизм формирования органоминерального комплекса будущих протоклеток на комплексе минералов позволяет ответить на основные вопросы проблемы происхождения жизни: как и почему клетки являются апериодическими, гомеостатическими, самовоспроизводимыми жидкими кристаллами. Отпадают вопросы относительно синтеза протобиополимеров в воде и создания упорядоченной живой системы против законов термодинамики. Однонаправленность и одновременность синтеза НК и белков в объеме минералов по принципу «спираль в спираль» , по видимому, сыграла ключевую роль в возникновении структурной асимметрии мономерных звеньев и апериодичности биомолекул. В наследство от минералов будущие протоклетки получили: 1) матричный механизм синтеза на участке накопления дефектов в решетке апатита и сокристаллизующихся с ним минералов; 2) универсальный генетический код (единая мат рица апатита) и отсутствие универсальности в строении структур, участвующих в реализации этого кода; 3) особенности структуры ДНК эукариот (сателлитные, умеренно повторенные и уникальные гены); 4) невозможность удалить сателитные участки ДНК в силу необходимости поддержания возникшего гомеостатического состояния живых систем: 5) ключевую роль Са 2+ и РО 43 групп в жизнедеятельности живых систем; 5) белки, специфически связывающие только Са 2+ или другие ионы; 6) живые системы, использующие для построения сво его скелета только Sr, Si, Са, РО 43 ионы или другие эле менты. Мы предполагаем, что предложенный механизм является универсальным для возникновения органических веществ на Земле или других планетах типа Земля. Биомолекулы живых систем, гуминовый скелет почв, углеводороды нефти – все это результат взаимодействия компонентов газовой фазы с решетками апатита, карбонатапатита, кальцита, кристобалита, для гумуса почв – алюмосиликаты; для углеводородов нефти – минералы с различной структурой кристаллической решетки.

Послесловие. Предложенная теория позволяет сделать предположение о реальной связи жизненного потенциала будущих живых систем (прежде всего млекопитающих) с годом и месяцем формирования эмбриона, а не временем его рождения. Это хорошо согласуется с Восточным (12 годичный цикл) и Зодиакальным гороскопом (28 дневный цикл или лунный календарь). В соответствии с предложенной теорией и предположениями ряда авторов (Дж. Бернал – [31], Кэрнс Смит – [39]) живые клетки являются жидко кристаллическими комплексами. Хорошо известно, что жидкие кристаллы меняют свою конформацию под воздействием различных факторов, в том числе, таких как температура, электрическое, магнитное, электро магнитное излучение, силовые поля космического пространства. Такое воздействие наиболее эффективно проявляется на стадии формирования эмбриона после оплодотворения и характеризуется определенной цикличностью, связанной с постоянно действующими силовыми полями космического пространства. Эффективность действия таких полей определяется положением планет в нашей галактике, и вероятно, наиболее сильно – планет солнечной системы. Сформированный на первом этапе жидко кристаллический структурный остов клеток, становиться матрицей для дальнейшего роста и развития органов и тканей и как исходная матрица не может быть в дальнейшем изменен. Именно поэтому, жизненный потециал будущей живой системы в значительной степени становится зависимым от времени начала формирования эмбриона т. е. фактического воздействия в этот момент силовых полей космического пространства. Далее срабатывает механизм: структура – функция. При этом, мы не отбрасываем генентический потенциал заложенный в половых клетках родителей. Описанная выше зависимость формирования потенциала будущих живых систем от года и месяца рождения, подкрепленная огромной статистической информацией накопленной в литературе по зодиакальным гороскопам, может быть эффективно использована в судебно правовой практике, лечении нарко и алкогольной зависимости, использовании гипноза и других воздействий на организм человека.

Публикации Э. Я. Костецкого по проблеме возникновения жизни. § Э. Я. Костецкий, Алексаков С. А. О возможности синтеза нуклеопротеинов на матрице апатита //Докл. Акад. Наук СССР. 1981. Т. 260. С. 1013 1018. § Э. Я. Костецкий. О происхождении жизни и возможности формирования протоклеток и их структурных элементов на кристаллах апатита //Журн. Эволюц. Биохимии и Физиологии. 1999. Т. 35. С. 249 256. § EDUARD Y. KOSTETSKY. The Possibility of the Formation of Protocells and Their Structural Components on the Basis of the Apatite Matrix and Cocrystallizing Minerals // Journal of Biological Physics 2005. 31: 607– 638, § Э. Я. КОСТЕЦКИЙ. КАК ВОЗНИКЛА ЖИЗНЬ. ТЕОРИЯ ВОЗНИКНОВЕНИЯ ПРОТОКЛЕТОК И ИХ СТРУКТУРНЫХ КОМПОНЕНТОВ // ВЕСТНИК ТИХООКЕАНСКОГО ГОСУДАРСТВЕННОГО ЭКОНОМИЧЕСКОГО УНИВЕРСИТЕТА. ВЛАДИВОСТОК, 2008. N 1, ЧАСТЬ 1, С. 79 102; N 2, ЧАСТЬ 2, С. 86 104. § Э. Я. КОСТЕЦКИЙ. КАК ВОЗНИКЛА ЖИЗНЬ. ТЕОРИЯ ВОЗНИКНОВЕНИЯ ПРОТОКЛЕТОК И ИХ СТРУКТУРНЫХ КОМПОНЕНТОВ ПРИ УЧАСТИИ АПАТИТОВОЙ МАТРИЦЫ И СОКРИСТАЛИЗУЮЩИХСЯ С НЕЙ МИНЕРАЛОВ // ТРУДЫ ПРОФЕССОРСКОГО КЛУБА. ВЫПУСК 11. ВЛАДИВОСТОК, 2008. С. 53 77.

Презентация БПИ-3 Для доклада.ppt