

Lekts_Dykhanie_-2_stom.ppt
- Количество слайдов: 29
 Кафедра нормальной физиологии СГМУ Лекция Транспорт газов кровью. Регуляция дыхания Лекция для студентов стоматологического факультета Доцент кафедры норм. физиологии, к. м. н. Шерстенникова Александра Константиновна 2011 год
Кафедра нормальной физиологии СГМУ Лекция Транспорт газов кровью. Регуляция дыхания Лекция для студентов стоматологического факультета Доцент кафедры норм. физиологии, к. м. н. Шерстенникова Александра Константиновна 2011 год
 • 1) Транспорт") 3 этап дыхания -ТРАНСПОРТ ГАЗОВ КРОВЬЮ. (главный вид транспорта - конвекция) • 1) Транспорт кислорода кровью • О 2 находится в крови в двух состояниях: физически растворенном и химически связанном. В 100 мл крови растворяется 0, 003 мл О 2. Остальная часть кислорода сражу же диффундирует в эритроциты, связываясь с гемоглобином, образуя оксигемоглобин Нb. О 2
3 этап дыхания -ТРАНСПОРТ ГАЗОВ КРОВЬЮ. (главный вид транспорта - конвекция) • 1) Транспорт кислорода кровью • О 2 находится в крови в двух состояниях: физически растворенном и химически связанном. В 100 мл крови растворяется 0, 003 мл О 2. Остальная часть кислорода сражу же диффундирует в эритроциты, связываясь с гемоглобином, образуя оксигемоглобин Нb. О 2
 • 1 г гемоглобина может связать 1, 34 мл кислорода. Следовательно, при содержании гемоглобина 140 -150 г/л, в каждом литре крови максимально возможное содержание кислорода в химически связанной форме составит 190 -200 мл О 2/л. Это так называемая кислородная емкость крови - величина, отражающая количество кислорода, которое может связаться с гемоглобином.
• 1 г гемоглобина может связать 1, 34 мл кислорода. Следовательно, при содержании гемоглобина 140 -150 г/л, в каждом литре крови максимально возможное содержание кислорода в химически связанной форме составит 190 -200 мл О 2/л. Это так называемая кислородная емкость крови - величина, отражающая количество кислорода, которое может связаться с гемоглобином.
 зависит от парциального напряжения кислорода в крови") Превращение Hb в Hb. O 2 (оксигемоглобин) зависит от парциального напряжения кислорода в крови (PO 2). Графически эта запись выражается кривой диссоциации оксигемоглобина и имеет Sобразную форму, где по оси абсцисс откладывается РО 2 в крови, по ординате насыщение гемоглобина кислородом (Hb. O 2) в %. Hb. O 2 %. РО 2 мм. рт. ст
Превращение Hb в Hb. O 2 (оксигемоглобин) зависит от парциального напряжения кислорода в крови (PO 2). Графически эта запись выражается кривой диссоциации оксигемоглобина и имеет Sобразную форму, где по оси абсцисс откладывается РО 2 в крови, по ординате насыщение гемоглобина кислородом (Hb. O 2) в %. Hb. O 2 %. РО 2 мм. рт. ст
 • Когда напряжение кислорода в крови равно нулю, в крови находится только восстановленный гемоглобин. Повышение напряжения кислорода приводит к увеличению оксигемоглобина. Особенно быстро уровень оксигемоглобина возрастает при напряжении от 10 -40 мм рт. ст. - до 75%. , Hb. O 2 %. РО 2 мм. рт. ст
• Когда напряжение кислорода в крови равно нулю, в крови находится только восстановленный гемоглобин. Повышение напряжения кислорода приводит к увеличению оксигемоглобина. Особенно быстро уровень оксигемоглобина возрастает при напряжении от 10 -40 мм рт. ст. - до 75%. , Hb. O 2 %. РО 2 мм. рт. ст
 • при напряжении О 2 60 мм рт. ст. насыщение гемоглобина кислородом составляет 90%. При дальнейшем повышении напряжения кислорода насыщение гемоглобина идет очень медленно. Плато кривой характерно для насыщенной О 2 артериальной крови, а крутая нисходящая часть кривой - для венозной.
• при напряжении О 2 60 мм рт. ст. насыщение гемоглобина кислородом составляет 90%. При дальнейшем повышении напряжения кислорода насыщение гемоглобина идет очень медленно. Плато кривой характерно для насыщенной О 2 артериальной крови, а крутая нисходящая часть кривой - для венозной.
 На диссоциацию оксигемоглобина влияют следующие факторы крови: Сдвиг графика вправо происходит при повышении: - РСО 2; - t 0, - 2, 3 -дифосфоглицерат, - Н+ (снижении р. Н). Это значительно увеличивает скорость распада оксигемоглобина, улучшая его отдачу в тканях (немного затрудняя его присоединение в легких), и уменьшает сродство гемоглобина к О 2.
На диссоциацию оксигемоглобина влияют следующие факторы крови: Сдвиг графика вправо происходит при повышении: - РСО 2; - t 0, - 2, 3 -дифосфоглицерат, - Н+ (снижении р. Н). Это значительно увеличивает скорость распада оксигемоглобина, улучшая его отдачу в тканях (немного затрудняя его присоединение в легких), и уменьшает сродство гемоглобина к О 2.
 • Сдвиг графика влево происходит при снижении: - РСО 2; - t 0, - 2, 3 -дифосфоглицерат, - Н+ (повышении р. Н). Это увеличивает сродство гемоглобина к О 2, ухудшая его отдачу в тканях (немного улучшая его присоединение в легких). Таким образом, транспорт кислорода обеспечивается, в основном, за счет химической связи его с гемоглобином.
• Сдвиг графика влево происходит при снижении: - РСО 2; - t 0, - 2, 3 -дифосфоглицерат, - Н+ (повышении р. Н). Это увеличивает сродство гемоглобина к О 2, ухудшая его отдачу в тканях (немного улучшая его присоединение в легких). Таким образом, транспорт кислорода обеспечивается, в основном, за счет химической связи его с гемоглобином.
Транспорт углекислого газа. • • Углекислый газ находится в крови в 3 фракциях. •") 2)Транспорт углекислого газа. • • Углекислый газ находится в крови в 3 фракциях. • Если из венозной крови можно извлечь около 58 об% углекислого газа (или из 1 л крови 580 мл углекислого газа), то
2)Транспорт углекислого газа. • • Углекислый газ находится в крови в 3 фракциях. • Если из венозной крови можно извлечь около 58 об% углекислого газа (или из 1 л крови 580 мл углекислого газа), то
Транспорт углекислого газа 58 % СО 2 3% СО 2 5% Hb. СО 2") 2)Транспорт углекислого газа 58 % СО 2 3% СО 2 5% Hb. СО 2 50 % бикарбонат 34 % Na. HСО 2 17 % KHСО 2
2)Транспорт углекислого газа 58 % СО 2 3% СО 2 5% Hb. СО 2 50 % бикарбонат 34 % Na. HСО 2 17 % KHСО 2
 • СО 2, согласно градиенту напряжения, переходит из клетки в эритроциты и соединяется там с водой. под влиянием фермента - карбоангидразы образуется угольная кислота. • В плазме также образуется небольшое количество углекислоты, но эта реакция идет очень медленно, т. к в плазме нет фермента карбоангидразы, катализирующего образование углекислоты. А в эритроцитах под влиянием этого фермента карбоангидразы - она резко ускоряется (в десятки тысяч раз).
• СО 2, согласно градиенту напряжения, переходит из клетки в эритроциты и соединяется там с водой. под влиянием фермента - карбоангидразы образуется угольная кислота. • В плазме также образуется небольшое количество углекислоты, но эта реакция идет очень медленно, т. к в плазме нет фермента карбоангидразы, катализирующего образование углекислоты. А в эритроцитах под влиянием этого фермента карбоангидразы - она резко ускоряется (в десятки тысяч раз).
 • Угольная кислота диссоциирует на Н+ и НСО 3 -, • НСО 3 -, выходит в плазму и соединяется там с Na+, а взамен ему входит в эритроцит Cl-. • Н+ соединяется с Hb, который отходит от О 2 • О 2 идет в клетку
• Угольная кислота диссоциирует на Н+ и НСО 3 -, • НСО 3 -, выходит в плазму и соединяется там с Na+, а взамен ему входит в эритроцит Cl-. • Н+ соединяется с Hb, который отходит от О 2 • О 2 идет в клетку
 • В капиллярах малого круга кровообращения происходят обратные реакции. • Впервые И. М. Сеченов высказал предположение о том, что в эритроцитах содержится катализатор, который ускоряет процесс синтеза угольной кислоты.
• В капиллярах малого круга кровообращения происходят обратные реакции. • Впервые И. М. Сеченов высказал предположение о том, что в эритроцитах содержится катализатор, который ускоряет процесс синтеза угольной кислоты.
 Регуляция дыхания • Дыхательный центр – сеть многочисленных нейронов, расположенных на разных уровнях ЦНС и обеспечивающие газообмен между организмом и 1. окружающей 2. средой. УРОВНИ: Спинной (С 3 -5, Th 1 -6) Продолговатый 3. Подкорковые структуры: варолиев мост, ГТ, ЛС, КГМ
Регуляция дыхания • Дыхательный центр – сеть многочисленных нейронов, расположенных на разных уровнях ЦНС и обеспечивающие газообмен между организмом и 1. окружающей 2. средой. УРОВНИ: Спинной (С 3 -5, Th 1 -6) Продолговатый 3. Подкорковые структуры: варолиев мост, ГТ, ЛС, КГМ
 Главный дыхательный центр • - совокупность специфических дыхательных ядер продолговатого мозга и варолиева моста. • Структуры, необходимые для возникновения дыхательного ритма, были обнаружены в продолговатом мозге (Миславский Н. А. 1885 г. ).
Главный дыхательный центр • - совокупность специфических дыхательных ядер продолговатого мозга и варолиева моста. • Структуры, необходимые для возникновения дыхательного ритма, были обнаружены в продолговатом мозге (Миславский Н. А. 1885 г. ).
 В состав главного ДЦ входят 3 структуры: • 1. Дорсальная группа нейронов ПМ • 2. Вентральная группа нейронов ПМ • 3. Варолиев мост: пневмотаксический и апнейстический центр
В состав главного ДЦ входят 3 структуры: • 1. Дорсальная группа нейронов ПМ • 2. Вентральная группа нейронов ПМ • 3. Варолиев мост: пневмотаксический и апнейстический центр
 • Формирует") 1. Дорсальная группа нейронов ПМ • Является главной структурой ДЦ (инспираторный центр) • Формирует дыхательный ритм и посылает импульсы к инспираторным мышцам • Получает всю информацию, необходимую для дыхания (изменение р. Н, О 2, СО 2). • Расположена в пределах ядра одиночного пути, в которое поступают волокна блуждающего нерва
1. Дорсальная группа нейронов ПМ • Является главной структурой ДЦ (инспираторный центр) • Формирует дыхательный ритм и посылает импульсы к инспираторным мышцам • Получает всю информацию, необходимую для дыхания (изменение р. Н, О 2, СО 2). • Расположена в пределах ядра одиночного пути, в которое поступают волокна блуждающего нерва
 2. Вентральная группа нейронов ПМ • Включается при форсированном дыхании. Следовательно посылает импульсы к экспираторным мышцам.
2. Вентральная группа нейронов ПМ • Включается при форсированном дыхании. Следовательно посылает импульсы к экспираторным мышцам.
 Варолиев мост: Пневмотаксический центр моста • Расположен в верхней части моста • * Эффект: тормозит инспираторные (дорсальные) нейроны, вызывает укорочение вдоха и тем самым делая дыхание более поверхностным и частым. Апнейстический центр расположен в нижней части моста, оказывает постоянную стимуляцию инспираторных нейронов, способствует вдоху и пролонгирует его.
Варолиев мост: Пневмотаксический центр моста • Расположен в верхней части моста • * Эффект: тормозит инспираторные (дорсальные) нейроны, вызывает укорочение вдоха и тем самым делая дыхание более поверхностным и частым. Апнейстический центр расположен в нижней части моста, оказывает постоянную стимуляцию инспираторных нейронов, способствует вдоху и пролонгирует его.
 Происхождение дыхательного ритма • Вдох - осуществляется активно, за счет сокращения дыхательных мышц. Следовательно, во время вдоха ДЦ посылает возбуждающие импульсы к мотонейронам инспираторных мышц. • Выдох - пассивный процесс. Следовательно, при выдохе достаточно затормозить нейроны, посылающие эти возбуждающие импульсы.
Происхождение дыхательного ритма • Вдох - осуществляется активно, за счет сокращения дыхательных мышц. Следовательно, во время вдоха ДЦ посылает возбуждающие импульсы к мотонейронам инспираторных мышц. • Выдох - пассивный процесс. Следовательно, при выдохе достаточно затормозить нейроны, посылающие эти возбуждающие импульсы.
 Инспираторный нейрон обладает автоматизмом • При возбуждении инспираторный нейрон посылает импульс к инспираторным мышцам и одновременно к тормозному нейрону. • Тормозной нейрон, получающий импульс от инспираторного нейрона тормозит его. • Вдох прекращается и действие тормозного нейрона угасает. В результате инспираторный нейрон начинает вновь генерировать – и начинается новый вдох.
Инспираторный нейрон обладает автоматизмом • При возбуждении инспираторный нейрон посылает импульс к инспираторным мышцам и одновременно к тормозному нейрону. • Тормозной нейрон, получающий импульс от инспираторного нейрона тормозит его. • Вдох прекращается и действие тормозного нейрона угасает. В результате инспираторный нейрон начинает вновь генерировать – и начинается новый вдох.
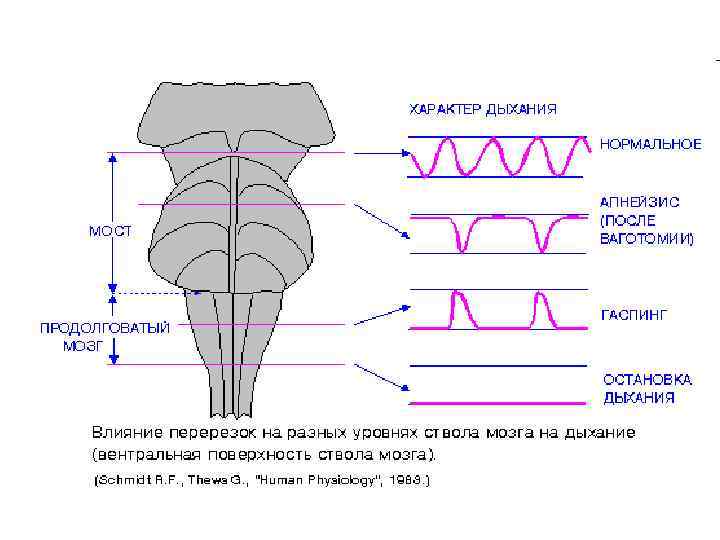
 • При нанесении перерезки выше моста характер дыхания не изменяется. • Отделение продолговатого мозга от спинного приводит к полной остановке дыхания. • Если нанести перерезку на границе между верхней и средней третью моста и одновременно пересечь оба блуждающих нерва, то дыхание остановится в фазе вдоха, лишь иногда прерываемой экспираторными движениями. Такой тип дыхания называется апнейзис. Очевидно, что после подобной перерезки устраняются тормозные влияния верхних отделов моста на нейроны, отвечающие за вдох. • После перерезки ствола мозга ниже моста дыхательные движения сохраняются, однако их ритм становится неправильным. Иногда возникает дыхание типа гаспинга, при котором длительный выдох периодически прерывается короткими вдохами. • Таким образом, изолированный продолговатый мозг способен генерировать дыхательный ритм. Однако для стабилизации и координации этого ритма необходимо участие нервных образований, расположенных в среднем и верхнем отделах моста.
• При нанесении перерезки выше моста характер дыхания не изменяется. • Отделение продолговатого мозга от спинного приводит к полной остановке дыхания. • Если нанести перерезку на границе между верхней и средней третью моста и одновременно пересечь оба блуждающих нерва, то дыхание остановится в фазе вдоха, лишь иногда прерываемой экспираторными движениями. Такой тип дыхания называется апнейзис. Очевидно, что после подобной перерезки устраняются тормозные влияния верхних отделов моста на нейроны, отвечающие за вдох. • После перерезки ствола мозга ниже моста дыхательные движения сохраняются, однако их ритм становится неправильным. Иногда возникает дыхание типа гаспинга, при котором длительный выдох периодически прерывается короткими вдохами. • Таким образом, изолированный продолговатый мозг способен генерировать дыхательный ритм. Однако для стабилизации и координации этого ритма необходимо участие нервных образований, расположенных в среднем и верхнем отделах моста.
 Рефлекторная регуляция дыхания. Механорецепторы легких информируют ДЦ о вентиляци легких. • Рецепторы растяжения легких (медленно адаптируются) расположены в гладких мышцах трахеи и бронхах, реагируют на увеличение объема легких при вдохе; с них возникает инспираторнотормозящий рефлекс Геринга - Брейера. • Ирритантные рецепторы расположены в эпителии бронхов, реагируют на быстрое изменение объема легких, на механические воздействия (пыль) и на пары химических веществ: * это быстро адаптирующиеся рецепторы. С них формируются рефлексы кашля. • Юкстаальвеолярные рецепторы (J-рецепторы) локализуются в интерстиции альвеол у капилляров, реагируют на давление жидкости в межклеточном пространстве легких, с них формируется одышка.
Рефлекторная регуляция дыхания. Механорецепторы легких информируют ДЦ о вентиляци легких. • Рецепторы растяжения легких (медленно адаптируются) расположены в гладких мышцах трахеи и бронхах, реагируют на увеличение объема легких при вдохе; с них возникает инспираторнотормозящий рефлекс Геринга - Брейера. • Ирритантные рецепторы расположены в эпителии бронхов, реагируют на быстрое изменение объема легких, на механические воздействия (пыль) и на пары химических веществ: * это быстро адаптирующиеся рецепторы. С них формируются рефлексы кашля. • Юкстаальвеолярные рецепторы (J-рецепторы) локализуются в интерстиции альвеол у капилляров, реагируют на давление жидкости в межклеточном пространстве легких, с них формируется одышка.
 Рефлекс Геринга-Брейера. • Дуга этого рефлекса начинается от рецепторов растяжения легких. • Аффрентные волокна от рецепторов идут в составе n. vagus в ДЦ. • От ДЦ импульсы идут в спинной мозг к мотонейронам, иннервирующим дыхательные мышцы (С 3 -5 диафрагму, Th 1 -6 межреберные мышцы)
Рефлекс Геринга-Брейера. • Дуга этого рефлекса начинается от рецепторов растяжения легких. • Аффрентные волокна от рецепторов идут в составе n. vagus в ДЦ. • От ДЦ импульсы идут в спинной мозг к мотонейронам, иннервирующим дыхательные мышцы (С 3 -5 диафрагму, Th 1 -6 межреберные мышцы)
, тканях внутренних") Гуморальная регуляция дыхания Периферические хеморецепторы: • локализуются в сосудах (особенно в артериях), тканях внутренних органов, их концентрация максимальна в синокаротидной и аортальной зонах; • афферентная импульсация от них проводится по блуждающему нерву; • имеют высокую чувствительность к изменению Р 02 в артериальной крови (особенно к его снижению) – гипоксемии.
Гуморальная регуляция дыхания Периферические хеморецепторы: • локализуются в сосудах (особенно в артериях), тканях внутренних органов, их концентрация максимальна в синокаротидной и аортальной зонах; • афферентная импульсация от них проводится по блуждающему нерву; • имеют высокую чувствительность к изменению Р 02 в артериальной крови (особенно к его снижению) – гипоксемии.
 Центральные хеморецепторы: локализуются на переднебоковой поверхности продолговатого мозга и моста в виде трех пар скопления нейронов; • отличаются высокой чувствительностью к изменению р. Н и Рсо 2 - гиперкапния;
Центральные хеморецепторы: локализуются на переднебоковой поверхности продолговатого мозга и моста в виде трех пар скопления нейронов; • отличаются высокой чувствительностью к изменению р. Н и Рсо 2 - гиперкапния;
 Влияние на дыхательный центр высших отделов ЦНС Гипоталамус осуществляет: • связь дыхания с обменом веществ и терморегуляцией в организме; • регуляцию дыхания для обеспечения поведенческих актов, направленных на удовлетворение биологических потребностей (агрессивно-оборонительной, пищевой, половой и др. ). Лимбическая система осуществляет связь дыхания с вегетативной регуляцией органов и с эмоциями.
Влияние на дыхательный центр высших отделов ЦНС Гипоталамус осуществляет: • связь дыхания с обменом веществ и терморегуляцией в организме; • регуляцию дыхания для обеспечения поведенческих актов, направленных на удовлетворение биологических потребностей (агрессивно-оборонительной, пищевой, половой и др. ). Лимбическая система осуществляет связь дыхания с вегетативной регуляцией органов и с эмоциями.
 Кора больших полушарий осуществляет: • связь дыхания с изменениями внешней и внутренней среды (через сенсорную кору) • условно-рефлекторную и произвольную регуляцию дыхания; • корковое дублирование автоматии дыхательного центра; • регуляцию дыхания для обеспечения социальных форм поведения; • особенности дыхания при образовании речи (участие в механизмах фонации и артикуляции.
Кора больших полушарий осуществляет: • связь дыхания с изменениями внешней и внутренней среды (через сенсорную кору) • условно-рефлекторную и произвольную регуляцию дыхания; • корковое дублирование автоматии дыхательного центра; • регуляцию дыхания для обеспечения социальных форм поведения; • особенности дыхания при образовании речи (участие в механизмах фонации и артикуляции.