

2Изменчивость характера наследования и проявления признаков в зависимости.ppt
- Количество слайдов: 50
 Изменчивость характера наследования и проявления признаков в зависимости от действия и взаимодействия генов После 1900 г. исследуется влияние генотипа на проявление признаков. Полученные результаты скрещиваний не соответствуют числовым соотношениям Менделя. в точности Для объяснения выдвинуты новые гипотезы и разработаны специальные проверочные системы скрещиваний.
Изменчивость характера наследования и проявления признаков в зависимости от действия и взаимодействия генов После 1900 г. исследуется влияние генотипа на проявление признаков. Полученные результаты скрещиваний не соответствуют числовым соотношениям Менделя. в точности Для объяснения выдвинуты новые гипотезы и разработаны специальные проверочные системы скрещиваний.
 Связь между геном и признаком может быть простой и однозначной: v Расщепление в F 2 идет в соотношении Развитие одного признака контролируется одним геном (моногенный контороль). 3 : 1 и 9 : 3 : 1 Моногибридное скрещивание Дигибридное скрещивание Одна пара аллелей Две пары аллелей
Связь между геном и признаком может быть простой и однозначной: v Расщепление в F 2 идет в соотношении Развитие одного признака контролируется одним геном (моногенный контороль). 3 : 1 и 9 : 3 : 1 Моногибридное скрещивание Дигибридное скрещивание Одна пара аллелей Две пары аллелей
 После 1900 г. было показано, что между генами существуют сложные взаимоотношения: один признак контролируется разными генами, взаимодействующими между собой • Комплементарность (взаимодополнение) • Эпистаз • Полимерия • Гены-модификаторы один и тот же ген может оказывать влияние на несколько различных признаков Плейотропия
После 1900 г. было показано, что между генами существуют сложные взаимоотношения: один признак контролируется разными генами, взаимодействующими между собой • Комплементарность (взаимодополнение) • Эпистаз • Полимерия • Гены-модификаторы один и тот же ген может оказывать влияние на несколько различных признаков Плейотропия
. Одно из возможных структурных состояний гена. Аллели дикого типа") Аллельное взаимодействие генов Аллель (allele). Одно из возможных структурных состояний гена. Аллели дикого типа – аллели, которые обычны для данной популяции. Обычно они доминантны и определяют фенотип дикого типа. Новые аллели есть результат мутации, приводит к изменениям фенотипа. Мутантный аллель – кодирует часто измененный генный продукт. Обычно мутация вызывает утрату функции. которую в норме кодирует аллель дикого типа.
Аллельное взаимодействие генов Аллель (allele). Одно из возможных структурных состояний гена. Аллели дикого типа – аллели, которые обычны для данной популяции. Обычно они доминантны и определяют фенотип дикого типа. Новые аллели есть результат мутации, приводит к изменениям фенотипа. Мутантный аллель – кодирует часто измененный генный продукт. Обычно мутация вызывает утрату функции. которую в норме кодирует аллель дикого типа.
 Аллельное взаимодействие генов Взаимоотношение двух аллелей одного гена называют аллельным взаимодействие генов. 1. Доминантность или доминирование 2. Сверхдоминирование или сверхдоминантность 3. Кодоминирование 4. Множественный аллелизм
Аллельное взаимодействие генов Взаимоотношение двух аллелей одного гена называют аллельным взаимодействие генов. 1. Доминантность или доминирование 2. Сверхдоминирование или сверхдоминантность 3. Кодоминирование 4. Множественный аллелизм
 Аллельное взаимодействие генов 1. Доминантность или доминирование - явление, при котором один из аллелей гетерозиготы (Аа) оказывает более сильное влияние на контролируемый признак, чем другой аллель. Господствующий аллель называется доминантным (А), а подавляемый рецессивный (а). Доминирование Полное 3 : 1 Неполное 1 : 2 : 1 У Аа – промежуточное прояление признака ( окраска цветка у львиный зева , ночной красавицы)
Аллельное взаимодействие генов 1. Доминантность или доминирование - явление, при котором один из аллелей гетерозиготы (Аа) оказывает более сильное влияние на контролируемый признак, чем другой аллель. Господствующий аллель называется доминантным (А), а подавляемый рецессивный (а). Доминирование Полное 3 : 1 Неполное 1 : 2 : 1 У Аа – промежуточное прояление признака ( окраска цветка у львиный зева , ночной красавицы)
 Расположение аллелей одной аллельной пары Аллели одного гена локализованы в одном локусе гомологичных хромосом. У диплоидных организмов в каждом локусе может быть только два аллеля (АА, Аа, аа). локус ____А _______А______ гомозигота по доминантному аллелю ____А______а______ гетерозигота ____а___________ гомозигота по рецессивному аллелю
Расположение аллелей одной аллельной пары Аллели одного гена локализованы в одном локусе гомологичных хромосом. У диплоидных организмов в каждом локусе может быть только два аллеля (АА, Аа, аа). локус ____А _______А______ гомозигота по доминантному аллелю ____А______а______ гетерозигота ____а___________ гомозигота по рецессивному аллелю
 превосходит по выраженности признака гомозигот") Аллельное взаимодействие генов 2. Сверхдоминирование или сверхдоминантность Гетерозигота (Аа) превосходит по выраженности признака гомозигот по доминантному (АА) и рецессивному аллелям (аа). Сверхдоминирование играет огромную роль в природе, с ним связано явление гетерозиса.
Аллельное взаимодействие генов 2. Сверхдоминирование или сверхдоминантность Гетерозигота (Аа) превосходит по выраженности признака гомозигот по доминантному (АА) и рецессивному аллелям (аа). Сверхдоминирование играет огромную роль в природе, с ним связано явление гетерозиса.
 Аллельное взаимодействие генов 3. Кодоминирование Ø Два аллеля одного гена кодируют два различных геных продукта. Ø Каждый из двух алеллей гетерозиготы влияет на проявление признака, отсутствуют доминантнорецессивные отношения. Пример: генетический контроль групп крови у человека – АВО система. Группы крови открыты Ландштейнером (1868 -1943). в 1900 г. Карлом
Аллельное взаимодействие генов 3. Кодоминирование Ø Два аллеля одного гена кодируют два различных геных продукта. Ø Каждый из двух алеллей гетерозиготы влияет на проявление признака, отсутствуют доминантнорецессивные отношения. Пример: генетический контроль групп крови у человека – АВО система. Группы крови открыты Ландштейнером (1868 -1943). в 1900 г. Карлом
, А (II), B (III), AB (IV), контролируемых") Кодоминирование Известны четыре группы крови: O (I), А (II), B (III), AB (IV), контролируемых геном I, имеющего три аллеля – i. О, IА, IВ. Аллели IА и IВ доминантны по отношению к аллелю i. О, но кодоминантны по отношению друг к другу. Генотип Группа Наличие антигенов и крови антител i. О O (I) -, IА IА, IА i. О А (II) А, Тип взаимодействия α, ß ß полное доминирование IВ IВ, IВ i. О B (III) В, α полное доминирование IА IВ AB (IV) А и В, - кодоминирование Кодоминирование проявляется в основном по признакам биохимическим и иммунологическим.
Кодоминирование Известны четыре группы крови: O (I), А (II), B (III), AB (IV), контролируемых геном I, имеющего три аллеля – i. О, IА, IВ. Аллели IА и IВ доминантны по отношению к аллелю i. О, но кодоминантны по отношению друг к другу. Генотип Группа Наличие антигенов и крови антител i. О O (I) -, IА IА, IА i. О А (II) А, Тип взаимодействия α, ß ß полное доминирование IВ IВ, IВ i. О B (III) В, α полное доминирование IА IВ AB (IV) А и В, - кодоминирование Кодоминирование проявляется в основном по признакам биохимическим и иммунологическим.
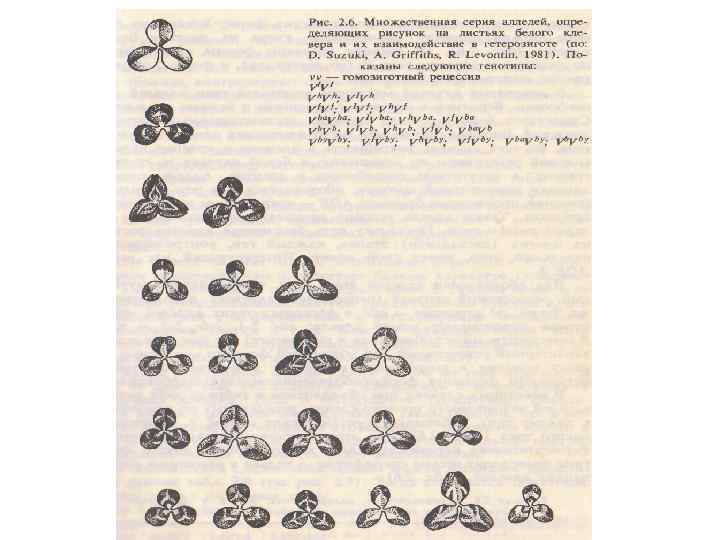
 Аллельное взаимодействие генов 4. Множественный аллелизм - локус представлен более чем двумя аллелями. § Аллельные гены возникают в результате мутации. § Многообразие состояний гена в сумме составляет серию аллелей. каждый из членов серии может быть полностью или неполностью доминировать над другим ее членом. § Явление множественного аллелизма широко распространено в природе (окраска шерсти у животных, окраска глаз у дрозофилы, форма рисунка на листьях клевера, тип совместимости при опылении у высших растений).
Аллельное взаимодействие генов 4. Множественный аллелизм - локус представлен более чем двумя аллелями. § Аллельные гены возникают в результате мутации. § Многообразие состояний гена в сумме составляет серию аллелей. каждый из членов серии может быть полностью или неполностью доминировать над другим ее членом. § Явление множественного аллелизма широко распространено в природе (окраска шерсти у животных, окраска глаз у дрозофилы, форма рисунка на листьях клевера, тип совместимости при опылении у высших растений).
 Множественный аллелизм Классический пример – окраска меха у кроликов. Серия из четырех аллелей гена с определяет окраску меха – с+ > ссh > сa. Аллель ссh является рецессивным по отношению к с+, но доминантным к сh и сa. Гетерозиготы, содержащие разные аллели, называются компаундами.
Множественный аллелизм Классический пример – окраска меха у кроликов. Серия из четырех аллелей гена с определяет окраску меха – с+ > ссh > сa. Аллель ссh является рецессивным по отношению к с+, но доминантным к сh и сa. Гетерозиготы, содержащие разные аллели, называются компаундами.
 Генетическая детерминация окраски меха у кроликов Аллель Генотип Фенотип c+ с+с+, с+сch, с+сh, с +с Агути (серая окраска), дикий тип cch сchсch Шиншилла (светлее дикого типа) сchсh сchсa Светлая шиншилла (светло-серая окраска) ch сhсh, сhсa ca с aс a Гималайский тип (белый мех, кроме лапок, хвоста, ушей и кончика носа) Альбинос (белый мех, глаза красные)
Генетическая детерминация окраски меха у кроликов Аллель Генотип Фенотип c+ с+с+, с+сch, с+сh, с +с Агути (серая окраска), дикий тип cch сchсch Шиншилла (светлее дикого типа) сchсh сchсa Светлая шиншилла (светло-серая окраска) ch сhсh, сhсa ca с aс a Гималайский тип (белый мех, кроме лапок, хвоста, ушей и кончика носа) Альбинос (белый мех, глаза красные)

 Тип совместимости при опылении у высших растений Рис. 1. Механизм влияния генов самостерильности на процессы прорастания пыльцы и оплодотворения (S. Bогojevic, К. Вогojеvic, 1976. — Р. 146)
Тип совместимости при опылении у высших растений Рис. 1. Механизм влияния генов самостерильности на процессы прорастания пыльцы и оплодотворения (S. Bогojevic, К. Вогojеvic, 1976. — Р. 146)
 Неаллельное взаимодействие генов - это взаимодействие генов, локализованных в разных негомологичных хромосомах и контролирующих развитие одного и того же признака. Термин взаимодействие означает влияет несколько генов. что на отдельный признак v Неаллельное взаимодействие генов проявляется в нескольких формах и приводит к своеобразным изменениям дигибридного расщепления 9 А В _: 3 А_ вв : 3 аа В_ : 1 аавв Ø Комплементарность (взаимодополнение) Ø Эпистаз Ø Полимерия Ø Гены-модификаторы
Неаллельное взаимодействие генов - это взаимодействие генов, локализованных в разных негомологичных хромосомах и контролирующих развитие одного и того же признака. Термин взаимодействие означает влияет несколько генов. что на отдельный признак v Неаллельное взаимодействие генов проявляется в нескольких формах и приводит к своеобразным изменениям дигибридного расщепления 9 А В _: 3 А_ вв : 3 аа В_ : 1 аавв Ø Комплементарность (взаимодополнение) Ø Эпистаз Ø Полимерия Ø Гены-модификаторы
 Комплементарное взаимодействие Комплементарность заключается в том, что два и более генов, взаимодействуя между собой, вызывают изменение в проявлении признака, которые не обнаруживаются у родительских форм. Для данного типа взаимодействия характерны следующие формулы расщепления: а) 9 : 3 : 1 б) 9 : 6 : 1 в) 9 : 7 г) 9 : 3 : 4
Комплементарное взаимодействие Комплементарность заключается в том, что два и более генов, взаимодействуя между собой, вызывают изменение в проявлении признака, которые не обнаруживаются у родительских форм. Для данного типа взаимодействия характерны следующие формулы расщепления: а) 9 : 3 : 1 б) 9 : 6 : 1 в) 9 : 7 г) 9 : 3 : 4
:") Комплементарное взаимодействие а б в Рис. 2 . Четыре типа греблей у кур (петухи): а - простой- б- гороховидный; в- ореховидный; г - розовидный; (Ваиг, 1930. - Р. 120) г
Комплементарное взаимодействие а б в Рис. 2 . Четыре типа греблей у кур (петухи): а - простой- б- гороховидный; в- ореховидный; г - розовидный; (Ваиг, 1930. - Р. 120) г
 F 1 х Гороховидный") Комплементарное взаимодействие Р ААвв аа. ВВ х Розовидный гребень (виандотты) F 1 х Гороховидный гребень (порода брамма, куры европейского происхождения) Аа. Вв Ореховидный гребень (куры малайского происхождения) F 2 9 А В _: 3 А_ вв : 3 аа В_ : 1 аавв орех розов горох лист Каждый ген имеет собственное фенотипическое проявление, розовидная и гороховидная формы гребня контролируются каждым доминантным геном в отдельности, Ав и а. В соответственно. Ореховидная форма - результат взаимодействия двух доминантных генов (АВ). Листовидная форма гребня есть результат взаимодействия двух рецессивных генов (ав).
Комплементарное взаимодействие Р ААвв аа. ВВ х Розовидный гребень (виандотты) F 1 х Гороховидный гребень (порода брамма, куры европейского происхождения) Аа. Вв Ореховидный гребень (куры малайского происхождения) F 2 9 А В _: 3 А_ вв : 3 аа В_ : 1 аавв орех розов горох лист Каждый ген имеет собственное фенотипическое проявление, розовидная и гороховидная формы гребня контролируются каждым доминантным геном в отдельности, Ав и а. В соответственно. Ореховидная форма - результат взаимодействия двух доминантных генов (АВ). Листовидная форма гребня есть результат взаимодействия двух рецессивных генов (ав).
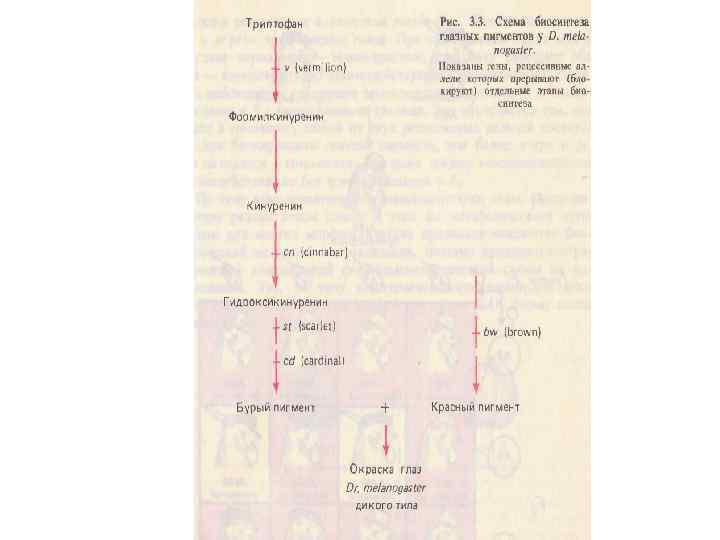
 Комплементарное взаимодействие Р bw+ st st Ярко- красные глаза х bw bw st+st+ Коричневые F 1 bw+ bw st +st Темно-красные (дикий тип) F 2 9 bw+ bw st+st : 3 bw+_ st st : 3 bw bw st+ _ : 1 bw bw st st Темно-красные Ярко- красные Коричневые Белые
Комплементарное взаимодействие Р bw+ st st Ярко- красные глаза х bw bw st+st+ Коричневые F 1 bw+ bw st +st Темно-красные (дикий тип) F 2 9 bw+ bw st+st : 3 bw+_ st st : 3 bw bw st+ _ : 1 bw bw st st Темно-красные Ярко- красные Коричневые Белые
 9: 7 Каждый доминантный ген в отдельности не обуславливает развитие окраски") Комплементарное взаимодействие б) 9: 7 Каждый доминантный ген в отдельности не обуславливает развитие окраски (Ав, а. В) Взаимодействие двух доминантных генов обуславливает развитие красной окраски (АВ). Отсутствие доминантных генов А и В одновременно также не обуславливает развитие окраски (ав). F 2 9 красных А_В_ : 7 белых [3 А_ вв + 3 аа. В_ + 1 аавв] Рис. 3. Наследование окраски цветков душистого горошка (Lathyrus odoratus) при взаимодействии двух пар генов (Лобашев, 1967. - С. 158)
Комплементарное взаимодействие б) 9: 7 Каждый доминантный ген в отдельности не обуславливает развитие окраски (Ав, а. В) Взаимодействие двух доминантных генов обуславливает развитие красной окраски (АВ). Отсутствие доминантных генов А и В одновременно также не обуславливает развитие окраски (ав). F 2 9 красных А_В_ : 7 белых [3 А_ вв + 3 аа. В_ + 1 аавв] Рис. 3. Наследование окраски цветков душистого горошка (Lathyrus odoratus) при взаимодействии двух пар генов (Лобашев, 1967. - С. 158)
 х Р 2 аа. ВВ (белые цветки) F 1") Р 1 ААbb (белые цветки) х Р 2 аа. ВВ (белые цветки) F 1 Аа. Вb - цветки пурпурной окраски АВ Ав а. В ав AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 9 пурпурных А_В_ : 7 белых [3 А_ вв + 3 аа. В_ + 1 аавв]
Р 1 ААbb (белые цветки) х Р 2 аа. ВВ (белые цветки) F 1 Аа. Вb - цветки пурпурной окраски АВ Ав а. В ав AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 9 пурпурных А_В_ : 7 белых [3 А_ вв + 3 аа. В_ + 1 аавв]
 9: 3: 4 § по окраске чешуи лука – 9 красных") Комплементарное взаимодействие в) 9: 3: 4 § по окраске чешуи лука – 9 красных : 3 желтых : 4 белых; § по окраске шерсти мышей – 9 агути, рыжевато-серая : 3 черных : 4 белых. v для хлорофильных мутаций у ячменя – 9 зеленых : 3 желтых «xantha» : 4 белых «albina» . ü Нормальный процесс образование хлорофилла требует присутствия серии доминантных аллелей (АВ). ü При отсутствии доминантного аллеля В (Ав) листья будут желтыми. ü При отсутствии доминантного аллеля А (а. В) листья будут белыми. ü Хлорофилл не будет синтезироваться и при одновременном отсутствии доминантных аллелей А и В (ав). Ø Тип потомства с одним доминантным геном (3 аа. В_ ) неотличим от двойного рецессива (1 аавв).
Комплементарное взаимодействие в) 9: 3: 4 § по окраске чешуи лука – 9 красных : 3 желтых : 4 белых; § по окраске шерсти мышей – 9 агути, рыжевато-серая : 3 черных : 4 белых. v для хлорофильных мутаций у ячменя – 9 зеленых : 3 желтых «xantha» : 4 белых «albina» . ü Нормальный процесс образование хлорофилла требует присутствия серии доминантных аллелей (АВ). ü При отсутствии доминантного аллеля В (Ав) листья будут желтыми. ü При отсутствии доминантного аллеля А (а. В) листья будут белыми. ü Хлорофилл не будет синтезироваться и при одновременном отсутствии доминантных аллелей А и В (ав). Ø Тип потомства с одним доминантным геном (3 аа. В_ ) неотличим от двойного рецессива (1 аавв).
 Характер расщепления при скрещивании различных форм ячменя, различающихся по окраске листьев Р 1 ААbb (жёлтые листья) х Р 2 аа. ВВ (белые листья) F 1 Аа. Вb - зелёные листья AB Ab a. B ab AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 9 зеленых А_ В_ : 3 желтых «xantha» А_ вв : 4 белых «albina» [3 аа. В_ + 1 аавв]
Характер расщепления при скрещивании различных форм ячменя, различающихся по окраске листьев Р 1 ААbb (жёлтые листья) х Р 2 аа. ВВ (белые листья) F 1 Аа. Вb - зелёные листья AB Ab a. B ab AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 9 зеленых А_ В_ : 3 желтых «xantha» А_ вв : 4 белых «albina» [3 аа. В_ + 1 аавв]
 9 : 6 : 1 - по данной формуле наследуется форма") Комплементарное взаимодействие г) 9 : 6 : 1 - по данной формуле наследуется форма плодов у тыквы. Р ААвв аа. ВВ Сферическая х Сферическая F 1 Аа. Вв Дисковидная F 2 9 А_ В_ дисков. : 6 сферич. [3 А_ вв + 3 аа. В_ ] : 1 аавв удлиненная Каждый доминантный ген в отдельности (Ав и а. В) дает сходный фенотипический эффект, что приводит к изменению характера расщепления в F 2, вследствие объединения в один фенотипический класс двух типов потомств (3 А_вв + 3 аа. В_). Взаимодействие доминантных генов (АВ) приводит к новообразованию и взаимодействие двух рецессивных генов (ав) также приводит к образованию новой формы плодов.
Комплементарное взаимодействие г) 9 : 6 : 1 - по данной формуле наследуется форма плодов у тыквы. Р ААвв аа. ВВ Сферическая х Сферическая F 1 Аа. Вв Дисковидная F 2 9 А_ В_ дисков. : 6 сферич. [3 А_ вв + 3 аа. В_ ] : 1 аавв удлиненная Каждый доминантный ген в отдельности (Ав и а. В) дает сходный фенотипический эффект, что приводит к изменению характера расщепления в F 2, вследствие объединения в один фенотипический класс двух типов потомств (3 А_вв + 3 аа. В_). Взаимодействие доминантных генов (АВ) приводит к новообразованию и взаимодействие двух рецессивных генов (ав) также приводит к образованию новой формы плодов.
 при взамодействии") Комплементарное взаимодействие Рис. 4. Наследование формы плода у фигурной тыквы (Cucurbita реро) при взамодействии двух пар генов (Лобашев, 1967. - С. I 64)
Комплементарное взаимодействие Рис. 4. Наследование формы плода у фигурной тыквы (Cucurbita реро) при взамодействии двух пар генов (Лобашев, 1967. - С. I 64)
 v Комплементарный тип взаимодействия генов иллюстрирует один из путей возникновения комбинативной изменчивости и имеет отношение к явлению гетерозиса.
v Комплементарный тип взаимодействия генов иллюстрирует один из путей возникновения комбинативной изменчивости и имеет отношение к явлению гетерозиса.
 – это такое взаимодействие неаллельных, когда один ген подавляет действие другого") Эпистаз (эпистатическое взаимодействие) – это такое взаимодействие неаллельных, когда один ген подавляет действие другого гена. Ген, подавляющий действие другого неаллельного гена и не имеющий собственного фенотипического проявления, называется ингибитором или супрессором и обозначается буквами I и S. У растений чаще ингибирующее действие оказывает доминантный ген неаллельной пары, имеющий собственное фенотипическое проявление эпистатический, а подавляемый – гипостатический. Для данного типа взаимодействия формулы расщепления: а) 12 : 3 : 1 б) 13 : 3 характерны следующие
Эпистаз (эпистатическое взаимодействие) – это такое взаимодействие неаллельных, когда один ген подавляет действие другого гена. Ген, подавляющий действие другого неаллельного гена и не имеющий собственного фенотипического проявления, называется ингибитором или супрессором и обозначается буквами I и S. У растений чаще ингибирующее действие оказывает доминантный ген неаллельной пары, имеющий собственное фенотипическое проявление эпистатический, а подавляемый – гипостатический. Для данного типа взаимодействия формулы расщепления: а) 12 : 3 : 1 б) 13 : 3 характерны следующие
 12 : 3 : 1 По типу эпистаза наследуется окраска плодов у") Эпистаз а) 12 : 3 : 1 По типу эпистаза наследуется окраска плодов у тыквы, окраска зерновки у кукурузы, форма лепестков у льна и др. Р F 1 ААвв Черная х Аа. Вв Черная аа. ВВ Серая (А > В) гипостатический эпистатический
Эпистаз а) 12 : 3 : 1 По типу эпистаза наследуется окраска плодов у тыквы, окраска зерновки у кукурузы, форма лепестков у льна и др. Р F 1 ААвв Черная х Аа. Вв Черная аа. ВВ Серая (А > В) гипостатический эпистатический
 АВ Ав а. В ав AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 12 [9 А_ В_ + 3 А_ вв] черных : 3 аа. В_ серых : 1 аавв белых Рассмотренное явление носит название доминантного эпистаза.
АВ Ав а. В ав AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 12 [9 А_ В_ + 3 А_ вв] черных : 3 аа. В_ серых : 1 аавв белых Рассмотренное явление носит название доминантного эпистаза.
 13 : 3 Один и тот же признак определяется независимыми доминантными и") Эпистаз в) 13 : 3 Один и тот же признак определяется независимыми доминантными и рецессивными генами. IС – белые (I > С) Iс - белые i. С - окрашенные iс - белые Р I IСС х Белые леггорны iiсс Белые плимутроки (минорки, виандотты) F 1 Ii. Сс Белые
Эпистаз в) 13 : 3 Один и тот же признак определяется независимыми доминантными и рецессивными генами. IС – белые (I > С) Iс - белые i. С - окрашенные iс - белые Р I IСС х Белые леггорны iiсс Белые плимутроки (минорки, виандотты) F 1 Ii. Сс Белые
 АВ Ав а. В ав AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 13 белые [9 I_С_ + 3 I_ сс + 1 iiсс] С_ : 3 окрашенные i i
АВ Ав а. В ав AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb F 2 13 белые [9 I_С_ + 3 I_ сс + 1 iiсс] С_ : 3 окрашенные i i
 Криптомерия - в ряде случаев отдельные гены не проявляют своего действия фенотипически, если не взаимодействуют с другими неаллельными генами. Ген, присутствие которого в генотипе необходимо для фенотипического проявления признака, можно назвать геном– проявителем. Так у льна доминантный аллель В определяет голубую окраску, рецессивный аллель в – розовую. Но для проявления необходимо наличие в генотипе доминантного аллеля А (проявителя). Его отсутствие в генотипе определяет белую окраску цветков. Р ААвв х аа. ВВ Розовоцветковые Белоцветковые растения F 1 Аа. Вв Голубоцветковые растения F 2 9 А_ В_ голубоцветковые : 3 розовоцветковые А_ вв : 4 розовоцветковые [3 аа. В_ + 1 аавв ]
Криптомерия - в ряде случаев отдельные гены не проявляют своего действия фенотипически, если не взаимодействуют с другими неаллельными генами. Ген, присутствие которого в генотипе необходимо для фенотипического проявления признака, можно назвать геном– проявителем. Так у льна доминантный аллель В определяет голубую окраску, рецессивный аллель в – розовую. Но для проявления необходимо наличие в генотипе доминантного аллеля А (проявителя). Его отсутствие в генотипе определяет белую окраску цветков. Р ААвв х аа. ВВ Розовоцветковые Белоцветковые растения F 1 Аа. Вв Голубоцветковые растения F 2 9 А_ В_ голубоцветковые : 3 розовоцветковые А_ вв : 4 розовоцветковые [3 аа. В_ + 1 аавв ]
 неаллельные гены, вызывающие сходные эффекты по проявлению одного") Полимерия (от греч. рolimeria – многомерность) неаллельные гены, вызывающие сходные эффекты по проявлению одного и того же признака, называются полимерными. Ø С данным типом взаимодействия генов связано наследование количественных признаков. Ø Явление открыто и экспериментально изучено шведским генетиком Нильссоном-Эле в 1909 г. Ø Полимерные гены принято обозначать их одной и той же буквой с указанием индекса для разных членов: R 1, R 2 и r 1, r 2.
Полимерия (от греч. рolimeria – многомерность) неаллельные гены, вызывающие сходные эффекты по проявлению одного и того же признака, называются полимерными. Ø С данным типом взаимодействия генов связано наследование количественных признаков. Ø Явление открыто и экспериментально изучено шведским генетиком Нильссоном-Эле в 1909 г. Ø Полимерные гены принято обозначать их одной и той же буквой с указанием индекса для разных членов: R 1, R 2 и r 1, r 2.
 Полимерия Р F 1 F 2 R 1 R 1 R 2 х r 1 r 1 r 2 темно-красное белое R 1 r 1 R 2 r 2 светло-красное 15 : 1 1/16 R 1 R 1 R 2 - темно-красных (два доминантных гена в двойной дозе) 4/16 R 1 R 2 r 2 - красных (три доминантных аллеля) 6/16 R 1 r 1 R 2 r 2 - светло-красных (два доминантных аллеля) 4/16 R 1 r 2 r 2 - бледно-красных (один доминантный аллель) 1/16 r 1 r 2 - белых (отсутствуют доминантные аллели) Интенсивность окраски зависит от дозы или числа доминантных аллелей. Гибриды F 1 обычно характеризуются промежуточным проявлением признака, в F 2 имеются плавные переходы между крайними вариантами, невозможно разграничить различные категории потомства.
Полимерия Р F 1 F 2 R 1 R 1 R 2 х r 1 r 1 r 2 темно-красное белое R 1 r 1 R 2 r 2 светло-красное 15 : 1 1/16 R 1 R 1 R 2 - темно-красных (два доминантных гена в двойной дозе) 4/16 R 1 R 2 r 2 - красных (три доминантных аллеля) 6/16 R 1 r 1 R 2 r 2 - светло-красных (два доминантных аллеля) 4/16 R 1 r 2 r 2 - бледно-красных (один доминантный аллель) 1/16 r 1 r 2 - белых (отсутствуют доминантные аллели) Интенсивность окраски зависит от дозы или числа доминантных аллелей. Гибриды F 1 обычно характеризуются промежуточным проявлением признака, в F 2 имеются плавные переходы между крайними вариантами, невозможно разграничить различные категории потомства.
 R 1 R 2 R 1 r 2 r 1 R 2 r 1 r 2 R 1 R 1 R 2 R 2 R 1 R 1 R 2 r 2 R 1 r 1 R 2 R 2 R 1 r 1 R 2 r 2 R 1 R 1 R 2 r 2 R 1 R 1 r 2 r 2 R 1 r 1 R 2 r 2 R 1 r 1 r 2 r 2 r 1 R 2 R 1 r 1 R 2 r 2 r 1 r 1 R 2 R 2 r 1 r 1 R 2 r 2 R 1 r 1 R 2 r 2 r 1 r 1 r 2 r 2 R 1 R 2 r 1 r 2 R 1 r 1 r 2 r 2 При дигенном контроле признака ряд распределения фенотипических классов имеет следующий вид - 1 : 4 : 6 : 4 : 1, что соответствует частотам генотипов с разным количеством доминантных генов, .
R 1 R 2 R 1 r 2 r 1 R 2 r 1 r 2 R 1 R 1 R 2 R 2 R 1 R 1 R 2 r 2 R 1 r 1 R 2 R 2 R 1 r 1 R 2 r 2 R 1 R 1 R 2 r 2 R 1 R 1 r 2 r 2 R 1 r 1 R 2 r 2 R 1 r 1 r 2 r 2 r 1 R 2 R 1 r 1 R 2 r 2 r 1 r 1 R 2 R 2 r 1 r 1 R 2 r 2 R 1 r 1 R 2 r 2 r 1 r 1 r 2 r 2 R 1 R 2 r 1 r 2 R 1 r 1 r 2 r 2 При дигенном контроле признака ряд распределения фенотипических классов имеет следующий вид - 1 : 4 : 6 : 4 : 1, что соответствует частотам генотипов с разным количеством доминантных генов, .
 Частоты генотипов с разным количеством доминантных генов Количество Частота доминантн рецессивных генотипов ых аллелей Фенотип Частота фенотипов 4 0 1 темно-красные 1 3 1 4 красные 4 2 2 6 светло-красное 6 1 3 4 Бледно-красные 4 0 4 1 белые 1
Частоты генотипов с разным количеством доминантных генов Количество Частота доминантн рецессивных генотипов ых аллелей Фенотип Частота фенотипов 4 0 1 темно-красные 1 3 1 4 красные 4 2 2 6 светло-красное 6 1 3 4 Бледно-красные 4 0 4 1 белые 1
 Полимерия Если признак «окраска зерна» контролируется тремя генами R, то характер расщепления в F 2 будет 63 : 1, а ряд распределения будет следующим 1 : 6 : 15 : 20 : 15 : 6 : 1. § Распределение фенотипических классов находится в соответствии с коэффициентом разложения бинома Ньютона (p +q)n , где n – число неаллельных генов. § По мере возрастания n, распределение коэффициентов разложения будет все более приближаться к нормальному распределению, потомство гибрида будет образовывать непрерывный вариационный ряд по фенотипическому проявлению данного признака, различия между разными фенотипическим классами будут менее заметными, которые стираются условиями внешней среды. § Большинство хозяйственно-ценных признаков растений и животных контролируется полимерными генами: урожайность, высота растений, число зерен, качество зерна, устойчивость к неблагоприятным факторам среды, полевая устойчивость к болезням, жирность молока, молочность скота, яйценоскость и т. д.
Полимерия Если признак «окраска зерна» контролируется тремя генами R, то характер расщепления в F 2 будет 63 : 1, а ряд распределения будет следующим 1 : 6 : 15 : 20 : 15 : 6 : 1. § Распределение фенотипических классов находится в соответствии с коэффициентом разложения бинома Ньютона (p +q)n , где n – число неаллельных генов. § По мере возрастания n, распределение коэффициентов разложения будет все более приближаться к нормальному распределению, потомство гибрида будет образовывать непрерывный вариационный ряд по фенотипическому проявлению данного признака, различия между разными фенотипическим классами будут менее заметными, которые стираются условиями внешней среды. § Большинство хозяйственно-ценных признаков растений и животных контролируется полимерными генами: урожайность, высота растений, число зерен, качество зерна, устойчивость к неблагоприятным факторам среды, полевая устойчивость к болезням, жирность молока, молочность скота, яйценоскость и т. д.
 Некумулятивная полимерия Пример: наследование формы плода у пастушьей сумки в соотношении 15 треугольных : 1 овальная. стручки треугольной формы х стручки овальной формы F 1 плоды F 2 треугольной формы 15 треугольных : 1 овальных. Для наличия треугольной формы достаточно хотя бы одного из доминантных генов А 1 и А 2 в одиночной дозе. А 1 А 1 А 2 А 2 А 1 А 2 а 2 А 1 А 1 а 2 а 2 А 1 а 1 А 2 а 2 А 1 а 1 а 2 а 2 а 1 а 1 а 2 а 2 - треугольная форма (15 /16) плоды будут овальными (1/16).
Некумулятивная полимерия Пример: наследование формы плода у пастушьей сумки в соотношении 15 треугольных : 1 овальная. стручки треугольной формы х стручки овальной формы F 1 плоды F 2 треугольной формы 15 треугольных : 1 овальных. Для наличия треугольной формы достаточно хотя бы одного из доминантных генов А 1 и А 2 в одиночной дозе. А 1 А 1 А 2 А 2 А 1 А 2 а 2 А 1 А 1 а 2 а 2 А 1 а 1 А 2 а 2 А 1 а 1 а 2 а 2 а 1 а 1 а 2 а 2 - треугольная форма (15 /16) плоды будут овальными (1/16).
 или ослабляющие (супрессоры) действие главного гена. Главные гены") Гены-модификаторы – это гены усиливающие (интенсификаторы) или ослабляющие (супрессоры) действие главного гена. Главные гены – это гены, определяющие развитие признака и свойства, например, выработку пигментов, наличие или отсутствие цианида, устойчивость или чувствительность к заболеваниям и т. д. У томатов рецессивный ген lsls вызывает в гомозиготном состоянии прекращение верхушечного роста после образования первого соцветия. Действие этого гена зависит от генотипической среды. lsls. ААВВ – ген работает lslsаа. ВВ – ген не работает Гены-модификаторы участвуют в создании небольших, но важных различий в таких признаках цветка, как окраска, форма или аромат, изменяют также внешний вид, вкус и аромат плодов. На действие среды. генов модификаторов могут оказывать влияние условия
Гены-модификаторы – это гены усиливающие (интенсификаторы) или ослабляющие (супрессоры) действие главного гена. Главные гены – это гены, определяющие развитие признака и свойства, например, выработку пигментов, наличие или отсутствие цианида, устойчивость или чувствительность к заболеваниям и т. д. У томатов рецессивный ген lsls вызывает в гомозиготном состоянии прекращение верхушечного роста после образования первого соцветия. Действие этого гена зависит от генотипической среды. lsls. ААВВ – ген работает lslsаа. ВВ – ген не работает Гены-модификаторы участвуют в создании небольших, но важных различий в таких признаках цветка, как окраска, форма или аромат, изменяют также внешний вид, вкус и аромат плодов. На действие среды. генов модификаторов могут оказывать влияние условия
 действие генов, вызывающее увеличение или уменьшение выраженности признака у") Трансгрессия – это аддитивное (суммирующее) действие генов, вызывающее увеличение или уменьшение выраженности признака у гибридов в сравнении с родительскими формами. Как правило, трансгрессия проявляется по количественным признакам. Р ААвв х аа. ВВ Длинный колос, Короткий колос, малоцветковый колосок многоцветковый колосок F 1 Аа. Вв Длинный колос, многоцветковый колосок F 2 ААВВ ……………. аавв Длинный колос, Короткий колос, многоцветковый колосок малоцветковый колосок положительная трансгрессия отрицательная трансгрессия Когда одинаковое проявление признака у родителей зависит о разных положительно действующих полимерных генов, то при скрещивании форм внешне не отличимых друг от друга можно ожидать у потомства усиление свойств или увеличение выраженности признака.
Трансгрессия – это аддитивное (суммирующее) действие генов, вызывающее увеличение или уменьшение выраженности признака у гибридов в сравнении с родительскими формами. Как правило, трансгрессия проявляется по количественным признакам. Р ААвв х аа. ВВ Длинный колос, Короткий колос, малоцветковый колосок многоцветковый колосок F 1 Аа. Вв Длинный колос, многоцветковый колосок F 2 ААВВ ……………. аавв Длинный колос, Короткий колос, многоцветковый колосок малоцветковый колосок положительная трансгрессия отрицательная трансгрессия Когда одинаковое проявление признака у родителей зависит о разных положительно действующих полимерных генов, то при скрещивании форм внешне не отличимых друг от друга можно ожидать у потомства усиление свойств или увеличение выраженности признака.
 – это свойство гена оказывать влияние на развитие") Плейотропия (от греч. рleistos – наибольший) – это свойство гена оказывать влияние на развитие нескольких признаков одновременно. Явление распространено как у растений, так и у животных. Многие гены плейотропны в отношении морфологических признаков. Ø Мендель обнаружил, что красная окраска цветков, красные пятна на прилистниках и серая окраска семенной кожуры детерминируются одним геном. Ø У высших растений окраска цветков нередко коррелирует с окраской стебля: сорта с красными цветками имеют красный стебель, тогда как у сортов с белыми цветами стебель зеленый. Ø У ячменя ген, контролирующий рыхлый колос, одновременно вызывает сокращение длины верхнего междоузлия. Ø Н. И. Вавилов и О. В. Якушкина при изучении генетических особенностей персидской пшеницы установили, что доминантный ген черной окраски всегда вызывает и сильное опушение чешуй. Ø Доминантный ген, вызывающий аномалию пальцев у человека (синдром Марфана) «паучьи пальцы» , обуславливает одновременно дефект хрусталика глаза и порок сердца.
Плейотропия (от греч. рleistos – наибольший) – это свойство гена оказывать влияние на развитие нескольких признаков одновременно. Явление распространено как у растений, так и у животных. Многие гены плейотропны в отношении морфологических признаков. Ø Мендель обнаружил, что красная окраска цветков, красные пятна на прилистниках и серая окраска семенной кожуры детерминируются одним геном. Ø У высших растений окраска цветков нередко коррелирует с окраской стебля: сорта с красными цветками имеют красный стебель, тогда как у сортов с белыми цветами стебель зеленый. Ø У ячменя ген, контролирующий рыхлый колос, одновременно вызывает сокращение длины верхнего междоузлия. Ø Н. И. Вавилов и О. В. Якушкина при изучении генетических особенностей персидской пшеницы установили, что доминантный ген черной окраски всегда вызывает и сильное опушение чешуй. Ø Доминантный ген, вызывающий аномалию пальцев у человека (синдром Марфана) «паучьи пальцы» , обуславливает одновременно дефект хрусталика глаза и порок сердца.
 Плейотропия Особенно часто встречается плейотропия, при которой гены одновременно оказывают влияние на жизнеспособность особи. q У мышей желтая (окраска) пигментация всегда сочетается с определенными изменениями физиологических функций: пониженная плодовитость, склонность к ожирению. Все три признака детерминированы одним геном. q У человека генетическое заболевание серповидноклеточная анемия вызывается гомозиготностью по рецессивному аллелю Hb. S. Отличие аномального гемоглобина от нормального: глутаминовая кислота в положении 6 заменена валином. Больные гемолитической анемией страдают и от частых болей в кишечнике и скелетных мышцах. Через несколько лет пониженное кровоснабжение приводит к некрозу органов. Такие симптомы, как анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга могут наблюдаться либо по отдельности, либо во всех возможных комбинациях. Все плейотропные эффекты обусловлены одним биохимическим действием гена.
Плейотропия Особенно часто встречается плейотропия, при которой гены одновременно оказывают влияние на жизнеспособность особи. q У мышей желтая (окраска) пигментация всегда сочетается с определенными изменениями физиологических функций: пониженная плодовитость, склонность к ожирению. Все три признака детерминированы одним геном. q У человека генетическое заболевание серповидноклеточная анемия вызывается гомозиготностью по рецессивному аллелю Hb. S. Отличие аномального гемоглобина от нормального: глутаминовая кислота в положении 6 заменена валином. Больные гемолитической анемией страдают и от частых болей в кишечнике и скелетных мышцах. Через несколько лет пониженное кровоснабжение приводит к некрозу органов. Такие симптомы, как анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга могут наблюдаться либо по отдельности, либо во всех возможных комбинациях. Все плейотропные эффекты обусловлены одним биохимическим действием гена.
, который кодирует гликопротеин фибрилин-1,") Синдром Марфана вызывается мутациями в гене FBN 1 (15 хромосома), который кодирует гликопротеин фибрилин-1, являющийся компонентом внеклеточного матрикса. Белок фибрилин-1 имеет важное значение для правильного формирования внеклеточного матрикса, играет определенную роль при биогенезе и влияет на функционирование эластичных волокон.
Синдром Марфана вызывается мутациями в гене FBN 1 (15 хромосома), который кодирует гликопротеин фибрилин-1, являющийся компонентом внеклеточного матрикса. Белок фибрилин-1 имеет важное значение для правильного формирования внеклеточного матрикса, играет определенную роль при биогенезе и влияет на функционирование эластичных волокон.
 Пенетрантность и экспрессивность Пенетрантность способность гена или комбинации генов проявлять себя тем или иным образом фенотипически Пенетрантность выражается долей особей (%) у которых исследуемый ген проявился фенотипически от числа особей, у которых он должен проявиться. При полной пенетрантности (100%) мутантный ген проявляет свое действие у каждой особи, обладающей им, при неполной пенетрантности (<100%) ген проявляет свой фенотипический эффект не у всех особей. При скрещивании красноцветковой примулы с белоцветковой в F 2 происходит моногибридное расщепление только при t = +15+25 3 : 1, при t = +30 -35 все цветки белоцветковые. В зависимости от температуры количество белых цветков у примулы колеблется от 25 до 100%. Ген, детерминирующий красноцветковость, фенотипически проявляется не у всех особей, в генотипе которых он присутствует. Неполно пенетрировать могут как рецессивные, так и доминантные мутации в гомо- и гетерозиготном состоянии.
Пенетрантность и экспрессивность Пенетрантность способность гена или комбинации генов проявлять себя тем или иным образом фенотипически Пенетрантность выражается долей особей (%) у которых исследуемый ген проявился фенотипически от числа особей, у которых он должен проявиться. При полной пенетрантности (100%) мутантный ген проявляет свое действие у каждой особи, обладающей им, при неполной пенетрантности (<100%) ген проявляет свой фенотипический эффект не у всех особей. При скрещивании красноцветковой примулы с белоцветковой в F 2 происходит моногибридное расщепление только при t = +15+25 3 : 1, при t = +30 -35 все цветки белоцветковые. В зависимости от температуры количество белых цветков у примулы колеблется от 25 до 100%. Ген, детерминирующий красноцветковость, фенотипически проявляется не у всех особей, в генотипе которых он присутствует. Неполно пенетрировать могут как рецессивные, так и доминантные мутации в гомо- и гетерозиготном состоянии.
 Пенетрантность и экспрессивность • Фенотипическое проявление гена может варьировать по степени выраженности признака. Для описания характера или степени выраженности (признака) гена, оказавшегося пенетрантным, употребляется термин экспрессивность. • Экспрессивность – это степень выраженности признака (предложил Н. В. Тимофеев-Ресовский в 1927 г. ). Экспрессивность обычно выражают количественно в зависимости от уклонения признака от дикого типа. • У дрозофилы имеется мутация редуцированным числом фасеток. еyeless «безглазая» с сильно У потомства одной из родительской пары глаза одних мух почти полностью лишены фасеток, тогда как у других мух число фасеток в глазах достигает половины нормального числа, то есть наблюдается различная степень редукции числа фасеток глаза, и может варьировать в больших пределах, достигая половины нормального числа фасеток.
Пенетрантность и экспрессивность • Фенотипическое проявление гена может варьировать по степени выраженности признака. Для описания характера или степени выраженности (признака) гена, оказавшегося пенетрантным, употребляется термин экспрессивность. • Экспрессивность – это степень выраженности признака (предложил Н. В. Тимофеев-Ресовский в 1927 г. ). Экспрессивность обычно выражают количественно в зависимости от уклонения признака от дикого типа. • У дрозофилы имеется мутация редуцированным числом фасеток. еyeless «безглазая» с сильно У потомства одной из родительской пары глаза одних мух почти полностью лишены фасеток, тогда как у других мух число фасеток в глазах достигает половины нормального числа, то есть наблюдается различная степень редукции числа фасеток глаза, и может варьировать в больших пределах, достигая половины нормального числа фасеток.
 Норма реакции генотипа – это специфическая способность генотипа тем или иным образом реагировать на изменения условий внешней среды, ограниченных возможностями данного организма. Амплитуда возможной выражает норму реакции. изменчивости в реализации генотипа Для каждого из генотипов норма реакция достаточно определенная. Выражение хозяйственно-ценных признаков сильно зависит от температурного, светового, пищевого, водного и других режимов. Например, сорт мягкой яровой пшеницы Саратовская 29 в засуху дает урожайность 12 ц/га, а в нормальных условиях – 32 ц/га. Варьирование урожайности обусловлено различными условиями, в которых происходила реализация генотипа, это и есть норма реакции.
Норма реакции генотипа – это специфическая способность генотипа тем или иным образом реагировать на изменения условий внешней среды, ограниченных возможностями данного организма. Амплитуда возможной выражает норму реакции. изменчивости в реализации генотипа Для каждого из генотипов норма реакция достаточно определенная. Выражение хозяйственно-ценных признаков сильно зависит от температурного, светового, пищевого, водного и других режимов. Например, сорт мягкой яровой пшеницы Саратовская 29 в засуху дает урожайность 12 ц/га, а в нормальных условиях – 32 ц/га. Варьирование урожайности обусловлено различными условиями, в которых происходила реализация генотипа, это и есть норма реакции.
 Выводы 1. Генотип представляет собой систему взаимодействующих генов, которые проявляются фенотипически от условий генотипической среды и условий внешний среды. Эта сложная система может быть разложена путем менделеевского на элементарные признаки фены и тем самым идентифицировать отдельные дискретные единицы – гены. 2. Между аллелями одного гена и между аллелями разных генов существуют сложные взаимодействия, которые приводят к изменению наследования признаков, их выраженности. 3. Все виды взаимодействия вызывают новообразования в проявлении признаков, имеющие важное селекционное и эволюционное значение.
Выводы 1. Генотип представляет собой систему взаимодействующих генов, которые проявляются фенотипически от условий генотипической среды и условий внешний среды. Эта сложная система может быть разложена путем менделеевского на элементарные признаки фены и тем самым идентифицировать отдельные дискретные единицы – гены. 2. Между аллелями одного гена и между аллелями разных генов существуют сложные взаимодействия, которые приводят к изменению наследования признаков, их выраженности. 3. Все виды взаимодействия вызывают новообразования в проявлении признаков, имеющие важное селекционное и эволюционное значение.