

Лек 3.ppt
- Количество слайдов: 54
 Изготовление парафиновых срезов
Изготовление парафиновых срезов
 Изготовление ультратонких срезов
Изготовление ультратонких срезов
 Лекция 3 Метод замороженных срезов Иммуногистохимия Криоэлектронная микроскопия Электронно-микроскопическая томография Атомно-силовая микроскопия Клетка Биомембраны Плазматическая мембрана (плазмалемма)
Лекция 3 Метод замороженных срезов Иммуногистохимия Криоэлектронная микроскопия Электронно-микроскопическая томография Атомно-силовая микроскопия Клетка Биомембраны Плазматическая мембрана (плазмалемма)
 Метод замороженных срезов Экспресс-диагностика Выявление ферментов Флуоресцентная иммуногистохимия Криостаты и замораживающие микротомы Микротом - криостат НМ 520 Leica LN 22 Замораживающий микротом на жидком азоте Замораживающий микротом Leica CM 1100 Сухая Портативный мобильный углекислота Криостат для рутинных работ, температура до -30°С
Метод замороженных срезов Экспресс-диагностика Выявление ферментов Флуоресцентная иммуногистохимия Криостаты и замораживающие микротомы Микротом - криостат НМ 520 Leica LN 22 Замораживающий микротом на жидком азоте Замораживающий микротом Leica CM 1100 Сухая Портативный мобильный углекислота Криостат для рутинных работ, температура до -30°С
 Приготовление криостатных срезов Криостатный срез нейронов кролика в культуре. Зеленый – дифференцированный нейрон ( «зрелый» ) Красные –клетки-предшественники Криостатный срез (16 мкм) почки мыши, обработка различными флуорохромами
Приготовление криостатных срезов Криостатный срез нейронов кролика в культуре. Зеленый – дифференцированный нейрон ( «зрелый» ) Красные –клетки-предшественники Криостатный срез (16 мкм) почки мыши, обработка различными флуорохромами
 Гистохимия ферментов. Гистохимическое выявление СДГ и НАДФНд на криостатных срезах мышц Выявление аквапорина-8 в клетках поджелудочной железы
Гистохимия ферментов. Гистохимическое выявление СДГ и НАДФНд на криостатных срезах мышц Выявление аквапорина-8 в клетках поджелудочной железы
 ИММУННОМЕЧЕНИЕ Иммуноцитохимия – выявление антигенов в клетках Иммуногистохимия – выявление антигенов в тканях и органах Иммунофлуоресцентная микроскопия Позволяет быстро выявить антигены в культуре клеток и тканях Яркая картинка Не позволяет точно дифференцировать клетки Светооптическая иммуномикроскопия Позволяет одновременно изучать локализацию антигена и структуру ткани, патологические изменения Позволяет точно идентифицировать клетки Интенсивность и цвет окрашивания антигена зависит от применяемых реагентов
ИММУННОМЕЧЕНИЕ Иммуноцитохимия – выявление антигенов в клетках Иммуногистохимия – выявление антигенов в тканях и органах Иммунофлуоресцентная микроскопия Позволяет быстро выявить антигены в культуре клеток и тканях Яркая картинка Не позволяет точно дифференцировать клетки Светооптическая иммуномикроскопия Позволяет одновременно изучать локализацию антигена и структуру ткани, патологические изменения Позволяет точно идентифицировать клетки Интенсивность и цвет окрашивания антигена зависит от применяемых реагентов
 Суть любой иммуногистохимической реакции – визуализация комплекса антиген-антитело Прямой метод Фермент Антитело Антиген связывается с антителом, которое конъюгировано с ферментом. В результате ферментативной реакции образуется окрашенный (видимый) продукт. Пероксидаза, кислая фосфатаза. Иммунофлуоресцентный вариант.
Суть любой иммуногистохимической реакции – визуализация комплекса антиген-антитело Прямой метод Фермент Антитело Антиген связывается с антителом, которое конъюгировано с ферментом. В результате ферментативной реакции образуется окрашенный (видимый) продукт. Пероксидаза, кислая фосфатаза. Иммунофлуоресцентный вариант.
.") Двойной непрямой метод Антиген фермент Антитела 1. Немеченые АТ связываются с АГ (первичные антитела). 2. Меченые ферментом вторичные АТ связываются с первичными АТ (они теперь выступают в качестве антигена)
Двойной непрямой метод Антиген фермент Антитела 1. Немеченые АТ связываются с АГ (первичные антитела). 2. Меченые ферментом вторичные АТ связываются с первичными АТ (они теперь выступают в качестве антигена)
 Тройной непрямой метод Антитела III Фермент Антитела II Антитела I Антиген Антитела II и III конъюгированы с одним и темже ферментом. Усиливается чувствительность, но возрастает количество ложно-положительных реакций.
Тройной непрямой метод Антитела III Фермент Антитела II Антитела I Антиген Антитела II и III конъюгированы с одним и темже ферментом. Усиливается чувствительность, но возрастает количество ложно-положительных реакций.
 Авидин-биотиновые методы Фермент Авидин Антитело II Биотин Антитело I Антиген Резко повышают чувствительность ИГХ реакции за счет высокой аффинности авидина (стрептавидина) к биотину. Авидин – белок, содержащийся в сырых куриных яйцах, конъюгирован с ферментом. Биотин – водорастворимый витамин Н (B 7), при биотинилировании ковалентно присоединяется к антителам
Авидин-биотиновые методы Фермент Авидин Антитело II Биотин Антитело I Антиген Резко повышают чувствительность ИГХ реакции за счет высокой аффинности авидина (стрептавидина) к биотину. Авидин – белок, содержащийся в сырых куриных яйцах, конъюгирован с ферментом. Биотин – водорастворимый витамин Н (B 7), при биотинилировании ковалентно присоединяется к антителам
 Негативная реакция На антигены ортопоксвирусов Печень мыши, Штамм ЕР-2 ВОК Позитивная реакция На антигены ортопоксвирусов Печень мыши Штамм К-2 Вируса эктромелии Иммунопероксидазная реакция
Негативная реакция На антигены ортопоксвирусов Печень мыши, Штамм ЕР-2 ВОК Позитивная реакция На антигены ортопоксвирусов Печень мыши Штамм К-2 Вируса эктромелии Иммунопероксидазная реакция
 ИММУННО-ЭЛЕКТРОННАЯ МИКРОСКОПИЯ Баланс между сохранением антигенной структуры и качеством фиксации - трудоемкий и дорогостоящий метод - требует специальной быстрой фиксации и процедуры обезвоживания и заключения в заливочную среду - для каждого конкретного случая требуется подбор разведений первичных и вторичных антител - не все антитела, «работающие» в ИФА, «работают» в иммуно-электронной микроскопии - малая толщина срезов лимитирует интенсивность метки - отсутствие осмирования снижает качество изображения
ИММУННО-ЭЛЕКТРОННАЯ МИКРОСКОПИЯ Баланс между сохранением антигенной структуры и качеством фиксации - трудоемкий и дорогостоящий метод - требует специальной быстрой фиксации и процедуры обезвоживания и заключения в заливочную среду - для каждого конкретного случая требуется подбор разведений первичных и вторичных антител - не все антитела, «работающие» в ИФА, «работают» в иммуно-электронной микроскопии - малая толщина срезов лимитирует интенсивность метки - отсутствие осмирования снижает качество изображения
 Ферритин Коллоидное золото
Ферритин Коллоидное золото
 Криоэлектронная микроскопия Сохранность нативной структуры макромолекул в водной среде - замораживание образцов - витрификация воды-льда - исследование в слое замороженной суспензии - криосрезы Leica EM HPM 100 Прибор для криофиксации путем заморозки под высоким давлением Получение изображений - устойчивость к воздействию электронного пучка и вакуума - повышение контраста
Криоэлектронная микроскопия Сохранность нативной структуры макромолекул в водной среде - замораживание образцов - витрификация воды-льда - исследование в слое замороженной суспензии - криосрезы Leica EM HPM 100 Прибор для криофиксации путем заморозки под высоким давлением Получение изображений - устойчивость к воздействию электронного пучка и вакуума - повышение контраста
 Процессор Leica EM AFS 2 Обеспечивает процесс замораживания-замещения и постепенного понижения температуры, полимеризацию заливочной среды. Процессор Leica EM GP для Электронный микроскоп приготовления для криомикроскопии витрифицированных Tecnai G 2 20 S-TWIN. и очень тонких образцов. Образец замораживается в жидком этане, на сетке
Процессор Leica EM AFS 2 Обеспечивает процесс замораживания-замещения и постепенного понижения температуры, полимеризацию заливочной среды. Процессор Leica EM GP для Электронный микроскоп приготовления для криомикроскопии витрифицированных Tecnai G 2 20 S-TWIN. и очень тонких образцов. Образец замораживается в жидком этане, на сетке
 Вирус гриппа Негативное Ультратонкий Контрастир-е срез Витрифицированный криосрез") Вирус гриппа Криопрепарат (в слое воды) Вирус гриппа Негативное Ультратонкий Контрастир-е срез Витрифицированный криосрез Mycobacterium bovis
Вирус гриппа Криопрепарат (в слое воды) Вирус гриппа Негативное Ультратонкий Контрастир-е срез Витрифицированный криосрез Mycobacterium bovis
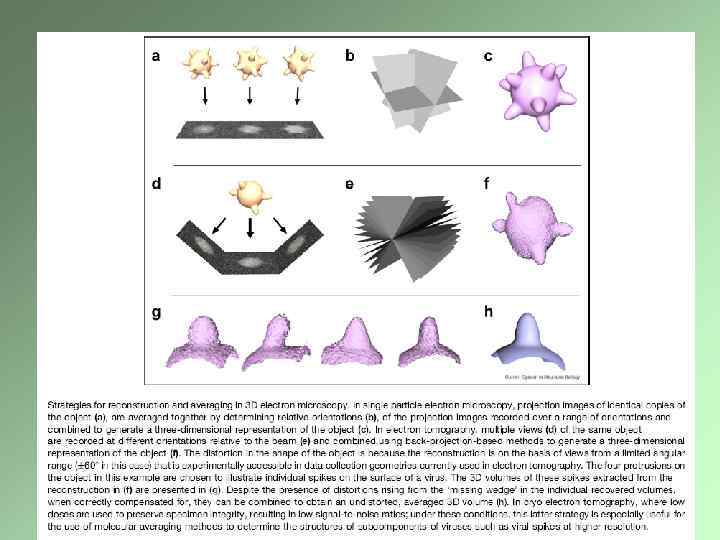
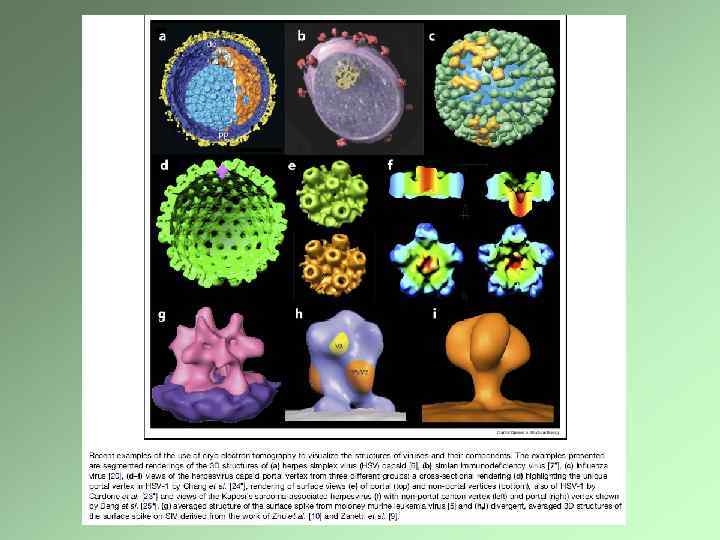
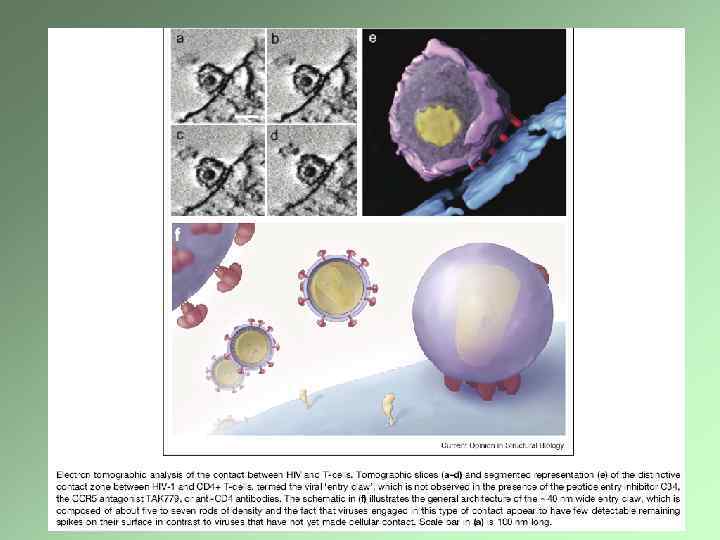
 Электронно-микроскопическая томография Создание трехмерного изображения на основе последовательных снимков, сделанных под разным углом Специальный гониометр Требования к препарату Компьютерное обеспечение Крио-электронная томография
Электронно-микроскопическая томография Создание трехмерного изображения на основе последовательных снимков, сделанных под разным углом Специальный гониометр Требования к препарату Компьютерное обеспечение Крио-электронная томография
 Атомно-силовая Микроскопия
Атомно-силовая Микроскопия
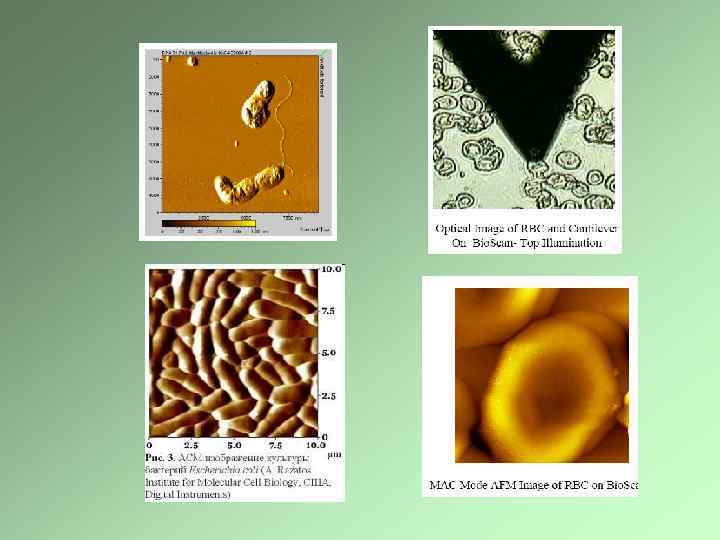
 Атомно-силовая микроскопия Вирус осповакцины Живые бактерии в водной среде
Атомно-силовая микроскопия Вирус осповакцины Живые бактерии в водной среде
 Изображение химической структуры молекулы пентацена, состоящей из 22 атомов углерода и 14 атомов водорода. Длина молекулы - 1, 4 нанометра, расстояние между соседними атомами углерода - 0, 14 нм. Получено специалистами из лаборатории IBM в Цюрихе методом бесконтактной атомно-силовой микроскопии. Микроскоп работал в сверхвысоком вакууме при - 268 °C. Необходимо было стабилизировать аппарат механически, так и термически, чтобы оба кончика AFM и молекула оставались в неизменном положении на протяжении более чем 20 часов.
Изображение химической структуры молекулы пентацена, состоящей из 22 атомов углерода и 14 атомов водорода. Длина молекулы - 1, 4 нанометра, расстояние между соседними атомами углерода - 0, 14 нм. Получено специалистами из лаборатории IBM в Цюрихе методом бесконтактной атомно-силовой микроскопии. Микроскоп работал в сверхвысоком вакууме при - 268 °C. Необходимо было стабилизировать аппарат механически, так и термически, чтобы оба кончика AFM и молекула оставались в неизменном положении на протяжении более чем 20 часов.
 Измерение силы взаимодействия между объектами Атомно-силовая спектроскопия
Измерение силы взаимодействия между объектами Атомно-силовая спектроскопия
 Выбор адекватного метода исследования – чрезвычайно важный момент, определяющий успех всего исследования И кто же это учил забивать гвозди микроскопом? ? !!
Выбор адекватного метода исследования – чрезвычайно важный момент, определяющий успех всего исследования И кто же это учил забивать гвозди микроскопом? ? !!
 Клетка 1665 г. - Роберт Гук - открыл клетки, наблюдая срез пробкового дерева в микроскоп, ввел понятие «клетка» от греч. «kytos» - полость «цитология»
Клетка 1665 г. - Роберт Гук - открыл клетки, наблюдая срез пробкового дерева в микроскоп, ввел понятие «клетка» от греч. «kytos» - полость «цитология»
 История развития клеточной теории 1665 г - Роберт Гук: открыл клетки, ввел понятие "клетка" 1680 г – А. Левенгук: открыл одноклеточные организмы 1831 г - Роберт Броун: открыл ядро в клетке 1838 г - Матиас Шлейден: ядро - неотъемлемая часть растительной клетки 1839 г - Томас Шванн: ядро - неотъемлемая часть животной клетки 1858 г - Рудольф Вирхов: суть происхождения клетки - деление ("Omnis cellula ex cellula" = «Каждая клетка - из клетки» )
История развития клеточной теории 1665 г - Роберт Гук: открыл клетки, ввел понятие "клетка" 1680 г – А. Левенгук: открыл одноклеточные организмы 1831 г - Роберт Броун: открыл ядро в клетке 1838 г - Матиас Шлейден: ядро - неотъемлемая часть растительной клетки 1839 г - Томас Шванн: ядро - неотъемлемая часть животной клетки 1858 г - Рудольф Вирхов: суть происхождения клетки - деление ("Omnis cellula ex cellula" = «Каждая клетка - из клетки» )
 Современная клеточная теория: 1. Все живые организмы состоят из клеток. Клетка – наименьшая структурная функциональная единица живого, основная единица строения и развития всех живых организмов; 2. Клетки всех одноклеточных и многоклеточных организмов сходны (гомологичны) по своему строению, химическому составу, основным проявлениям жизнедеятельности и обмену веществ. 3. Размножение клеток происходит путём их деления, и каждая новая клетка образуется в результате деления исходной клетки. 4. В многоклеточных организмах клетки специализированы по выполняемым ими функциям и образуют ткани; из тканей состоят органы. 5. Клеточное строение организмов – свидетельство, что все живые организмы имеют единое происхождение
Современная клеточная теория: 1. Все живые организмы состоят из клеток. Клетка – наименьшая структурная функциональная единица живого, основная единица строения и развития всех живых организмов; 2. Клетки всех одноклеточных и многоклеточных организмов сходны (гомологичны) по своему строению, химическому составу, основным проявлениям жизнедеятельности и обмену веществ. 3. Размножение клеток происходит путём их деления, и каждая новая клетка образуется в результате деления исходной клетки. 4. В многоклеточных организмах клетки специализированы по выполняемым ими функциям и образуют ткани; из тканей состоят органы. 5. Клеточное строение организмов – свидетельство, что все живые организмы имеют единое происхождение
 Клетка – это элементарная живая система, способная к самообновлению, саморегуляции и самовоспроизведению В природе НЕ СУЩЕСТВУЕТ «универсальной» клетки Наука «цитология» оперирует неким собирательным образом клетки Клеточные и неклеточные формы существования живой материи
Клетка – это элементарная живая система, способная к самообновлению, саморегуляции и самовоспроизведению В природе НЕ СУЩЕСТВУЕТ «универсальной» клетки Наука «цитология» оперирует неким собирательным образом клетки Клеточные и неклеточные формы существования живой материи
 событие биологической эволюции.") Появление эукариотической клетки – второе по значимости (после зарождения самой жизни) событие биологической эволюции. Важнейшее отличие эукариот от прокариот - более совершенная система регуляции генома (в этом - смысл появления клеточного ядра). Область активного метаболизма – цитоплазма – отделилась от области хранения, считывания, репликации генетической информации и, главное, - регуляции транскрипции и посттранскрипционных модификаций РНК. Благодаря этому резко возросла приспособляемость одноклеточных организмов, их способность адаптироваться к меняющимся условиям БЕЗ внесения наследственных изменений в геном. Эукариоты появились в результате симбиоза нескольких разновидностей прокариот (бактерий).
Появление эукариотической клетки – второе по значимости (после зарождения самой жизни) событие биологической эволюции. Важнейшее отличие эукариот от прокариот - более совершенная система регуляции генома (в этом - смысл появления клеточного ядра). Область активного метаболизма – цитоплазма – отделилась от области хранения, считывания, репликации генетической информации и, главное, - регуляции транскрипции и посттранскрипционных модификаций РНК. Благодаря этому резко возросла приспособляемость одноклеточных организмов, их способность адаптироваться к меняющимся условиям БЕЗ внесения наследственных изменений в геном. Эукариоты появились в результате симбиоза нескольких разновидностей прокариот (бактерий).
 Клетки эукариот имеют сложную пространственную организацию, В основе которой лежит принцип компартментализации Клетка должна поддерживать свою жизнедеятельность, адекватно отвечать на внешние сигналы. В клетке постоянно идут процессы перемещения структур, Процессы деградации и обновления органоидов. Ядро – репродукция клетки и синтез рибосом Синтез белков – рибосомы, прикрепленные и свободные Аппарат Гольджи – главная «сортировочная станция» , система коммуникаций Митохондрии – выработка энергии, дыхание Эндосомально-лизосомальный аппарат – уничтожение «состарившихся» структур и попавших в клетку веществ Цитоскелет – опорные структуры и структуры, обеспечивающие перемещение органоидов Эндоцитозные структуры – система селективного попадания в клетку макромолекул и их утилизации
Клетки эукариот имеют сложную пространственную организацию, В основе которой лежит принцип компартментализации Клетка должна поддерживать свою жизнедеятельность, адекватно отвечать на внешние сигналы. В клетке постоянно идут процессы перемещения структур, Процессы деградации и обновления органоидов. Ядро – репродукция клетки и синтез рибосом Синтез белков – рибосомы, прикрепленные и свободные Аппарат Гольджи – главная «сортировочная станция» , система коммуникаций Митохондрии – выработка энергии, дыхание Эндосомально-лизосомальный аппарат – уничтожение «состарившихся» структур и попавших в клетку веществ Цитоскелет – опорные структуры и структуры, обеспечивающие перемещение органоидов Эндоцитозные структуры – система селективного попадания в клетку макромолекул и их утилизации
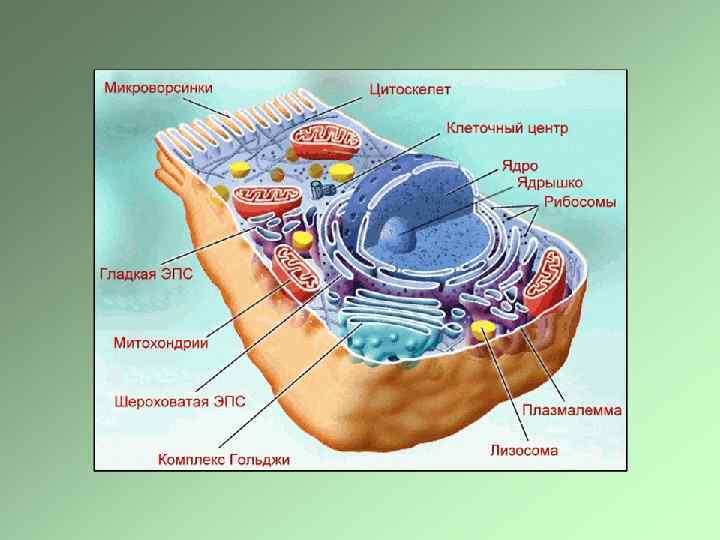
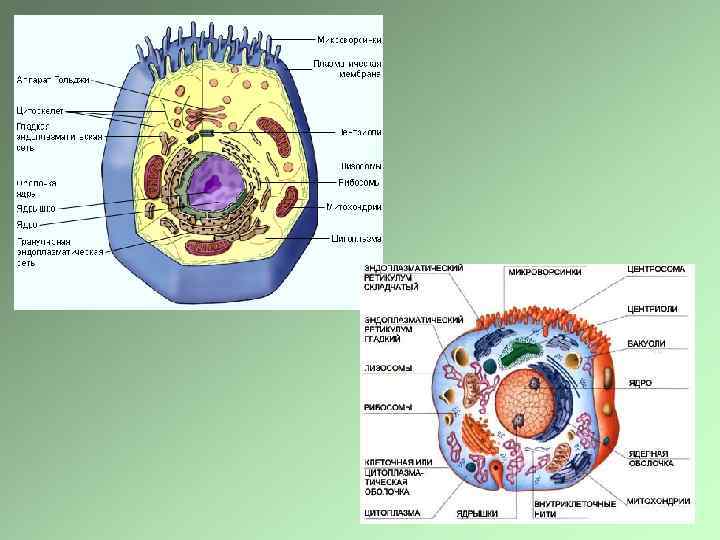
 Фосфолипидная мембрана – основа компартментализации живой материи, основа существования клетки
Фосфолипидная мембрана – основа компартментализации живой материи, основа существования клетки
 «Биологическая» мембрана
«Биологическая» мембрана
 – основная часть мембран. фосфатидилхолин фосфатидилэтаноламин фосфатидилсерин кардиолипин фосфатидилинозитол cфингомиелин") ФОСФОЛИПИДЫ (фосфоглицериды) – основная часть мембран. фосфатидилхолин фосфатидилэтаноламин фосфатидилсерин кардиолипин фосфатидилинозитол cфингомиелин
ФОСФОЛИПИДЫ (фосфоглицериды) – основная часть мембран. фосфатидилхолин фосфатидилэтаноламин фосфатидилсерин кардиолипин фосфатидилинозитол cфингомиелин
 Молекулы холестерола ориентируются в бислое так, чтобы их гидроксильные группы примыкали к") Холестерол (холестерин) Молекулы холестерола ориентируются в бислое так, чтобы их гидроксильные группы примыкали к полярным головам фосфолипидных молекул. При этом плоские стероидные кольца молекул холестерола частично иммобилизуют участки углеводородных цепей, непосредственно примыкающих к полярным головам. В мембране эритроцитов 23% холестерина
Холестерол (холестерин) Молекулы холестерола ориентируются в бислое так, чтобы их гидроксильные группы примыкали к полярным головам фосфолипидных молекул. При этом плоские стероидные кольца молекул холестерола частично иммобилизуют участки углеводородных цепей, непосредственно примыкающих к полярным головам. В мембране эритроцитов 23% холестерина
 Чистые фосфолипидные бислои спонтанно формируют замкнутые структуры, разграничивающие два водных компартмента. Пленки из фосфолипидных бислоев нестабильны, тогда как замкнутые сферические структуры устойчивы Замкнутые фосфолипидные структуры - липосомы Сложные системы на основе липосом – экспериментальные модели для изучения транспорта через мембрану и пр. Асимметрия состава внутри и вне липосом Липосомы – транспортные системы – переносчики
Чистые фосфолипидные бислои спонтанно формируют замкнутые структуры, разграничивающие два водных компартмента. Пленки из фосфолипидных бислоев нестабильны, тогда как замкнутые сферические структуры устойчивы Замкнутые фосфолипидные структуры - липосомы Сложные системы на основе липосом – экспериментальные модели для изучения транспорта через мембрану и пр. Асимметрия состава внутри и вне липосом Липосомы – транспортные системы – переносчики
 Липосомы Негативное контрастирование
Липосомы Негативное контрастирование
 Липосомы Негативное контрастирование
Липосомы Негативное контрастирование
 В основе всех клеточных мембран - фосфолипидный бислой Мембраны всегда ограничивают замкнутое пространство Толщина клеточных мембран 6 --12 нм
В основе всех клеточных мембран - фосфолипидный бислой Мембраны всегда ограничивают замкнутое пространство Толщина клеточных мембран 6 --12 нм
 Жидкостно-мозаичная модель строения мембраны
Жидкостно-мозаичная модель строения мембраны
 4 Цитоплазма 1 2 3 5 Способы «заякоривания» белков в мембране. 1. Монотопный белок, а-спиральный домен которого пронизывает мембрану. 2. Политопный белок, пронизывающий мембрану несколько раз. 3. Белок, присоединенный к мембране через жирную кислоту. 4. Белок, присоединенный к мембране через жирную кислоту и углеводную молекулу 5. Периферические белки не имеют гидрофобных областей, но содержат заряженные части, взаимодействующие с другими белками или фосфатными группами фосфолипидов.
4 Цитоплазма 1 2 3 5 Способы «заякоривания» белков в мембране. 1. Монотопный белок, а-спиральный домен которого пронизывает мембрану. 2. Политопный белок, пронизывающий мембрану несколько раз. 3. Белок, присоединенный к мембране через жирную кислоту. 4. Белок, присоединенный к мембране через жирную кислоту и углеводную молекулу 5. Периферические белки не имеют гидрофобных областей, но содержат заряженные части, взаимодействующие с другими белками или фосфатными группами фосфолипидов.
 строение. Четырехокись осмия связывается с полярными группами фосфолипидов") В ЭМ мембраны имеют трехлинейное (трехконтурное) строение. Четырехокись осмия связывается с полярными группами фосфолипидов по двойным связям, а гидрофобные концы молекул, формирующие светлую часть, не «окрашиваются» .
В ЭМ мембраны имеют трехлинейное (трехконтурное) строение. Четырехокись осмия связывается с полярными группами фосфолипидов по двойным связям, а гидрофобные концы молекул, формирующие светлую часть, не «окрашиваются» .
 Видны внутримембранные частицы.") Скол мембран микроворсинок (быстрое замораживание-скалывание) Видны внутримембранные частицы.
Скол мембран микроворсинок (быстрое замораживание-скалывание) Видны внутримембранные частицы.
 Гликофорин А содержится в мембранах эритроцитов, – первый изученный интегральный белок. Содержит 24 аминокислоты в а-спирали, которые формируют трансмембранный домен.
Гликофорин А содержится в мембранах эритроцитов, – первый изученный интегральный белок. Содержит 24 аминокислоты в а-спирали, которые формируют трансмембранный домен.
 Бактериородопсин
Бактериородопсин
 Некоторые свойства клеточных мембран В мембранах в среднем на одну молекулу белка приходится 50 молекул фосфолипидов, весовое соотношение – около 1: 1 Фосфолипиды и гликолипиды могут вращаться вокруг своей оси и перемещаться в латеральной плоскости. Средняя липидная молекула меняется местами с соседними около 107 раз в секунду, и перемещается за секунду на несколько мкм при 37 °C. Т. о. липидная молекула диффундирует по всей длине бактериальной клетки за 1 секунду, а по длине эукариотической клетки – за 20 сек. Липиды с ненасыщенными связями претерпевают фазовые переходы при более низких температурах, чем липиды с насыщенными связями. Гидрофобный внутренний слой мембран имеет небольшую вязкость и скорее жидкое, чем гелеобразное состояние. С уменьшением температуры, в мембранах повышается количество ненасыщенных жирных кислот – адаптация.
Некоторые свойства клеточных мембран В мембранах в среднем на одну молекулу белка приходится 50 молекул фосфолипидов, весовое соотношение – около 1: 1 Фосфолипиды и гликолипиды могут вращаться вокруг своей оси и перемещаться в латеральной плоскости. Средняя липидная молекула меняется местами с соседними около 107 раз в секунду, и перемещается за секунду на несколько мкм при 37 °C. Т. о. липидная молекула диффундирует по всей длине бактериальной клетки за 1 секунду, а по длине эукариотической клетки – за 20 сек. Липиды с ненасыщенными связями претерпевают фазовые переходы при более низких температурах, чем липиды с насыщенными связями. Гидрофобный внутренний слой мембран имеет небольшую вязкость и скорее жидкое, чем гелеобразное состояние. С уменьшением температуры, в мембранах повышается количество ненасыщенных жирных кислот – адаптация.
 Функции клеточных мембран 1. Барьерная функция: создание концентрационных градиентов, препятствие свободной диффузии. Мембрана принимает участие в механизмах электрогенеза: генерация потенциала покоя, потенциала действия, распространение биоэлектрических импульсов по однородной и неоднородной возбудимым структурам. 2. Регуляторная функция: тонкая регуляция внутриклеточного содержимого и внутриклеточных реакций путем рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и к запуску механизмов вторичных «месенджеров» ( «посредников» ). 3. Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах). 4. Высвобождение нейромедиаторов в синаптических окончаниях.
Функции клеточных мембран 1. Барьерная функция: создание концентрационных градиентов, препятствие свободной диффузии. Мембрана принимает участие в механизмах электрогенеза: генерация потенциала покоя, потенциала действия, распространение биоэлектрических импульсов по однородной и неоднородной возбудимым структурам. 2. Регуляторная функция: тонкая регуляция внутриклеточного содержимого и внутриклеточных реакций путем рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и к запуску механизмов вторичных «месенджеров» ( «посредников» ). 3. Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах). 4. Высвобождение нейромедиаторов в синаптических окончаниях.


