

Istoriya_vidkrittya_virusiv_Avtosokhranenny.pptx
- Количество слайдов: 30
 Історія відкриття вірусів. Віруси, віроїди, пріони. Морфологія та ультраструктура вірусів. Бактеріофаги
Історія відкриття вірусів. Віруси, віроїди, пріони. Морфологія та ультраструктура вірусів. Бактеріофаги
 Історія відкриття вірусів Перші згадки про вірусних хворобах людей і тварин в дійшли до нас з письмових джерел древніх народів. У них, зокрема, містяться відомості про епізоотії сказу у вовків, шакалів і собак і поліомієліті в Стародавньому Єгипті (II III тис. років до н. Е. . ). Про чорну віспу було відомо в Китаї за тисячу років до нашої ери. Давню історію має також жовта лихоманка, від якої протягом століть помирали білі першопрохідці у тропічній Африці і моряки. Перші описи вірусних хвороб у рослин відносяться до живописної «пестролепестности» тюльпанів, які вже близько 500 років вирощують голландські квітникарі. Початком становлення вірусології як науки можна вважати кінець XIX століття. У 80 х роках XIX століття, працюючи над створенням вакцини проти сказу, Луї Пастер (1822 1895) вперше застосував термін вірус (від лат. virus, отрута) для позначення інфекційного агента. Пастер був першим, хто почав використовувати лабораторних тварин в роботах з вивчення вірусу. Він проводив дослідження з інокуляції матеріалу, отриманого від хворих на сказ, в мозок кролика. Однак Пастер не робив відмінності між вірусами як такими і іншими інфекційними агентами.
Історія відкриття вірусів Перші згадки про вірусних хворобах людей і тварин в дійшли до нас з письмових джерел древніх народів. У них, зокрема, містяться відомості про епізоотії сказу у вовків, шакалів і собак і поліомієліті в Стародавньому Єгипті (II III тис. років до н. Е. . ). Про чорну віспу було відомо в Китаї за тисячу років до нашої ери. Давню історію має також жовта лихоманка, від якої протягом століть помирали білі першопрохідці у тропічній Африці і моряки. Перші описи вірусних хвороб у рослин відносяться до живописної «пестролепестности» тюльпанів, які вже близько 500 років вирощують голландські квітникарі. Початком становлення вірусології як науки можна вважати кінець XIX століття. У 80 х роках XIX століття, працюючи над створенням вакцини проти сказу, Луї Пастер (1822 1895) вперше застосував термін вірус (від лат. virus, отрута) для позначення інфекційного агента. Пастер був першим, хто почав використовувати лабораторних тварин в роботах з вивчення вірусу. Він проводив дослідження з інокуляції матеріалу, отриманого від хворих на сказ, в мозок кролика. Однак Пастер не робив відмінності між вірусами як такими і іншими інфекційними агентами.
 Луї Пастер Дмитро Іванович Івановський Першим, хто виділив віруси як конкретну групу інфекційних агентів, був російський вчений Дмитро Іванович Івановський (1864 1920). У 1892 році в результаті проведених досліджень Д. І. Івановський дійшов висновку, що мозаїчна хвороба тютюну викликається мікроорганізмами, що проходять через фільтр Шамберлана, які крім того не спроможні рости на штучних субстратах. Представлені дані про збудника тютюнової мозаїки (ВТМ) потім тривалий час були критеріями для віднесення збудників хвороб до «вірусів» : фільтрованість через «бактеріальні» фільтри, нездатність рости на штучних середовищах, загибель в субстраті бактерій і грибів в присутності вірусів. У 1898 році Мартін Бейерінк (1851 1931) підтвердив і розширив дослідження Івановського і сформулював першу повноцінну теорію про віруси як про новий клас мікроорганізмів.
Луї Пастер Дмитро Іванович Івановський Першим, хто виділив віруси як конкретну групу інфекційних агентів, був російський вчений Дмитро Іванович Івановський (1864 1920). У 1892 році в результаті проведених досліджень Д. І. Івановський дійшов висновку, що мозаїчна хвороба тютюну викликається мікроорганізмами, що проходять через фільтр Шамберлана, які крім того не спроможні рости на штучних субстратах. Представлені дані про збудника тютюнової мозаїки (ВТМ) потім тривалий час були критеріями для віднесення збудників хвороб до «вірусів» : фільтрованість через «бактеріальні» фільтри, нездатність рости на штучних середовищах, загибель в субстраті бактерій і грибів в присутності вірусів. У 1898 році Мартін Бейерінк (1851 1931) підтвердив і розширив дослідження Івановського і сформулював першу повноцінну теорію про віруси як про новий клас мікроорганізмів.
 Віруси — неклітинні форми життя, які являють собою автономні генетичні системи, нездатні до самостійного існування поза організмом або клітиною хазяїна, тобто є облігантними внутрішньоклітинними паразитами. Віруси займають суміжне положення між живою і неживою матерією. До основних рис, що відрізняють їх від живих організмів, належать: відсутність клітинної будови; відсутність власної білок синтезуючої системи; геном вірусів може бути представлений не тільки ДНК, але й РНК; деякі віруси можуть формувати всередині клітини кристали. Як і всі живі об’єкти, віруси здатні: до розмноження; успадковування ознак; генетичної і фенотипічної мінливості; адаптації до умов навколишнього середовища. Віруси класифікуються на ті, що містять ДНК (вірус простого герпесу) і ті, що містять РНК (вірус імунодефіциту людини). За структурою капсомерів. Ізометричні (кубічні), спіральні, змішані. За наявністю або відсутністю додаткової ліпопротеїнової оболонки (суперкапсиду) віруси поділяються на прості та складні. За клітинами хазяїнами
Віруси — неклітинні форми життя, які являють собою автономні генетичні системи, нездатні до самостійного існування поза організмом або клітиною хазяїна, тобто є облігантними внутрішньоклітинними паразитами. Віруси займають суміжне положення між живою і неживою матерією. До основних рис, що відрізняють їх від живих організмів, належать: відсутність клітинної будови; відсутність власної білок синтезуючої системи; геном вірусів може бути представлений не тільки ДНК, але й РНК; деякі віруси можуть формувати всередині клітини кристали. Як і всі живі об’єкти, віруси здатні: до розмноження; успадковування ознак; генетичної і фенотипічної мінливості; адаптації до умов навколишнього середовища. Віруси класифікуються на ті, що містять ДНК (вірус простого герпесу) і ті, що містять РНК (вірус імунодефіциту людини). За структурою капсомерів. Ізометричні (кубічні), спіральні, змішані. За наявністю або відсутністю додаткової ліпопротеїнової оболонки (суперкапсиду) віруси поділяються на прості та складні. За клітинами хазяїнами
 складаються з нуклеїнової кислоти, поміщеної в білкову або") БУДОВА ВІРУСІВ Зрілі вірусні частинки (віріони) складаються з нуклеїнової кислоти, поміщеної в білкову або ліпопротеїнову (білок у комплексі з ліпідами) оболонку. До складу вірусів входить один з двох видів нуклеїнових кислот — ДНК або РНК; ця ознака є основою їх класифікації на ДНК вмісні та РНК вмісні. У свою чергу, обидві групи поділяють на одноланцюгові і дволанцюгові. Низькомолекулярні білки зв’язуються з нуклеїновою кислотою, утворюючи чохол — капсид. Багато вірусів мають ще одну оболонку, розташовану зовні капсида — пеплос. Пеплос складається з високомолекулярних білків, організованих у вирости — пепломери, які служать для розпізнавання клітин мішеней. Крім білків, до складу пеплоса входять ліпіди та вуглеводи. Білки капсида і пеплоса виконують такі функції: стабілізують і захищають нуклеїнову кислоту; є ферментами, що беруть участь у відтворенні вірусної частинки; розпізнають відповідну клітину мішень.
БУДОВА ВІРУСІВ Зрілі вірусні частинки (віріони) складаються з нуклеїнової кислоти, поміщеної в білкову або ліпопротеїнову (білок у комплексі з ліпідами) оболонку. До складу вірусів входить один з двох видів нуклеїнових кислот — ДНК або РНК; ця ознака є основою їх класифікації на ДНК вмісні та РНК вмісні. У свою чергу, обидві групи поділяють на одноланцюгові і дволанцюгові. Низькомолекулярні білки зв’язуються з нуклеїновою кислотою, утворюючи чохол — капсид. Багато вірусів мають ще одну оболонку, розташовану зовні капсида — пеплос. Пеплос складається з високомолекулярних білків, організованих у вирости — пепломери, які служать для розпізнавання клітин мішеней. Крім білків, до складу пеплоса входять ліпіди та вуглеводи. Білки капсида і пеплоса виконують такі функції: стабілізують і захищають нуклеїнову кислоту; є ферментами, що беруть участь у відтворенні вірусної частинки; розпізнають відповідну клітину мішень.
, які називаються капсомерами. Їх кількість стабільна для кожного") Капсиди утворені з білкових субодиниць (поліпептидів), які називаються капсомерами. Їх кількість стабільна для кожного виду вірусів і використовується як таксономічна ознака. Віруси з таким типом будови називають простими. До простих належать найдрібніші з патогенних вірусів людини: поліовіруси, аденовіруси, паповавіруси.
Капсиди утворені з білкових субодиниць (поліпептидів), які називаються капсомерами. Їх кількість стабільна для кожного виду вірусів і використовується як таксономічна ознака. Віруси з таким типом будови називають простими. До простих належать найдрібніші з патогенних вірусів людини: поліовіруси, аденовіруси, паповавіруси.
 Більшість вірусів має ще одну оболонку – суперкапсидну, яка містить ліпіди. Вона пронизана вірусспецифічними білками глікопротеїдами, які на поверхні оболонки утворюють особливі стуктури, що називаються шипами. Такі віруси називають складними або оболонковими. Структурною одиницею суперкапсидної оболонки є пепломер. До них належать віруси сказу, герпесу, грипу, енцефалітів, імунодефіциту людини та ін.
Більшість вірусів має ще одну оболонку – суперкапсидну, яка містить ліпіди. Вона пронизана вірусспецифічними білками глікопротеїдами, які на поверхні оболонки утворюють особливі стуктури, що називаються шипами. Такі віруси називають складними або оболонковими. Структурною одиницею суперкапсидної оболонки є пепломер. До них належать віруси сказу, герпесу, грипу, енцефалітів, імунодефіциту людини та ін.
 Віріони характеризуються поняттям симетрії. Тип симетрії залежить від способу укладки нуклеїнової кислоти і, відповідно, розташування капсомерів навколо неї. Виділяють ізометричний (або кубічний), спіральний та змішаний типи симетрії. Кубічний тип характеризується тим, що капсомери утворюють багато гранник (найчастіше ікосаедр – 20 гранник). Форм У віріоні зі спіральною симетрією молекула нуклеїнової кислоти закручена разом із капсомерами в тугу спіраль. Такий тип симетрії мають віруси мозаїчної хвороби тютюну, грипу, кору, епідемічного паротиту Комбінований тип симетрії спостерігається у деяких бактеріофагів. При цьому головка бактеріофага має кубічний тип симетрії, а нуклеопротеїд, розміщений у хвості, укладається спірально.
Віріони характеризуються поняттям симетрії. Тип симетрії залежить від способу укладки нуклеїнової кислоти і, відповідно, розташування капсомерів навколо неї. Виділяють ізометричний (або кубічний), спіральний та змішаний типи симетрії. Кубічний тип характеризується тим, що капсомери утворюють багато гранник (найчастіше ікосаедр – 20 гранник). Форм У віріоні зі спіральною симетрією молекула нуклеїнової кислоти закручена разом із капсомерами в тугу спіраль. Такий тип симетрії мають віруси мозаїчної хвороби тютюну, грипу, кору, епідемічного паротиту Комбінований тип симетрії спостерігається у деяких бактеріофагів. При цьому головка бактеріофага має кубічний тип симетрії, а нуклеопротеїд, розміщений у хвості, укладається спірально.
, яка становить від 1") Віруси містять лише один тип нуклеїнової кислоти (ДНК або РНК), яка становить від 1 до 40 % маси віріона. Вірусні геноми містять інформацію, достатню для синтезу лише декількох білків. Віруси характеризуються надзвичайним розмаїттям форм геному. Він може бути представлений як односпіральними, так і двоспіральними молекулами, бути лінійним, циркулярним або фрагментованим. Білки вірусів (70 90 % маси віріона) поділяються на структурні та неструктурні. Структурними називають такі білки, які входять до складу зрілих позаклітинних вірусів. Вони виконують ряд важливих функцій: захищають нуклеїнову кислоту від зовнішнього пошкодження, взаємодіють з мембранами чутливих клітин, і забезпечують проникнення вірусу в клітину, мають РНК і ДНК полімеразну активність та ін. Неструктурні білки не входять до складу зрілих вірусів, однак утворюються під час їх репродукції. Вони забезпечують регуляцію експресії вірусного геному, є попередниками вірусних білків, здатні пригнічувати клітинний біосинтез. Залежно від розташування у віріоні, білки поділяються на капсидні, суперкапсидні, матриксні, білки серцевини та асоційовані з нуклеїновою кислотою.
Віруси містять лише один тип нуклеїнової кислоти (ДНК або РНК), яка становить від 1 до 40 % маси віріона. Вірусні геноми містять інформацію, достатню для синтезу лише декількох білків. Віруси характеризуються надзвичайним розмаїттям форм геному. Він може бути представлений як односпіральними, так і двоспіральними молекулами, бути лінійним, циркулярним або фрагментованим. Білки вірусів (70 90 % маси віріона) поділяються на структурні та неструктурні. Структурними називають такі білки, які входять до складу зрілих позаклітинних вірусів. Вони виконують ряд важливих функцій: захищають нуклеїнову кислоту від зовнішнього пошкодження, взаємодіють з мембранами чутливих клітин, і забезпечують проникнення вірусу в клітину, мають РНК і ДНК полімеразну активність та ін. Неструктурні білки не входять до складу зрілих вірусів, однак утворюються під час їх репродукції. Вони забезпечують регуляцію експресії вірусного геному, є попередниками вірусних білків, здатні пригнічувати клітинний біосинтез. Залежно від розташування у віріоні, білки поділяються на капсидні, суперкапсидні, матриксні, білки серцевини та асоційовані з нуклеїновою кислотою.
 Взаємодія вірусів і клітини Типи взаємодії. При продуктивній інфекції вірус функціонує в клітині автономно, а його репродукція відбувається незалежно від репродукції клітинного генетичного апарата. При цьому утворюється нове покоління інфекційних вірусів. Якщо цикл репродукції вірусів блокується на одній із стадій, а нові віруси не утворюються, такий тип взаємодії позначають як абортивний. Коли з клітиною взаємодіють онкогенні РНК та ДНК геномні віруси (СНІДу, лейкозу), нуклеїнова кислота інтегрується в клітинну хромосому та існує там у вигляді провірусу. В результаті може спричинятися зміна спадкових властивостей клітини. Такий тип взаємодії називають вірогенією. Стадії репродукції До ранніх належить адсорбція вірусів на поверхні клітини, проникнення (пенетрація) їх всередину кілтини та їх роздягання (депротеїнізація). Пізні стадії (стратегія вірусного генома) включають синтез вірусних нуклеїнових кислот, синтез білка, збирання віріонів та вихід вірусних часток із клітини.
Взаємодія вірусів і клітини Типи взаємодії. При продуктивній інфекції вірус функціонує в клітині автономно, а його репродукція відбувається незалежно від репродукції клітинного генетичного апарата. При цьому утворюється нове покоління інфекційних вірусів. Якщо цикл репродукції вірусів блокується на одній із стадій, а нові віруси не утворюються, такий тип взаємодії позначають як абортивний. Коли з клітиною взаємодіють онкогенні РНК та ДНК геномні віруси (СНІДу, лейкозу), нуклеїнова кислота інтегрується в клітинну хромосому та існує там у вигляді провірусу. В результаті може спричинятися зміна спадкових властивостей клітини. Такий тип взаємодії називають вірогенією. Стадії репродукції До ранніх належить адсорбція вірусів на поверхні клітини, проникнення (пенетрація) їх всередину кілтини та їх роздягання (депротеїнізація). Пізні стадії (стратегія вірусного генома) включають синтез вірусних нуклеїнових кислот, синтез білка, збирання віріонів та вихід вірусних часток із клітини.
 Прикріплення вірусів до поверхні клітини забезпечується двома механізмами: неспецифічним і специфічним. Неспецифічний визначається силами електростатичної взаємодії, на поверхні вірусів і клітин, які несуть різні заряди. Специфічний механізм (обернена та необернена адсорбція) зумовлюється комплементарними вірусними та клітинними рецепторами. Вони можуть мати білкову, вуглеводну, ліпідну природу. Наприклад, рецептором для вірусів грипу є сіалова кислота. Число рецепторів на ділянках адсорбції може сягати 3000. Проникнення вірусів всередину клітини відбувається за механізмом рецепторного ендоцитозу (варіант віропексису) на спеціальних ділянках клітинних мембран, які містять особливий білок з високою молекулярною масою клатрин. Мембрани інвагінуються, і утворюються вкриті клатрином внутрішньоклітинні вакуолі. Вакуолі, об’єднуючись, утворюють рецептосоми, а останні зливаються з лізосомами. Поверхневі білки вірусів взаємодіють із ферментами лізосом, а їх нуклеопротеїд виходить у цитоплазму. Однак існує ще один механізм проникнення вірусів у клітину індукція злиття мембран. Вона відбувається завдяки особливому вірусному білку злиття (F від fusion злиття).
Прикріплення вірусів до поверхні клітини забезпечується двома механізмами: неспецифічним і специфічним. Неспецифічний визначається силами електростатичної взаємодії, на поверхні вірусів і клітин, які несуть різні заряди. Специфічний механізм (обернена та необернена адсорбція) зумовлюється комплементарними вірусними та клітинними рецепторами. Вони можуть мати білкову, вуглеводну, ліпідну природу. Наприклад, рецептором для вірусів грипу є сіалова кислота. Число рецепторів на ділянках адсорбції може сягати 3000. Проникнення вірусів всередину клітини відбувається за механізмом рецепторного ендоцитозу (варіант віропексису) на спеціальних ділянках клітинних мембран, які містять особливий білок з високою молекулярною масою клатрин. Мембрани інвагінуються, і утворюються вкриті клатрином внутрішньоклітинні вакуолі. Вакуолі, об’єднуючись, утворюють рецептосоми, а останні зливаються з лізосомами. Поверхневі білки вірусів взаємодіють із ферментами лізосом, а їх нуклеопротеїд виходить у цитоплазму. Однак існує ще один механізм проникнення вірусів у клітину індукція злиття мембран. Вона відбувається завдяки особливому вірусному білку злиття (F від fusion злиття).
 Роздягання віріонів багатоступеневий процес, під час якого вивільнюється їх нуклеїновий апарат, зникають захисні оболонки, які гальмують експресію генома. Відбувається воно в спеціалізованих ділянках лізосомах, апараті Гольджі. Пізні стадії репродукції спрямовані на синтез вірусних нуклеїнових кислот та білка. Механізм реплікації (утворення вірусних геномів, які є точною копією попередника) залежить від особливостей нуклеїнової кислоти. У різних видів вірусів він неоднаковий. Виділяють 6 основних класів реплікації вірусів У двониткових ДНК містких вірусів (герепесвіруси, аденовіруси, віруси натуральної віспи) спочатку відбувається деспіралізація ДНК і розходження її ниток. На одній з них за принципом комплементарності синтезується нова нитка ДНК, на другій інформаційна (матрична) РНК (іРНК або м. РНК). Цей процес триває, поки в клітинах не утвориться достатня кількість нуклеїнових кислот. В однониткових ДНК геномних вірусів процес відбувається за умови утворення проміжної форми ДНК.
Роздягання віріонів багатоступеневий процес, під час якого вивільнюється їх нуклеїновий апарат, зникають захисні оболонки, які гальмують експресію генома. Відбувається воно в спеціалізованих ділянках лізосомах, апараті Гольджі. Пізні стадії репродукції спрямовані на синтез вірусних нуклеїнових кислот та білка. Механізм реплікації (утворення вірусних геномів, які є точною копією попередника) залежить від особливостей нуклеїнової кислоти. У різних видів вірусів він неоднаковий. Виділяють 6 основних класів реплікації вірусів У двониткових ДНК містких вірусів (герепесвіруси, аденовіруси, віруси натуральної віспи) спочатку відбувається деспіралізація ДНК і розходження її ниток. На одній з них за принципом комплементарності синтезується нова нитка ДНК, на другій інформаційна (матрична) РНК (іРНК або м. РНК). Цей процес триває, поки в клітинах не утвориться достатня кількість нуклеїнових кислот. В однониткових ДНК геномних вірусів процес відбувається за умови утворення проміжної форми ДНК.
 Репродукція вірусів у культурах клітин. Цей метод широко використовують для лабораторної діагностики вірусних інфекцій. У будь якій вірусологічній лабораторії є спеціальні лінії культур клітин: перещеплювані культури клітин He. La, HЕp 2, Vero, KB, первинно трипсинізовані культури ембріонів людини, нирок мавп, фібробластів ембріона курки, тощо. Первинно трипсинізовані культури клітин отримують із будь яких тканин тварини або людини шляхом їх дезинтеграції ферментативною обробкою. Для цього отримані тканини подрібнюють на невеликі шматочки і доливають до них 0, 25% розчин трипсину. Дезинтеграцію (руйнування зв’язків між клітинами) можна проводити при температурі 37 °С або 4 °С. Після цього суспензію центрифугують, отримані клітини поміщають у спеціальні середовища, а їх кількість визначають при підрахунку в камері Горяєва.
Репродукція вірусів у культурах клітин. Цей метод широко використовують для лабораторної діагностики вірусних інфекцій. У будь якій вірусологічній лабораторії є спеціальні лінії культур клітин: перещеплювані культури клітин He. La, HЕp 2, Vero, KB, первинно трипсинізовані культури ембріонів людини, нирок мавп, фібробластів ембріона курки, тощо. Первинно трипсинізовані культури клітин отримують із будь яких тканин тварини або людини шляхом їх дезинтеграції ферментативною обробкою. Для цього отримані тканини подрібнюють на невеликі шматочки і доливають до них 0, 25% розчин трипсину. Дезинтеграцію (руйнування зв’язків між клітинами) можна проводити при температурі 37 °С або 4 °С. Після цього суспензію центрифугують, отримані клітини поміщають у спеціальні середовища, а їх кількість визначають при підрахунку в камері Горяєва.
 Досліджуваний матеріал для вірусологічного методу: кров, змиви з носо і ротоглотки, харкотиння, рідина з пухирців, спинномозкова рідина, сеча, фекалії, шматочки органів і тканин людини, отриманих при дослі дженні трупів, матеріал від лабораторних тварин тощо. Одержаний матеріал зберігають при температурі 20 °С. Для транспортування вірусів можна використати термоси, які заповнюються сухим льодом. Дослідження на курячому ембріоні Існує два способи зараження курячих ембріонів – відкритий і закритий). Ембріони, у яких візуально немає пошкодження шкаралупи, просвічують за допомогою овоскопа і відмічають межу повітряного мішка. При відкритому способі шкаралупу над мішком обробляють спиртом і розчином йоду, зрізають шкаралупу, знімають верхній листок оболонки повітряного мішка і проводять зараження. Отвір закривають спеціальною скляною кришкою і герметизують стерильним розтопленим парафіном. При закритому способі зараження колючим інструментом роблять отвір у шкаралупі і вводять за допомогою шприца з товстою голкою 0, 1 0, 2 мл під контролем овоскопа матеріал з вірусом.
Досліджуваний матеріал для вірусологічного методу: кров, змиви з носо і ротоглотки, харкотиння, рідина з пухирців, спинномозкова рідина, сеча, фекалії, шматочки органів і тканин людини, отриманих при дослі дженні трупів, матеріал від лабораторних тварин тощо. Одержаний матеріал зберігають при температурі 20 °С. Для транспортування вірусів можна використати термоси, які заповнюються сухим льодом. Дослідження на курячому ембріоні Існує два способи зараження курячих ембріонів – відкритий і закритий). Ембріони, у яких візуально немає пошкодження шкаралупи, просвічують за допомогою овоскопа і відмічають межу повітряного мішка. При відкритому способі шкаралупу над мішком обробляють спиртом і розчином йоду, зрізають шкаралупу, знімають верхній листок оболонки повітряного мішка і проводять зараження. Отвір закривають спеціальною скляною кришкою і герметизують стерильним розтопленим парафіном. При закритому способі зараження колючим інструментом роблять отвір у шкаралупі і вводять за допомогою шприца з товстою голкою 0, 1 0, 2 мл під контролем овоскопа матеріал з вірусом.
 Зараження курячих ембріонів. Курячі ембріони 6 12 денного віку є зручною моделлю для виділення та подальшої ідентифікації вірусів. Після зараження їх інкубують у термостаті при температурі 36 38 °С протягом декількох днів.
Зараження курячих ембріонів. Курячі ембріони 6 12 денного віку є зручною моделлю для виділення та подальшої ідентифікації вірусів. Після зараження їх інкубують у термостаті при температурі 36 38 °С протягом декількох днів.
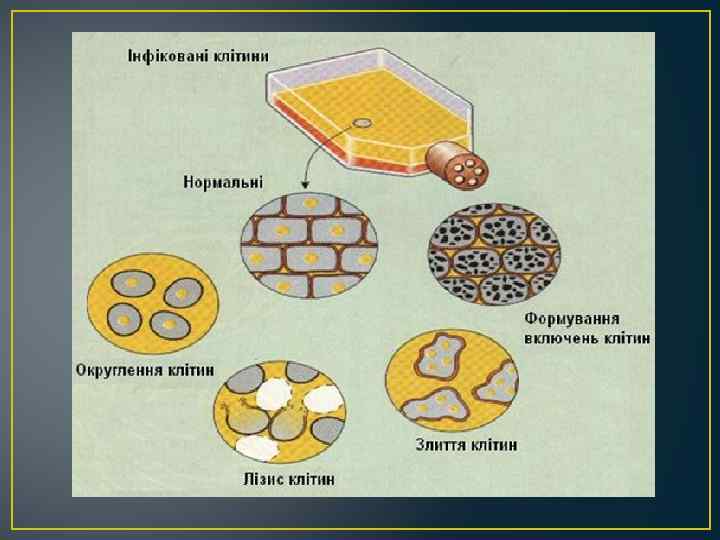
, або цитопатичною дією віруса (ЦПД). В") Деструкція клітин спричинена вірусом називається цитопатичним ефектом (ЦПЕ), або цитопатичною дією віруса (ЦПД). В зараженій клітині відбувається зміна метаболізму, що зумовлено синтезом вірусоспецифічних макромолекул, активацією або гальмуванням ферментних систем клітини, пошкодження її хромосомного апарату та органел. Вірус, який спричиняє цитопатогенним. цитопатичний ефект, називається Цитопатичні зміни інфікованих вірусом клітин • специфічні утворення вірусних включень і симпластів • Неспецифічні або мало специфічними мутне набухання клітини внаслідок порушення проникливості плазмолеми, маргинація хроматину, хромосомні аберації, пікноз ядер. Вакуолізація цитоплазми являється досить частою мало специфічною ознакою вірусної інфекції, проте в деяких випадках вона набуває настільки виражений і своєрідний характер, що вважається специфічною. Наприклад, одна із назв віруса SV 40 вакуолізуючий вірус.
Деструкція клітин спричинена вірусом називається цитопатичним ефектом (ЦПЕ), або цитопатичною дією віруса (ЦПД). В зараженій клітині відбувається зміна метаболізму, що зумовлено синтезом вірусоспецифічних макромолекул, активацією або гальмуванням ферментних систем клітини, пошкодження її хромосомного апарату та органел. Вірус, який спричиняє цитопатогенним. цитопатичний ефект, називається Цитопатичні зміни інфікованих вірусом клітин • специфічні утворення вірусних включень і симпластів • Неспецифічні або мало специфічними мутне набухання клітини внаслідок порушення проникливості плазмолеми, маргинація хроматину, хромосомні аберації, пікноз ядер. Вакуолізація цитоплазми являється досить частою мало специфічною ознакою вірусної інфекції, проте в деяких випадках вона набуває настільки виражений і своєрідний характер, що вважається специфічною. Наприклад, одна із назв віруса SV 40 вакуолізуючий вірус.
 Методи ідентифікації вірусів
Методи ідентифікації вірусів
 Реакція гемаглютинації – феномен склеювання еритроцитів під впливом вірусів. Позитивна реакції – осад у вигляді «парасольки» , негативна – у вигляді «гудзика» .
Реакція гемаглютинації – феномен склеювання еритроцитів під впливом вірусів. Позитивна реакції – осад у вигляді «парасольки» , негативна – у вигляді «гудзика» .
. Реакція базується на здатності блокувати гемаглютинуючі властивості вірусів за допомогою") Реакція гальмування гемаглютинації (РГГА). Реакція базується на здатності блокувати гемаглютинуючі властивості вірусів за допомогою специфічних антитіл. У результаті цього спостерігається затримка аглютинації еритроцитів.
Реакція гальмування гемаглютинації (РГГА). Реакція базується на здатності блокувати гемаглютинуючі властивості вірусів за допомогою специфічних антитіл. У результаті цього спостерігається затримка аглютинації еритроцитів.
 — віруси,") Бактеріофа ги або фа ги (від др. грец. φᾰγω — «пожираю» ) — віруси, які вибірково вражають бактеріальні клітини. Найбільше бактеріофаги розмножуються в бактеріях та викликають їх лізис. Структура фага Т 2. 1 головка, 2 відросток, 3 нуклеїнова кислота, 4 капсид, 5 комірець, 6 чохол, 7 хвостві нитки, 8 шипи, 9 базальна пластинка
Бактеріофа ги або фа ги (від др. грец. φᾰγω — «пожираю» ) — віруси, які вибірково вражають бактеріальні клітини. Найбільше бактеріофаги розмножуються в бактеріях та викликають їх лізис. Структура фага Т 2. 1 головка, 2 відросток, 3 нуклеїнова кислота, 4 капсид, 5 комірець, 6 чохол, 7 хвостві нитки, 8 шипи, 9 базальна пластинка
 Хімічний склад бактеріофагів, як інших вірусів, представлений нуклеїновою кислотою, білками, невеликою кількістю ліпідів у оболонці. Більшість фагів містить ДНК і окремі – РНК. За своїм складом фагові нуклеїнові кислоти не відрізняються від аналогічних структур інших мікроорганізмів і вірусів. Всередині голівки є невелика кількість “внутрішнього білка”. В дистальній частині відростка, під чохлом, міститься фермент лізоцим, який відіграє роль у проникненні фагової нуклеїнової кислоти в бактеріальну клітину. Бактеріофаги на поверхні клітини
Хімічний склад бактеріофагів, як інших вірусів, представлений нуклеїновою кислотою, білками, невеликою кількістю ліпідів у оболонці. Більшість фагів містить ДНК і окремі – РНК. За своїм складом фагові нуклеїнові кислоти не відрізняються від аналогічних структур інших мікроорганізмів і вірусів. Всередині голівки є невелика кількість “внутрішнього білка”. В дистальній частині відростка, під чохлом, міститься фермент лізоцим, який відіграє роль у проникненні фагової нуклеїнової кислоти в бактеріальну клітину. Бактеріофаги на поверхні клітини
 Залежно від наслідків взаємодії фагів з бактеріальною клітиною, вони поді ляються на вірулентні та помірні. Вірулентні бактеріофаги проникають всередину клітини, спричиняючи її лізис. Взаємодія їх з клітиною складається з ряду етапів, притаманних практично всім вірусам. Коли з клітиною взаємодіють помірні бактеріофаги, частина клітин залишається неушкодженою ними, тому що спостерігається явище лізогенії – інтеграції генома бактеріофага в геном клітини
Залежно від наслідків взаємодії фагів з бактеріальною клітиною, вони поді ляються на вірулентні та помірні. Вірулентні бактеріофаги проникають всередину клітини, спричиняючи її лізис. Взаємодія їх з клітиною складається з ряду етапів, притаманних практично всім вірусам. Коли з клітиною взаємодіють помірні бактеріофаги, частина клітин залишається неушкодженою ними, тому що спостерігається явище лізогенії – інтеграції генома бактеріофага в геном клітини
 Віроїди Назва «віроїд» була запропонована в 1971 р. Т. Дінером. Від вірусів віроїди відрізняються за такими ознаками: 1. Відсутня білкова оболонка, віроїди складаються тільки із інфекційної РНК. 2. Віроїди – найменші, здатні до розмноження частинки, які відомі у природі. Їх РНК становить тільки 300 400 нуклеотидів. 3. Віроїди – це однокільцеві РНК. 4. Молекули РНК віроїдів не кодують власних білків. З 1971 р. виявлено біля 10 видів віроїдів, які відрізняються за первинною структурою, хазяїнами, яких вони вражають, сиптомами хвороб. Вони вражають рослинні клітини.
Віроїди Назва «віроїд» була запропонована в 1971 р. Т. Дінером. Від вірусів віроїди відрізняються за такими ознаками: 1. Відсутня білкова оболонка, віроїди складаються тільки із інфекційної РНК. 2. Віроїди – найменші, здатні до розмноження частинки, які відомі у природі. Їх РНК становить тільки 300 400 нуклеотидів. 3. Віроїди – це однокільцеві РНК. 4. Молекули РНК віроїдів не кодують власних білків. З 1971 р. виявлено біля 10 видів віроїдів, які відрізняються за первинною структурою, хазяїнами, яких вони вражають, сиптомами хвороб. Вони вражають рослинні клітини.
 Пріони. Таку назву запропонував, С. Прузінер, який їх відкрив у 1982 р. Це низькомолекулярні білки, які не містять нуклеїнових кислот. Пріони складаються із особливого білка, який існує у вигляді двох ізомерів. Один з них – нормальний клітинний протеїн. Інший ізомер – аномальний. Від нормального він відрізняється вторинною структурою. Його вміст у хворих тварин в 10 раз більший, ніж у здорових. Відомо біля 12 пріонових хвороб, шість з яких спостерігають у тварин. Шість хвороб описані і в людей (Коров’ячий сказ або хвороба Крейтцфельда Якоба, куру, синдром Альперса). Це хвороби із сповільненою дією, які часто призводять до порушень діяльності ЦНС та смерті. Подібність до вірусу: малі розміри, нездатність розмножуватися на штучних поживних середовищах, специфічне коло хазяїнів. Відмінність від вірусу: у пріонів немає власного геному, отже, вони не можуть розглядатися, як живі істоти, вони не провокують імунної відповіді, набагато стійкіші за віруси до температури, дії УФ опромінення, іонізуючої радіації.
Пріони. Таку назву запропонував, С. Прузінер, який їх відкрив у 1982 р. Це низькомолекулярні білки, які не містять нуклеїнових кислот. Пріони складаються із особливого білка, який існує у вигляді двох ізомерів. Один з них – нормальний клітинний протеїн. Інший ізомер – аномальний. Від нормального він відрізняється вторинною структурою. Його вміст у хворих тварин в 10 раз більший, ніж у здорових. Відомо біля 12 пріонових хвороб, шість з яких спостерігають у тварин. Шість хвороб описані і в людей (Коров’ячий сказ або хвороба Крейтцфельда Якоба, куру, синдром Альперса). Це хвороби із сповільненою дією, які часто призводять до порушень діяльності ЦНС та смерті. Подібність до вірусу: малі розміри, нездатність розмножуватися на штучних поживних середовищах, специфічне коло хазяїнів. Відмінність від вірусу: у пріонів немає власного геному, отже, вони не можуть розглядатися, як живі істоти, вони не провокують імунної відповіді, набагато стійкіші за віруси до температури, дії УФ опромінення, іонізуючої радіації.
 Залежно від наявності чи відсутності зовнішніх проявів при ураженні організму вірусами розрізняють: гострі вірусні інфекції, що повністю проявляються клінічно, латентні вірусні інфекції з пері одичними загостреннями (чи без них), у проміжках між ними вірус не виявляється; хронічні вірусні інфекції з вірусом, що постій но виділяється, з більш менш вираженими ознаками захворювання організму; повільні вірусні інфекції, що характеризуються тривалим, іноді багаторічним інкубаційним періодом з наступним неухильним розвитком захворювання, що спричиняє загибель організму. Персистенція вірусів, є безсимптомними чи з маловираженими ознаками ушкодженнями, називають також вірусоносійством. Особливу епідеміологічну небезпеку становлять персистентні вірусні інфекції, при яких у навколишнє середовище виділяється вірус.
Залежно від наявності чи відсутності зовнішніх проявів при ураженні організму вірусами розрізняють: гострі вірусні інфекції, що повністю проявляються клінічно, латентні вірусні інфекції з пері одичними загостреннями (чи без них), у проміжках між ними вірус не виявляється; хронічні вірусні інфекції з вірусом, що постій но виділяється, з більш менш вираженими ознаками захворювання організму; повільні вірусні інфекції, що характеризуються тривалим, іноді багаторічним інкубаційним періодом з наступним неухильним розвитком захворювання, що спричиняє загибель організму. Персистенція вірусів, є безсимптомними чи з маловираженими ознаками ушкодженнями, називають також вірусоносійством. Особливу епідеміологічну небезпеку становлять персистентні вірусні інфекції, при яких у навколишнє середовище виділяється вірус.
 Вірусний канцерогенез Спочатку деякі віруси розглядалися в якості інфекційних агентів, що спонукають клітини до нерегульованого розмноженню. На противагу цьому Л. А. Зільбер (1945) розробив теорію, згідно з якою геном вірусу інтегрується в геном нормальної клітини, перетворюючи її в пухлинну, тобто канцерогенні віруси по своїй дії принципово відрізняються від інфекційних. У 70 ті роки в канцерогенних РНК вмісних вірусах виявлені гени, необхідні для перетворення нормальної клітини в пухлинну, – трансформують гени або онкогени (v onc – вірусні онкогени). У подальшому копії або аналогії онкогенів були виявлені в нормальних клітинах різних тварин н людини (з опс – «cellular» клітинні онкогени), потім була доведена здатність онкогена вбудовуватися в геном вірусу. Онкогени нині ідентифіковані, визначена їх хімічна структура, локалізація в хромосомах. Ідентифіковано також білки – продукти діяльності цих генів, кожен з них синтезує свій специфічний білок.
Вірусний канцерогенез Спочатку деякі віруси розглядалися в якості інфекційних агентів, що спонукають клітини до нерегульованого розмноженню. На противагу цьому Л. А. Зільбер (1945) розробив теорію, згідно з якою геном вірусу інтегрується в геном нормальної клітини, перетворюючи її в пухлинну, тобто канцерогенні віруси по своїй дії принципово відрізняються від інфекційних. У 70 ті роки в канцерогенних РНК вмісних вірусах виявлені гени, необхідні для перетворення нормальної клітини в пухлинну, – трансформують гени або онкогени (v onc – вірусні онкогени). У подальшому копії або аналогії онкогенів були виявлені в нормальних клітинах різних тварин н людини (з опс – «cellular» клітинні онкогени), потім була доведена здатність онкогена вбудовуватися в геном вірусу. Онкогени нині ідентифіковані, визначена їх хімічна структура, локалізація в хромосомах. Ідентифіковано також білки – продукти діяльності цих генів, кожен з них синтезує свій специфічний білок.
 Противірусний набутий імунітет специфічний і в спадщину не передається. Розрізняють активно і пасивно набутий імунітет. Перший з них розвивається в результаті природного (клінічно вираженого чи безсимптомного) інфікування чи після вакцинації живими чи убитими вакцинами. Пасивно набутий імунітет може бути створений штучно шляхом введення в організм імунних сироваток, гамма глобулінів, імунолактону, а також при введенні ефекторних клітин, одержаних від сингенного імунного донора, чи передається від матері плоду через плаценту і при вживанні немовлям молозива в перші дні життя.
Противірусний набутий імунітет специфічний і в спадщину не передається. Розрізняють активно і пасивно набутий імунітет. Перший з них розвивається в результаті природного (клінічно вираженого чи безсимптомного) інфікування чи після вакцинації живими чи убитими вакцинами. Пасивно набутий імунітет може бути створений штучно шляхом введення в організм імунних сироваток, гамма глобулінів, імунолактону, а також при введенні ефекторних клітин, одержаних від сингенного імунного донора, чи передається від матері плоду через плаценту і при вживанні немовлям молозива в перші дні життя.
 Клітинний противірусний імунітет Т клітини мають на поверхні антигенспецифічні рецептори. При стимуляції антигеном Т клітини здійснюють три різні функції: 1) специфічно убивають чужорідні клітини чи власні клітини організму, інфіковані вірусами; 2) допомагають специфічним Т чи В лімфоцитам відповідати на антиген і можуть активізувати деякі інші клітини (не лімфоцити), наприклад макрофаги; 3) пригнічують (супресія) відповіді специфічних Т чи В лімфоцитів. Ці різні функції здійснюються різними субпопуляціями Т клітин, цитотоксичними Т клітинами(кілерами), Т клітинами хелперами (чи індукторами) і Т клітинами супресорами відповідно. Цитотоксичні Т лімфоцити убивають клітини, інфіковані вірусами. Т хелпери активізують макрофаги шляхом секреції лімфокінів (чи інтерлейкінів — посередників між лейкоцитами). Одним з найбільш вивчених лімфокінів є фактор, який інгібує міграцію макрофагів (MIF — macrophage migration inhibiting factor). У результаті останні починають більш активно фагоцитувати клітини уражені вірусами.
Клітинний противірусний імунітет Т клітини мають на поверхні антигенспецифічні рецептори. При стимуляції антигеном Т клітини здійснюють три різні функції: 1) специфічно убивають чужорідні клітини чи власні клітини організму, інфіковані вірусами; 2) допомагають специфічним Т чи В лімфоцитам відповідати на антиген і можуть активізувати деякі інші клітини (не лімфоцити), наприклад макрофаги; 3) пригнічують (супресія) відповіді специфічних Т чи В лімфоцитів. Ці різні функції здійснюються різними субпопуляціями Т клітин, цитотоксичними Т клітинами(кілерами), Т клітинами хелперами (чи індукторами) і Т клітинами супресорами відповідно. Цитотоксичні Т лімфоцити убивають клітини, інфіковані вірусами. Т хелпери активізують макрофаги шляхом секреції лімфокінів (чи інтерлейкінів — посередників між лейкоцитами). Одним з найбільш вивчених лімфокінів є фактор, який інгібує міграцію макрофагів (MIF — macrophage migration inhibiting factor). У результаті останні починають більш активно фагоцитувати клітини уражені вірусами.
 Механізми противірусного імунітету. За Ройтом А. , 2000.
Механізми противірусного імунітету. За Ройтом А. , 2000.