

spatial lecture2015-2-4v.ppt
- Количество слайдов: 169
. Клетки места. Лекция 2 Плескачева М. Г. 2015") История изучения пространственного поведения (продолжение). Клетки места. Лекция 2 Плескачева М. Г. 2015
История изучения пространственного поведения (продолжение). Клетки места. Лекция 2 Плескачева М. Г. 2015
 Edward C. Tolman. Когнитивные карты Идея «когнитивной карты» . “We believe that in the course of learning something like a field map of the environment gets established in the rat’s brain” “Cognitive maps in rats and men”, Psychol Rev. 1948; 55: 189 -208 1886 -1959 University of California in Berkeley Критик классического бихевиоризма (Уотсона), упрощенного представления о поведении как о совокупности элементарных ответов на стимулы. Purposive Behavior in Animals and Men, 1932 Studies in spatial learning: Orientation and the short-cut. J. Exp Psychol. 1946; 36: 13 -24 Studies in spatial learning. II. Place learning versus response learning. J. of Exp Psych. 1946, 36: 221 -229.
Edward C. Tolman. Когнитивные карты Идея «когнитивной карты» . “We believe that in the course of learning something like a field map of the environment gets established in the rat’s brain” “Cognitive maps in rats and men”, Psychol Rev. 1948; 55: 189 -208 1886 -1959 University of California in Berkeley Критик классического бихевиоризма (Уотсона), упрощенного представления о поведении как о совокупности элементарных ответов на стимулы. Purposive Behavior in Animals and Men, 1932 Studies in spatial learning: Orientation and the short-cut. J. Exp Psychol. 1946; 36: 13 -24 Studies in spatial learning. II. Place learning versus response learning. J. of Exp Psych. 1946, 36: 221 -229.
 Edward C. Tolman. Когнитивные карты «В процессе обучения в мозге крысы образуется нечто подобное карте окружающей среды. …Поступающие стимулы перерабатываются в центре управления в предполагаемую когнитивную карту окружающей среды. . . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным. » 1886 -1959 University of California in Berkeley Критик классического бихевиоризма, упрощенного представления о поведении как о совокупности элементарных ответов на стимулы по типу стимул-реакция. Идея «когнитивной карты» “Cognitive maps in rats and men”, Psychol Rev. 1948 + +
Edward C. Tolman. Когнитивные карты «В процессе обучения в мозге крысы образуется нечто подобное карте окружающей среды. …Поступающие стимулы перерабатываются в центре управления в предполагаемую когнитивную карту окружающей среды. . . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным. » 1886 -1959 University of California in Berkeley Критик классического бихевиоризма, упрощенного представления о поведении как о совокупности элементарных ответов на стимулы по типу стимул-реакция. Идея «когнитивной карты» “Cognitive maps in rats and men”, Psychol Rev. 1948 + +
 1. На основе исследования и запоминания элементов среды формируется ментальное представление, «карта» этой среды 2. Представление – интегрированная информация из разных источников 3. В мозге существует «центр управления» , где обрабатывается информация 4. Навигация в реальной среде осуществляется с использованием ментального представления 5. Такая навигация обеспечивает достижение цели с разных стартовых позиций и не нарушается при частичном изменении среды
1. На основе исследования и запоминания элементов среды формируется ментальное представление, «карта» этой среды 2. Представление – интегрированная информация из разных источников 3. В мозге существует «центр управления» , где обрабатывается информация 4. Навигация в реальной среде осуществляется с использованием ментального представления 5. Такая навигация обеспечивает достижение цели с разных стартовых позиций и не нарушается при частичном изменении среды
 Изучение механизмов пространственного поведения в России Обучался в С. -Петербургском университете (естественное отд. физ. -мат. фак-та). Студентом начал экспериментальную работу в физиологической лаборатории Николая Евгеньевича Введенского по проблеме спинно -мозговой координации рефлекторных движений. Оставлен Н. Е. Введенским при кафедре, стажировался в Голландии (Утрехт, у Р. Магнуса) – для изучения методики опытов на теплокровных животных. 1885 -1974 Бериташвили Иван Соломонович. 1915 -1919 — приват-доцент Новороссийского университета. 1919 — профессор Тбилисского университета С 1935 — директор института физиологии при Тбилисском университете 1959
Изучение механизмов пространственного поведения в России Обучался в С. -Петербургском университете (естественное отд. физ. -мат. фак-та). Студентом начал экспериментальную работу в физиологической лаборатории Николая Евгеньевича Введенского по проблеме спинно -мозговой координации рефлекторных движений. Оставлен Н. Е. Введенским при кафедре, стажировался в Голландии (Утрехт, у Р. Магнуса) – для изучения методики опытов на теплокровных животных. 1885 -1974 Бериташвили Иван Соломонович. 1915 -1919 — приват-доцент Новороссийского университета. 1919 — профессор Тбилисского университета С 1935 — директор института физиологии при Тбилисском университете 1959
 И. С. Бериташвили. Исследование пространственной ориентации животных «Более 25 лет прошло с тех пор, как мы начали изучать высшую форму поведения животных, которая основывается на индивидуальном опыте и характеризуется тем, что направляется воспроизведенным образом или конкретным представлением жизненно-важных объектов, проецируемых в определенной внешней среде. » Предисловие к книге (1959 г. ), стр. 1 Первые сообщения об этой форме поведения сделаны в 1932 -1933 г. У животных «под влиянием того или другого компонента знакомой среды возникают в мозгу приблизительно такие же образы с такой же локализацией их во внешней среде, как это было впервые при его жизненной деятельности в данной среде. Вследствие этого животные ориентируются в пространстве на основании этих конкретных представлений, как при непосредственном восприятии соответствующей среды. » стр. 23
И. С. Бериташвили. Исследование пространственной ориентации животных «Более 25 лет прошло с тех пор, как мы начали изучать высшую форму поведения животных, которая основывается на индивидуальном опыте и характеризуется тем, что направляется воспроизведенным образом или конкретным представлением жизненно-важных объектов, проецируемых в определенной внешней среде. » Предисловие к книге (1959 г. ), стр. 1 Первые сообщения об этой форме поведения сделаны в 1932 -1933 г. У животных «под влиянием того или другого компонента знакомой среды возникают в мозгу приблизительно такие же образы с такой же локализацией их во внешней среде, как это было впервые при его жизненной деятельности в данной среде. Вследствие этого животные ориентируются в пространстве на основании этих конкретных представлений, как при непосредственном восприятии соответствующей среды. » стр. 23
 И. С. Бериташвили. Исследование пространственной ориентации животных О наличии у животных «конкретного представления» И. С. Бериташвили судил по поведенческим показателям, полученным в ходе хорошо продуманных экспериментов. Идея о существовании «психонервного процесса образа или конкретного представления жизненно-важного объекта» «… «идеационная деятельность» [ по Хеббу] мозга (т. е. та деятельность, которая выявляется в представлениях) играет большую роль в поведении высших позвоночных животных» стр. 8 Психонервный комплекс представления среды включает зрительные, звуковые, обонятельные, вкусовые и кинестетические «восприятия» , а также информацию от «внутренних» рецепторов. Важная роль коры головного мозга в этих процессах Экспериментально доказал важную роль вестибулярной информации в процессах ориентации животных
И. С. Бериташвили. Исследование пространственной ориентации животных О наличии у животных «конкретного представления» И. С. Бериташвили судил по поведенческим показателям, полученным в ходе хорошо продуманных экспериментов. Идея о существовании «психонервного процесса образа или конкретного представления жизненно-важного объекта» «… «идеационная деятельность» [ по Хеббу] мозга (т. е. та деятельность, которая выявляется в представлениях) играет большую роль в поведении высших позвоночных животных» стр. 8 Психонервный комплекс представления среды включает зрительные, звуковые, обонятельные, вкусовые и кинестетические «восприятия» , а также информацию от «внутренних» рецепторов. Важная роль коры головного мозга в этих процессах Экспериментально доказал важную роль вестибулярной информации в процессах ориентации животных
 Схема экспериментальной комнаты для работы с кошками и собаками
Схема экспериментальной комнаты для работы с кошками и собаками
 Исследование локализации функций у животных и человека. Ввел представления об ансамблях нейронов Donald Olding Hebb (1904 -1985) “…the assembly is a system that is organized in the first place by a particular sensory event but is capable of continuing its activity after that stimulation has ceased. …. the system may be excited by other similar systems in the total absence of the sensory event that originally organized it. It thus meets the requirements of an ideational process. The very essence of an “idea” is that a brain activity is occurring in the absence of the environmental event it corresponds to. ” 1972 PHD (рук. К. Lashley, Chicago, Разработка лабиринта Хебба-Уильямса "Hebb. Harvard) Williams" maze (Hebb & Williams, 1946) Postdoc (рук. W. Penfield, Montreal Neurological Institute) Studies of the organization of behavior. I Behavior of the rat in a field orientation. 1938 Changes in the field orientation of the rat after cortical destruction. 1938 The Organization of Behavior (1949)
Исследование локализации функций у животных и человека. Ввел представления об ансамблях нейронов Donald Olding Hebb (1904 -1985) “…the assembly is a system that is organized in the first place by a particular sensory event but is capable of continuing its activity after that stimulation has ceased. …. the system may be excited by other similar systems in the total absence of the sensory event that originally organized it. It thus meets the requirements of an ideational process. The very essence of an “idea” is that a brain activity is occurring in the absence of the environmental event it corresponds to. ” 1972 PHD (рук. К. Lashley, Chicago, Разработка лабиринта Хебба-Уильямса "Hebb. Harvard) Williams" maze (Hebb & Williams, 1946) Postdoc (рук. W. Penfield, Montreal Neurological Institute) Studies of the organization of behavior. I Behavior of the rat in a field orientation. 1938 Changes in the field orientation of the rat after cortical destruction. 1938 The Organization of Behavior (1949)
 Лабиринт Хебба-Уильямса Обучение Тест, оценка ошибочных заходов в зоны, которые не ведут к корму Hebb, D. O. , K. A. Williams. 1946. A method of rating animal intelligence. J. Gen. Psychol. 34: 59 -65.
Лабиринт Хебба-Уильямса Обучение Тест, оценка ошибочных заходов в зоны, которые не ведут к корму Hebb, D. O. , K. A. Williams. 1946. A method of rating animal intelligence. J. Gen. Psychol. 34: 59 -65.
 Нейробиологическая основа когнитивной карты
Нейробиологическая основа когнитивной карты
 Гиппокамп как когнитивная карта
Гиппокамп как когнитивная карта
 обнаружены нейроны,") Гиппокамп John O’Keefe В 1971 году в гиппокампе крысы (поле CA 1) обнаружены нейроны, которые начинали активно генерировать спайки, только тогда, когда крыса пересекала определенное место экспериментального пространства. Когда крыса покидала эту область, активность нейронов снижалась и прекращалась. Клетки, обладающие такой функцией были названы «клетками места, place cells»
Гиппокамп John O’Keefe В 1971 году в гиппокампе крысы (поле CA 1) обнаружены нейроны, которые начинали активно генерировать спайки, только тогда, когда крыса пересекала определенное место экспериментального пространства. Когда крыса покидала эту область, активность нейронов снижалась и прекращалась. Клетки, обладающие такой функцией были названы «клетками места, place cells»
. The hippocampus as") Гиппокамп как когнитивная карта O'KEEFE, J. , and DOSTROVSKY, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34, 171 -5. O'KEEFE, J. (1976). Place units in the hippocampus of the freely moving rat. Exp. Neurol. 51, 78 -109. Открытие клеток места и анализ данных по удалению гиппокампа привели к предположению, что гиппокамп – структура, нейронный субстрат для создания ментального представления (когнитивной карты) среды. Исследовательская активность животного – необходимый компонент создания и контроля когнитивной карты.
Гиппокамп как когнитивная карта O'KEEFE, J. , and DOSTROVSKY, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34, 171 -5. O'KEEFE, J. (1976). Place units in the hippocampus of the freely moving rat. Exp. Neurol. 51, 78 -109. Открытие клеток места и анализ данных по удалению гиппокампа привели к предположению, что гиппокамп – структура, нейронный субстрат для создания ментального представления (когнитивной карты) среды. Исследовательская активность животного – необходимый компонент создания и контроля когнитивной карты.
 O’Keefe, Nadel, постулаты о пространстве «We shall postulate that, in most of the brain, space is represented in the relative manner; that is, referenced to the organism and built up through experience. We shall refer to these egocentric spatial systems as taxon systems, and this type of space will be called taxon space. Included within this category are all the sensory and motor systems. In Kantian, or locale, space, representations are located within a Euclidean system, yielding a space which does not depend for its existence on particular objects but which serves as a framework for relating these objects to each other independent of the observer. » P. 60 Постулировали наличие двух принципиально различных пространственных систем: 1. Пространство таксона (эгоцентрическая система). Пространство представлено через отношение к организму, привязано к нему и выстраивается через опыт. Включает все сенсорные и моторные системы 2. Локальное пространство (среда), представления помещены внутри Эвклидовой системы. Такое пространство служит основой (системой координат, framework), независимой от наблюдателя, для связи объектов друг с другом. Не является эгоцентрическим.
O’Keefe, Nadel, постулаты о пространстве «We shall postulate that, in most of the brain, space is represented in the relative manner; that is, referenced to the organism and built up through experience. We shall refer to these egocentric spatial systems as taxon systems, and this type of space will be called taxon space. Included within this category are all the sensory and motor systems. In Kantian, or locale, space, representations are located within a Euclidean system, yielding a space which does not depend for its existence on particular objects but which serves as a framework for relating these objects to each other independent of the observer. » P. 60 Постулировали наличие двух принципиально различных пространственных систем: 1. Пространство таксона (эгоцентрическая система). Пространство представлено через отношение к организму, привязано к нему и выстраивается через опыт. Включает все сенсорные и моторные системы 2. Локальное пространство (среда), представления помещены внутри Эвклидовой системы. Такое пространство служит основой (системой координат, framework), независимой от наблюдателя, для связи объектов друг с другом. Не является эгоцентрическим.
 Определение понятия «карты» по O’Keefe, Nadel, 1978 The simplest definition of a map is that it is the representation (usually two dimensional) of a part of space. The constituents of space are places, and thus an alternative definition of a map is the representation of a set of connected places which are systematically related to each other by a group of spatial transformation rules. Простейшее определение карты: это представление (обычно двухмерное) части пространства. Места – это компоненты пространства и, следовательно, альтернативное определение карты - это представление набора связанных мест
Определение понятия «карты» по O’Keefe, Nadel, 1978 The simplest definition of a map is that it is the representation (usually two dimensional) of a part of space. The constituents of space are places, and thus an alternative definition of a map is the representation of a set of connected places which are systematically related to each other by a group of spatial transformation rules. Простейшее определение карты: это представление (обычно двухмерное) части пространства. Места – это компоненты пространства и, следовательно, альтернативное определение карты - это представление набора связанных мест
 Когнитивные карты. Свойства. Предположили, что способность объективно представлять мир в трехмерном пространстве, т. е. строить когнитивные карты, должна быть врожденной. Система когнитивного картирования содержит карту для каждой среды, которая известна организму. Когнитивные карты могут быть мультимодальны. Свойство безопасности или опасности должно быть атрибутом среды или места внутри среды.
Когнитивные карты. Свойства. Предположили, что способность объективно представлять мир в трехмерном пространстве, т. е. строить когнитивные карты, должна быть врожденной. Система когнитивного картирования содержит карту для каждой среды, которая известна организму. Когнитивные карты могут быть мультимодальны. Свойство безопасности или опасности должно быть атрибутом среды или места внутри среды.
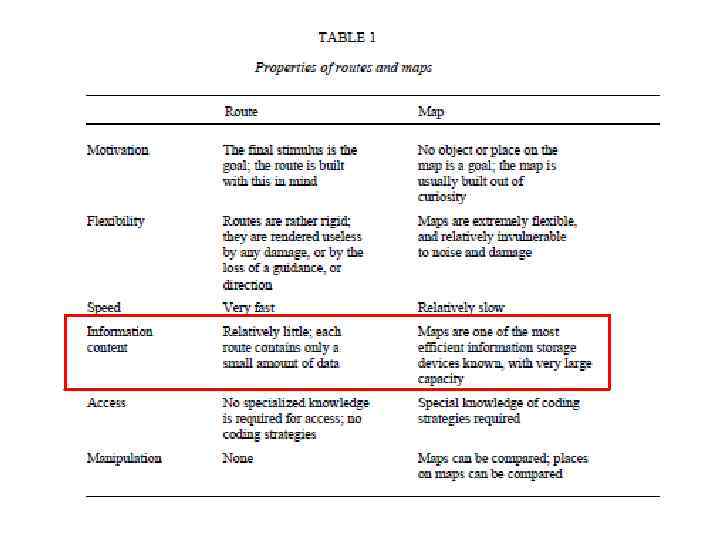
 принципиально") « Карты» и «маршруты» , разные механизмы передвижения в пространстве «Карты» (maps) принципиально отличаются по механизмам от «маршрутов» (routes). В последнем случае поведение основано на системах таксонов (taxons), специфические ответы направляются ориентирами. “The term route, with its implication of landmarks and specific responses guided by these, serves as a useful metaphor in describing taxon behaviours”. O’Keefe, Nadel, 1978
« Карты» и «маршруты» , разные механизмы передвижения в пространстве «Карты» (maps) принципиально отличаются по механизмам от «маршрутов» (routes). В последнем случае поведение основано на системах таксонов (taxons), специфические ответы направляются ориентирами. “The term route, with its implication of landmarks and specific responses guided by these, serves as a useful metaphor in describing taxon behaviours”. O’Keefe, Nadel, 1978
 и «карт» (maps), O’Keefe, Nadel, 1978 Маршруты • привязанность к") Некоторые характеристики «маршрутов» (routes) и «карт» (maps), O’Keefe, Nadel, 1978 Маршруты • привязанность к специфическому сигналу или элементу среды. Моторная система работает на приближение или на удаление от этого сигнала. • Скорее ригидны и становятся бесполезны при потере сигнала или направления • Скорость быстрая Карты (представления части пространства) • нет целевых объектов, строится на любознательности • гибкие и относительно устойчивые к «шумам и повреждениям» • Скорость скорее медленная • Именно карты контролируются гиппокампом
Некоторые характеристики «маршрутов» (routes) и «карт» (maps), O’Keefe, Nadel, 1978 Маршруты • привязанность к специфическому сигналу или элементу среды. Моторная система работает на приближение или на удаление от этого сигнала. • Скорее ригидны и становятся бесполезны при потере сигнала или направления • Скорость быстрая Карты (представления части пространства) • нет целевых объектов, строится на любознательности • гибкие и относительно устойчивые к «шумам и повреждениям» • Скорость скорее медленная • Именно карты контролируются гиппокампом
 According to theory, exploration is behaviour designed initially to build") Роль исследовательской активности (exploration) According to theory, exploration is behaviour designed initially to build and subsequently to update cognitive maps. • Исследовательская активность – поведение предназначенное вначале строить, а впоследствии обновлять когнитивные карты • При несовпадении текущего и предсказанного представлением сенсорного входа переключается набор misplace детекторов, выход из которых активирует и направляет моторные системы, вовлеченные в исследовательское поведение. Новая информация о месте включается в карту. • В новой среде все misplace детекторы активированы и исследовательская активность будет продолжаться до тех пор, пока вся информация не будет включена в карту данной среды.
Роль исследовательской активности (exploration) According to theory, exploration is behaviour designed initially to build and subsequently to update cognitive maps. • Исследовательская активность – поведение предназначенное вначале строить, а впоследствии обновлять когнитивные карты • При несовпадении текущего и предсказанного представлением сенсорного входа переключается набор misplace детекторов, выход из которых активирует и направляет моторные системы, вовлеченные в исследовательское поведение. Новая информация о месте включается в карту. • В новой среде все misplace детекторы активированы и исследовательская активность будет продолжаться до тех пор, пока вся информация не будет включена в карту данной среды.
, которая генерирует") Гиппокамп и карта Гиппокамп действует как когнитивная картирующая система (называемая local system), которая генерирует представление о месте и исследовательскую активность. Потеря этой системы ведет к использованию животным оставшейся экстра-гиппокампальной системы
Гиппокамп и карта Гиппокамп действует как когнитивная картирующая система (называемая local system), которая генерирует представление о месте и исследовательскую активность. Потеря этой системы ведет к использованию животным оставшейся экстра-гиппокампальной системы
 обнаружены нейроны,") Гиппокамп John O’Keefe В 1971 году в гиппокампе крысы (поле CA 1) обнаружены нейроны, которые начинали активно генерировать спайки, только тогда, когда крыса пересекала определенное место экспериментального пространства. Когда крыса покидала эту область, активность нейронов снижалась и прекращалась. Клетки, обладающие такой функцией были названы «клетками места, place cells»
Гиппокамп John O’Keefe В 1971 году в гиппокампе крысы (поле CA 1) обнаружены нейроны, которые начинали активно генерировать спайки, только тогда, когда крыса пересекала определенное место экспериментального пространства. Когда крыса покидала эту область, активность нейронов снижалась и прекращалась. Клетки, обладающие такой функцией были названы «клетками места, place cells»
 гиппокампа.") Клетки места, основные характеристики Впервые обнаружены в поле CA 1 дорзального (ростральная область) гиппокампа. Позже найдены в поле CA 3, в зубчатой фасции, в том числе и в более каудальных отделах гиппокампа. Не все клетки гиппокампа являются клетками места. Клетки места – пирамидные клетки с низкой частотой разряда и с характерным типом активности Другие клетки – интернейроны, другой характер разряда, высокая частота разряда, theta cells
Клетки места, основные характеристики Впервые обнаружены в поле CA 1 дорзального (ростральная область) гиппокампа. Позже найдены в поле CA 3, в зубчатой фасции, в том числе и в более каудальных отделах гиппокампа. Не все клетки гиппокампа являются клетками места. Клетки места – пирамидные клетки с низкой частотой разряда и с характерным типом активности Другие клетки – интернейроны, другой характер разряда, высокая частота разряда, theta cells
 Срез мозга крысы, ростральная область гиппокампа CA 1 CA 3 Зубчатая фасция, DG
Срез мозга крысы, ростральная область гиппокампа CA 1 CA 3 Зубчатая фасция, DG
 Основные связи в гиппокампальной формации Медиальн. Латеральн. септум Зубчатая фасция Энторинальная кора
Основные связи в гиппокампальной формации Медиальн. Латеральн. септум Зубчатая фасция Энторинальная кора
 Клетки места, основные характеристики Разряды клетки места “complex spike burst” – вспышка, серия спайков с короткими интервалами- характерна для многих клеток места Цвет элементов изображения арены отражает частоту разряда клетки места Muller, Kubie, Ranck (1987)
Клетки места, основные характеристики Разряды клетки места “complex spike burst” – вспышка, серия спайков с короткими интервалами- характерна для многих клеток места Цвет элементов изображения арены отражает частоту разряда клетки места Muller, Kubie, Ranck (1987)
 Клетки места гиппокампа Поле разряда клетки места, область, где эта клетка активна Поля разрядов 4 клеток места
Клетки места гиппокампа Поле разряда клетки места, область, где эта клетка активна Поля разрядов 4 клеток места
 Ансамбли клеток поля CA 1 гиппокампа, картирование Поля разрядов ансамбля клеток места (одновременная регистрация 80 клеток). Не все клетки места активны в экспериментальной арене, активность только 30 -40% нейронов Стабильное положение поля разряда устанавливалось приблизительно через 10 мин Клетки места интернейроны
Ансамбли клеток поля CA 1 гиппокампа, картирование Поля разрядов ансамбля клеток места (одновременная регистрация 80 клеток). Не все клетки места активны в экспериментальной арене, активность только 30 -40% нейронов Стабильное положение поля разряда устанавливалось приблизительно через 10 мин Клетки места интернейроны
 Свойства клеток места 1. Пространственная специфичность 2. В условиях стабильности экспериментальной среды положение поля разряда клетки стабильно и воспроизводится при повторном помещении крысы в арену. 3. Активность клеток места на связана с какой-либо формой поведения, запускается только «по месту» Из Eric Hargreaves, 2007 I: Place Cells in the Hippocampus. htm
Свойства клеток места 1. Пространственная специфичность 2. В условиях стабильности экспериментальной среды положение поля разряда клетки стабильно и воспроизводится при повторном помещении крысы в арену. 3. Активность клеток места на связана с какой-либо формой поведения, запускается только «по месту» Из Eric Hargreaves, 2007 I: Place Cells in the Hippocampus. htm
 Новизна среды и клетки места • Новая среда вызывает исследовательскую активность животных • Процесс сопровождается «картированием» (mapping) среды: формированием стабильных полей разряда клеток места (по местоположению, площади и интенсивности генерации спайков). «Карта» формируется при первом попадании животного на данную территорию. • При неизменности среды поля этих клеток воспроизводятся на прежнем месте. Установленная «карта» стабильна если нет изменений окружающей среды.
Новизна среды и клетки места • Новая среда вызывает исследовательскую активность животных • Процесс сопровождается «картированием» (mapping) среды: формированием стабильных полей разряда клеток места (по местоположению, площади и интенсивности генерации спайков). «Карта» формируется при первом попадании животного на данную территорию. • При неизменности среды поля этих клеток воспроизводятся на прежнем месте. Установленная «карта» стабильна если нет изменений окружающей среды.
 Изменение среды и клетки места • Изменение знакомой среды запускает исследовательскую активность и процессы «перекартирования» (remapping ) ансамбля клеток места: активируются клетки места, ранее неактивные, изменяется положение полей и частота разряда уже работавших клеток
Изменение среды и клетки места • Изменение знакомой среды запускает исследовательскую активность и процессы «перекартирования» (remapping ) ансамбля клеток места: активируются клетки места, ранее неактивные, изменяется положение полей и частота разряда уже работавших клеток
 Трансформация полей клеток места при изменении среды Исходная арена Измененная среда Деформация формы поля разряда Изменение частоты разряда Глобальное перекартирование Derdikman, Moser, 2010
Трансформация полей клеток места при изменении среды Исходная арена Измененная среда Деформация формы поля разряда Изменение частоты разряда Глобальное перекартирование Derdikman, Moser, 2010
 1. Внешние стимулы Дистантные зрительные ориентиры") Информация, определяющая где будет разряжаться клетка места (PC) 1. Внешние стимулы Дистантные зрительные ориентиры – один из основных источников информации, поддерживающий активность PC у крыс. Запахи, звуки, проксимальные (внутри аппарата, арены, лабиринта) зрительные стимулы Взаимодействие стимулов разной модальности.
Информация, определяющая где будет разряжаться клетка места (PC) 1. Внешние стимулы Дистантные зрительные ориентиры – один из основных источников информации, поддерживающий активность PC у крыс. Запахи, звуки, проксимальные (внутри аппарата, арены, лабиринта) зрительные стимулы Взаимодействие стимулов разной модальности.
") Дистантные зрительные ориентиры и клетки места крыс Экспериментальные установки (лабиринты разного типа, Т-образный, радиальный) окружали занавесом, что позволяло контролировать зрительные и другие дистантные ориентиры в комнате (например, лампа, белая карточка, и др. , O’Keefe, Conway, 1978). Одновременное вращение вокруг лабиринта всех предметов вызывало соответствующий поворот местоположения клеток места.
Дистантные зрительные ориентиры и клетки места крыс Экспериментальные установки (лабиринты разного типа, Т-образный, радиальный) окружали занавесом, что позволяло контролировать зрительные и другие дистантные ориентиры в комнате (например, лампа, белая карточка, и др. , O’Keefe, Conway, 1978). Одновременное вращение вокруг лабиринта всех предметов вызывало соответствующий поворот местоположения клеток места.
 Зрительные стимулы внутри арены и клетки места Местоположение поля связано со зрительным сигналом. Изменение положения карточки изменяет положение поля PC Удаление карточки не всегда изменяет положение поля PC Местоположение поля определяется не только по зрительному ориентиру Muller, Kubie, 1987
Зрительные стимулы внутри арены и клетки места Местоположение поля связано со зрительным сигналом. Изменение положения карточки изменяет положение поля PC Удаление карточки не всегда изменяет положение поля PC Местоположение поля определяется не только по зрительному ориентиру Muller, Kubie, 1987
 Изменение геометрии среды – один из основных факторов, вызывающих") Свойства клеток места (геометрия ) Изменение геометрии среды – один из основных факторов, вызывающих значительное изменение поля PC. Появление барьера приводит к исчезновению поля PC Muller, Kubie, 1987 Клетка (А) изменяет форму поля разряда, когда убирают преграду, закрывающую доступ в новый отсек При устранении барьера появляется поле на новом месте Wilson, Mc. Naughton, 1993
Свойства клеток места (геометрия ) Изменение геометрии среды – один из основных факторов, вызывающих значительное изменение поля PC. Появление барьера приводит к исчезновению поля PC Muller, Kubie, 1987 Клетка (А) изменяет форму поля разряда, когда убирают преграду, закрывающую доступ в новый отсек При устранении барьера появляется поле на новом месте Wilson, Mc. Naughton, 1993
 Геометрические детерминанты полей клеток места CA 1 и CA 3 рострального гиппокампа крыс Поля разрядов 4 клеток места в аренах разной формы. Крыс тестировали в аренах с непрозрачными высокими стенками. Каркасы арен и пол меняли, чтобы устранить влияние запахов. O’Keefe, Burgess, 1996 Положение поля клетки места в таких условиях определялось расстоянием от стенок арен. Изменения длины стенок приводят к перекартированию (вытягиванию, исчезновению поля, появлению дополнительного поля
Геометрические детерминанты полей клеток места CA 1 и CA 3 рострального гиппокампа крыс Поля разрядов 4 клеток места в аренах разной формы. Крыс тестировали в аренах с непрозрачными высокими стенками. Каркасы арен и пол меняли, чтобы устранить влияние запахов. O’Keefe, Burgess, 1996 Положение поля клетки места в таких условиях определялось расстоянием от стенок арен. Изменения длины стенок приводят к перекартированию (вытягиванию, исчезновению поля, появлению дополнительного поля
 Ключевые факторы: ч. стенки") Клетки места, влияние контекста (цвет стенок и/или цвет пола арены) Ключевые факторы: ч. стенки цвет пола б. цвет пола Jeffrey et al. , 2003
Клетки места, влияние контекста (цвет стенок и/или цвет пола арены) Ключевые факторы: ч. стенки цвет пола б. цвет пола Jeffrey et al. , 2003
 Изменение поля клеток места СА 1 при увеличении размера арены диаметр 76 см 152 см Изменение размеров арены и карточки не изменяет положения поля PC, но увеличивает его размеры Muller, Kubie, 1987
Изменение поля клеток места СА 1 при увеличении размера арены диаметр 76 см 152 см Изменение размеров арены и карточки не изменяет положения поля PC, но увеличивает его размеры Muller, Kubie, 1987
 Влияние размера экспериментального пространства на характеристики клеток места рострального гиппокампа Диам. 76 см 1. 8 x 1. 4 м Сравнивали характеристики клеток места в знакомых аренах разного размера. В арене большего размера клетка места демонстрирует множественные поля разрядов, в отличие от малой арены. Размер поля разряда в больших аренах больше. Но: арены различались по форме Park, Dvorak, Fenton, 2011
Влияние размера экспериментального пространства на характеристики клеток места рострального гиппокампа Диам. 76 см 1. 8 x 1. 4 м Сравнивали характеристики клеток места в знакомых аренах разного размера. В арене большего размера клетка места демонстрирует множественные поля разрядов, в отличие от малой арены. Размер поля разряда в больших аренах больше. Но: арены различались по форме Park, Dvorak, Fenton, 2011
 Клетки места в арене без стенок Hayman, Donnett, Jeffrey, 2008 Множественные поля разрядов клеток места в условиях отсутствия стенок. Функциональные границы среды меньше определяют положение клеток места, чем реальные барьеры
Клетки места в арене без стенок Hayman, Donnett, Jeffrey, 2008 Множественные поля разрядов клеток места в условиях отсутствия стенок. Функциональные границы среды меньше определяют положение клеток места, чем реальные барьеры
 Клетки места формируют «карту» у слепых крыс Поля разрядов 8 клеток места у слепых крыс Частота разряда клеток ниже, чем у контрольных животных Стабильное воспроизведение В арену помещены 3 разных предмета Поле связано с предметами и воспроизводится там же при повороте арены Поле проявляется только тогда, когда крыса доходит до предмета и опознает его Save, E. et al. J. Neurosci. 1998; 18: 1818 -1826
Клетки места формируют «карту» у слепых крыс Поля разрядов 8 клеток места у слепых крыс Частота разряда клеток ниже, чем у контрольных животных Стабильное воспроизведение В арену помещены 3 разных предмета Поле связано с предметами и воспроизводится там же при повороте арены Поле проявляется только тогда, когда крыса доходит до предмета и опознает его Save, E. et al. J. Neurosci. 1998; 18: 1818 -1826
 5 серий") Перекартирование. Роль обонятельных и зрительных сигналов (ростр. CA 1 и CA 3) 5 серий по 16 мин Арена 75 см диам. Свет +/-, промывка пола (обоняние) +/Зрит. ориентир (карточка на стенке арены -
Перекартирование. Роль обонятельных и зрительных сигналов (ростр. CA 1 и CA 3) 5 серий по 16 мин Арена 75 см диам. Свет +/-, промывка пола (обоняние) +/Зрит. ориентир (карточка на стенке арены -
 Обнаружена важная") Перекартирование. Роль обонятельных и зрительных сигналов (ростр. CA 1 и CA 3) Обнаружена важная роль стабильности обонятельной и зрительной информации для сохранения характеристик клеток места
Перекартирование. Роль обонятельных и зрительных сигналов (ростр. CA 1 и CA 3) Обнаружена важная роль стабильности обонятельной и зрительной информации для сохранения характеристик клеток места
 Регистрация разрядов клеток места при обучении мыши в виртуальном пространстве C. D. Harvey, F. Collman, D. A. Dombeck, D. W. Tank. Princeton, NJ. Nature, 2009, 461, 941
Регистрация разрядов клеток места при обучении мыши в виртуальном пространстве C. D. Harvey, F. Collman, D. A. Dombeck, D. W. Tank. Princeton, NJ. Nature, 2009, 461, 941
") Мышь в виртуальной реальности (видео)
Мышь в виртуальной реальности (видео)
 Клетки места в виртуальной реальности 0 90 180 см Пространственная избирательность 5 клеток места мыши, по оси Y – частота разрядов вода Мыши успешно обучались бегать по прямому маршруту от одного конца «лабиринта» к другому в поисках воды
Клетки места в виртуальной реальности 0 90 180 см Пространственная избирательность 5 клеток места мыши, по оси Y – частота разрядов вода Мыши успешно обучались бегать по прямому маршруту от одного конца «лабиринта» к другому в поисках воды
 Оптическая визуализация клеток места у мыши, обучающейся в виртуальном лабиринте Двухфотонный лазерный микроскоп Dombeck, Harvey…Tank, 2010
Оптическая визуализация клеток места у мыши, обучающейся в виртуальном лабиринте Двухфотонный лазерный микроскоп Dombeck, Harvey…Tank, 2010
 у мыши, обучающейся в виртуальном лабиринте") Визуализация клеток места (пирамидный слой поля CA 1) у мыши, обучающейся в виртуальном лабиринте Активность нейронов оценивали по уровню флуоресценции белка GCa. MP 3, который белка служил индикатором наличия кальция в клетке Предварительно проводили инъекцию в гиппокамп аденоассоциированного вируса AAV 2/1 -synapsin-1 -GCa. MP 3, который нес ген, кодирующий этот белок. Образцы пространственного расположения клеток Разным цветом показаны группы клеток места, разряжающихся на разных участках трека
Визуализация клеток места (пирамидный слой поля CA 1) у мыши, обучающейся в виртуальном лабиринте Активность нейронов оценивали по уровню флуоресценции белка GCa. MP 3, который белка служил индикатором наличия кальция в клетке Предварительно проводили инъекцию в гиппокамп аденоассоциированного вируса AAV 2/1 -synapsin-1 -GCa. MP 3, который нес ген, кодирующий этот белок. Образцы пространственного расположения клеток Разным цветом показаны группы клеток места, разряжающихся на разных участках трека
 Соседние клетки CA 1 кодируют разные области виртуального пространства!!!
Соседние клетки CA 1 кодируют разные области виртуального пространства!!!
 Внутренние стимулы и клетки места
Внутренние стимулы и клетки места
 Характеристики клеток места изменялись, если животное не могло само передвигаться в пространстве (Foster et al. , 1989; Terrazas et al. , 2005 –поле CA 1) Крыс обучали передвигаться к корму или бегом, или сидя на платформе, движение которой они запускали, нажимая на рычаг. Если крыса ехала на платформе, то клеток места было зарегистрировано меньше. В таких клетках поля разрядов были больше, чем в том случае, когда животное передвигалось самостоятельно. Терялась пространственная специфичность (Terrazas et al. , 2005)
Характеристики клеток места изменялись, если животное не могло само передвигаться в пространстве (Foster et al. , 1989; Terrazas et al. , 2005 –поле CA 1) Крыс обучали передвигаться к корму или бегом, или сидя на платформе, движение которой они запускали, нажимая на рычаг. Если крыса ехала на платформе, то клеток места было зарегистрировано меньше. В таких клетках поля разрядов были больше, чем в том случае, когда животное передвигалось самостоятельно. Терялась пространственная специфичность (Terrazas et al. , 2005)
 Полукружные каналы (передний, задний вертикальные и горизонтальный каналы) –") Вестибулярная информация nervus vestibulocochlearis (VIII) Полукружные каналы (передний, задний вертикальные и горизонтальный каналы) – сенсорная информация об угловом ускорении Отолитовые органы (утрикулус и саккулус) – гравитация и линейное ускорение
Вестибулярная информация nervus vestibulocochlearis (VIII) Полукружные каналы (передний, задний вертикальные и горизонтальный каналы) – сенсорная информация об угловом ускорении Отолитовые органы (утрикулус и саккулус) – гравитация и линейное ускорение
 Вестибулярная информация и гиппокамп контроль Smith et al. , 2005 Вестибулярная деафферентация нарушает формирование и поддержание клеток места CA 1
Вестибулярная информация и гиппокамп контроль Smith et al. , 2005 Вестибулярная деафферентация нарушает формирование и поддержание клеток места CA 1
 Блокирование вестибулярного сигнала нарушает функционирование клеток места Эффект нарушения притока вестибулярной информации у трех клеток места. Введение через барабанную перепонку тетродотоксина выключало вестибулярный сигнал на двое суток. Воздействие обратимо. Крысы могли передвигаться (чаще кругами) по арене и собирать крошки Stackman, Clark, Taube, 2002
Блокирование вестибулярного сигнала нарушает функционирование клеток места Эффект нарушения притока вестибулярной информации у трех клеток места. Введение через барабанную перепонку тетродотоксина выключало вестибулярный сигнал на двое суток. Воздействие обратимо. Крысы могли передвигаться (чаще кругами) по арене и собирать крошки Stackman, Clark, Taube, 2002
. Активное передвижение провоцировали стимуляцией") КЛЕТКИ МЕСТА В НЕВЕСОМОСТИ Escher лестничный трек (Escher staircase track). Активное передвижение провоцировали стимуляцией через электроды зон подкрепления (medial forebrain bandle) Гиппокамп может создавать представление пространства в условиях невесомости, т. е. при ограничении стимуляции от отолитовых органов Knierim, J. J. , Mc. Naughton, B. L. , Poe, G. R. (2000). Threedimensional spatial selectivity of hippocampal neurons during space flight. Nat. Neurosci. 3, 209– 210.
КЛЕТКИ МЕСТА В НЕВЕСОМОСТИ Escher лестничный трек (Escher staircase track). Активное передвижение провоцировали стимуляцией через электроды зон подкрепления (medial forebrain bandle) Гиппокамп может создавать представление пространства в условиях невесомости, т. е. при ограничении стимуляции от отолитовых органов Knierim, J. J. , Mc. Naughton, B. L. , Poe, G. R. (2000). Threedimensional spatial selectivity of hippocampal neurons during space flight. Nat. Neurosci. 3, 209– 210.
 Факторы, влияющие на характеристики полей разрядов клеток места при картировании новой среды и при перекартировании (после ее изменения) Стабильность факторов среды Внешние и внутренние стимулы Размер и геометрия среды, размерность (линейная, 2 D, 3 D) Область гиппокампа (септальная/ростральная – темпоральная/каудальная) • Область гиппокампа (поле CA 1 или CA 3, зубчатая фасция) • Видоспецифические особенности • Индивидуальный опыт животного • •
Факторы, влияющие на характеристики полей разрядов клеток места при картировании новой среды и при перекартировании (после ее изменения) Стабильность факторов среды Внешние и внутренние стимулы Размер и геометрия среды, размерность (линейная, 2 D, 3 D) Область гиппокампа (септальная/ростральная – темпоральная/каудальная) • Область гиппокампа (поле CA 1 или CA 3, зубчатая фасция) • Видоспецифические особенности • Индивидуальный опыт животного • •
, расположение на разных уровнях рострокаудальной") Поля гиппокампа (CA 1, CA 3 и зубчатая фасция), расположение на разных уровнях рострокаудальной оси
Поля гиппокампа (CA 1, CA 3 и зубчатая фасция), расположение на разных уровнях рострокаудальной оси
 Свойства клеток места разных полей гиппокампа Перенесение арены в другую комнату вызвало перекартирование в СА 3 и не изменило характеристик клетки в CA 1. Но изменение формы арены вызвало изменение положения поля разряда клетки места СА 1 S. Leutgeb , J. K. Leutgeb Learn. Mem. 2007 14: 745 -757
Свойства клеток места разных полей гиппокампа Перенесение арены в другую комнату вызвало перекартирование в СА 3 и не изменило характеристик клетки в CA 1. Но изменение формы арены вызвало изменение положения поля разряда клетки места СА 1 S. Leutgeb , J. K. Leutgeb Learn. Mem. 2007 14: 745 -757
 Клетки места гиппокампа крыс разряжаются по-разному на горизонтальной и вертикальной плоскости На плоскости, стоящей вертикально, снижается точность кодирования местоположения крысы Hayman et al. , 2011
Клетки места гиппокампа крыс разряжаются по-разному на горизонтальной и вертикальной плоскости На плоскости, стоящей вертикально, снижается точность кодирования местоположения крысы Hayman et al. , 2011
 Обучение и клетки места Клетки места гиппокампа могут изменять свое положение в ходе обучения
Обучение и клетки места Клетки места гиппокампа могут изменять свое положение в ходе обучения
 В ходе обучения клетки поля CA 1 изменяли исходное местоположение поля на более близкое к цели (кормушке). Поле воспроизводилось и в пробной попытке, без корма Dupret et al. , 2010, Csicsvari, Dupret, 2014
В ходе обучения клетки поля CA 1 изменяли исходное местоположение поля на более близкое к цели (кормушке). Поле воспроизводилось и в пробной попытке, без корма Dupret et al. , 2010, Csicsvari, Dupret, 2014
 в области цели при обучении в") Концентрация полей клеток места (d. CA 1 ) в области цели при обучении в кольцевом водном тесте Цель(платформа) Время в целевом (черная линия) и других сегментах во время тестовых попыток Наибольшее число клеток места зарегистрировано в целевой области Hollup, …. Moser, 2001
Концентрация полей клеток места (d. CA 1 ) в области цели при обучении в кольцевом водном тесте Цель(платформа) Время в целевом (черная линия) и других сегментах во время тестовых попыток Наибольшее число клеток места зарегистрировано в целевой области Hollup, …. Moser, 2001
 цель A. Поля 7 клеток места в пробную попытку B. Во время обучения клетка активна в момент перед обнаружением платформы, но не тогда, когда крыса забралась на нее Hollup, …. Moser, 2001
цель A. Поля 7 клеток места в пробную попытку B. Во время обучения клетка активна в момент перед обнаружением платформы, но не тогда, когда крыса забралась на нее Hollup, …. Moser, 2001
 Формирование дополнительной зоны активности клеток места в целевой области, участие в планировании пути старт Диам 75 см, цель 10 см Дополнительная область разряда PC в зоне ожидания, Для 4 клеток места Hok, …Poucet, 2007 Крыс обучали заходить в целевую зону и оставаться там на 2 сек. После этого давали пищевую таблетку, которую бросали случайным образом на арену. Методика позволяла разделять место подкрепления и целевую зону.
Формирование дополнительной зоны активности клеток места в целевой области, участие в планировании пути старт Диам 75 см, цель 10 см Дополнительная область разряда PC в зоне ожидания, Для 4 клеток места Hok, …Poucet, 2007 Крыс обучали заходить в целевую зону и оставаться там на 2 сек. После этого давали пищевую таблетку, которую бросали случайным образом на арену. Методика позволяла разделять место подкрепления и целевую зону.
 Клетки места у представителей других видов
Клетки места у представителей других видов
 Клетки места у летучей мыши Образцы клеток места Клетки регистрировали в септальной части поля CA 1 у летучей мыши (Бурый кожан, Eptisicus fuscus, вес 15 -17 г ) в камере 70 х70 см. Мышь ползала по арене, наклоненной под углом 70 градусов, и собирала мучных червей. Ulanovsky, Moss, 2007, 2011 Также как и у грызунов, при неизменности среды, сохраняется стабильность разряда клеток места
Клетки места у летучей мыши Образцы клеток места Клетки регистрировали в септальной части поля CA 1 у летучей мыши (Бурый кожан, Eptisicus fuscus, вес 15 -17 г ) в камере 70 х70 см. Мышь ползала по арене, наклоненной под углом 70 градусов, и собирала мучных червей. Ulanovsky, Moss, 2007, 2011 Также как и у грызунов, при неизменности среды, сохраняется стабильность разряда клеток места
 Форма поля разряда клетки места и биология вида ? ? ? Hayman et al. , 2011 Ulanovsky 2011 http: //www. youtube. com/watch? v=jw. Prn 2 Pa-Q 0 Nachum Ulanovsky, Depart. of Neurobiology, Weizmann Institute of Science, Israel Будет ли сферическим поле разряда клетки места у летучей мыши в полете?
Форма поля разряда клетки места и биология вида ? ? ? Hayman et al. , 2011 Ulanovsky 2011 http: //www. youtube. com/watch? v=jw. Prn 2 Pa-Q 0 Nachum Ulanovsky, Depart. of Neurobiology, Weizmann Institute of Science, Israel Будет ли сферическим поле разряда клетки места у летучей мыши в полете?
 были") Объемные поля клеток места в гиппокампе крыланов в полете Клетки места (CA 1) были активны в ограниченном 3 D пространстве. Размер поля 0. 5 -2 м. Все три оси кодировались со сходным разрешением. Yartsev, Ulanovsky, 2013
Объемные поля клеток места в гиппокампе крыланов в полете Клетки места (CA 1) были активны в ограниченном 3 D пространстве. Размер поля 0. 5 -2 м. Все три оси кодировались со сходным разрешением. Yartsev, Ulanovsky, 2013
,") Кодирование пространства у обезьян Саймири свободно передвигались по камере (1. 5 х 1. 2), собирали фрукты, прикрепленные на стенах камеры В поле CA 1 обнаружены пространственно специфичные клетки Ludvig et al. , 2004
Кодирование пространства у обезьян Саймири свободно передвигались по камере (1. 5 х 1. 2), собирали фрукты, прикрепленные на стенах камеры В поле CA 1 обнаружены пространственно специфичные клетки Ludvig et al. , 2004
 Гиппокамп у мыши и голубя Butler et al. , 2006
Гиппокамп у мыши и голубя Butler et al. , 2006
 Пространственно-специфичные клетки гиппокампа почтового голубя Siegel, J. J. , Nitz, D. , & Bingman, V. P. (2005). Spatial specificity of single-units in the hippocampal formation of freely moving homing pigeons. Hippocampus, 15, 26 -40.
Пространственно-специфичные клетки гиппокампа почтового голубя Siegel, J. J. , Nitz, D. , & Bingman, V. P. (2005). Spatial specificity of single-units in the hippocampal formation of freely moving homing pigeons. Hippocampus, 15, 26 -40.
 Путешествие по") Человек (пациенты с вживленными по медицинским показаниям электродами в область эпилептического очага) Путешествие по виртуальному городу (игра Yellow Cab). Слева - вид сверху, красным выделены целевые объекты Поля клеток места гиппокампа при передвижении по виртуальному городу. Jacobs et al. , 2010
Человек (пациенты с вживленными по медицинским показаниям электродами в область эпилептического очага) Путешествие по виртуальному городу (игра Yellow Cab). Слева - вид сверху, красным выделены целевые объекты Поля клеток места гиппокампа при передвижении по виртуальному городу. Jacobs et al. , 2010
 оси") Функциональная неоднородность гиппокампа вдоль септотемпоральной (рострокаудальной для грызунов) оси
Функциональная неоднородность гиппокампа вдоль септотемпоральной (рострокаудальной для грызунов) оси
 Височный") Гиппокамп вдоль рострокаудальной оси неоднороден по своим функциям Септальный полюс (ростральный =дорзальный гиппокамп) Височный полюс (каудальный=вентральный гипокамп)
Гиппокамп вдоль рострокаудальной оси неоднороден по своим функциям Септальный полюс (ростральный =дорзальный гиппокамп) Височный полюс (каудальный=вентральный гипокамп)
, расположение на разных уровнях рострокаудальной") Поля гиппокампа (CA 1, CA 3 и зубчатая фасция), расположение на разных уровнях рострокаудальной оси
Поля гиппокампа (CA 1, CA 3 и зубчатая фасция), расположение на разных уровнях рострокаудальной оси
 Молекулярные домены гиппокампа C помощью набора генетических маркеров выявлены 3 домена: Вид с латеральной стороны 1. Дорзальный (включает CA 1 d и CA 3 d) 2. Промежуточный (CA 1 i и CA 3 i) 3. Вентральный (CA 1 v и CA 3 v) Вид с медиальной стороны По маркерам различаются и поля CA 1 и CA 3 Fanselow, Dong, 2010
Молекулярные домены гиппокампа C помощью набора генетических маркеров выявлены 3 домена: Вид с латеральной стороны 1. Дорзальный (включает CA 1 d и CA 3 d) 2. Промежуточный (CA 1 i и CA 3 i) 3. Вентральный (CA 1 v и CA 3 v) Вид с медиальной стороны По маркерам различаются и поля CA 1 и CA 3 Fanselow, Dong, 2010
 Молекулярные домены гиппокампа C помощью набора генетических маркеров выявлены 3 функциональных домена: 1. Дорзальный (включает CA 1 d и CA 3 d) – больше связан с когнитивными функциями, обучением и памятью 2. Промежуточный (CA 1 i и CA 3 i) ? ? 3. Вентральный (CA 1 v и CA 3 v) – с эмоциями, мотивационным контролем Fanselow, Dong, 2010
Молекулярные домены гиппокампа C помощью набора генетических маркеров выявлены 3 функциональных домена: 1. Дорзальный (включает CA 1 d и CA 3 d) – больше связан с когнитивными функциями, обучением и памятью 2. Промежуточный (CA 1 i и CA 3 i) ? ? 3. Вентральный (CA 1 v и CA 3 v) – с эмоциями, мотивационным контролем Fanselow, Dong, 2010
 Основные связи в гиппокампальной формации Медиальн. Латеральн. септум Зубчатая фасция Энторинальная кора
Основные связи в гиппокампальной формации Медиальн. Латеральн. септум Зубчатая фасция Энторинальная кора
 Hartman, Lever, Burgess, O’Keefe, 2014
Hartman, Lever, Burgess, O’Keefe, 2014
 Отделы коры, непосредственно связанные с гиппокампом rs LEA – латеральная энторнинальная кора, MEA – медиальная энторнинальная кора, PR – периринальная кора, POR – постринальная кора, rs – ринальная борозда к ы
Отделы коры, непосредственно связанные с гиппокампом rs LEA – латеральная энторнинальная кора, MEA – медиальная энторнинальная кора, PR – периринальная кора, POR – постринальная кора, rs – ринальная борозда к ы
 Особенности входов в разные области энторинальной коры Корковые входы в ЭК организованны по «полоскам» , особенно в медиальной энторинальной коре. Латеральная полоска, которая иннервирует септальную область DG, получает полимодальную информацию зрительной и пространственной природы. Средние и медиальные полоски, иннервирующие более темпоральные уровни DG получают входы от обонятельных областей
Особенности входов в разные области энторинальной коры Корковые входы в ЭК организованны по «полоскам» , особенно в медиальной энторинальной коре. Латеральная полоска, которая иннервирует септальную область DG, получает полимодальную информацию зрительной и пространственной природы. Средние и медиальные полоски, иннервирующие более темпоральные уровни DG получают входы от обонятельных областей
 в зубчатую извилину A – полушария (мозжечок снят), MEA") Топология проекций энторинальной коры (ЭК) в зубчатую извилину A – полушария (мозжечок снят), MEA – медиальная ЭК, LEA – латеральная ЭК. – B – схема ЭК, латеральная (LB), промежуточная (IB) и медиальная полоски (MB). C – схема зубчатой извилины (DG), s/d – септальный полюс, t/v – темпоральный полюс Ростральные 2/3 DG получают проекции из латеральной полоски ЭК, а более каудальные темпоральные области DG - от медиальной полоски ЭК Kerr et al. , 2007
Топология проекций энторинальной коры (ЭК) в зубчатую извилину A – полушария (мозжечок снят), MEA – медиальная ЭК, LEA – латеральная ЭК. – B – схема ЭК, латеральная (LB), промежуточная (IB) и медиальная полоски (MB). C – схема зубчатой извилины (DG), s/d – септальный полюс, t/v – темпоральный полюс Ростральные 2/3 DG получают проекции из латеральной полоски ЭК, а более каудальные темпоральные области DG - от медиальной полоски ЭК Kerr et al. , 2007
 Функциональная неоднородность гиппокампа Bast, 2007
Функциональная неоднородность гиппокампа Bast, 2007
 Свойства клеток места в разных субрегионах гиппокампа вдоль септотемпоральной оси Пространственно-специфичные нейроны обнаружены в медиальной и вентральной части гиппокампа. Характеристики клеток места отличаются в разных субрегионах Пространственная избирательность снижается от ростральной до каудальной области гиппокампа. Предполагается, что эти области гиппокампа по-разному контролируют пространственное поведение
Свойства клеток места в разных субрегионах гиппокампа вдоль септотемпоральной оси Пространственно-специфичные нейроны обнаружены в медиальной и вентральной части гиппокампа. Характеристики клеток места отличаются в разных субрегионах Пространственная избирательность снижается от ростральной до каудальной области гиппокампа. Предполагается, что эти области гиппокампа по-разному контролируют пространственное поведение
 Клетки места при передвижении на большие расстояния 18 м Kjelstrup et al. , 2008
Клетки места при передвижении на большие расстояния 18 м Kjelstrup et al. , 2008
 Характеристики клеток места вдоль септотемпоральной оси гиппокампа 8 клеток места поля CA 3 крысы, бежавшей по дорожке длиной 18 м. Размеры поля разряда минимальны в септальной (ростральной) части и увеличиваются в средней и каудальной части. Размеры поля разряда в ростральной части на дорожке больше, чем обычно наблюдали в стандартных аренах Характеристики полей зависели от направления движения крысы. Авторы предположили, что в гиппокампе пространственная информация кодируется с разной степенью разрешения.
Характеристики клеток места вдоль септотемпоральной оси гиппокампа 8 клеток места поля CA 3 крысы, бежавшей по дорожке длиной 18 м. Размеры поля разряда минимальны в септальной (ростральной) части и увеличиваются в средней и каудальной части. Размеры поля разряда в ростральной части на дорожке больше, чем обычно наблюдали в стандартных аренах Характеристики полей зависели от направления движения крысы. Авторы предположили, что в гиппокампе пространственная информация кодируется с разной степенью разрешения.
 Поля дорзального и вентрального гиппокампа различаются по пространственной специфичности Royer, Sirota, Patel, Buzsaki The Journal of Neuroscience, 2010 • 30(5): 1777– 1787
Поля дорзального и вентрального гиппокампа различаются по пространственной специфичности Royer, Sirota, Patel, Buzsaki The Journal of Neuroscience, 2010 • 30(5): 1777– 1787
 гиппокампа Клетки места каудального (вентрального) гиппокампа имеют более широкие") Особенности клеток места каудального (вентрального) гиппокампа Клетки места каудального (вентрального) гиппокампа имеют более широкие поля разрядов (меньшую пространственную специфичность). В радиальном лабиринте одна клетка избирательно разряжалась в концах радиального лабиринта, и только в том случае, когда крыса шла в направлении из центра Клетка места рострального гиппокампа картировала место в одном из лучей лабиринта Royer, Sirota, Patel, Buzsaki The Journal of Neuroscience, 2010 • 30(5): 1777– 1787
Особенности клеток места каудального (вентрального) гиппокампа Клетки места каудального (вентрального) гиппокампа имеют более широкие поля разрядов (меньшую пространственную специфичность). В радиальном лабиринте одна клетка избирательно разряжалась в концах радиального лабиринта, и только в том случае, когда крыса шла в направлении из центра Клетка места рострального гиппокампа картировала место в одном из лучей лабиринта Royer, Sirota, Patel, Buzsaki The Journal of Neuroscience, 2010 • 30(5): 1777– 1787
 Особенности клеток места каудального гиппокампа Кодирование мотивационных и эмоциональных характеристик экспериментального пространства Интернейроны Пирамидные клетки и, в меньшей степени, интернейроны специфически разряжались в рукавах без стенок. Такая специфичность проявлялась только при движении крысы от центра лабиринта. От центра К центру Royer, Sirota, Patel, Buzsaki The Journal of Neuroscience, 2010 • 30(5): 1777– 1787
Особенности клеток места каудального гиппокампа Кодирование мотивационных и эмоциональных характеристик экспериментального пространства Интернейроны Пирамидные клетки и, в меньшей степени, интернейроны специфически разряжались в рукавах без стенок. Такая специфичность проявлялась только при движении крысы от центра лабиринта. От центра К центру Royer, Sirota, Patel, Buzsaki The Journal of Neuroscience, 2010 • 30(5): 1777– 1787
 Более каудально расположенные CA 3 нейроны гиппокампа имеют большие поля, чем ростральные. Обнаружено участие нейронов средней и каудальной области в контроле контекстных характеристик среды
Более каудально расположенные CA 3 нейроны гиппокампа имеют большие поля, чем ростральные. Обнаружено участие нейронов средней и каудальной области в контроле контекстных характеристик среды
 Клетки дорзального гиппокампа сразу начинают избирательно активироваться в одном из контекстов Клетки промежуточной и вентральной области гиппокампа обучаются точно кодировать контекст, но обладают меньшей простраственной избирательностью
Клетки дорзального гиппокампа сразу начинают избирательно активироваться в одном из контекстов Клетки промежуточной и вентральной области гиппокампа обучаются точно кодировать контекст, но обладают меньшей простраственной избирательностью
 Картирование иммуногистохимическим методом (по уровню экспрессии") Функциональные особенности каудальных областей гиппокампа грызунов (наши данные) Картирование иммуногистохимическим методом (по уровню экспрессии с. Fos) активации разных областей гиппокампа вдоль септотемпоральной (рострокаудальной) оси после обследования мышами и полевками нового радиального лабиринта или арены «открытого поля» .
Функциональные особенности каудальных областей гиппокампа грызунов (наши данные) Картирование иммуногистохимическим методом (по уровню экспрессии с. Fos) активации разных областей гиппокампа вдоль септотемпоральной (рострокаудальной) оси после обследования мышами и полевками нового радиального лабиринта или арены «открытого поля» .
 Pavel Kuptsov Ilya Lebedev Marina Pleskacheva Исследование функциональной гетерогенности гиппокампа в лаборатории физиологии и генетики поведения каф. ВНД
Pavel Kuptsov Ilya Lebedev Marina Pleskacheva Исследование функциональной гетерогенности гиппокампа в лаборатории физиологии и генетики поведения каф. ВНД
 V. M. Malygin") Collaborators K. V. Anokhin R. Deacon (Oxford, UK) V. M. Malygin
Collaborators K. V. Anokhin R. Deacon (Oxford, UK) V. M. Malygin
 лабиринт рукав Области каудального гиппокампа значительно активировались у мышей и полевок, исследовавших арену или лабиринт (Купцов и др. 2005; Купцов, 2006, Купцов, Плескачева, Анохин, 2012)
лабиринт рукав Области каудального гиппокампа значительно активировались у мышей и полевок, исследовавших арену или лабиринт (Купцов и др. 2005; Купцов, 2006, Купцов, Плескачева, Анохин, 2012)
 C 57 BL/6 Рыжая полевка Активное передвижение животных по арене «открытого поля» увеличивает активацию средних и задних отделов гиппокампа лабораторных мышей и рыжих полевок Купцов, 2006
C 57 BL/6 Рыжая полевка Активное передвижение животных по арене «открытого поля» увеличивает активацию средних и задних отделов гиппокампа лабораторных мышей и рыжих полевок Купцов, 2006
 усиливает") Плотность с-Fos положит. клеток Введение новых зрительных ориентиров (синяя, красная и зеленая линия) усиливает активацию только в ростральных отделах поля CA 3 мышей. В целом, исследование показало, что области рострального и каудального гиппокампа по-разному участвуют в контроле пространственного поведения. Область каудального гиппокампа особенно активируется при исследовательской активности животных Купцов, 2006
Плотность с-Fos положит. клеток Введение новых зрительных ориентиров (синяя, красная и зеленая линия) усиливает активацию только в ростральных отделах поля CA 3 мышей. В целом, исследование показало, что области рострального и каудального гиппокампа по-разному участвуют в контроле пространственного поведения. Область каудального гиппокампа особенно активируется при исследовательской активности животных Купцов, 2006
 Эксперименты с аренами разных размеров Ø 35 см Ø 150 см Ø 75 см Ø 220 см Размер арены влияет на характер передвижения мышей C 57 BL/6, увеличивается длина пути, вертикальная исследовательская активность, изменяется характер использования пространства и структура траектории Лебедев, Плескачева, Анохин, ЖВНД, 2012
Эксперименты с аренами разных размеров Ø 35 см Ø 150 см Ø 75 см Ø 220 см Размер арены влияет на характер передвижения мышей C 57 BL/6, увеличивается длина пути, вертикальная исследовательская активность, изменяется характер использования пространства и структура траектории Лебедев, Плескачева, Анохин, ЖВНД, 2012
 Размер арены влиял на интенсивность экспрессии с-Fos в каудальной области гиппокампа Плотность с. Fos позит. клеток Кауд. Ростр. * * CA 1 CA 3 * DG * 35 150 75 220 35 75 220 150 ** ** ** Размер арены повлиял на интенсивность экспрессии с-Fos только в каудальной области гиппокампа. Наиболее высокая активность обнаружена у мышей, осваивавших пространства большого размера. Этот эффект отмечен во всех полях, однако оказался наиболее выражен в зубчатой фасции. Ø cm Лебедев, 2012, Лебедев и др.
Размер арены влиял на интенсивность экспрессии с-Fos в каудальной области гиппокампа Плотность с. Fos позит. клеток Кауд. Ростр. * * CA 1 CA 3 * DG * 35 150 75 220 35 75 220 150 ** ** ** Размер арены повлиял на интенсивность экспрессии с-Fos только в каудальной области гиппокампа. Наиболее высокая активность обнаружена у мышей, осваивавших пространства большого размера. Этот эффект отмечен во всех полях, однако оказался наиболее выражен в зубчатой фасции. Ø cm Лебедев, 2012, Лебедев и др.
 Эксперименты с повреждением каудального гиппокампа мышей SS SL Small LS LL Large Apparatus: mice were tested in two open field arenas: st 1 trial, 75 cm diameter (N(controls)=10, N(lesions)=10) 220 cm diameter (N(controls)=11, N(lesions)=12) 20 min 2 nd trial, 20 min (ITI=24 h)
Эксперименты с повреждением каудального гиппокампа мышей SS SL Small LS LL Large Apparatus: mice were tested in two open field arenas: st 1 trial, 75 cm diameter (N(controls)=10, N(lesions)=10) 220 cm diameter (N(controls)=11, N(lesions)=12) 20 min 2 nd trial, 20 min (ITI=24 h)
 Эксперименты с повреждением каудального гиппокампа мышей Br -3. 64 mm Maximum and minimum lesion area Coordinates from Paxinos, Franklin, 2001 Частичное цитотоксическое билатеральное повреждение гиппокампа мышей с помощью введения NMDA было проведено до экспериментов
Эксперименты с повреждением каудального гиппокампа мышей Br -3. 64 mm Maximum and minimum lesion area Coordinates from Paxinos, Franklin, 2001 Частичное цитотоксическое билатеральное повреждение гиппокампа мышей с помощью введения NMDA было проведено до экспериментов
 Частичное повреждение гиппокампа повлияло на интенсивность вертикальной активности при слабом воздействии на горизонтальную активность Снижение числа вертикальных стоек в малом «открытом поле» ранее отмечено Р. Диконом при полном удалении гиппокампа мышей (Deacon et al. , 2002). Control (sham lesion) Lesion
Частичное повреждение гиппокампа повлияло на интенсивность вертикальной активности при слабом воздействии на горизонтальную активность Снижение числа вертикальных стоек в малом «открытом поле» ранее отмечено Р. Диконом при полном удалении гиппокампа мышей (Deacon et al. , 2002). Control (sham lesion) Lesion
 В большой арене частичное повреждение гиппокампа вызвало нарушение динамики угашения горизонтальной активности 1 st trial Distance, m * ** * ## # Large ## Small Arena: F(1, 410)=406, p<. 001 Group: F(1, 410)=23. 9, p<. 001 Arena. XGroup: F(1, 410)=5. 7, p<. 02
В большой арене частичное повреждение гиппокампа вызвало нарушение динамики угашения горизонтальной активности 1 st trial Distance, m * ** * ## # Large ## Small Arena: F(1, 410)=406, p<. 001 Group: F(1, 410)=23. 9, p<. 001 Arena. XGroup: F(1, 410)=5. 7, p<. 02
 Повреждение гиппокампа изменило характеристики передвижения, особенно в большой арене Мыши с поврежденным гиппокампом в большой арене больше передвигались вне зоны стенки арены, чем контрольные животные Повреждение гиппокампа Траектория передвижения снижало извилистость оперированных мышей в траектории большой арене была менее сегментирована
Повреждение гиппокампа изменило характеристики передвижения, особенно в большой арене Мыши с поврежденным гиппокампом в большой арене больше передвигались вне зоны стенки арены, чем контрольные животные Повреждение гиппокампа Траектория передвижения снижало извилистость оперированных мышей в траектории большой арене была менее сегментирована
 Удаление гиппокампа увеличило число скоростных протяженных сегментов у животных, передвигающихся в большой арене Class Maximum speed 1 V<5 cm/sec 2 5≤V<10 cm/sec 3 10≤V<40 cm/sec 4 40≤V<70 cm/sec 5 V≥ 70 cm/sec + - comparison with another arena * - comparison with another group длительность Segment Analyzer (Anokhin, Mukhina, 2005)
Удаление гиппокампа увеличило число скоростных протяженных сегментов у животных, передвигающихся в большой арене Class Maximum speed 1 V<5 cm/sec 2 5≤V<10 cm/sec 3 10≤V<40 cm/sec 4 40≤V<70 cm/sec 5 V≥ 70 cm/sec + - comparison with another arena * - comparison with another group длительность Segment Analyzer (Anokhin, Mukhina, 2005)
 Изменение сегментной структуры траектории при удалении каудального гиппокампа у мышей и рыжих полевок Mice Voles Samples of animal tracks in large arena (diam. 220 cm), Segment Analyzer software (Anokhin, Mukhina). Red lines mark high-speed segments
Изменение сегментной структуры траектории при удалении каудального гиппокампа у мышей и рыжих полевок Mice Voles Samples of animal tracks in large arena (diam. 220 cm), Segment Analyzer software (Anokhin, Mukhina). Red lines mark high-speed segments
 Изменение характеристик сегментов пути после повреждения каудальной части гиппокампа у мышей: увеличивается доля протяженных высокоскоростных сегментов Yellow area marks 95%percentile for control group. The portion of segments located out of that area is higher in lesioned mice: 150 cm: p<0. 05 220 cm: p<0. 01 (Mann–Whitney U-test) Portion of segments out of yellow area in arena 220 cm: 5, 8 % in controls 9, 3 % in lesions
Изменение характеристик сегментов пути после повреждения каудальной части гиппокампа у мышей: увеличивается доля протяженных высокоскоростных сегментов Yellow area marks 95%percentile for control group. The portion of segments located out of that area is higher in lesioned mice: 150 cm: p<0. 05 220 cm: p<0. 01 (Mann–Whitney U-test) Portion of segments out of yellow area in arena 220 cm: 5, 8 % in controls 9, 3 % in lesions
 Спасибо за внимание
Спасибо за внимание
 Три проекции поля CA 1 1 Дорзальная часть CA 1 2 Промежуточная и вентральная часть CA 1 3 CA 1 Sub EC EC Ретросплениальная кора (Цингулярная кора) Зрительные, соматосенсорные, обонятельные, вкусовые, висцеральные корковые области, базолатеральный комплекс амигдалы, инсулярные и орбитальные области коры Система свода Префронтальные, орбитальные, обонятельные области коры
Три проекции поля CA 1 1 Дорзальная часть CA 1 2 Промежуточная и вентральная часть CA 1 3 CA 1 Sub EC EC Ретросплениальная кора (Цингулярная кора) Зрительные, соматосенсорные, обонятельные, вкусовые, висцеральные корковые области, базолатеральный комплекс амигдалы, инсулярные и орбитальные области коры Система свода Префронтальные, орбитальные, обонятельные области коры
 Ансамбли клеток поля CA 1 гиппокампа, реконструкция траектории передвижения крысы Поля разрядов ансамбля клеток места
Ансамбли клеток поля CA 1 гиппокампа, реконструкция траектории передвижения крысы Поля разрядов ансамбля клеток места
 Особенности “пространственной карты” в гиппокампе грызунов • Установленная «карта» стабильна (воспроизводится при повторном попадании животного на данную территорию), если нет изменений окружающей среды. • Изменение среды (появление новых объектов на территории, дистантных зрительных сигналов, появление барьеров, изменение формы, расширение экспериментальной территории) вызывает «перекартирование» : активируются клетки места, ранее неактивные, изменяется положение полей и частота разряда уже работавших клеток
Особенности “пространственной карты” в гиппокампе грызунов • Установленная «карта» стабильна (воспроизводится при повторном попадании животного на данную территорию), если нет изменений окружающей среды. • Изменение среды (появление новых объектов на территории, дистантных зрительных сигналов, появление барьеров, изменение формы, расширение экспериментальной территории) вызывает «перекартирование» : активируются клетки места, ранее неактивные, изменяется положение полей и частота разряда уже работавших клеток
 Краткое содержание предыдущей серии…. Ч. 1 1. Эдвард Толмен: идея когнитивной карты 1. 1 Ментальное представление среды 1. 2 Интегрирование информации о среде 1. 3 В мозге существует «интегратор» 1. 4 «Широкая» карта «. . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным» 1. 5 «Широкая» карта дает возможность выбирать оптимальный путь с любых стартовых позиций , «узкая» карта при изменении условий среды становиться непригодной 2. O’Keefe, Nadel «Гиппокамп как когнитивная карта» 2. 1 В гиппокампе функционирует система, создающая ментальное представление (когнитивную карту) среды. Гиппокамп как интегратор 2. 2 Карты (представление набора связанных мест, части среды, гибкие, устойчивы к помехам) и маршруты (привязанность к специфическому сигналу или элементу среды, ригидны)
Краткое содержание предыдущей серии…. Ч. 1 1. Эдвард Толмен: идея когнитивной карты 1. 1 Ментальное представление среды 1. 2 Интегрирование информации о среде 1. 3 В мозге существует «интегратор» 1. 4 «Широкая» карта «. . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным» 1. 5 «Широкая» карта дает возможность выбирать оптимальный путь с любых стартовых позиций , «узкая» карта при изменении условий среды становиться непригодной 2. O’Keefe, Nadel «Гиппокамп как когнитивная карта» 2. 1 В гиппокампе функционирует система, создающая ментальное представление (когнитивную карту) среды. Гиппокамп как интегратор 2. 2 Карты (представление набора связанных мест, части среды, гибкие, устойчивы к помехам) и маршруты (привязанность к специфическому сигналу или элементу среды, ригидны)
 Когнитивные карты. Представления Толмена В процессе обучения в мозге крысы образуется нечто подобное карте окружающей среды. …Крыса в процессе пробежки по лабиринту подвергается воздействию стимулов и в результате этого воздействия появляются ответные реакции. Однако вмешивающиеся мозговые процессы являются более сложными, более структурированными и часто, …более автономными, чем об этом говорят психологи, придерживающиеся теории «стимул-реакция» … Поступающие стимулы перерабатываются в центральной диспетчерской в предполагаемую когнитивную карту окружающей среды. . . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным. Карты могут быть «узкие» (простой одиночный путь от данной позиции животного до цели и «широкие» (обширная область среды). Если стартовая позиция животного изменится, широкая карта позволит ему достаточно корректно перемещаться и выбрать оптимальный новый путь. …При изменении условий окружающей среды узкая карта окажется непригодной
Когнитивные карты. Представления Толмена В процессе обучения в мозге крысы образуется нечто подобное карте окружающей среды. …Крыса в процессе пробежки по лабиринту подвергается воздействию стимулов и в результате этого воздействия появляются ответные реакции. Однако вмешивающиеся мозговые процессы являются более сложными, более структурированными и часто, …более автономными, чем об этом говорят психологи, придерживающиеся теории «стимул-реакция» … Поступающие стимулы перерабатываются в центральной диспетчерской в предполагаемую когнитивную карту окружающей среды. . . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным. Карты могут быть «узкие» (простой одиночный путь от данной позиции животного до цели и «широкие» (обширная область среды). Если стартовая позиция животного изменится, широкая карта позволит ему достаточно корректно перемещаться и выбрать оптимальный новый путь. …При изменении условий окружающей среды узкая карта окажется непригодной
 Краткое содержание предыдущей серии…. Ч. 2 2. 3 Исследовательская активность – необходимый компонент построения и контроля карты 2. 4 Клетки места – функциональные элементы карты 3. Клетки места (PC). Свойства 3. 1 Пространство контролируется ансамблем клеток места (КМ 3. 2 Картирование (Mapping) – формирование стабильных по положению в пространстве и частоте разряда полей PC в новой среде 3. 4 Перекартирование (remapping) – изменение характеристик PC при изменениях в знакомой среде 3. 5 Факторы внешней среды и внутренние сигналы (вестибулярные и кинестетические) могут влиять на характеристики PC
Краткое содержание предыдущей серии…. Ч. 2 2. 3 Исследовательская активность – необходимый компонент построения и контроля карты 2. 4 Клетки места – функциональные элементы карты 3. Клетки места (PC). Свойства 3. 1 Пространство контролируется ансамблем клеток места (КМ 3. 2 Картирование (Mapping) – формирование стабильных по положению в пространстве и частоте разряда полей PC в новой среде 3. 4 Перекартирование (remapping) – изменение характеристик PC при изменениях в знакомой среде 3. 5 Факторы внешней среды и внутренние сигналы (вестибулярные и кинестетические) могут влиять на характеристики PC
 Edward C. Tolman. Когнитивные карты «В процессе обучения в мозге крысы образуется нечто подобное карте окружающей среды. …Поступающие стимулы перерабатываются в центральной диспетчерской в предполагаемую когнитивную карту окружающей среды. . . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным. » 1886 -1959 University of California in Berkeley Критик классического бихевиоризма (Уотсона), упрощенного представления о поведении как о совокупности элементарных ответов на стимулы. Идея «когнитивной карты» “Cognitive maps in rats and men”, Psychol Rev. 1948; 55: 189 -208 + +
Edward C. Tolman. Когнитивные карты «В процессе обучения в мозге крысы образуется нечто подобное карте окружающей среды. …Поступающие стимулы перерабатываются в центральной диспетчерской в предполагаемую когнитивную карту окружающей среды. . . Эта карта, указывая маршруты, пути и взаимосвязи элементов окружающей среды, окончательно определяет, какие ответные реакции …будут выполняться животным. » 1886 -1959 University of California in Berkeley Критик классического бихевиоризма (Уотсона), упрощенного представления о поведении как о совокупности элементарных ответов на стимулы. Идея «когнитивной карты» “Cognitive maps in rats and men”, Psychol Rev. 1948; 55: 189 -208 + +
 Первые эксперименты, регистрация активности клетки, разряжающейся в определенном месте пространства лабиринта O’Keefe, 1976
Первые эксперименты, регистрация активности клетки, разряжающейся в определенном месте пространства лабиринта O’Keefe, 1976
 Place representation как часть когнитивной карты Строятся с использованием двух типов стимулов : На основе набора внешних стимулов среды На основе внутренних стимулов от организма (dead reckoning=path integration), дополняющих информацию от набора внешних стимулов. Взаимодействие систем важно при движении организма.
Place representation как часть когнитивной карты Строятся с использованием двух типов стимулов : На основе набора внешних стимулов среды На основе внутренних стимулов от организма (dead reckoning=path integration), дополняющих информацию от набора внешних стимулов. Взаимодействие систем важно при движении организма.
 Определение понятия «карты» по O’Keefe, Nadel, 1978 The simplest definition of a map is that it is the representation (usually two dimensional) of a part of space. The constituents of space are places, and thus an alternative definition of a map is the representation of a set of connected places which are systematically related to each other by a group of spatial transformation rules. Простейшее определение карты: это представление (обычно двухмерное) части пространства. Места – это компоненты пространства и, следовательно, альтернативное определение карты - это представление набора связанных мест The location of an object in this space is … a function (although only indirectly) of the organism's movements in space relative to that object. Местоположение объекта в локальном пространстве есть функция (хотя и не напрямую) передвижений организма в пространстве относительно этого объекта КАРТИРОВАНИЕ ПРОИСХОДИТ ЧЕРЕЗ ПЕРЕДВИЖЕНИЕ
Определение понятия «карты» по O’Keefe, Nadel, 1978 The simplest definition of a map is that it is the representation (usually two dimensional) of a part of space. The constituents of space are places, and thus an alternative definition of a map is the representation of a set of connected places which are systematically related to each other by a group of spatial transformation rules. Простейшее определение карты: это представление (обычно двухмерное) части пространства. Места – это компоненты пространства и, следовательно, альтернативное определение карты - это представление набора связанных мест The location of an object in this space is … a function (although only indirectly) of the organism's movements in space relative to that object. Местоположение объекта в локальном пространстве есть функция (хотя и не напрямую) передвижений организма в пространстве относительно этого объекта КАРТИРОВАНИЕ ПРОИСХОДИТ ЧЕРЕЗ ПЕРЕДВИЖЕНИЕ
. The hippocampus as") Гиппокамп как когнитивная карта O'KEEFE, J. , and DOSTROVSKY, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34, 171 -5. O'KEEFE, J. (1976). Place units in the hippocampus of the freely moving rat. Exp. Neurol. 51, 78 -109. В гиппокампе обнаружены клетки, разряжающиеся только тогда, когда крысы находились в определенном месте экспериментального пространства.
Гиппокамп как когнитивная карта O'KEEFE, J. , and DOSTROVSKY, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34, 171 -5. O'KEEFE, J. (1976). Place units in the hippocampus of the freely moving rat. Exp. Neurol. 51, 78 -109. В гиппокампе обнаружены клетки, разряжающиеся только тогда, когда крысы находились в определенном месте экспериментального пространства.
 «Когда животное двигается, «внутренняя навигационная» система будет переключать фокус") Когнитивные карты (O’Keefe, Nadel, 1978) «Когда животное двигается, «внутренняя навигационная» система будет переключать фокус возбуждения внутри карты к другим представлениям места, соответствующим другим местоположениям. Это предположительно происходит на основе информации о дистанциях, сгенерированной от сенсорных и моторных входов. »
Когнитивные карты (O’Keefe, Nadel, 1978) «Когда животное двигается, «внутренняя навигационная» система будет переключать фокус возбуждения внутри карты к другим представлениям места, соответствующим другим местоположениям. Это предположительно происходит на основе информации о дистанциях, сгенерированной от сенсорных и моторных входов. »
, упрощенного представления о") Edward C. Tolman. Когнитивные карты Когнитивный бихевиорист, критик классического бихевиоризма (Уотсона), упрощенного представления о поведении как о совокупности элементарных ответов на стимулы. Идея «когнитивной карты» Purposive Behavior in Animals and Men, 1932 1886 -1959 University of California in Berkeley “Cognitive maps in rats and men”, Psychol Rev. 1948; 55: 189 -208 Tolman, E. C. ; Ritchie, B. F. ; Kalish, D. : Studies in spatial learning: Orientation and the short-cut. J. Exp Psychol. 1946; 36: 13 -24 Studies in spatial learning. II. Place learning versus response learning. J. of Exp Psych. 1946, 36: 221 -229.
Edward C. Tolman. Когнитивные карты Когнитивный бихевиорист, критик классического бихевиоризма (Уотсона), упрощенного представления о поведении как о совокупности элементарных ответов на стимулы. Идея «когнитивной карты» Purposive Behavior in Animals and Men, 1932 1886 -1959 University of California in Berkeley “Cognitive maps in rats and men”, Psychol Rev. 1948; 55: 189 -208 Tolman, E. C. ; Ritchie, B. F. ; Kalish, D. : Studies in spatial learning: Orientation and the short-cut. J. Exp Psychol. 1946; 36: 13 -24 Studies in spatial learning. II. Place learning versus response learning. J. of Exp Psych. 1946, 36: 221 -229.
 • Беспорядочная навигация (random navigation). Нет") Способы навигации при поиске целевого объекта (Redish, 1999) • Беспорядочная навигация (random navigation). Нет информации, где расположен целевой объект. Случайный поиск. • Навигация по ориентиру (taxon navigation). Виден сам объект или заметный ориентир около него • Праксисная навигация (praxis navigation). Использование выученных моторных программ. Удобна при стабильном положении объекта и места старта • Маршрутная навигация (route navigation). Животное обучается ассоциировать нужное направление с каким-либо сенсорным сигналом (цепь последовательных перемещений). Очевидно, совмещает принципы двух предыдущих стратегий • Навигация по «карте» (locale navigation). Животное должно выучить «карту» местности. Если ему известно свое местоположение на данной известной территории и положение объекта, тогда может быть спланировано и осуществлено передвижение к объекту по оптимальной траектории.
Способы навигации при поиске целевого объекта (Redish, 1999) • Беспорядочная навигация (random navigation). Нет информации, где расположен целевой объект. Случайный поиск. • Навигация по ориентиру (taxon navigation). Виден сам объект или заметный ориентир около него • Праксисная навигация (praxis navigation). Использование выученных моторных программ. Удобна при стабильном положении объекта и места старта • Маршрутная навигация (route navigation). Животное обучается ассоциировать нужное направление с каким-либо сенсорным сигналом (цепь последовательных перемещений). Очевидно, совмещает принципы двух предыдущих стратегий • Навигация по «карте» (locale navigation). Животное должно выучить «карту» местности. Если ему известно свое местоположение на данной известной территории и положение объекта, тогда может быть спланировано и осуществлено передвижение к объекту по оптимальной траектории.
 Праксисная навигация Требуется запомнить последовательность поворотов в точках разветвления туннелей: ЛППЛ Последовательности поворотов и пройденная дистанция на каждом отрезке (7 Л 6 Л 4 П 6 П 8 Л)
Праксисная навигация Требуется запомнить последовательность поворотов в точках разветвления туннелей: ЛППЛ Последовательности поворотов и пройденная дистанция на каждом отрезке (7 Л 6 Л 4 П 6 П 8 Л)
 Таксонная навигация Движение направлено в сторону видимого ориентира, находящегося рядом с местом финиша
Таксонная навигация Движение направлено в сторону видимого ориентира, находящегося рядом с местом финиша
 Маршрутная навигация Дойти до вырубки и налево, до конца вырубки, Л 4, П до пня, от пня направо до нарядной елки, от нее налево. Снижаются требования к контролю дистанции и направления, но необходим контроль зрительных ориентиров
Маршрутная навигация Дойти до вырубки и налево, до конца вырубки, Л 4, П до пня, от пня направо до нарядной елки, от нее налево. Снижаются требования к контролю дистанции и направления, но необходим контроль зрительных ориентиров
 Навигация по карте Необходимо наличие нескольких дистантных ориентиров. Требует знания об их местоположении и возможности оценки своего местоположения относительно этих ориентиров. Требуется знание о местоположении цели относительно дистантных ориентиров. Позволяет достигать цели из разных точек по оптимальному пути.
Навигация по карте Необходимо наличие нескольких дистантных ориентиров. Требует знания об их местоположении и возможности оценки своего местоположения относительно этих ориентиров. Требуется знание о местоположении цели относительно дистантных ориентиров. Позволяет достигать цели из разных точек по оптимальному пути.
 ”Keefe, Nadel, 978 while burrowing. Rats were allowed to run through an opaque tube placed in a sandbox, leading from start (A) to goal (B). Following this, they were required to burrow from A to B under either of two conditions: with or without sight of the visual landmarks associated with start and goal at the initiation of a trial. Under both conditions rats succeeded in getting to the goal through the sand, though visual cues facilitated performance noticeably. The authors excluded the possibility that magnetic, olfactory, or visual cues were used during the actual burrowing. Further, the routes taken by the rats did not match the shape of the routes used in the training tubes, eliminating the possibility that fixed sequences of responses were involved. Thus, they concluded that 'the burrowing rats use a complex system of kinesthetic-vestibular cues for orientation coupled with a previously acquired mental map' (p. 667). The fact that accurate burrowing could be accomplished without visual cues indicates
”Keefe, Nadel, 978 while burrowing. Rats were allowed to run through an opaque tube placed in a sandbox, leading from start (A) to goal (B). Following this, they were required to burrow from A to B under either of two conditions: with or without sight of the visual landmarks associated with start and goal at the initiation of a trial. Under both conditions rats succeeded in getting to the goal through the sand, though visual cues facilitated performance noticeably. The authors excluded the possibility that magnetic, olfactory, or visual cues were used during the actual burrowing. Further, the routes taken by the rats did not match the shape of the routes used in the training tubes, eliminating the possibility that fixed sequences of responses were involved. Thus, they concluded that 'the burrowing rats use a complex system of kinesthetic-vestibular cues for orientation coupled with a previously acquired mental map' (p. 667). The fact that accurate burrowing could be accomplished without visual cues indicates
 has described hunting behaviour in wolf-packs which, he feels, necessitates the use") Peters (1973) has described hunting behaviour in wolf-packs which, he feels, necessitates the use of the cognitive mapping concept. He cites three pieces of evidence: (1) wolves can take intentional short cuts, or detours; (2) packs can split up and re-group at some distant point, beyond the effective range of howling, such that some idea of distance and direction is required; (3) wolves can return to a rendezvous point where pups have been left from any direction. All this strongly implies a map-like organization of their psychological space. Peters stresses that these maps are likely to be heavily dependent upon smells, an important point as it extends the notion of mapping to other modalities. We assume, in fact, that all cognitive maps are at least potentially multi -modal. ∗ ∗ Peters also discusses why maps would provide a firmer basis for behaviour than routes. We shall be discussing this in the next section.
Peters (1973) has described hunting behaviour in wolf-packs which, he feels, necessitates the use of the cognitive mapping concept. He cites three pieces of evidence: (1) wolves can take intentional short cuts, or detours; (2) packs can split up and re-group at some distant point, beyond the effective range of howling, such that some idea of distance and direction is required; (3) wolves can return to a rendezvous point where pups have been left from any direction. All this strongly implies a map-like organization of their psychological space. Peters stresses that these maps are likely to be heavily dependent upon smells, an important point as it extends the notion of mapping to other modalities. We assume, in fact, that all cognitive maps are at least potentially multi -modal. ∗ ∗ Peters also discusses why maps would provide a firmer basis for behaviour than routes. We shall be discussing this in the next section.
 Стратегии навигации. Интеграция пути
Стратегии навигации. Интеграция пути
: Прослеживание своего местоположения от референтной") • Интеграция пути (path integration=dead reckoning, счисление пути): Прослеживание своего местоположения от референтной точки (напр. места старта) по внутренним сигналам, которые генерируются при передвижении Не требует внешних ориентиров.
• Интеграция пути (path integration=dead reckoning, счисление пути): Прослеживание своего местоположения от референтной точки (напр. места старта) по внутренним сигналам, которые генерируются при передвижении Не требует внешних ориентиров.
 Навигация на основе «интеграции пути» (path integration=dead reckoning ( Mittelstaedt, Mittelstaedt 1973, Gallistel, 1990; Etienne et al. , 1996) • Навигация реализуется за счет внутренних сигналов, поступающих от самого движущегося животного (кинестетическая, вестибулярная и др. информация) • Обеспечивает целенаправленное передвижение, когда зрительная информация недоступна • Невысокая точность навигации, с увеличением длины пути отклонение от планируемого пути увеличивается
Навигация на основе «интеграции пути» (path integration=dead reckoning ( Mittelstaedt, Mittelstaedt 1973, Gallistel, 1990; Etienne et al. , 1996) • Навигация реализуется за счет внутренних сигналов, поступающих от самого движущегося животного (кинестетическая, вестибулярная и др. информация) • Обеспечивает целенаправленное передвижение, когда зрительная информация недоступна • Невысокая точность навигации, с увеличением длины пути отклонение от планируемого пути увеличивается
 Врангель Фердинанд Петрович (17961870), адмирал,") История экспериментального изучения пространственного поведения Ч. Дарвин (1809 -1882) Врангель Фердинанд Петрович (17961870), адмирал, российский полярный исследователь. "Путешествие по северным берегам Сибири по Ледовитому морю, 18201824" Darwin, C. 1873. Origin of certain instincts. Nature, 7, 417– 418.
История экспериментального изучения пространственного поведения Ч. Дарвин (1809 -1882) Врангель Фердинанд Петрович (17961870), адмирал, российский полярный исследователь. "Путешествие по северным берегам Сибири по Ледовитому морю, 18201824" Darwin, C. 1873. Origin of certain instincts. Nature, 7, 417– 418.
 Murphy JJ. 1873. Instinct: a mechanical analogy. Nature 7: 483.
Murphy JJ. 1873. Instinct: a mechanical analogy. Nature 7: 483.
 Эгоцентрическая навигационная стратегия • Определение своего местоположения и направления передвижения только на основе «внутренних» сигналов (вестибулярных, проприоцептивных, а также motor efference? моторной эфферентации и др. ) самого передвигающегося животного.
Эгоцентрическая навигационная стратегия • Определение своего местоположения и направления передвижения только на основе «внутренних» сигналов (вестибулярных, проприоцептивных, а также motor efference? моторной эфферентации и др. ) самого передвигающегося животного.
 История экспериментального изучения функций мозга В 1825 г. Предложил экспериментальный метод удаления частей мозга у животных (кролики и голуби) для исследования локализации функций. Разные части мозга (полушария, мозжечок, ствол …. ) отвечают за разные функции. Исследовал последствия повреждения полукружных каналов Jean Pierre Flourens (1794 -1867) Франция Обнаружил анестезирующий эффект хлороформа
История экспериментального изучения функций мозга В 1825 г. Предложил экспериментальный метод удаления частей мозга у животных (кролики и голуби) для исследования локализации функций. Разные части мозга (полушария, мозжечок, ствол …. ) отвечают за разные функции. Исследовал последствия повреждения полукружных каналов Jean Pierre Flourens (1794 -1867) Франция Обнаружил анестезирующий эффект хлороформа
 Рецепторы растяжения в скелетных мышцах у млекопитающих - мышечные") Проприоцептивная информация Орган Гольджи (сухожилия) Рецепторы растяжения в скелетных мышцах у млекопитающих - мышечные веретена
Проприоцептивная информация Орган Гольджи (сухожилия) Рецепторы растяжения в скелетных мышцах у млекопитающих - мышечные веретена
ЦЕПТОРЫ (англ. proprioceptors; от лат. proprius — собственный + receptor —") • • ПРОПРИО(РЕ)ЦЕПТОРЫ (англ. proprioceptors; от лат. proprius — собственный + receptor — принимающий) — чувствительные аппараты (рецепторы) мышц, сухожилий и суставов. Наиболее изучены мышечные П. (т. н. мышечные веретена) и сухожильные П. (тельца Гольджи). Установлено, что мышечные веретена регистрируют расслабления и удлинения мышц, а рецепторы сухожилий — напряжения при сокращении и растяжении мышц. В суставных сумках обнаружены свободные нервные окончания, рецепторы типа окончаний Руффини, тельца Гольджи и тельца Пачини. Импульсы от П. по центростремительным путям поступают через задние корешки спинного мозга в подкорковые ядра (таламус) и затем в заднюю часть постцентральной извилины коры, образующую корковое ядро полисенсорного двигательного анализатора. Др. часть проприоцепторных импульсов по коллатералям передается в ретикулярную систему ствола мозга и мозжечок.
• • ПРОПРИО(РЕ)ЦЕПТОРЫ (англ. proprioceptors; от лат. proprius — собственный + receptor — принимающий) — чувствительные аппараты (рецепторы) мышц, сухожилий и суставов. Наиболее изучены мышечные П. (т. н. мышечные веретена) и сухожильные П. (тельца Гольджи). Установлено, что мышечные веретена регистрируют расслабления и удлинения мышц, а рецепторы сухожилий — напряжения при сокращении и растяжении мышц. В суставных сумках обнаружены свободные нервные окончания, рецепторы типа окончаний Руффини, тельца Гольджи и тельца Пачини. Импульсы от П. по центростремительным путям поступают через задние корешки спинного мозга в подкорковые ядра (таламус) и затем в заднюю часть постцентральной извилины коры, образующую корковое ядро полисенсорного двигательного анализатора. Др. часть проприоцепторных импульсов по коллатералям передается в ретикулярную систему ствола мозга и мозжечок.
 Навигационные стратегии, при которых используются сигналы из окружающей среды
Навигационные стратегии, при которых используются сигналы из окружающей среды
 Смена навигационных стратегий в ходе опыта, от новой до хорошо знакомой среды Мыши выходили из гнезда на арену (диам. 1 м) и перетаскивали (возвращали) мышат из центра арены в гнездо. Оценивали траекторию передвижения при изменении местоположения гнезда 10 проб 100 проб На ранних этапах обучения мышат возвращали в гнездо на новое место. Эгоцентрическая стратегия (интеграция пути), внешние ориентиры или предметы на арене игнорировались. После длительного обучения мыши искали гнездо на старом месте. Пространственная аллоцентрическая стратегия (дистантные зрительные ориентиры). Расположение предметов на арене не влияло на выбор пути.
Смена навигационных стратегий в ходе опыта, от новой до хорошо знакомой среды Мыши выходили из гнезда на арену (диам. 1 м) и перетаскивали (возвращали) мышат из центра арены в гнездо. Оценивали траекторию передвижения при изменении местоположения гнезда 10 проб 100 проб На ранних этапах обучения мышат возвращали в гнездо на новое место. Эгоцентрическая стратегия (интеграция пути), внешние ориентиры или предметы на арене игнорировались. После длительного обучения мыши искали гнездо на старом месте. Пространственная аллоцентрическая стратегия (дистантные зрительные ориентиры). Расположение предметов на арене не влияло на выбор пути.
 Иерархия навигационных стратегий 1. В условиях доступности дистантных зрительных ориентиров Alyan, Jander, 1994
Иерархия навигационных стратегий 1. В условиях доступности дистантных зрительных ориентиров Alyan, Jander, 1994
 Схема областей мозга, содержащих клетки, связанные с контролем навигации животного ADN- передне-дорзальное ядро переднего таламуса, DTN – дорзальное тегментальное ядро, Ento. cortex – энторинальная кора, LDN –латеральное дорзальное ядро таламуса, LMN – латеральное маммилярное ядро, Post – постсубикулюм, Retro – ретросплениальная кора Sharp, 2010
Схема областей мозга, содержащих клетки, связанные с контролем навигации животного ADN- передне-дорзальное ядро переднего таламуса, DTN – дорзальное тегментальное ядро, Ento. cortex – энторинальная кора, LDN –латеральное дорзальное ядро таламуса, LMN – латеральное маммилярное ядро, Post – постсубикулюм, Retro – ретросплениальная кора Sharp, 2010
") Пути, связывающие структуры, где обнаружены HD клетки Вестибулярные сигналы из MVN (мед. вестиб. ядро) поступают в n. prepositus, далее в дорз. тегменальное ядро (DTN), которое проецируется в латеральное мамилл. ядро (LMN), которое связано с передним дорзальным таламическим ядром (ADN). Оно дает проекции в постсубикулюм (Po. S). Oc 1 и Oc 2 – зрительная кора, Rs. A и Rs. G – ретросплениальная кора. Taube, Basset, 2003
Пути, связывающие структуры, где обнаружены HD клетки Вестибулярные сигналы из MVN (мед. вестиб. ядро) поступают в n. prepositus, далее в дорз. тегменальное ядро (DTN), которое проецируется в латеральное мамилл. ядро (LMN), которое связано с передним дорзальным таламическим ядром (ADN). Оно дает проекции в постсубикулюм (Po. S). Oc 1 и Oc 2 – зрительная кора, Rs. A и Rs. G – ретросплениальная кора. Taube, Basset, 2003
 Навигация и вестибулярная система Yoder, Taube, 2014
Навигация и вестибулярная система Yoder, Taube, 2014
 Интеграция пути у золотистых хомячков Гнездо Etienne, A. S. , Maurer, R. , Boulens, V. , Levy, A. and Rowe, T. (2004). Resetting the path integrator: a basic condition for route-based navigation. J. Exp. Biol. 207, 1491 -1508. Гнездо
Интеграция пути у золотистых хомячков Гнездо Etienne, A. S. , Maurer, R. , Boulens, V. , Levy, A. and Rowe, T. (2004). Resetting the path integrator: a basic condition for route-based navigation. J. Exp. Biol. 207, 1491 -1508. Гнездо
 до места, где была кормушка") Ошибки интеграции пути Хомячков выманивали приманкой из гнезда (D) до места, где была кормушка (P). Обратно они возвращались по прямому маршруту, но с угловой ошибкой. Эксперименты проводили при инфракрасной подсветке.
Ошибки интеграции пути Хомячков выманивали приманкой из гнезда (D) до места, где была кормушка (P). Обратно они возвращались по прямому маршруту, но с угловой ошибкой. Эксперименты проводили при инфракрасной подсветке.
: 1011– 1026 Fractionating dead") Гиппокамп и интеграция пути • Naturwissenschaften. 2008 November ; 95(11): 1011– 1026 Fractionating dead reckoning: role of the compass, odometer, logbook, and home base establishment in spatial orientation Douglas G. Wallace, Megan M. Martin, and Shawn S. Winter Department of Psychology, Northern Illinois University, De Kalb, IL 60115 -2892, USA
Гиппокамп и интеграция пути • Naturwissenschaften. 2008 November ; 95(11): 1011– 1026 Fractionating dead reckoning: role of the compass, odometer, logbook, and home base establishment in spatial orientation Douglas G. Wallace, Megan M. Martin, and Shawn S. Winter Department of Psychology, Northern Illinois University, De Kalb, IL 60115 -2892, USA
 9: 219– 228 DOI 10. 1007/s 10071 -006 -0023 -x ORIGINAL") Anim Cogn (2006) 9: 219– 228 DOI 10. 1007/s 10071 -006 -0023 -x ORIGINAL ARTICLE Douglas G. Wallace · Derek A. Hamilton · Ian Q. Whishaw Movement characteristics support a role for dead reckoning in organizing exploratory behavior
Anim Cogn (2006) 9: 219– 228 DOI 10. 1007/s 10071 -006 -0023 -x ORIGINAL ARTICLE Douglas G. Wallace · Derek A. Hamilton · Ian Q. Whishaw Movement characteristics support a role for dead reckoning in organizing exploratory behavior
 Path integration Вестибулярная информация Проприоцептивная Моторная Зрительный поток
Path integration Вестибулярная информация Проприоцептивная Моторная Зрительный поток

 КЛЕТКИ МЕСТА В НЕВЕСОМОСТИ Гиппокамп может создавать представление пространства в условиях невесомости, т. е. при ограничении стимуляции от отолитовых органов Knierim, J. J. , Mc. Naughton, B. L. , Poe, G. R. (2000). Threedimensional spatial selectivity of hippocampal neurons during space flight. Nat. Neurosci. 3, 209– 210.
КЛЕТКИ МЕСТА В НЕВЕСОМОСТИ Гиппокамп может создавать представление пространства в условиях невесомости, т. е. при ограничении стимуляции от отолитовых органов Knierim, J. J. , Mc. Naughton, B. L. , Poe, G. R. (2000). Threedimensional spatial selectivity of hippocampal neurons during space flight. Nat. Neurosci. 3, 209– 210.

 Отличия hd в разных структурах Постсубикулуюм – есть реакция на место
Отличия hd в разных структурах Постсубикулуюм – есть реакция на место
 • • • When the shape of the animal's environment is changed, for example, from a cylinder to a rectangle, a cell's preferred direction frequently shifts to a new direction without e€ecting its peak ®ring rate or directional ®ring range (Taube et al. , 1990 b). Sometimes a cell's preferred direction will be unaffected by a change in the shape of one enclosure (e. g. , going from a cylinder to a square), but will be a€ected when going to another shaped enclosure (e. g. , rectangle). Taube, 1998
• • • When the shape of the animal's environment is changed, for example, from a cylinder to a rectangle, a cell's preferred direction frequently shifts to a new direction without e€ecting its peak ®ring rate or directional ®ring range (Taube et al. , 1990 b). Sometimes a cell's preferred direction will be unaffected by a change in the shape of one enclosure (e. g. , going from a cylinder to a square), but will be a€ected when going to another shaped enclosure (e. g. , rectangle). Taube, 1998
 делятся на 3 основных типа: • 1") Проприоцепция • Проприорецепторы (механорецепторы двигательной сенсорной системы) делятся на 3 основных типа: • 1 - тельца Гольджи (оплетают сухожильные волокна мышц или свободнолежащие), • 2 - Тельца Пачини (залегают в фасциях, сухожилиях и капсулах суставов), • 3 - нервно-мышечные и нервно-сухожильные веретена (имеют удлиненную форму и лежат в толще мышц). Эти рецепторы состоят из капсулы и проходящих внутри нее интрафузальных волокон (остальные волокна мышцы - экстрафузальные). • Рецепторы первого и второго типов возбуждаются при сокращении мышцы, а веретена - при расслаблении. Поток импульсов обратной связи, поступающих от всех типов проприорецепторов, информируют ЦНС при любом состоянии мышц, при всех, даже самых малейших изменениях мышечного тонуса.
Проприоцепция • Проприорецепторы (механорецепторы двигательной сенсорной системы) делятся на 3 основных типа: • 1 - тельца Гольджи (оплетают сухожильные волокна мышц или свободнолежащие), • 2 - Тельца Пачини (залегают в фасциях, сухожилиях и капсулах суставов), • 3 - нервно-мышечные и нервно-сухожильные веретена (имеют удлиненную форму и лежат в толще мышц). Эти рецепторы состоят из капсулы и проходящих внутри нее интрафузальных волокон (остальные волокна мышцы - экстрафузальные). • Рецепторы первого и второго типов возбуждаются при сокращении мышцы, а веретена - при расслаблении. Поток импульсов обратной связи, поступающих от всех типов проприорецепторов, информируют ЦНС при любом состоянии мышц, при всех, даже самых малейших изменениях мышечного тонуса.

 Изучение механизмов пространственного поведения в России Обучался в С. -Петербургском университете (естественное отд. физ. -мат. фак-та). Студентом начал экспериментальную работу в физиологической лаборатории Николая Евгеньевича Введенского по проблеме спинно -мозговой координации рефлекторных движений. Оставлен Н. Е. Введенским при кафедре, стажировался в Голландии (Утрехт, у Р. Магнуса) – для изучения методики опытов на теплокровных животных. 1885 -1974 Бериташвили Иван Соломонович. 1915 -1919 — приват-доцент Новороссийского университета. 1919 — профессор Тбилисского университета С 1935 — директор института физиологии при Тбилисском университете 1959
Изучение механизмов пространственного поведения в России Обучался в С. -Петербургском университете (естественное отд. физ. -мат. фак-та). Студентом начал экспериментальную работу в физиологической лаборатории Николая Евгеньевича Введенского по проблеме спинно -мозговой координации рефлекторных движений. Оставлен Н. Е. Введенским при кафедре, стажировался в Голландии (Утрехт, у Р. Магнуса) – для изучения методики опытов на теплокровных животных. 1885 -1974 Бериташвили Иван Соломонович. 1915 -1919 — приват-доцент Новороссийского университета. 1919 — профессор Тбилисского университета С 1935 — директор института физиологии при Тбилисском университете 1959
 Ориентация и навигация Основные механизмы, которые вовлечены в топографическую навигацию: 1. 2. 3. Направленная ориентация (guided orientation=taxon): локализованные стимулы (объекты) непосредственно направляют движение к стимулам или от них Ориентация путем интеграции пути (path integration) Навигация по ориентирам (landmarks), основана на распознавании и запоминании конфигураций индивидуальных стационарных объектов или их совокупностей. Основная сенсорная модальность – зрение. Позволяет животному выбирать произвольное направление и дистанцию передвижения относительно определенной совокупности ориентиров. Alyan, Jander, 1994
Ориентация и навигация Основные механизмы, которые вовлечены в топографическую навигацию: 1. 2. 3. Направленная ориентация (guided orientation=taxon): локализованные стимулы (объекты) непосредственно направляют движение к стимулам или от них Ориентация путем интеграции пути (path integration) Навигация по ориентирам (landmarks), основана на распознавании и запоминании конфигураций индивидуальных стационарных объектов или их совокупностей. Основная сенсорная модальность – зрение. Позволяет животному выбирать произвольное направление и дистанцию передвижения относительно определенной совокупности ориентиров. Alyan, Jander, 1994
 • • • Беспорядочная навигация (random") Способы навигации при поиске целевого объекта (Redish, 1999) • • • Беспорядочная навигация (random navigation). Нет информации, где расположен целевой объект. Случайный поиск. ЭГОЦЕНТРИЧЕСКАЯ Навигация по ориентиру (taxon navigation). Виден сам объект или заметный ориентир около него АЛЛОЦЕНТРИЧЕСКАЯ Праксисная навигация (praxis navigation). Использование выученных моторных программ. Удобна при стабильном положении объекта и места старта ЭГОЦЕНТРИЧЕСКАЯ Маршрутная навигация (route navigation). Животное обучается ассоциировать нужное направление с каким-либо сенсорным сигналом (цепь последовательных перемещений). Очевидно, совмещает принципы двух предыдущих стратегий АЛЛОЦЕНТРИЧЕСКАЯ+ ЭГОЦЕНТРИЧЕСКАЯ Навигация по «карте» (locale navigation). Животное должно выучить «карту» местности. Если ему известно свое местоположение на данной известной территории и положение объекта, тогда может быть спланировано и осуществлено передвижение к объекту по оптимальной траектории. ЭГОЦЕНТРИЧЕСКАЯ НА НОВОЙ ТЕРРИТОРИИ, СМЕШАННАЯ НА ЗНАКОМОЙ
Способы навигации при поиске целевого объекта (Redish, 1999) • • • Беспорядочная навигация (random navigation). Нет информации, где расположен целевой объект. Случайный поиск. ЭГОЦЕНТРИЧЕСКАЯ Навигация по ориентиру (taxon navigation). Виден сам объект или заметный ориентир около него АЛЛОЦЕНТРИЧЕСКАЯ Праксисная навигация (praxis navigation). Использование выученных моторных программ. Удобна при стабильном положении объекта и места старта ЭГОЦЕНТРИЧЕСКАЯ Маршрутная навигация (route navigation). Животное обучается ассоциировать нужное направление с каким-либо сенсорным сигналом (цепь последовательных перемещений). Очевидно, совмещает принципы двух предыдущих стратегий АЛЛОЦЕНТРИЧЕСКАЯ+ ЭГОЦЕНТРИЧЕСКАЯ Навигация по «карте» (locale navigation). Животное должно выучить «карту» местности. Если ему известно свое местоположение на данной известной территории и положение объекта, тогда может быть спланировано и осуществлено передвижение к объекту по оптимальной траектории. ЭГОЦЕНТРИЧЕСКАЯ НА НОВОЙ ТЕРРИТОРИИ, СМЕШАННАЯ НА ЗНАКОМОЙ
 Информация, влияющая на функционирование клетки места 1. Дистантная зрительная информация 2. Проксимальная сенсорная информация (введение барьеров и др. ) 3. Клетки места могут формировать поля и при отсутствии зрительной информации, т. е возможно использование внутренней информации (вестибулярной, кинестетической) 4. Память и мотивация. Характеристики полей могут изменяться в ходе обучения. Best, White, Minai, 2001 и др.
Информация, влияющая на функционирование клетки места 1. Дистантная зрительная информация 2. Проксимальная сенсорная информация (введение барьеров и др. ) 3. Клетки места могут формировать поля и при отсутствии зрительной информации, т. е возможно использование внутренней информации (вестибулярной, кинестетической) 4. Память и мотивация. Характеристики полей могут изменяться в ходе обучения. Best, White, Minai, 2001 и др.
 Система когнитивного картирования содержит карту для каждой среды, которая известна организму. Гиппокамп действует как когнитивная картирующая система (называемая local system), которая генерирует представление о месте и исследовательскую активность Потеря этой системы ведет к использованию животным оставшейся экстра-гиппокампальной системы the hippocampus acts as a cognitive mapping system, which we shall call the locale system and which generates place hypotheses and exploration. Loss of this system forces an animal to rely on the remaining extra-hippocampal systems.
Система когнитивного картирования содержит карту для каждой среды, которая известна организму. Гиппокамп действует как когнитивная картирующая система (называемая local system), которая генерирует представление о месте и исследовательскую активность Потеря этой системы ведет к использованию животным оставшейся экстра-гиппокампальной системы the hippocampus acts as a cognitive mapping system, which we shall call the locale system and which generates place hypotheses and exploration. Loss of this system forces an animal to rely on the remaining extra-hippocampal systems.
 Система когнитивного картирования содержит карту для каждой среды, которая известна организму. Когнитивные карты могут быть мультимодальны Safety or danger can b Свойство безопасности или опасности должно быть атрибутом environments, or place среды или места внутри среды, environment; approac responses can be dire Гиппокамп действует как когнитивная картирующая система away from, places; pla (называемая local system), которая генерирует представление о from any other place, месте и исследовательскую активность behaviours, even whe unavailable; new path Потеря этой системы ведет к использованию животным when old ones are clo оставшейся экстра-гиппокампальной системы behaviour.
Система когнитивного картирования содержит карту для каждой среды, которая известна организму. Когнитивные карты могут быть мультимодальны Safety or danger can b Свойство безопасности или опасности должно быть атрибутом environments, or place среды или места внутри среды, environment; approac responses can be dire Гиппокамп действует как когнитивная картирующая система away from, places; pla (называемая local system), которая генерирует представление о from any other place, месте и исследовательскую активность behaviours, even whe unavailable; new path Потеря этой системы ведет к использованию животным when old ones are clo оставшейся экстра-гиппокампальной системы behaviour.
 Праксисная навигация Ego-навигация") Навигационные стратегии: ориентация от субъекта или ориентация по внешним сигналам (ориентирам) Праксисная навигация Ego-навигация Маршрутная навигация Навигация по карте Allo-навигация Таксонная навигация
Навигационные стратегии: ориентация от субъекта или ориентация по внешним сигналам (ориентирам) Праксисная навигация Ego-навигация Маршрутная навигация Навигация по карте Allo-навигация Таксонная навигация
 The cognitive-mapping system is assumed to contain") Как строятся когнитивные карты (O’Keefe, Nadel, 1978) The cognitive-mapping system is assumed to contain a map for each environment the organism has experienced. These maps are built up in the following way: The animal brings to a new situation a tabula rasa of potential place representations. One of these is chosen to represent a specific location in that environment; this automatically determines the way in which the remainder of the locations in the environment will be represented. As the animal moves, the 'internal navigation' system will shift the focus of excitation within the map to other place representations appropriate to other locations. This is done, as we have suggested, on the basis of information about distances generated from both sensory and motor inputs. In order for the distances to be 'correct' in both the internal sense (that is, purely between the locations) and in the external sense (that is, between the framework of the observer and the locations) both these sources of input must contribute. There need be no representation within the map itself of the behaviours which moved the organism through the environment and which were instrumental in constructing the map.
Как строятся когнитивные карты (O’Keefe, Nadel, 1978) The cognitive-mapping system is assumed to contain a map for each environment the organism has experienced. These maps are built up in the following way: The animal brings to a new situation a tabula rasa of potential place representations. One of these is chosen to represent a specific location in that environment; this automatically determines the way in which the remainder of the locations in the environment will be represented. As the animal moves, the 'internal navigation' system will shift the focus of excitation within the map to other place representations appropriate to other locations. This is done, as we have suggested, on the basis of information about distances generated from both sensory and motor inputs. In order for the distances to be 'correct' in both the internal sense (that is, purely between the locations) and in the external sense (that is, between the framework of the observer and the locations) both these sources of input must contribute. There need be no representation within the map itself of the behaviours which moved the organism through the environment and which were instrumental in constructing the map.
 Place representation как часть когнитивной карты Строятся с использованием двух типов стимулов : На основе набора внешних стимулов среды На основе внутренних стимулов от организма (dead reckoning=path integration), дополняющих информацию от набора внешних стимулов. Взаимодействие систем важно при движении организма. Во время построения карты моторная возвратная информация существенна присвоении (приписке) представлений вещей к соответствующему их «месту» в локальном пространстве. Когда животное в знакомой среде, активация системы без реального передвижения даст возможность животному предсказывать, что появится в этом месте During the building of a map motor feedback information is crucial in assigning representations of things to their appropriate 'place' in locale space. When an animal is in a familiar environment the activation of the system, without real movement, will enable the animal to predict what should appear in what place.
Place representation как часть когнитивной карты Строятся с использованием двух типов стимулов : На основе набора внешних стимулов среды На основе внутренних стимулов от организма (dead reckoning=path integration), дополняющих информацию от набора внешних стимулов. Взаимодействие систем важно при движении организма. Во время построения карты моторная возвратная информация существенна присвоении (приписке) представлений вещей к соответствующему их «месту» в локальном пространстве. Когда животное в знакомой среде, активация системы без реального передвижения даст возможность животному предсказывать, что появится в этом месте During the building of a map motor feedback information is crucial in assigning representations of things to their appropriate 'place' in locale space. When an animal is in a familiar environment the activation of the system, without real movement, will enable the animal to predict what should appear in what place.
 Животное приносит в новую ситуацию а tabula") Как строятся когнитивные карты (O’Keefe, Nadel, 1978) Животное приносит в новую ситуацию а tabula rasa потенциальных представлений места. Одно из них выбирается чтобы представить специфическое местоположение в этой среде, это автоматически детерминирует путь в котором будет представлен remainder? ? ? места в среде. Когда животное двигается, «внутренняя навигационная» система будет переключать фокус возбуждения внутри карты к другим представлениям места, соответствующим другим местоположениям. Это предположительно происходит на основе информации о дистанциях, сгенерированной от сенсорных и моторных входов.
Как строятся когнитивные карты (O’Keefe, Nadel, 1978) Животное приносит в новую ситуацию а tabula rasa потенциальных представлений места. Одно из них выбирается чтобы представить специфическое местоположение в этой среде, это автоматически детерминирует путь в котором будет представлен remainder? ? ? места в среде. Когда животное двигается, «внутренняя навигационная» система будет переключать фокус возбуждения внутри карты к другим представлениям места, соответствующим другим местоположениям. Это предположительно происходит на основе информации о дистанциях, сгенерированной от сенсорных и моторных входов.