Лекция 5. Историческая биогеография.ppt
- Количество слайдов: 36



Историческая биогеография
: Великое оледенение. Ледниковая теория. Перигляциальные сообщества и мамонтовая фауна • Четвертичный")
Четвертичный период (антропоген): Великое оледенение. Ледниковая теория. Перигляциальные сообщества и мамонтовая фауна • Четвертичный период, или антропоген, — самый последний отрезок кайнозоя, начавшийся около 2 млн лет назад. • Наиболее существенная черта этого времени — существование в высоких широтах нашей планеты покровных оледенений; во время ледниковых эпох они распространялись на юг до 40 -х широт, а во время межледниковий — «съеживались» до примерно нынешнего состояния (когда ими покрыта лишь Антарктида в Южном полушарии и Гренландия — в Северном). • Оледенение Северного полушария • а) в наши дни; б) в последнюю ледниковую эпоху
 и начавшийся 10– 12 тыс.")
• Разделение четвертичного периода на плейстоцен (Великое оледенение) и начавшийся 10– 12 тыс. лет назад голоцен (время, в которое мы живем) в значительной степени условно: часто говорят, что на самом деле голоцен — это просто-напросто одно из плейстоценовых межледниковий, причем даже не самое крупное. • Вероятно, вам доводилось встречать где-нибудь на краях полей и на лесных опушках окатанные гранитные валуны, иногда с характерной «штриховкой» , хотя никаких скальных выходов в окрестностях нет и в помине. По нынешнему времени любой школьник знает, что эти эрратические (т. е. блуждающие) валуны, часто достигающие размеров танка, были перенесены от мест выхода соответствующих скальных пород движением древнего ледника, покрывавшего некогда обширные территории Европы и Северной Америки.

Однако в 1837 году, когда швейцарский геолог Л. Агассис выдвинул свою теорию существования в истории Земли ледникового периода, основываясь именно на сходстве между эрратическими валунами равнинной Европы и теми окатанными штрихованными булыжниками, которые на наших глазах вытаивают из-под краев альпийских ледников, его разве что не подняли на смех. Тогда не только широкая публика, но и геологи не сомневались, что все эти валуны разносились чудовищными потоками воды и грязи во время библейского Всемирного потопа. L. Agassiz.

Биогеография — наука о закономерностях географического распространения живых существ. • Вполне очевидно, что всякое живое существо экологически связано с определенным типом ландшафта, а если смотреть шире — с одной из природных (ландшафтно-климатических) зон: белый медведь — с арктическими льдами, лось — с тайгой, верблюд — с пустыней. • С другой стороны, ареал (область распространения) некоей группы организмов в той или иной степени отражает и историю его формирования: район происхождения, время расселения, действие преград (кавиморфные грызуны, широконосые обезьяны и неполнозубые ограничены Южной Америкой, что отражает долгую изоляцию этого континента, нарушенную лишь недавно возникновением Панамского перешейка). • В соответствии с этими двумя аспектами биогеографию подразделяют на ландшафтную (которая является фактически разделом экологии) и историческую.
, или дромеда р")
Например, верблюды Одногорбый верблюд (лат. Camelus распространены ныне в пустынях dromedarius), или дромеда р Азии — Центральной (двугорбый бактриан) и Передней (одногорбый дромадер). Ландшафтная биогеография призвана ответить на вопрос о том, почему верблюды живут в зоне пустынь? ? (а не в субтропических лесах или, скажем, в тайге), Историческая — почему верблюды распространены лишь в Азии? ? ? и отсутствуют в Австралии и Северной Америке (где есть вполне вроде бы подходящие для них пустыни). Двугорбый верблюд, или бактриа н

• Возможных вариантов тут два: то ли верблюды по каким-то причинам не достигли этих континентов, то ли они здесь раньше жили, но вымерли. • Однако можно ли прийти к определенным историческим выводам, исходя лишь из особенностей их современного распространения? • Оказывается, можно. Ламы, ближайшие родственники верблюдов, распространены в Южной Америке, так что каким бы путем ни расселялись мозоленогие (хоть из Азии в Южную Америку, хоть наоборот — из Южной Америки в Азию), миновать Северную Америку они никак не могли. • Соответственно, на этом материке мозоленогие наверняка жили, но вымерли, а вот Австралии этот подотряд, видимо, не достиг — так же, как все прочие плацентарные млекопитающие (кроме летучих мышей). • Ископаемые находки подтверждают эту реконструкцию (верблюды произошли в Северной Америке и исчезли там совсем недавно), однако так бывает далеко не всегда.

Современное распространение сумчатых — Австралия и Южная Америка (опоссум проник в Северную Америку из Южной недавно, по панамскому мосту). В прежние годы, когда положение материков принималось неизменным, приходилось предполагать, что сумчатые возникли в Северном полушарии, затем проникли в Австралию (из Азии) и Южную Америку (из Северной), после чего вымерли на северных материках. По нынешним же временам логично предположить, что группа эта никогда не проникала на территорию Арктогеи (ну, кроме опоссума), будучи на протяжении всей своей истории изолированной на южных материках. Она возникла в конце мела, когда фрагмент Гондваны, объединявший Южную Америку, Австралию и Антарктиду, был уже изолирован от остального мира, и расселилась по нему до эоцена, когда от него отделилась Австралия.

• Ископаемые находки, однако, рисуют иную картину. • Сумчатые действительно появились в Южной Америке в позднем мелу, а потом через Антарктиду проникли в Австралию; но родиной их является Северная Америка, откуда они — через Гренландию — заселили также Западную Евразию, где дожили до неогена, так и не проникнув ни в Юго. Восточную Азию, ни в Африку.

САМОЕ ДРЕВНЕЕ СУМЧАТОЕ ОБИТАЛО В КИТАЕ Отлично сохранившиеся окаменелости, возраст которых порядка 125 млн. лет, были обнаружены на западе Китая. Эти останки установили новый рекорд самых старых известных сумчатых. Окаменелость древнего сумчатого, у которой сохранилась большая часть скелета, говорит о том, что оно было размером с мышь и имело обширный волосяной покров. Оно жило в начале расцвета эпохи царствования динозавров. О том, что это было сумчатое, говорит особое строение запястья и костей лодыжки, а также некоторые из зубов. Обнаруженный вид назвали Sinodelphys szalayi. Вероятно, это существо обитало в густой растительности поблизости от водоемов пресной воды. Скорее всего эта древняя сумчатая мышь питалась насекомыми и червяками. Строение тела у этого существа было предназначено для обитания на деревьях и в кустарниках.
 Тилакосмил обитал во времена миоцена на территории Южной")
• Сумчатый саблезубый тигр (тилакосмил) Тилакосмил обитал во времена миоцена на территории Южной Америки и Евразии. Его главной отличительной особенностью были огромные постоянно растущие верхние клыки, а также защитные лопасти на верхней челюсти. Размерами сумчатый саблезубый тигр был схож с теперешними ягуарами и охотился на крупных травоядных. 200 000 years old.

территории Северной Австралии Обитал Marsupial Lion. 200 000 years old.

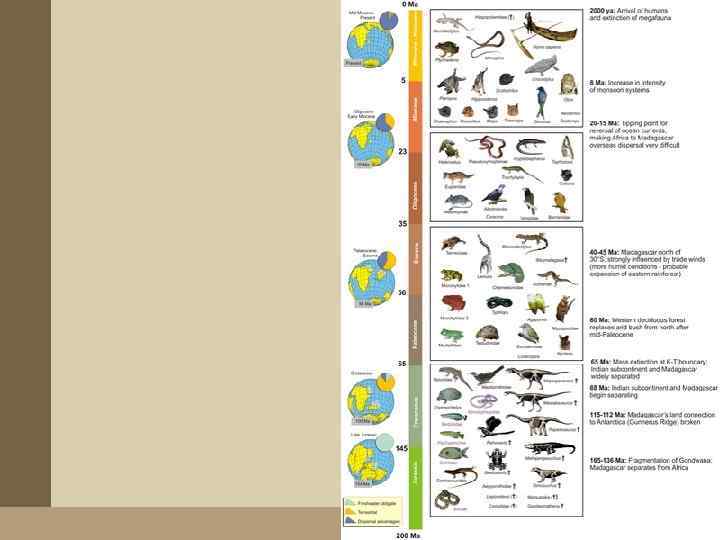
• Рассмотренная ситуация наглядно демонстрирует два момента. • Во-первых, насколько опасно в историко-биогеографических реконструкциях опираться лишь на современное распространение группы (что делается биогеографами сплошь и рядом). • Во-вторых, насколько эти реконструкции зависят от концепций, господствующих в геологии, — о корректности таких концепций биогеографы (естественно!) профессионально судить не могут и вынуждены многое принимать «на веру» . • В этом смысле весьма показательна история • изучения дизъюнктивных (разорванных) трансокеанических ареалов.

• Многие животные и растения распространены лишь на южных оконечностях материков Южного полушария, разобщенных ныне океанами. • Происхождение этих дизъюнкций издавна интриговало биогеографов. В XIX веке предполагали, что эти живые существа проникали из одной части их современного ареала в другую по неким «временным сухопутным мостам» — обширным участкам суши между континентами (расположение последних считалось тогда неизменным), которые впоследствии погрузились в океан. Типичный пример такого рода реконструкций — Лемурия, гипотетическая суша в Индийском океане, «созданная» биогеографами для того, чтобы напрямую «провести» из Африки в Индию лемуров, слонов, панголинов и т. п. Надо заметить, что сторонники этого направления быстро вошли во вкус и, по едкому замечанию Ч. Дарвина, принялись печь подобные «временные мосты» , как блины.
. • На огромном")
Гукеровская концепция • Таким последователем «мостов» был ботаник Джозеф Гукер (1860). • На огромном материале продемонстрировал высокое единство биоты материков Южного полушария (нотофагусовые леса на юге Австралии, в Новой Зеландии и в Чили). • Последователи Гукера предполагали автохтонное (на месте своего нынешнего распространения) развитие биоты южных континентов — изолированных от северных и соединенных между собой некими сухопутными связями; предполагалось, что одним из основных источников формирования этой биоты могла служить Антарктида (еще не покрытая тогда ледником и имевшая существенно больший размер, чем ныне).

Юг Южной Америки и архипелаг Огненная Земля, север в Чили Горные леса из нотофагусов в Новой Зеландии Нотофагус Домби (южный бук) Березовый бук Огненная Земля
.")
Уолесовская концепция • Иной взгляд на природу южнополушарных дизъюнкций высказывал зоолог А. Уоллес (1876). • Он полагал, что основные формообразовательные процессы идут на блоке северных континентов, откуда более молодые и продвинутые формы животных и растений оттесняют более древних и архаичных все дальше к югу. • Южные оконечности материков Южного полушария являются, таким образом, «последней линией обороны» этих реликтовых (сохранившихся от более древних эпох) групп, которые в прежние времена имели гораздо более широкое, обычно панконтинентальное, распространение. • С этой точки зрения нет принципиальной разницы между трансокеаническими дизъюнкциями, расположенными на одном материке (например, европейско-дальневосточными, как у жерлянок): и те и другие возникают в результате сокращения исходного ареала и вымирания «соединительных звеньев» .
 подтверждали ископаемые находки")
• Концепцию Уоллеса (ее позднее назвали теорией «оттесненных реликтов» ) подтверждали ископаемые находки — многие «крайне южные» ныне животные и растения (араукарии, саговники, двоякодышащие рыбы, бескилевые птицы и т. д. ) действительно обитали раньше на северных континентах. • Пауки-археиды, например, были первоначально открыты в ископаемом состоянии — в эоценовом балтийском янтаре, и лишь спустя несколько десятков лет в Южном полушарии были найдены их современные представители. • В рамках этой концепции (в отличие от гукеровской) акцент переносился на архаичность и реликтовый характер южной биоты. • Модель Уоллеса, хорошо обоснованная палеонтологическими данными и не вводящая таких «избыточных сущностей» , как сухопутные межматериковые соединения непонятной геологической природы, долго считалась главенствующей.
 по современным материкам и б) при нанесении")
Пример южнополушарной дизъюнкции: распространение родов пауков-археид а) по современным материкам и б) при нанесении его на карту Гондваны Ископаемые рода (из эоценового балтийского янтаря и из юры Казахстана) помечены крестиком (по Еськову, 1984)

• Однако с тех пор, как в геологии возобладала тектоника плит, среди биогеографов возродился интерес к гукеровской модели: ведь теперь уже не было нужды воздвигать между материками какие-то «сухопутные мосты» — достаточно сдвинуть между собой сами материки. • Эта модель позволяет рассматривать разнообразные южнополушарные дизъюнкции как прямой результат раскола Гондваны и сохранения на ее разошедшихся в результате континентального дрейфа фрагментах представителей некогда единой биоты этого мезозойского праматерика (рис. б пауки-архаиды)

Викариантная биогеография • Гукеровские построения считают одним из основных источников наиболее популярной ныне биогеографической концепции — викариантной биогеографии. • Ее задача состоит в том, чтобы согласовать последовательность ветвления филогенетического древа таксона с рисуемой геофизиками последовательностью расхождения фрагментов Пангеи.

Схема, отражающая степень родства между • Один из основоположников этого комарами-звонцами, обитающими в Южной Америке (ЮА), Австралии (АВ), Новой направления, энтомолог Л. Брундин (1966), (НЗ), на юге и на изучал комаров-звонцов (Chironomidae) Южного Зеландии континентов, Африки (АФ)(ЛВ). блоке северных в Лавразии полушария. Порядок ветвлений кладограммы действительно совпадает с порядком, в • Он показал, что они резко отличны от всех котором теряли между собой связь фрагменты северополушарных (что отражает давнюю их Пангеи (по Brundin, 1966) изоляцию на территории Гондваны); • среди южных хирономид наиболее архаичные, уклоняющиеся, формы обитают в Южной Африке, а наиболее близки между собой обитатели Австралии и Южной Америки. • Эта последовательность родственных связей вполне соответствует порядку, в котором расходились фрагменты Пангеи: • Раньше всего произошел раскол на Лавразию и Гондвану (триас). • Внутри Гондваны же последовательно теряли связь друг с другом (через Антарктиду) Южная Африка (юра), Новая Зеландия (мел), и наконец Австралия и Южная Америка (эоцен и олигоцен). Позже такие последовательности были построены для множества южнополушарных (гондванских) групп, главным образом для насекомых и иных наземных и пресноводных беспозвоночных.

Деревья - буки Южного полушария Nothofagus

Однако ……. Веские подтверждения получила в последнее время и уоллесовская теория «оттесненных реликтов» . По мере изучения мезозойских и палеогеновых насекомых стало ясно, что многие, если не большинство, из «крайне южных» групп, которыми оперируют сторонники викариантной биогеографии (в том числе те хирономиды, с которыми имел дело Брундин), в действительности были ранее широко распространены в Северном полушарии. Если искать современные аналоги той энтомофауне, что содержится в верхнемеловых янтарях Сибири и Канады, то на память приходит Новая Зеландия. Накапливается все больше аргументов в пользу того, что экзотическая биота, которая занимает ныне изолированные участки на крайнем юге южных континентов, в мезозое имела почти всесветное распространение. Сторонники теории «оттесненных реликтов» не отрицают того, что некоторые из южнополушарных ареалов могли возникнуть в результате раскола Гондваны и расхождения ее фрагментов.

Они лишь подчеркивают, что распознать такие случаи среди дизъюнкций, достоверно возникших в результате вымирания «связующих звеньев» на северных материках, в принципе невозможно. Модели Уоллеса и Гукера, возникшие из в общем-то частной проблемы происхождения биоты южных материков и акцентировавшие внимание в первом случае на ее архаичности и реликтовости, а во втором — на ее единстве (и обеспечивающих это единство механизмах), позже трансформировались в две альтернативные научные традиции. К этим двум линиям в принципе может быть сведено все многообразие концепций и моделей, существующих в исторической биогеографии. Таким образом «линия Уоллеса» и «линия Гукера» не взаимоисключающи, а взаимодополнительны: они просто рассматривают предмет исследования (ареал таксона) с разных сторон и лишь в паре дают о нем адекватное представление. В гукеровской модели таксон принимается как неизменная данность, а предметом анализа становится его расселение (преграды и их преодоление); в уоллесовской же модели исследуют таксонообразование и вымирание, полагая расселительный процесс относительно малосущественной деталью. Можно сказать, что уоллесовская модель пытается спроецировать время в пространство, а гукеровская — пространство во время.

• В своей исходной версии уоллесовская модель предполагала, что основной центр формообразования находится на блоке северных континентов, откуда всяческая архаика вытесняется на юг. • Позже это было отвергнуто; в частности, было установлено, что представления о глубокой древности и консервативности тропической биоты не соответствуют действительности • На самом деле реликты концентрируются во внетропических областях (главным образом в субтропиках) обоих полушарий, хотя в Южном, по ряду причин, сильнее, чем в Северном. • Реликтовый ареал таксона, распространенного во внетропических областях обоих полушарий, называют биполярным или амфитропическим.

• Итак, ключевой проблемой для уоллесовской схемы оказывается механизм образования такого исходного ареала, из которого можно вывести все остальные: биполярного, биполярной пары «сестринских» таксонов (таковы, например, буки и нотофагусы), или же такого типа распространения, когда более продвинутые субтаксоны приурочены к тропикам, а более архаичные — к внетропическим областям. • Необходимое для этого вымирание таксона на территории тропиков — с замещением его более продвинутыми формами или без такового — находит объяснение в двух альтернативных моделях: «экваториальной помпы» и «зональной стратификации» . деревья - буки Южного полушария Nothofagus

«Экваториальная помпа» • Термин принадлежит Ф. Дарлингтону и предполагает, что именно в тропических экосистемах с их стабильностью среды и идет постоянное и наиболее активное формообразование. • При этом реликты «выдавливаются» (помпа!) и на Север, и на Юг за счет конкуренции со стороны более молодых, продвинутых таксонов. Именно тропический пояс является главной ареной макроэволюционных событий. Отсюда сформировавшиеся таксоны при потеплениях «растекаются» во внеэкваториальные широты (фитоспрединг), где при последующих похолоданиях они либо вымирают, либо эволюционируют в более узких (обычно внутриродовых) таксономических рамках; Для внетропических областей на протяжении всей истории характерно длительное переживание архаических форм (внетропическое персистирование). При этом процесс этот строго однонаправленный: вселения бореальных групп в тропики при похолоданиях не происходит никогда.

«Зональная стратификация» • Альтернативный взгляд на природу амфитропических ареалов высказывал В. В. Жерихин (1978). • Он основан на том, что в мезозое и раннем палеогене климатическая зональность в ее современном виде отсутствовала и климат практически по всей планете был близок к нынешнему субтропическому. В конце палеогена произошла климатическая перестройка, в результате которой тепло стало распределяться по поверхности планеты менее равномерно: на полюсах стало заметно холоднее, чем прежде, а на экваторе — жарче.

• В итоге единую «теплоумеренно-субтропическую» биоту Земли охватил процесс «зональной стратификации» . • Он заключался в том, что в экваториальных и приполярных широтах (где сменился климат) начались экосистемные перестройки, приведшие к достаточно массовому вымиранию мезозойских (райский климат) и раннепалеогеновых групп. Эти группы имели наилучшие шансы выжить в двух амфитропических «кольцах» с субтропическим и теплоумеренным климатом, господствовавшим ранее по всей планете — где они и поныне существуют в статусе реликтов. • Т. е если в некий момент на полюсах стало «слишком холодно» , а на экваторе «слишком жарко» , то реликты предыдущей эпохи имеют наилучшие шансы выжить прежде всего в субтропиках, господствующих ранее по всей Земле (например, буки). • Заключение Жерихина связано с тем, что тропические сообщества, наряду с бореальными эволюционно очень молоды и принадлежат к числу самых молодых на Земле.

• Сравнение моделей «экваториальной помпы» и «зональной стратификации» позволяет прийти к достаточно неожиданному выводу о том, что они в действительности не альтернативны, а взаимодополнительны. • Фитоспрединг — это универсальный механизм флоро– и фауногенеза; он имеет три режима работы, определяемых глобальным термическим градиентом, и режимы эти формируют замкнутый трехчленный цикл: • — «экваториальная помпа» (поздний палеозой, криоэра) • — «диффузные центры формообразования» (мезозой — ранний палеоген, термоэра) • — «зональная стратификация» (поздний палеоген) — • снова «экваториальная помпа» (неоген, криоэра) и т. д.

Третичные млекопитающие Южной Америки: а — неполнозубые; б, в, г — «южноамериканские копытные» ; д, е — хищные сумчатые а — глиптодонт; б — литоптерна Thoatherium; в — литоптерна Toxodon; г — нотоунгулят Macrauchenia; д — сумчатый волк Prothylacynus, е — сумчатый саблезубый тигр Thylacosmilus; ж — череп Thylacosmilus; з — череп «северной» саблезубой кошки Smilodon (в одном масштабе)

Раннетретичные растительноядные млекопитающие Арктогеи: а — кондиляртра (группа, исходная и для «северных» , и для «южноамериканских» копытных) Phenacodus; б — диноцерат Uintotherium; в, г — парнокопытные; д, е — непарнокопытные (по Фентон, 1997, и Norman, 1994); в — «жирафовый верблюд» Aepicamelus; г — гигантская (высота в холке 2, 1 м) свинья Dinochyus; д — халикотерий Moropus; е — гигантский безрогий носорог индрикотерий

• • • Раннетретичные хищники Арктогеи: а — креодонт Patriofelis (длина около 1, 5 м); б — собакоподобная мезонихида Synoplotherium; в — голова гигантской мезонихиды Andrewsarchus (рядом, в том же масштабе, изображена голова бурого медведя); г — диатрима (по Фентон, 1997, и Norman, 1994)

• 24 вятских глухаря приехали в Польшу! Мы завершаем труднейший проект 2016 года по укреплению популяции глухарей в Пуще Августовской. За трое суток транспорт с птицами прошел 2270 км, Прошлой ночью мы готовили новоселов к новым местам обитания (взвешивание, новые кольца и радиопередатчики). Теперь идем бриться, мыться и отсыпаться. . . • 10 часов бюрократических баталий по обе стороны границы, и - птицы в ЕС • Трейлер с глухарями и машина сопровождения • в таких вольерах глухари проживут еще 30 -дневный карантин, потом - на волю
Лекция 5. Историческая биогеография.ppt