Общая презентация.ppt
- Количество слайдов: 73



ИСТОЧНИКИ ЭНЕРГИИ ДЛЯ МЕТАБОЛИЗМА






БИОТЕРМОДИНАМИКА БИОЛОГИЧЕСКИХ СИСТЕМ

Биотермодинамика: поток энергии


Биотермодинамика: законы

Биотермодинамика: законы Энтропия


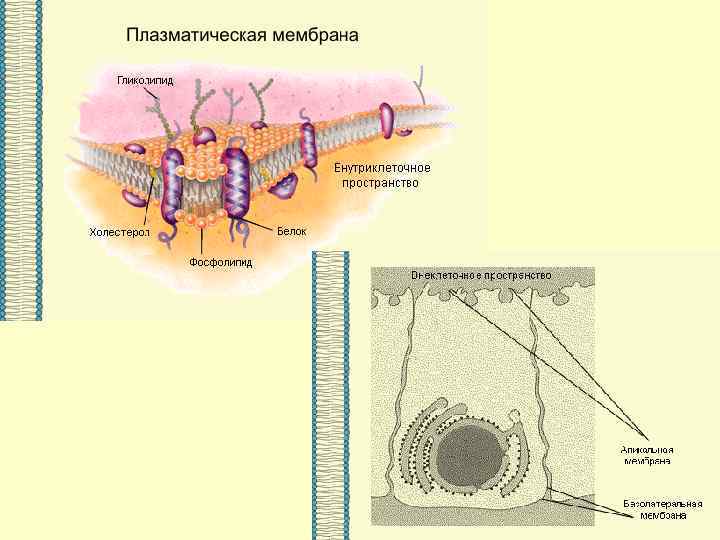
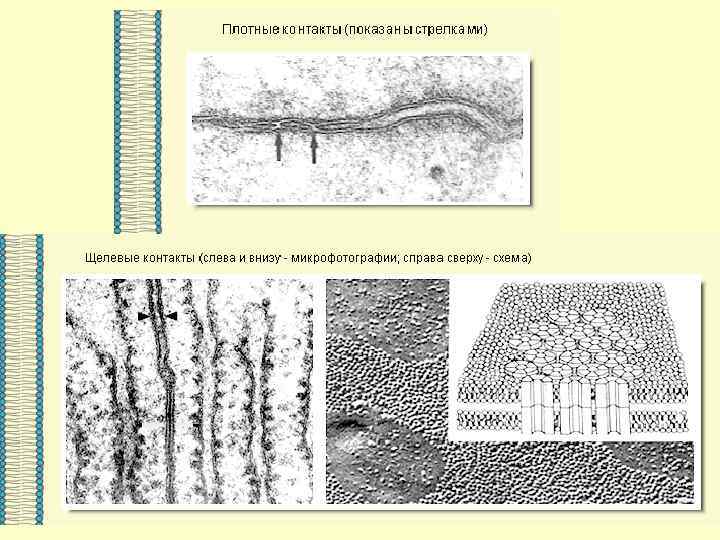
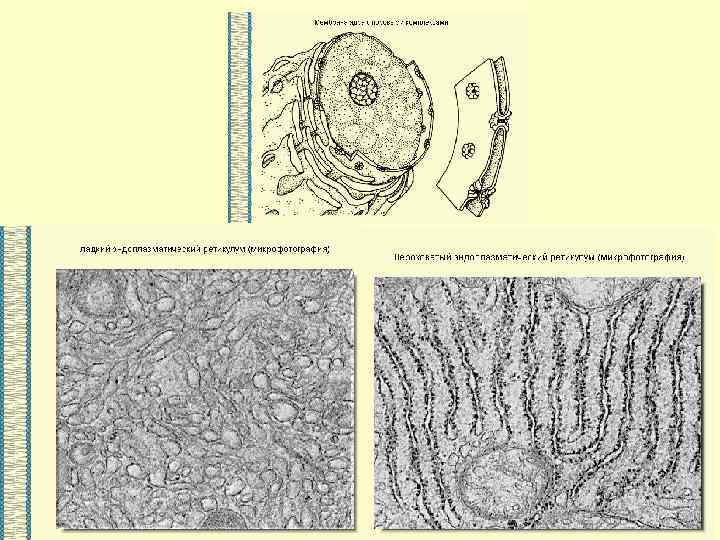
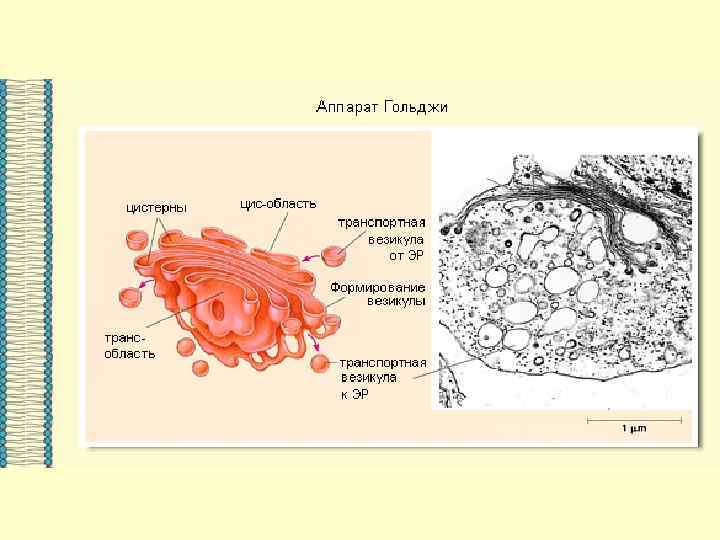
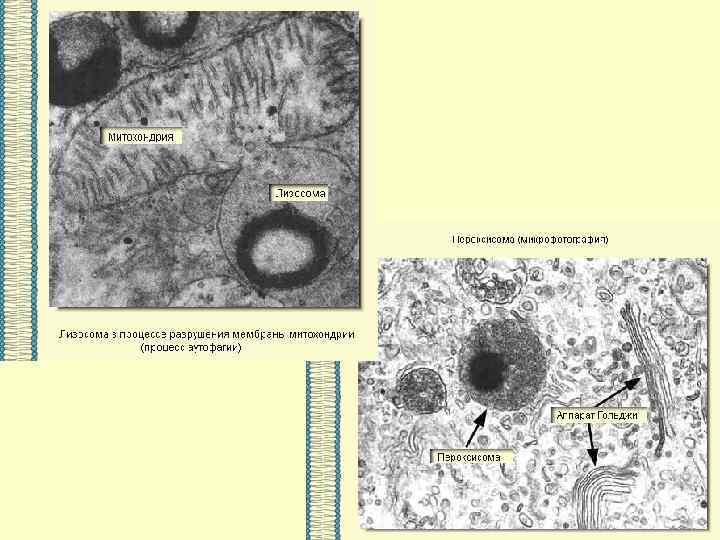
ВЕЗДЕСУЩАЯ И НЕЗАМЕНИМАЯ… МЕМБРАНА

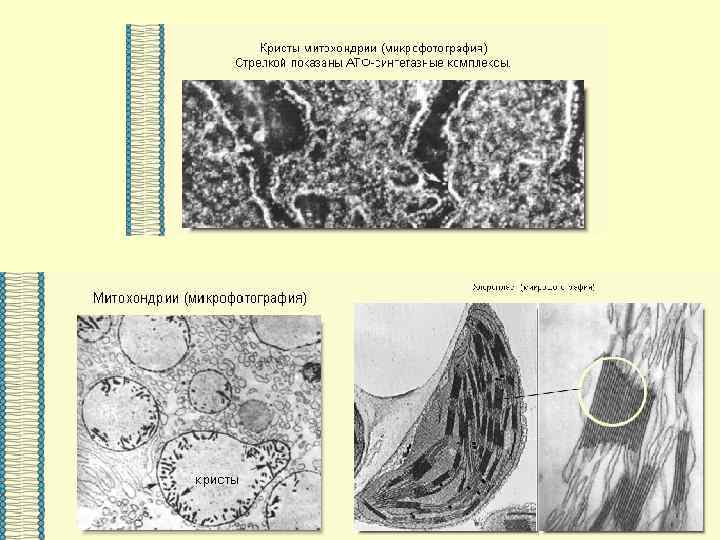
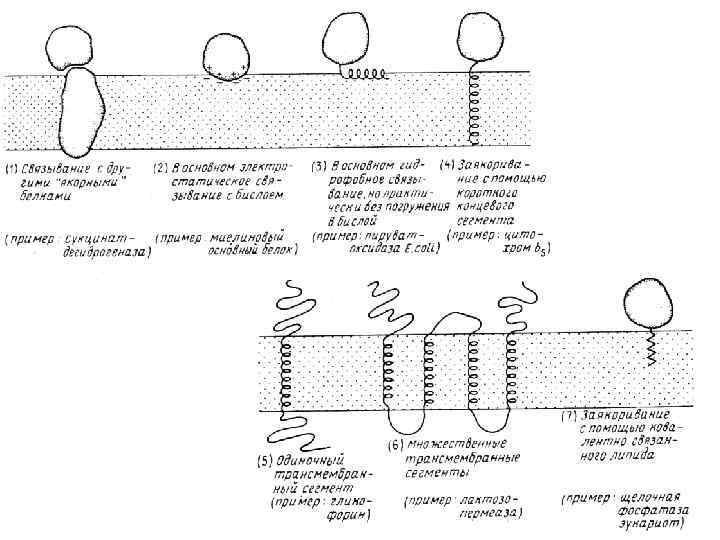
Основными компонентами мембран являются белки и липиды. На долю углеводов может приходиться около 10% массы мембран, при этом они всегда входят в состав гликолипидов или гликопротеинов. Соотношение между белками и липидами в мембранах значительно варьирует - от 20% (по массе сух. вещ-ва) белка в миелине до 80% в митохондриях. Плотность мембран прямо пропорциональна содержанию в них белка; чем выше содержание белка в мембране, тем больше ее плотность.



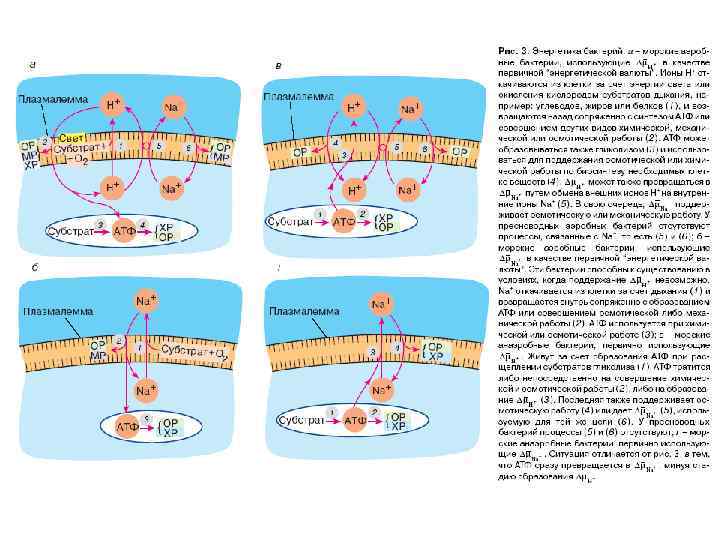
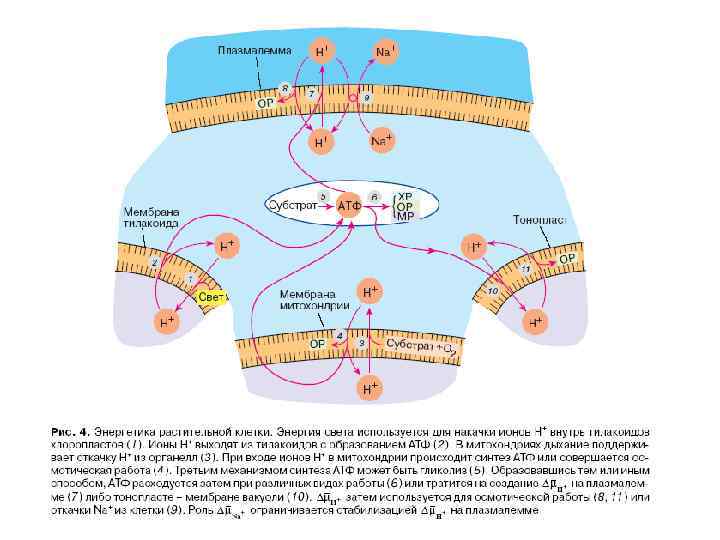
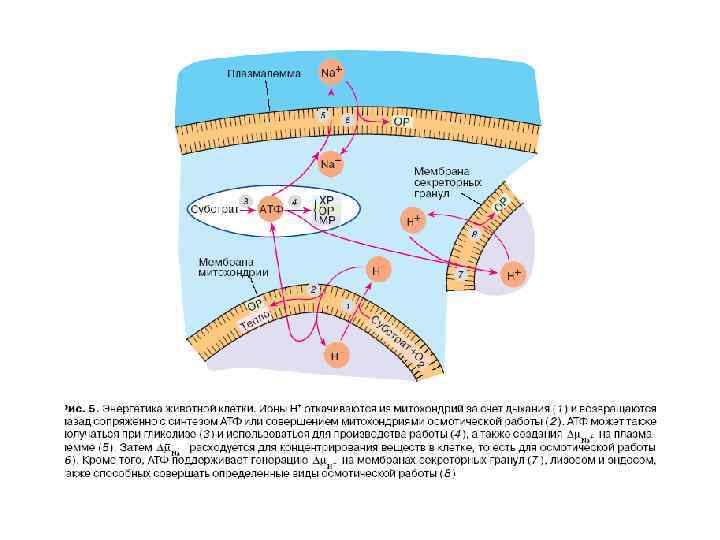
СИСТЕМЫ ПЕРВИЧНОГО АКТИВНОГО ТРАНСПОРТА

Системы первичного активного транспорта: Na/K-АТФаза

Системы первичного активного транспорта: Na/K-АТФаза

Системы первичного активного транспорта: Na/K-АТФаза

Системы первичного активного транспорта: Na/K-АТФаза

Системы первичного активного транспорта: Са-АТФаза

Системы первичного активного транспорта: Са-АТФаза

ДРУГИЕ КЛАССЫ АКТИВНЫХ ПЕРЕНОСЧИКОВ

Помимо АТР-зависимых активных переносчиков E 1 E 2 - и F 1 Fo-типов есть еще три класса активных переносчиков, использующих свободную энергию гидролиза макроэргических фосфатных связей. О реальных механизмах транспорта или сопряжения в этих системах известно немного. Бактериальные фосфотрансферазы. Транспорт сахаров и сахароспиртов у бактерий. Регуляция транспорта – фосфорилирование переносимого соединения. Источник энергии – фосфоенолпируват. Бактериальные периплазматические транспортные системы. Транспорт у бактерий аминокислот и сахаров. Регуляция – растворимый связывающий белок. Возможно также доступность АТФ. Вакуолярные Н -АТРазы эукариотов. Транспорт протонов. Механизм регуляции не установлен.

Локализация Cyt С - оксидаза Структура Функция Перенос электрона сопровождается трансмембранным переносом двух протонов из матрикса в цитозольное пространство и появлением

БАКТЕРИОРОДОПСИН

Гомологичность некоторых транспортных белков внутренней митохондриальной мембраны свидетельствует об их близком родстве: по всей вероятности, они произошли от общего предка в результате дивергентной эволюции. Имеются по меньшей мере три представителя этой группы: 1) ADP/ATP-транслоказа; 2) переносчик фосфата и 3) разобщающий белок. ADP/ATP-транслоказа катализирует транспорт ADP и АТР через бислой. При физиологических условиях АТР транспортируется из митохондрий, а ADP переносится в матрикс. Переносчик фосфата осуществляет одновременно и симпорт Н. Этот белок катализирует транспорт фосфата внутрь митохондрий. Благодаря симпорту Н, процесс в целом является электронейтральным и не зависит от трансмембранного потенциала. Разобщающий белок был обнаружен в митохондриях из клеток бурого жира млекопитающих; его функция заключается в диссипации протонного электрохимического градиента, создаваемого при функционировании дыхательной цепи, в результате чего генерируется тепло. Разобщающий белок может также катализировать транспорт анионов, например Сl. Все три переносчика, а возможно, еще и а-кетоглутарат/малат-транслоказа, имеют сходное строение; этот вывод был сделан на основе данных об их аминокислотной последовательности. Все они имеют мол. массу около 33000 Да и состоят из трех гомо-логичных доменов, каждый из которых содержит ~ 100 аминокислот. По всей вероятности, эти три домена образовались в результате утроения единственного гена. Была построена модель, согласно которой каждый из гомологичных доменов дважды пересекает мембрану, а вся субъединица содержит шесть трансмембранных а-спиралей.

АТФ-зависимый транспорт белков через мембрану

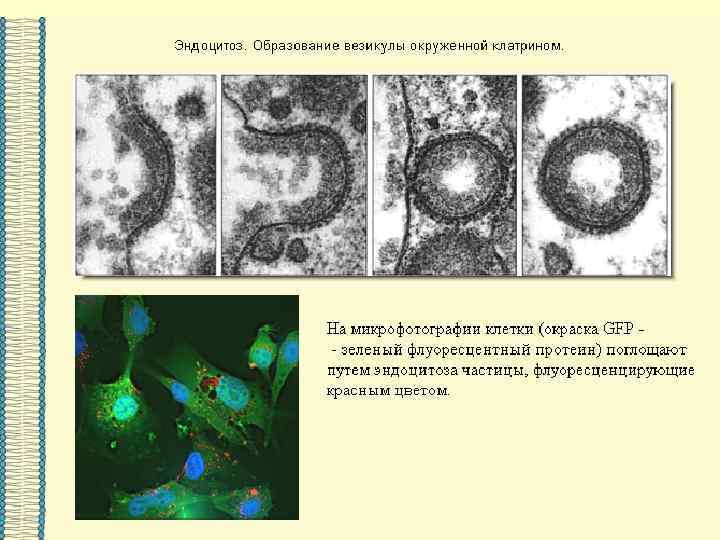
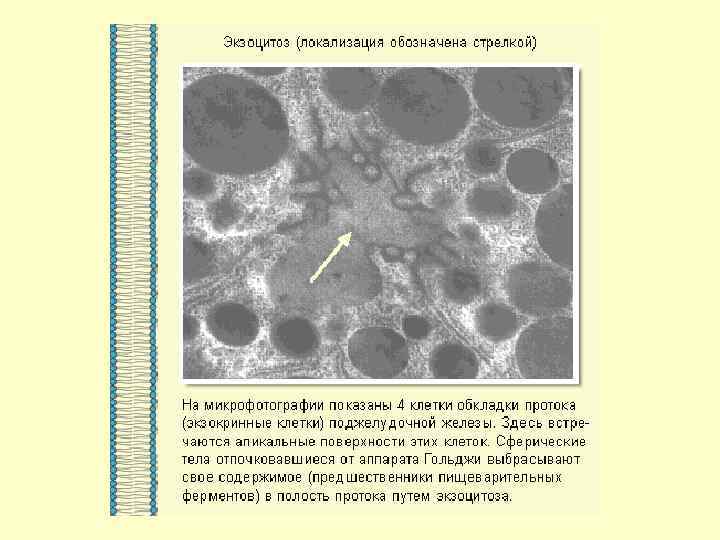
ТРАНСПОРТ ЧЕРЕЗ ПОРЫ и КАНАЛЫ


ПОТЕНЦИАЛЗАВИСИМЫЕ КАНАЛЫ Гипотетическая модель натриевого канала

Гипотетическая модель поринового тримера ПОРИНЫ

Модель мальтопорина
")
Модель Omp. F-порина. (порины семьи Omp’s)

Ионофоры

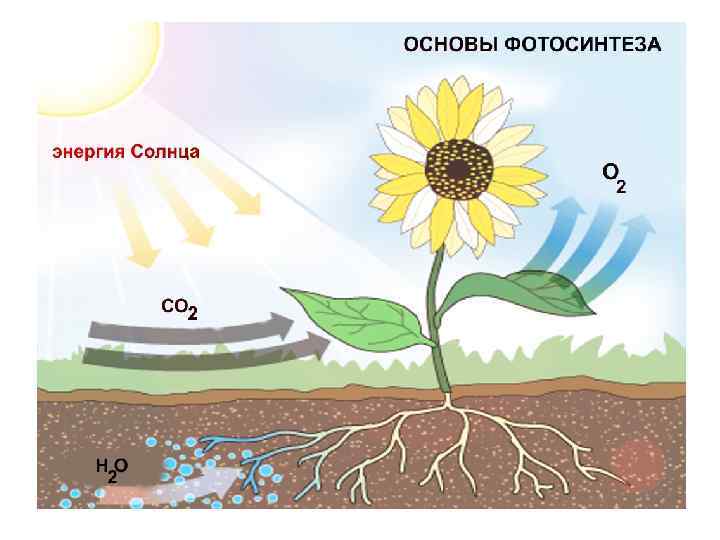

Наиболее частые заблуждения о фотосинтезе: • Все фотосинтетические организмы – выделяют кислород • Поглощение углекислого газа и выделение кислорода – связанные процессы • Результатом фотосинтеза является выделение молекулярного кислорода • Кислород берет начало из молекулы углекислого газа • Продукт фотосинтеза – глюкоза

Зеленые клетки листьев высших растений принадлежат к продуцентам кислорода. Для восстановления двуокиси углерода они используют в качестве донора водорода воду и в ходе этого процесса выделяют молекулярный кислород, согласно уравнению: Фотосинтезирующие бактерии кислорода не образуют (кроме цианобактерий). Более того, многие фотосинтезирующие бактерии являются облигатными анаэробами, т. е. вообще не переносят кислорода. В качестве доноров водорода некоторые фотосинтезирующие бактерии используют неорганические соединения. Зеленым серным бактериям, например, донором водорода служит сероводород согласно уравнению:

Общий взгляд… Фотосинтез зеленых растений протекает в 2 стадии. Первая из них объединяет световые реакции, идущие реакции только на свету, а вторая – темновые реакции, реакции которые могут идти как в темноте, так и на свету. Образование кислорода, происходящее только на свету, и восстановление двуокиси углерода, для которого свет не требуется, представляют собой два раздельных процесса.

Вглубь зеленого листа

Место действия – хлоропласт

Еще глубже - пигменты

Еще глубже - пигменты

Хлорофиллы С Хлорофилл d
 — тетрапиррольные")
Фикобилины (от греч. φύκος — водоросли и лат. bilis — желчь) — тетрапиррольные пигменты (билины) красных водорослей, криптофит и цианобактерий. Фикобилины являются хромофорной группой фикобилипротеинов — кислых водорастворимых глобулярных хромопротеинов светособирающего комплекса фотосинтетических структур водорослей. В водорослях наиболее распространены красные фикоэритрины с максимумом поглощения λmax = 560— 570 нм и синие фикоцианины с λmax = 610 нм, при этом большинство видов содержат как фикоэритрины, так и фикоцианины. Отношение количеств фикоэритринов и фикоцианинов зависят от спектрального состава света в среде обитания водорослей: преобладание зеленого света в освещении при росте на глубине вследствие поглощения водой красного участка спектра ведет к синтезу поглощающего в этом участке спектра фикоэритрина, при достаточном красном освещении у поверхности воды преобладает синтез фикоцианинов. Фитоэритробилин

Итак: У эукариотических организмов процесс фотосинтеза происходит в хлоропластах в две стадии. Световые реакции осуществляются в тилакоидах, где обнаружены молекулы хлорофиллов и других пигментов. тов Серия реакций, с помощью которых энергия переносится к углеродсодержащим соединениям, происходит в строме, которая окружает тилакоиды (темновая фаза).

Световые реакции фотосинтеза. Когда пигменты поглощают свет и электроны поднимаются на более высокий энергетический уровень, дальнейшие события могут развиваться следующим образом: энергия электрона рассеивается в виде тепла; немедленно выделяется в виде световой энергии большей длины волны (явление флуоресценции, однако когда происходит испускание света с некоторой задержкой, то данное явление называют фосфоресценцией); энергия запасается в форме химических связей, что и происходит в процессе фотосинтеза.

Световые реакции фотосинтеза

Световые реакции фотосинтеза

Темновые реакции фотосинтеза На этом этапе химическая энергия, запасенная в световых реакциях, используется для восстановления углерода. Углерод, доступный для фотосинтезирующих клеток, поставляется в виде двуокиси углерода. У большинства растений двуокись поступает к фотосинтезирующим клеткам через специальные отверстия – устьица, которые находятся в листьях. Восстановление углерода происходит в строме хлоропласта в цикле реакций, которые известны как цикл Кальвина.

Наиболее частые заблуждения о фотосинтезе: • Все фотосинтетические организмы – выделяют кислород (нет; встречаются даже облигатные анаэробы) • Поглощение углекислого газа и выделение кислорода – связанные процессы (нет; кислород – продукт фотолиза, а углекислый газ – поглощается в светонезависимой фазе) • Результатом фотосинтеза является выделение молекулярного кислорода (нет; накопление АТФ и. НАДФН и синтез глицеральдегид-3 -фосфата) • Кислород берет начало из молекулы углекислого газа (нет; кислород – продукт фотохимического расщепления воды) • Продукт фотосинтеза – глюкоза (нет; углеводородный скелет глицеральдегид-3 -фосфат)

Цикл Кальвина – не единственный путь фиксации углерода в темновых реакциях. У некоторых растений первый продукт фиксации – не трех. С молекула 3 -фосфоглицерата, а четырех. С соединение – оксалоацетат. Растения, которые осуществляют этот путь, называются соответственно С 4 -растениями (С 4 -путь называют также циклом Хэтча-Слэка в честь австралийских физиологов растений, которые сыграли решающую роль в его открытии) Пространственное разделение цикла Кальвина (т. е. фиксации углерода) и захвата двуокиси углерода
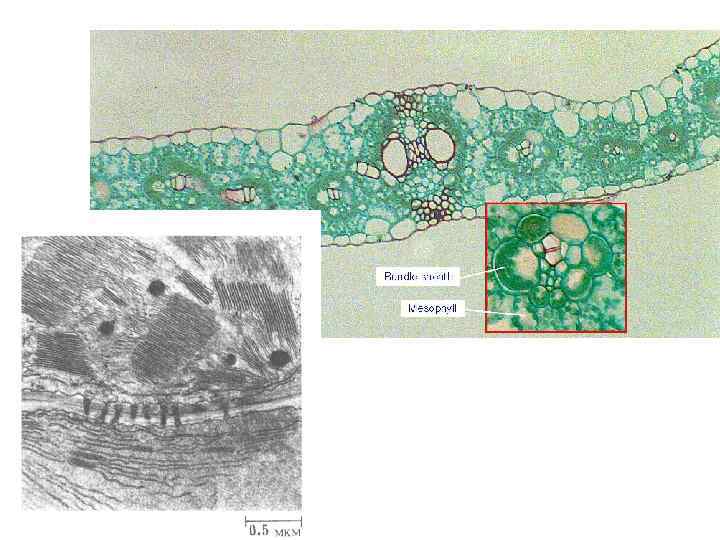

Важно! Фиксация СО 2 у С 4 растений требует более высоких энергетических затрат, чем у С В С 4 -пути на каждую ассимилированную молекулу СО 2 должна образовываться мол ФЭП при этом тратятся две высокоэнергетические фосфатные группы АТФ. С 4 -растениям нужно затратить пять молекул АТФ на фиксацию одной молекулы СО 2 а С 3 -растениям – только три. Почему у С 4 -растений в процессе эволюции возник такой сложный и энергетич дорогой способ обеспечения цикла Кальвина углекислотой? Это станет ясно, если уточнить, что фотосинтез у Сз-растений всегда сопровождается фотодыханием – «расточительным» процессом, в ходе которого на свету потребляется кислород и выделяется СО 2. Этот процесс нельзя приписать митохондриям, т. к. он лишь очен незначительно подавляется цианидом, ингибитором митохондриальной цитохромокс Важно, что по сравнению с митохондриальным дыханием фотодыхание не сопровож окислительным фосфорилированием и не дает выхода в виде АТФ. При нормальных атмосферных условиях до 50% углерода, ассимилированного при фотосинтезе у С 3 -растений, может реокисляться до СО 2 в процессе фотодыхания. Таким образом, столь активное фотодыхание у С 3 -растений ограничивает в значительной степени их эффективность, а у С 4 -растений оно практически отсутствует.

ФОТОДЫХАНИЕ: РДФ-карбоксилаза может способствовать взаимодействию РБФ либо с СО 2, либо с О 2. Когда концентрация СО 2, высокая, а кислорода - относительно низкая, то РДФ-карбоксилаза фиксирует СО 2 на РБФ с образованием фосфоглицерата. В случае низкой концентрации СО 2, и относительно высокой концентрации О 2 фермент проявляет оксигеназную активность и «сажает» О 2 на РБФ. При этом образуется фосфогликолевая кислота и фосфоглицерат вместо двух молекул фосфоглицерата, образуемых в норме при карбоксилировании. Напротив, высокая концентрация СО 2 и низкая О 2 ограничивают фотодыхание. Соответственно С 4 -растения имеют преимущество перед С 3 -растениями, поскольку СО 2 фиксируемый в С 4 -пути, перекачивается активно из клеток мезофилла в клетки обкладки тем самым поддерживая высокое отношение СО 2/О 2 в участках, где сосредоточена актив РДФ-карбоксилазы. Возможна рефиксация СО 2 при закрытых устьицах в клетках мезофи
 клетки – т. е. процессы захвата и фиксации")
САМ-метаболизм проходит в пределах одной (!) клетки – т. е. процессы захвата и фиксации СО 2 разделены здесь временными рамками

Примеры С 3 -растений: пшеница, рожь, овес, рис С 4 -растений: сахарный тростник, сорго, кукуруза САМ-растений: ананас, толстянка, каланхое, хоия, сансевьера, вельвитчия

Сорго С 4 -растения Сахарный тростник

Толстянка Ананас Вельвичия САМ-растения
Общая презентация.ppt