Презентинтмет.ppt
- Количество слайдов: 41



Интеграция метаболических путей служит обеспечению жизнедеятельности организма: производство энергии в процессе окислительного распада пищевых веществ биосинтез веществ и структур. Производство ключевых метаболитов и восстановленных коферментов, необходимых для биосинтезов.

Принципиальные составляющие интеграции метаболизма: 1. Наличие общих промежуточных продуктов в большей части метаболических путей üГлюкоза-6 -фосфат üПируват üАцетил-Ко. А
: 2. Возможность взаимопревращений через общие метаболиты")
Принципиальные составляющие интеграции метаболизма (продолжение): 2. Возможность взаимопревращений через общие метаболиты

: 3. Использование общих коферментов и необходимость их постоянной циркуляции")
Принципиальные составляющие интеграции метаболизма (продолжение): 3. Использование общих коферментов и необходимость их постоянной циркуляции НАДФН. Н+, НАД+
: 4. Наличие общего пути катаболизма и единой системы освобождения")
Принципиальные составляющие интеграции метаболизма (продолжение): 4. Наличие общего пути катаболизма и единой системы освобождения и использования энергии

: 5. Наличие сходных механизмов регуляции v. Обеспечение субстратами v.")
Принципиальные составляющие интеграции метаболизма (продолжение): 5. Наличие сходных механизмов регуляции v. Обеспечение субстратами v. Аллостерические взаимодействия v. Ковалентная модификация v. Уровень фермента v. Разделение метаболических процессов по отдельным отсекам (компартментам)
 клетки")
Разделение метаболических процессов по отдельным отсекам (компартментам) клетки

v. Органная специализация Основные особенности метаболизма в печени в состоянии после принятия пищи: Поддержание уровня глюкозы на постоянном уровне: После принятия пищи концентрация глюкозы в крови достигает 6 и выше ммоль/л, она поступает в печень и превращается там в глюкоза 6 -фосфат, так как: противоположно миоцитам и адипоцитам инсулин не оказывает здесь прямого влияния на поглощение глюкозы. интенсивность фосфорилирования глюкозы в печени пропорциональна концентрации глюкозы в крови. в глюкоза-6 -фосфат превращаются в печени и другие моносахариды, подвергшиеся всасыванию в кишечнике (фруктоза, галактоза, манноза).
: увеличенная утилизация аминокислот")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): увеличенная утилизация аминокислот для энергетических целей, усиленный гликолиз и активность пируватдегидрогеназы, увеличение синтеза гликогена для пополнения его запасов, усиление синтеза жирных кислот. Жирные кислоты используются для синтеза триацилглицеролов, фосфолипидов, которые в составе липопротеинов секретируются в кровоток
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: гликогенсинтетазы (активирует глюкоза-6 -Ф);
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: фосфорилазы (ингибирует глюкоза и глюкоза-6 -Ф).
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: фосфофруктокиназы-1 (активирует фруктоза-2, 6 ФФн);
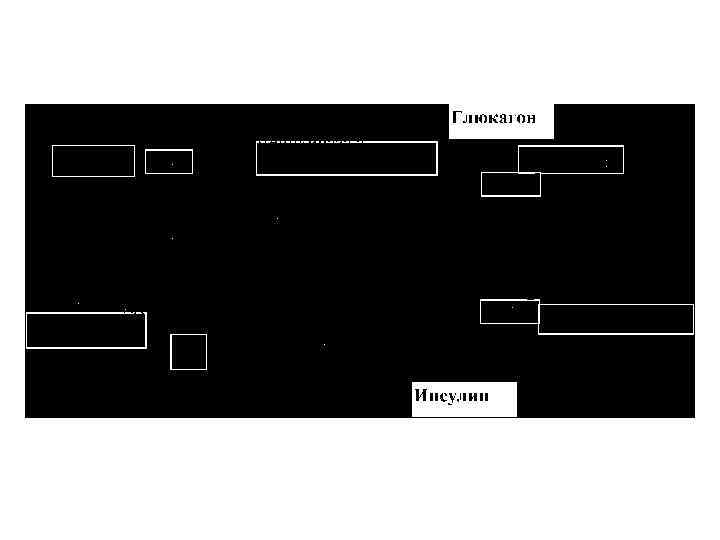
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: фруктоза-бисфосфатазы-1 (ингибирует фруктоза-2, 6 ФФн);
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: пируваткиназы (активирует фруктоза-1, 6 -ФФ);
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: пируватдегидрогеназы (активирует пируват);
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: ацетил-Ко. А карбоксилазы (активирует цитрат);
: Аллостерическая регуляция ферментов:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Аллостерическая регуляция ферментов: карнитин-ацилтрансферазы (ингибирует малонил-Ко. А);
: Ковалентную модификацию ферментов")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Ковалентную модификацию ферментов обеспечивает инсулин - дефосфорилирование белков (снижается ц. АМФ, увеличивается активность протеинфосфатазы) Увеличивается активность: фосфофруктокиназы-2
: Ковалентную модификацию ферментов")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): Ковалентную модификацию ферментов обеспечивает инсулин - дефосфорилирование белков (снижается ц. АМФ, увеличивается активность протеинфосфатазы) Увеличивается активность: пируваткиназы, пируватдегидрогеназы, ацетил-Ко. А карбоксилазы.
: инсулин индуцирует синтез:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): инсулин индуцирует синтез: üглюкокиназы, üглюкоза-6 -фосфат дегидрогеназы,
: инсулин индуцирует синтез:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): инсулин индуцирует синтез: üпируваткиназы, üцитратлиазы,
: инсулин индуцирует синтез:")
Основные особенности метаболизма в печени в состоянии после принятия пищи (продолжение): инсулин индуцирует синтез: üацетил-Ко. А карбоксилазы, üсинтетазы жирных кислот. инсулин тормозит синтез: фосфоенолпируваткарбоксикиназы глюкоза-6 -фосфатазы
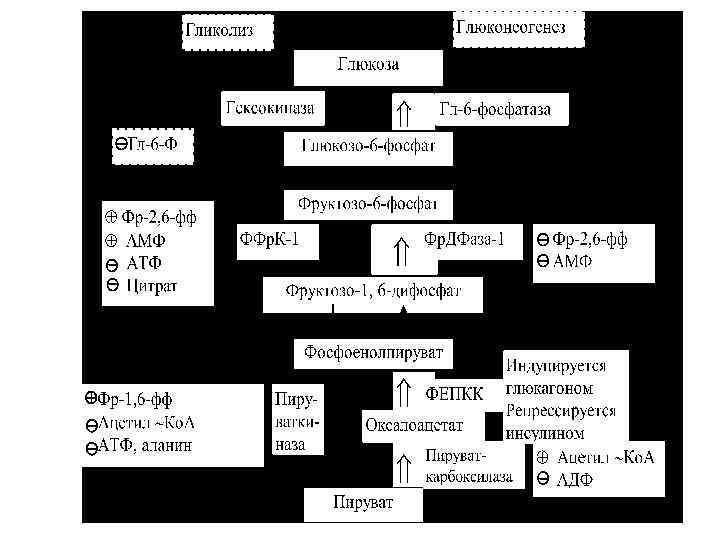

v. Органная специализация В мышцах после приема пищи: üГлюкоза используется для образования гликогена üОсновной источник энергии - жирные кислоты üАминокислоты с разветвленной цепью, подвергаясь дезаминированию, превращаются в кетокислоты, которые окисляются в цикле Кребса

v. Органная специализация после приема пищи: Для энергетических потребностей мозга интенсивно используются глюкоза и аминокислоты с разветвленной цепью. В эритроцитах глюкоза окисляется до лактата В почках основными источниками энергии выступают жирные кислоты и молочная кислота

v. Органная специализация В период натощак: • концентрация глюкозы в крови снижается до 4 ммоль/л , вследствие этого: • снижается секреция инсулина • уменьшается поглощение глюкозы всеми тканями, кроме нервной. • Низкая концентрация глюкозы в крови является сигналом для секреции глюкагона и кортизола
 Печень препятствует дальнейшему падению концентрации глюкозы в крови за счет:")
v. Органная специализация (натощак) Печень препятствует дальнейшему падению концентрации глюкозы в крови за счет: • расщепления гликогена и выхода глюкозы в кровь. • лактат, продукт анаэробного расщепления глюкозы в мышцах, поступает в печень • Спустя 12 ч после приема пищи основным источником глюкозы становится глюконеогенез из аминокислот (аланина и глутамина). Основные события в печени - активация глюконеогенеза и усиленное окисление жирных кислот!!!
 Аллостерическая регуляция: фруктозобисфосфатазы (активность усиливается за счет снятия ингибирования")
v. Органная специализация (печень, натощак) Аллостерическая регуляция: фруктозобисфосфатазы (активность усиливается за счет снятия ингибирования фруктоза-2, 6 -ФФ); фосфофруктокиназы-1 (активность снижается за счет снятия активации посредством фруктоза-2, 6 -ФФ); пируваткарбоксилазы (активируется ацетил-Ко. А); пируватдегидрогеназа (ингибируется ацетил-Ко. А); -КГ дегидрогеназы (в цикле Кребса ингибируется НАДН. Н+) ацетил-Ко. А карбоксилазы (ингибируется ацил-Ко. А).
 Ковалентная модификация (обеспечивает глюкагон, за счет увеличения уровня ц.")
v. Органная специализация (печень, натощак) Ковалентная модификация (обеспечивает глюкагон, за счет увеличения уровня ц. АМФ) Активность: фруктозобисфосфатазы-2 (усиливается), пируваткиназы (уменьшается), пируватдегидрогеназы (уменьшается); ацетил-Ко. А карбоксилазы (уменьшается); гликогенфосфорилазы (усиливается) гликогенсинтетазы (снижается).
 ц. АМФ и глюкокортикоиды индуцируют синтез: vглюкоза-6 -фосфатазы vфосфоенолпируваткарбоксикиназы.")
v. Органная специализация (печень, натощак) ц. АМФ и глюкокортикоиды индуцируют синтез: vглюкоза-6 -фосфатазы vфосфоенолпируваткарбоксикиназы.
")
v. Органная специализация (натощак)
 Плазма крови Уменьшение соотношения")
Межорганный метаболизм в динамике голодания (12 ч после приема пищи) Плазма крови Уменьшение соотношения инсулин/глюкагон; снижение уровня глюкозы; увеличение в 4 раза уровня жирных кислот Адипоциты Не извлекают глюкозу из кровотока, ускорена мобилизация ТАГ Мозг Продолжает использовать глюкозу в качестве источника энергии
 Мышцы Распад белков; высвобождение")
Межорганный метаболизм в динамике голодания (12 ч после приема пищи) Мышцы Распад белков; высвобождение большого количества глутамина и аланина; снижение использования глюкозы Почки Используют жирные кислоты и кетоновые тела в качестве предпочтительного источника энергии
 Плазма крови Инсулин/глюкагон (~")
Межорганный метаболизм в динамике голодания (3 суток после приема пищи) Плазма крови Инсулин/глюкагон (~ 10% от состояния сразу после приема пищи); увеличивается секреция кортизола; увеличен уровень жирных кислот (в 8 -10 раз); снижена концентрация глюкозы (~ 60%); увеличен уровень кетоновых тел (~ в 20 раз). Адипоциты Увеличена активность гормон-чувствительной липазы, растет высвобождение жирных кислот и глицерола Печень Гликоген отсутствует; глюконеогенез (в качестве субстратов используются аминокислоты и глицерол); синтез кетоновых тел
 Мышцы Распад белков до")
Межорганный метаболизм в динамике голодания (3 суток после приема пищи) Мышцы Распад белков до аминокислот, используемых для глюконеогенеза; глюкоза не используется в качестве источника энергии; основные источники - жирные кислоты и кетоновые тела Мозг Продолжают использовать глюкозу в качестве основного источника энергии, нарастает поступление из крови кетоновых тел Почки Используют лактат и кетоновые тела в качестве источников энергии, активирован глюконеогенез
 Плазма крови Инсулин/глюкагон (в")
Межорганный метаболизм в динамике голодания (3 недели после приема пищи) Плазма крови Инсулин/глюкагон (в 10 раз ниже, чем в состоянии сразу после приема пищи); уровень кетоновых тел увеличен ~ в 100 раз, угроза развития метаболического ацидоза. Адипоциты ТАГ расщепляются до жирных кислот Мышцы Продолжается распад белка; преимущественно высвобождается глутамин
 Печень Продолжаются глюконеогенез и")
Межорганный метаболизм в динамике голодания (3 недели после приема пищи) Печень Продолжаются глюконеогенез и увеличенный кетогенез Мозг Около 2/3 энергии черпается из окисления кетоновых тел и 1/3 - глюкозы Почки Концентрация кетоновых тел в крови превышает почечный порог реабсорбции; кетоновые тела выводятся с мочой. Метаболический ацидоз ведет к потере организмом катионов. Усиленная утилизация глутамина с целью наработки способного удаляться из организма катиона (NH 4+)
 Почки")
Межорганный метаболизм в динамике голодания (3 недели после приема пищи) Почки
Презентинтмет.ppt