

Иммунологическая Толерантность.ppt
- Количество слайдов: 18
 Иммунологическая Толерантность
Иммунологическая Толерантность
специфическая это иммунологическая неотвечаемость") Иммунологическая толерантность В переводе с английского толерантность – ( «терпимость» )специфическая это иммунологическая неотвечаемость (ареактивность), приобретенная в результате контакта с антигеном на уровне отдельных клонов лимфоцитов, тогда как способность к иммунному ответу на другие антигены сохраняется.
Иммунологическая толерантность В переводе с английского толерантность – ( «терпимость» )специфическая это иммунологическая неотвечаемость (ареактивность), приобретенная в результате контакта с антигеном на уровне отдельных клонов лимфоцитов, тогда как способность к иммунному ответу на другие антигены сохраняется.
 Иммунологическая толерантность в настоящее время представляет интерес для теоретиков и практиков: преодоление тканевой несовместимости при пересадке органов и тканей, профилактика и лечение аутоиммунных заболеваний, лечение хронических инфекционных заболеваний, формирование эффективного противоопухолевого иммунитета. Решение этих проблем позволит эффективно управлять процессами сохранения, утраты и возобновления толерантности к тканевым, микробным антигенам и аллергенам.
Иммунологическая толерантность в настоящее время представляет интерес для теоретиков и практиков: преодоление тканевой несовместимости при пересадке органов и тканей, профилактика и лечение аутоиммунных заболеваний, лечение хронических инфекционных заболеваний, формирование эффективного противоопухолевого иммунитета. Решение этих проблем позволит эффективно управлять процессами сохранения, утраты и возобновления толерантности к тканевым, микробным антигенам и аллергенам.
 Условно при изучении толерантности удобно дифференцировать два проявления этого феномена: • Толерантность к своему (“self” tolerance); • Искусственно индуцированная толерантность к чужеродному антигену (“non-self” tolerance). В основе этих двух форм проявления толерантности лежат одни и те же клеточные механизмы.
Условно при изучении толерантности удобно дифференцировать два проявления этого феномена: • Толерантность к своему (“self” tolerance); • Искусственно индуцированная толерантность к чужеродному антигену (“non-self” tolerance). В основе этих двух форм проявления толерантности лежат одни и те же клеточные механизмы.
 Впервые толерантность к клеточным антигенам была обнаружена Оуеном в 1945 г. у дизиготных телят-близнецов. В процессе эмбрионального развития у телят устанавливается общий плацентарный кровоток, что приводит к обмену клетками крови между ними. В результате каждый теленок представляет собой химеру, обладая как собственными клетками крови, так и клетками близнеца. Такие телята оставались ареактивными к аллоантигенам партнера.
Впервые толерантность к клеточным антигенам была обнаружена Оуеном в 1945 г. у дизиготных телят-близнецов. В процессе эмбрионального развития у телят устанавливается общий плацентарный кровоток, что приводит к обмену клетками крови между ними. В результате каждый теленок представляет собой химеру, обладая как собственными клетками крови, так и клетками близнеца. Такие телята оставались ареактивными к аллоантигенам партнера.
 В 50 -х годах П. Медовар демонстрировал опыты по созданию иммунологической толерантности на инбредных линиях мышей. Эмбрионам мышей линии CBA (гаплотип H -2 k ) вводили клетки селезенки мышей линии А (гаплотип Н-2 а). После рождения мышам СВА пересаживали кожный трансплантат донора А. Такой аллотрансплантат приживался. Трансплантат от любой другой линии мышей отторгался.
В 50 -х годах П. Медовар демонстрировал опыты по созданию иммунологической толерантности на инбредных линиях мышей. Эмбрионам мышей линии CBA (гаплотип H -2 k ) вводили клетки селезенки мышей линии А (гаплотип Н-2 а). После рождения мышам СВА пересаживали кожный трансплантат донора А. Такой аллотрансплантат приживался. Трансплантат от любой другой линии мышей отторгался.
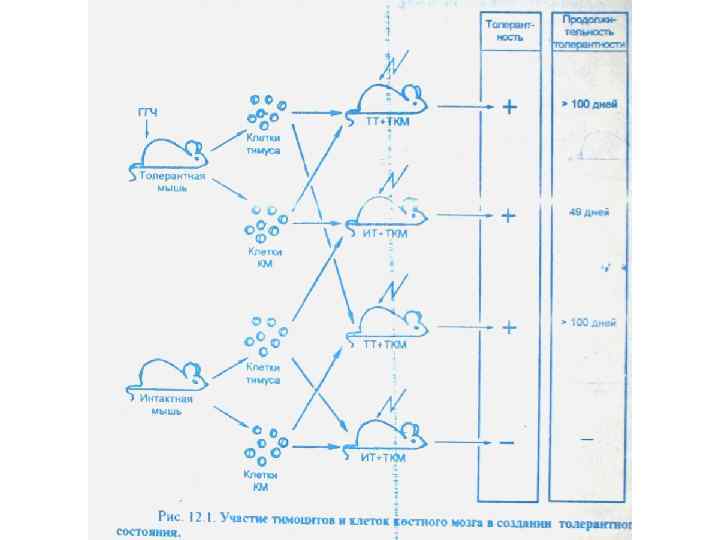
 Чешский исследователь К. Гашек индуцировал иммунологическую толерантность у кур путем трансплантации клеток донора на хориоаллантоисную мембрану эмбрионареципиента. После вылупления, цыпленок был толерантен к тканям донора. В дальнейшем было показано, что толерантность легче индуцировать в первые дни жизни введением аллогенных клеток.
Чешский исследователь К. Гашек индуцировал иммунологическую толерантность у кур путем трансплантации клеток донора на хориоаллантоисную мембрану эмбрионареципиента. После вылупления, цыпленок был толерантен к тканям донора. В дальнейшем было показано, что толерантность легче индуцировать в первые дни жизни введением аллогенных клеток.
") Толерантность можно создать и во взрослом состоянии, при условии повреждения иммунной системы (облучение, иммуносупрессия) Наилучшим условием для создания толерантности является взвесь клеток иммунной системы и кроветворных клеток. Это связано с двумя особенностями: длительному персистированию клеток в организме, их способностью к размножению (самоподдержанию) и высокой концентрацией на этих клетках МНС II класса.
Толерантность можно создать и во взрослом состоянии, при условии повреждения иммунной системы (облучение, иммуносупрессия) Наилучшим условием для создания толерантности является взвесь клеток иммунной системы и кроветворных клеток. Это связано с двумя особенностями: длительному персистированию клеток в организме, их способностью к размножению (самоподдержанию) и высокой концентрацией на этих клетках МНС II класса.
 Механизмы аутотолерантности • • Толерантность к «своему» формируется несколькими способами: Отрицательной селекцией в тимусе; Инактивацией на периферии;
Механизмы аутотолерантности • • Толерантность к «своему» формируется несколькими способами: Отрицательной селекцией в тимусе; Инактивацией на периферии;
 Механизмы центральной и периферической толерантности Клетки могут «уклониться» от негативной селекции в тимусе, но их активность подлежит контролю на периферии. У большинства здоровых индивидов аутотолерантность поддерживается с помощью ряда механизмов: секвестрации антигена, экспрессии в привилегированных участках, делеции или иммунорегуляции.
Механизмы центральной и периферической толерантности Клетки могут «уклониться» от негативной селекции в тимусе, но их активность подлежит контролю на периферии. У большинства здоровых индивидов аутотолерантность поддерживается с помощью ряда механизмов: секвестрации антигена, экспрессии в привилегированных участках, делеции или иммунорегуляции.
 Иммунологически привилегированные участки: головной мозг, передняя камера глаза, семенники, яичники, волосяные фолликулы, беременная матка. Называют их так потому, что вероятность отторжения трансплантата в этих тканях снижена. В этих участках имеется тканевой барьер и функция Т-лимфоцитов контролируется апоптозом (экспрессия Fas. L и TRAIL через Fas. R, CD 95), секрецией регуляторных цитокинов –TGFβ, ИЛ-10, а также дефицитом АПК. экспрессирующих молекулы МНС-П.
Иммунологически привилегированные участки: головной мозг, передняя камера глаза, семенники, яичники, волосяные фолликулы, беременная матка. Называют их так потому, что вероятность отторжения трансплантата в этих тканях снижена. В этих участках имеется тканевой барьер и функция Т-лимфоцитов контролируется апоптозом (экспрессия Fas. L и TRAIL через Fas. R, CD 95), секрецией регуляторных цитокинов –TGFβ, ИЛ-10, а также дефицитом АПК. экспрессирующих молекулы МНС-П.
 клеток при 2. контакте с антигеном в кортикомедуллярном") 1. Делеция клонов – гибель (апоптоз) клеток при 2. контакте с антигеном в кортикомедуллярном и мозговом слоях тимуса. Отрицательной селекции подвергаются тимоциты на стадиях CD 4+, CD 8+несущие высокоаффинный ТСR к аутоантигенам, когда они сохраняют высокую чувствительность к индукторам апоптоза (ингибируется транскрипция гена ИЛ-2). В- клетки элиминируют в результате взаимодействия со стромальными клетками костного мозга. Анергия клонов – такое состояние клеток, когда они жизнеспособны, но не могут осуществлять некоторые функции. Процесс выбраковки аутореактивных клонов продолжается на периферии. Ключевое значение в индукции анергии Т-клеток имеет отсутствие костимулирующей молекулы CD 80/86, CTLA-4.
1. Делеция клонов – гибель (апоптоз) клеток при 2. контакте с антигеном в кортикомедуллярном и мозговом слоях тимуса. Отрицательной селекции подвергаются тимоциты на стадиях CD 4+, CD 8+несущие высокоаффинный ТСR к аутоантигенам, когда они сохраняют высокую чувствительность к индукторам апоптоза (ингибируется транскрипция гена ИЛ-2). В- клетки элиминируют в результате взаимодействия со стромальными клетками костного мозга. Анергия клонов – такое состояние клеток, когда они жизнеспособны, но не могут осуществлять некоторые функции. Процесс выбраковки аутореактивных клонов продолжается на периферии. Ключевое значение в индукции анергии Т-клеток имеет отсутствие костимулирующей молекулы CD 80/86, CTLA-4.
 Роль гена Aire в иммунологической толерантности Aire стимулирует экспрессию органоспецифических генов в медуллярных эпителиальных клетках тимуса (m. TEC). Органоспецифические белки на поверхности m. TEC с помощью молекул МНС презентируются Тклеткам при их развитии в тимусе. Тимоциты, распознающие эти органоспецифические белки в контексте молекул МНС, подвергаются негативной селекции. В связи с этим роль Aire – ограничивать генерацию аутореактивных Т-клеток. Мутации в гене Aire влияют на экспрессию многих, но не всех эктопически экспрессированных белков, что указывает на существование других генов с подобной функцией.
Роль гена Aire в иммунологической толерантности Aire стимулирует экспрессию органоспецифических генов в медуллярных эпителиальных клетках тимуса (m. TEC). Органоспецифические белки на поверхности m. TEC с помощью молекул МНС презентируются Тклеткам при их развитии в тимусе. Тимоциты, распознающие эти органоспецифические белки в контексте молекул МНС, подвергаются негативной селекции. В связи с этим роль Aire – ограничивать генерацию аутореактивных Т-клеток. Мутации в гене Aire влияют на экспрессию многих, но не всех эктопически экспрессированных белков, что указывает на существование других генов с подобной функцией.
. В") 3. Супрессия клонов или антигенспецифическое подавление активации в результате действия вето-клеток (Т -супрессоров). В частности, супрессия клонов может осуществляться регуляторными цитокинами(ТФР-β, ИЛ– 10) 4. Оральная толерантность – индуцируется в печени 5. пищевыми антигенами. Значительная роль в запуске апоптоза отводится NK-клеткам печени, регуляторным Тклетками в лимфоидной ткани слизистых оболочек через ингибирующие цитокины (ТФР-β, PGE 2, ИЛ-10) Подавление инициации иммунного ответа неспецифическими клетками супрессорами разнообразного происхождения (в иммунопривилегированных местах – глиальные клетки сетчатки, децидуальные клетки эндометрия матки и др. )
3. Супрессия клонов или антигенспецифическое подавление активации в результате действия вето-клеток (Т -супрессоров). В частности, супрессия клонов может осуществляться регуляторными цитокинами(ТФР-β, ИЛ– 10) 4. Оральная толерантность – индуцируется в печени 5. пищевыми антигенами. Значительная роль в запуске апоптоза отводится NK-клеткам печени, регуляторным Тклетками в лимфоидной ткани слизистых оболочек через ингибирующие цитокины (ТФР-β, PGE 2, ИЛ-10) Подавление инициации иммунного ответа неспецифическими клетками супрессорами разнообразного происхождения (в иммунопривилегированных местах – глиальные клетки сетчатки, децидуальные клетки эндометрия матки и др. )
 CD 4+ регуляторные Т-клетки Тимус генерирует CD 25 - и CD 25+ регуляторные Т-клетки (Treg). Т-клетки отвечают на антиген (Ag), который презентируют зрелые DC. Затем Т-клетки дифференцируются в Т-клетки-эффекторы (Tэ), секретируя цитокины и оказывая «помощь» цитотоксическим Т-клеткам и Вклеткам. Антиген, презентируемый незрелыми DC, стимулирует дифференцировку клеток CD 25 -, в частности IL-10+Treg, которые супрессируют образование Tэ. CD 25+Treg тимусного происхождения блокируют экспансию популяции Tэ. CD 25+Fox. P 3+Treg могут также генерироваться из CD 25 - -прекурсоров в периферической лимфоидной ткани. Супрессивное действие разных популяций регуляторных клеток in vivo зависит от продукции цитокинов, таких как IL-10 и TGFβ, однако необходимость присутствия цитокинов зависит от природы ответа эффекторных Т-клеток.
CD 4+ регуляторные Т-клетки Тимус генерирует CD 25 - и CD 25+ регуляторные Т-клетки (Treg). Т-клетки отвечают на антиген (Ag), который презентируют зрелые DC. Затем Т-клетки дифференцируются в Т-клетки-эффекторы (Tэ), секретируя цитокины и оказывая «помощь» цитотоксическим Т-клеткам и Вклеткам. Антиген, презентируемый незрелыми DC, стимулирует дифференцировку клеток CD 25 -, в частности IL-10+Treg, которые супрессируют образование Tэ. CD 25+Treg тимусного происхождения блокируют экспансию популяции Tэ. CD 25+Fox. P 3+Treg могут также генерироваться из CD 25 - -прекурсоров в периферической лимфоидной ткани. Супрессивное действие разных популяций регуляторных клеток in vivo зависит от продукции цитокинов, таких как IL-10 и TGFβ, однако необходимость присутствия цитокинов зависит от природы ответа эффекторных Т-клеток.
 - это особая форма толерантности В-клеток, обусловленная дисбалансом между") Терминальная дифференцировка (исчерпание клона клеток) - это особая форма толерантности В-клеток, обусловленная дисбалансом между дифференцировкой и пролиферацией. ИЛ-6 при определенных условиях вызывает превращение В-лимфоцитов в плазматическую клетку, минуя стадию пролиферации, поскольку зрелая АОК имеет ограниченную продолжительность жизни, то клон В-лимфоцитов быстро истощается.
Терминальная дифференцировка (исчерпание клона клеток) - это особая форма толерантности В-клеток, обусловленная дисбалансом между дифференцировкой и пролиферацией. ИЛ-6 при определенных условиях вызывает превращение В-лимфоцитов в плазматическую клетку, минуя стадию пролиферации, поскольку зрелая АОК имеет ограниченную продолжительность жизни, то клон В-лимфоцитов быстро истощается.
 Отмена толерантности • Удаления из организма антигена • Иммунизация антигеном перекрестно реагирующим с антигеном, использованным для индукции толерантности. • Введение сингенных лимфоцитов от нетолерантных животных. • Воздействием, повреждающим лимфоциты с последующим восстановлением лимфоидной системы.
Отмена толерантности • Удаления из организма антигена • Иммунизация антигеном перекрестно реагирующим с антигеном, использованным для индукции толерантности. • Введение сингенных лимфоцитов от нетолерантных животных. • Воздействием, повреждающим лимфоциты с последующим восстановлением лимфоидной системы.