Лекция_15_17_дыхание_2013.ppt
- Количество слайдов: 74



«И дышит сад легко, глубоко, всей грудью яблонь, груш и слив» Вс. Рождественский ДЫХАНИЕ РАСТЕНИЙ «. . . Дыхание служит стимулом главнейших жизненных отправлений растений. . . подобно животным, они поглощают при переработке органических соединений в органические образования кислород, выделяют углекислоту и освобождают тепловую энергию. . . » А. С. Фаминцын

План лекции 1. Энергетический цикл жизни. История развития учения о дыхании. 2. Дыхание и его значение в жизни растений. 3. АТФ: структура и функции. 4. Строение и физиологические особенности митохондрий. 5. Величины, определяющие дыхание: интенсивность дыхания, дыхательный коэффициент. 6. Влияние внешних условий и внутренних факторов на дыхание. 7. Схема дыхания по В. И. Палладину 8. Особенности химизма и энергетики дыхания в сравнении с горением. 9. Ферменты дыхания. Пути дыхательного обмена растений. 10. Гликолитический (дихотомический) путь дыхательного обмена. 11. Генетическая связь дыхания и брожения. 12. Пентозофосфатный (апотомический) путь дыхательного обмена. 13. Глиоксилатный цикл. 14. Альтернативный путь дыхания

1. Энергетический цикл жизни С 6 Н 12 О 6 + 6 О 2 → 6 СО 2 + 6 Н 2 О + 2824 к. Дж

История развития учения о дыхании Дыхание было обнаружено более 200 лет назад (1771 -1779 гг. ) вместе с фотосинтезом как процесс «порчения воздуха» Пристли Джозеф (1733 -1804) Шееле Карл Вильгельм (1742 – 1786)
 А. Л. Лавуазье (1780) считал, что растения поглощают кислород для")
Антуан Лавуазье (1743 -1794) А. Л. Лавуазье (1780) считал, что растения поглощают кислород для сжигания пищи. При дыхании, как и при горении поглощается кислород и образуется СО 2, причем в обоих случаях выделяется теплота. Дыхание - это медленно текущее горение питательных веществ в живом организме. Лавуазье изображен во время проведения эксперимента по определению состава воды путем поджигания смеси водорода и кислорода электрической искрой (гравюра XIX в. ). УСТАНОВКА А. ЛАВУАЗЬЕ для определения состава воздуха.
, что растения исправляют воздух на свету и только зелеными")
Показал (1778 -80 гг. ), что растения исправляют воздух на свету и только зелеными частями, а незеленые части на свету и в темноте, зеленые в темноте – портят. «Когда солнце, поднявшееся над горизонтом, разбудит своими лучами заснувшие за ночь растения, оно сделает их способными исполнять свою целительную функцию – исправлять воздух для животных; во мраке ночи эта деятельность совсем прекращается; днем же совершается с тем большим оживлением, чем светлее день и чем выгоднее расположено растение в отношении солнечных лучей. Затененные высокими зданиями или другими растениями, они не исправляют воздух, а, наоборот, выделяют вредный для дыхания животных воздух. К концу дня выработка очищенного воздуха ослабевает и при заходе солнца совершенно прекращается» Ян Ингенхауз (1727 -1806) Основатель учения (1797 -1804 гг. ). Ввел термин, уравнение, установил соотношение СО 2/О 2, которое как правило равно 1. Этот газообмен и есть дыхание. При этом образуется вода. Николя-Теодор де Соссюр (1767 -1845)
 в 1838 г. утверждал, что растению не свойственно дыхание, как")
Ф. Ю. Мейен (Германия) в 1838 г. утверждал, что растению не свойственно дыхание, как у животных. Л. Гарро (Франция) – в 1850 -1851 гг. установил, что дыхание и фотосинтез – разные процессы. Ж. Б. Буссенго в 1848 г. предложил уравнение дыхания. Жан Батист Буссенго (1802– 1887)

Отто Варбург НОБЕЛЕВСКАЯ ПРЕМИЯ 1933 г. за результаты исследований связи кислорода и рака Ханс Адольф Кребс (1900 -1981) английский биохимик, НОБЕЛЕВСКАЯ ПРЕМИЯ 1953 г. (совместно с Ф. Липманом) по физиологии и медицине за открытие цикла трикарбоновых кислот. Альберт Сент-Дьёрдьи американский биохимик, НОБЕЛЕВСКАЯ ПРЕМИЯ 1937 г. по физиологии и медицине за работы по биологическому окислению.
 Автор ферментативной теории дыхания растений Бах")
Вклад русских ученых Палладин Владимир Иванович (1859 -1922) Автор ферментативной теории дыхания растений Бах Алексей Николаевич (1857 -1946) Автор перекисной теории медленного окисления Бородин Иван Парфеньевич (1847 -1930) В 1876 году защитил диссертацию «Физиологические исследования над дыханием листовых побегов» и получил степень магистра ботаники Костычев Сергей Павлович (1877 -1931) экспериментально обосновал теорию о генетической связи дыхания и брожения

2. Дыхание и его значение в жизни растений Дыхание – это процесс распада сложных органических соединений до СО 2 и воды при участии кислорода воздуха, сопровождаемый выделением энергии. С 6 Н 12 О 6+6 О 2→ 6 СО 2 + 6 Н 2 О + 2824 к. Дж

Значение дыхания 1. Энергетическое. Энергия, выделяющаяся при дыхании, используется эндорганические процессы: - синтезы, - поступление и транспорт веществ, - рост Ж. Б. Ламарк (1744 — 1829) - поддержание структур - терморегуляция. Дыхание → протонный потенциал → АТФ 2. Конструкционное 3. Образование воды
 Нобелевская премия по физиологии")
3. АТФ: структура и функции Липман Фриц Альберт (1899– 1986) Нобелевская премия по физиологии и медицине, 1953 г. АДФ + Фн → АТФ + Н 2 О

Энергия гидролиза фосфатов Вещество ΔG, к. Дж/моль ФЕП 1, 3 -Би. ФГК Креатинфосфат АТФ АДФ АМФ Глюкозо-6 -фосфат -62, 1 -49, 5 -43, 3 -30, 6 -13, 8

свет Орг. субстрат Биологические преобразователи энергии, ОВ цепи, связанные с мембранами Движение у прокариот Протонный потенциал Передача Е на расстояния Термогенез Активный транспорт АТФ Движение (механ. работа) Активный транспорт Передача Е на расстояния Биосинтезы

4. Строение и физиологические особенности митохондрий Диаметр 0, 5 - 1, 0 мкм длина 1 - 3 мкм Функции: - окислительное фосфорилирование -биосинтез соединений (некоторые аминокислоты) -протекание метаболических процессов

Геном растительной клетки

5. Величины, определяющие дыхание. Интенсивность дыхания. О 2 мм 3/ч 1 г сыр. массы Ананас 30 Аронник до 30000 Морковь, корни 25 -30 Морковь, мол. лист. 1100 Фасоль, растение 97 Фасоль, лист 260 -570 Элодея, побег 900 Яблоня, плод 20 -35 Ячмень, лист 250 Ячмень, семена 0, 06 Северные >южных Теневыносливые<светолюбивые высокогорные >равнинных корни <листья <цветки пестик > лепестки >листьев молодые >старые Камбий, флоэма >ксилема

Дыхательный коэффициент ДК = VСО 2 / VО 2 Органические вещества, разрушающиеся при дыхании – субстрат С 6 Н 12 О 6 +6 О 2 → 6 СО 2 + 6 Н 2 О 6 СО 2 / 6 О 2 =1 2 С 2 Н 2 О 4 + О 2 → 4 СО 2 + 2 Н 2 О 4 О 2 / 1 О 2 = 4 С 18 Н 36 О 2 + 26 О 2 → 18 СО 2 + 18 Н 2 О 8 СО 2/26 О 2= 0, 7 Факторы влияющие на ДК: 1. Качество дыхательного материала (степень его окисленности) 2. Концентрация кислорода 3. Расположение ткани 4. Повреждение мембран

6. Влияние внешних и внутренних факторов на интенсивность дыхания Внешние условия С 6 Н 12 О 6+6 О 2→ 6 СО 2 + 6 Н 2 О + 2824 к. Дж 1. Температура 2. Содержание кислорода 3. Содержание углекислого газа 4. Вода 5. Свет 6. Питательные соли 7. Поранение Внутренние факторы 1. Экологическая группа 2. Возраст 3. Фаза роста
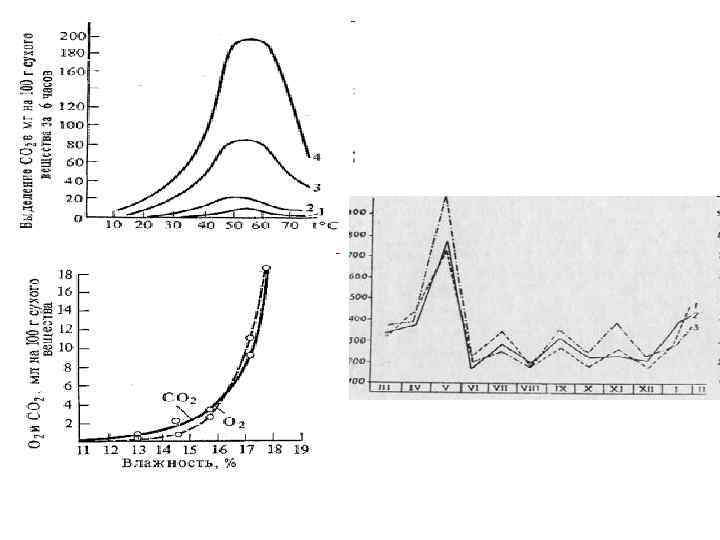

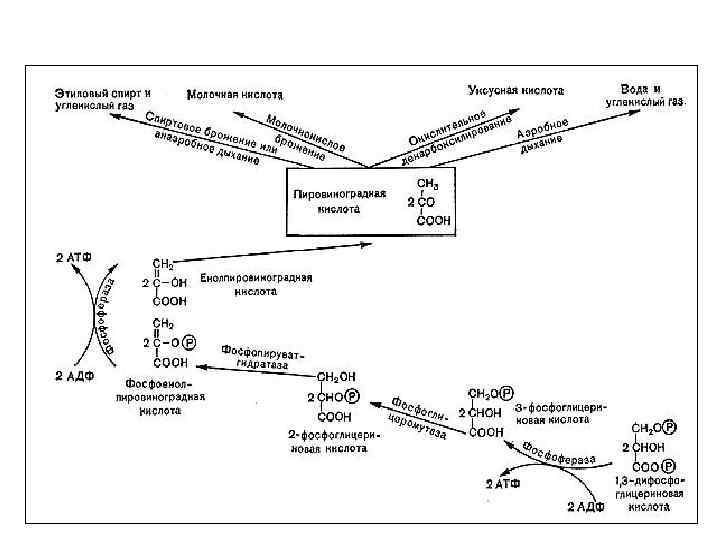
7. Схема дыхания по В. И. Палладину 1 ст. - анаэробный С 6 Н 12 О 6 + 6 Н 2 О + 12 R→ 6 СО 2+12 RН 2 2 ст. - аэробный 12 RН 2 + 6 О 2 → 12 R + 12 Н 2 О С 6 Н 12 О 6 + 6 О 2 → 6 СО 2 + 6 Н 2 О R – дыхательный пигмент (окраш. ) RН 2 – дыхательный хромоген (бесцвет)

8. Сравнение дыхания и горения Горение Дыхание Протекают по одному уравнению, выделяется одинаковое количество энергии Энергия выделяется сразу в большом количестве. Энергия выделяется в виде тепла и света Энергия выделяется в процессе присоединения кислорода. Энергия выделяется постепенно, многоступенчатый процесс. Большая часть энергии, заключенной в глюкозе, переходит в АТФ без перехода в тепло и используется организмом. Энергия выделяется в процессе отнятия водорода. Ферментативный процесс

9. Ферменты дыхания Оксидоредуктазы, участвующие в дыхании, делятся на следующие основные группы: • Анаэробные дегидрогеназы передают электроны промежуточным акцепторам, но не О 2. • Аэробные дегидрогеназы передают электроны различным акцепторам, в т. ч. и О 2. • Оксидазы передают электроны О 2.
. • • двухкомпонентные ферменты коферментом является НАД или НАДФ, которые имеют")
Анаэробные дегидрогеназы (пиридиновые). • • двухкомпонентные ферменты коферментом является НАД или НАДФ, которые имеют сходную структуру и после гидролиза образуют рибозу, аденин, никотинамид и фосфорную кислоту • НАД и НАДФ связаны с ферментом с помощью ионов металла и сульфгидрильных группировок • отнимают от субстрата 2 Н+ и 2 ē • Присоединяют 1 Н+ и 2 ē, 1 Н+ выделяется в среду (НАДН+Н+) • при окислении субстрата превращаются в восстановленную форму и легко отрываются от белковой части • в восстановленной форме находятся в цитоплазме в свободном состоянии, могут транспортироваться, являются универсальными восстановителями • в окисленной форме связаны с мембранами, являются универсальными окислителями. • различают более 150 пиридиновых дегидрогеназ: НАД (алкогольдегидрогеназа, лактатдегидрогеназа, малатдегидрогеназа); НАДФ (глюкозо-6 -фосфатдегидрогеназа, фосфоглюконатдегидрогеназа, изоцитратдегидрогеназа, )
 • • • двухкомпонентные ферменты простетическая группа прочно прикреплена к белковому")
Аэробные дегидрогеназы (флавиновые) • • • двухкомпонентные ферменты простетическая группа прочно прикреплена к белковому носителю простетической группой являются производные витамина В 2 (рибофлавин) — флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД) активной частью служит изоаллоксазиновое кольцо отнимают от субстрата два Н+ и два ē присоединяют 2 Н+и 2ē специфичность определяется белковой частью фермента содержат металлы: Fe, Cu, Mo, Mn доноры ē – анаэробные дегидрогеназы или субстрат акцепторы ē – хиноны, цитохромы, кислород Пример - сукцинатдегидрогеназа (ФАД). ФАД(ФМН) → ФАДН 2 (ФМН Н 2) окрашены (ФМН-желтый дых. фермент Варбурга) бесцветны

Оксидазы • • аэробные дегидрогеназы, передают электроны только на кислород при этом образуются вода (перенос 4ē), Н 2 О 2 -пероксид водорода (2ē) или О 2 -- супероксидный анион кислорода (ē) Примеры: цитохромоксидаза, полифенолоксидаза (вода), флавопротеиновые оксидазы (Н 2 О 2), ксантиноксидаза (О 2 -) • Fe-протеиды: геминовые: каталаза, пероксидаза, цитохромы; негеминовые: аконитаза, ксантиноксидаза • Cu-протеиды (полифенолоксидаза и аскорбатоксидаза) • Цитохромы: известно около 20 цитохромов (а, b, с, d, отличающихся между собой природой простетической группы): Fe 3+ ± ē ↔ Fe 2+ • цит. b→ цит. с1 → цит. с→ цит. аа 3→О 2
; Пентозофосфатный (апотомический).")
Пути дыхательного обмена растений Гликолитический (дихотомический); Пентозофосфатный (апотомический).
, 2 фаза — аэробная Локализованы в")
10. Гликолитический путь 1 фаза — анаэробная (гликолиз), 2 фаза — аэробная Локализованы в различных компартментах клетки: • анаэробная фаза (гликолиз) — в цитоплазме, • аэробная фаза — в митохондриях.
 Путь Г. Эмбдена – О. Мейергофа")
Гликолиз - (glycol – сахар, lysis – расщепление) Путь Г. Эмбдена – О. Мейергофа – Я. О. Парнаса С. П. Костычев, Л. А. Иванов, А. Гарден глюкоза → 2 ПВК (аэробный) глюкоза → 2 молочной кислоты (анаэробный) С 6 Н 12 О 6 → 2 С 3 Н 4 О 2 + 2 Н 2 O

Характеристика гликолиза • Универсальный, древний процесс. • Общий начальный этап дыхания и брожения. • С биологической точки зрения процесс достаточно примитивный, менее саморегулируемый. • Все реакции гликолиза протекают в цитозоле. • Включает 10 последовательных реакций, каждая из которых катализируется специфическим ферментом.
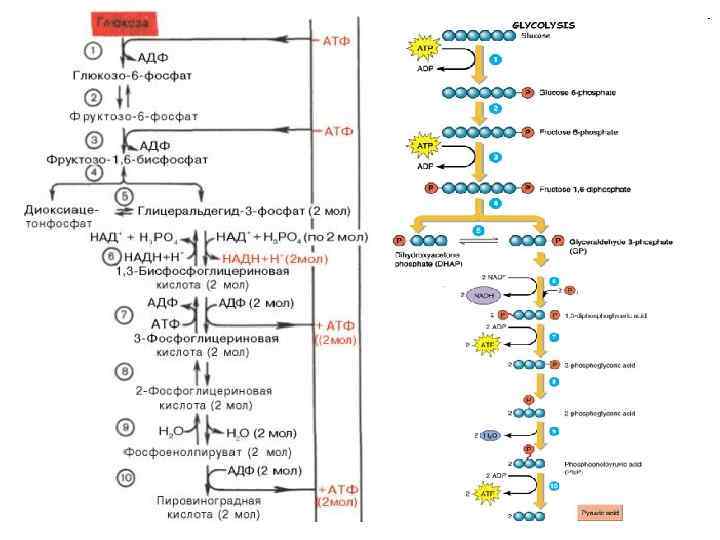
 АТФ С 6 Р 1 АТФ С 6 Р 2 С")
Глюкоза (С 6) АТФ С 6 Р 1 АТФ С 6 Р 2 С 3 Р 1 2 АТФ НАДН С 3 - ПВК

Значение гликолиза Энергетическое - образуется 2 молекулы АТФ и 2 молекулы НАДН+Н+ Метаболическое - образование промежуточных веществ, необходимых для синтетических процессов в клетке. Гликолиз в хлоропластах обеспечивает синтез АТФ, метаболизацию крахмала до триоз, которые экспортируются из хлоропласта. Первый подготовительный этап дыхания у аэробов.
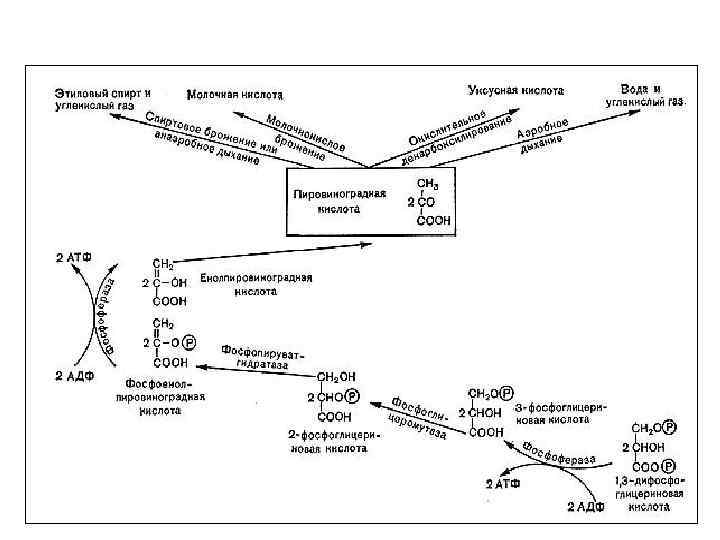

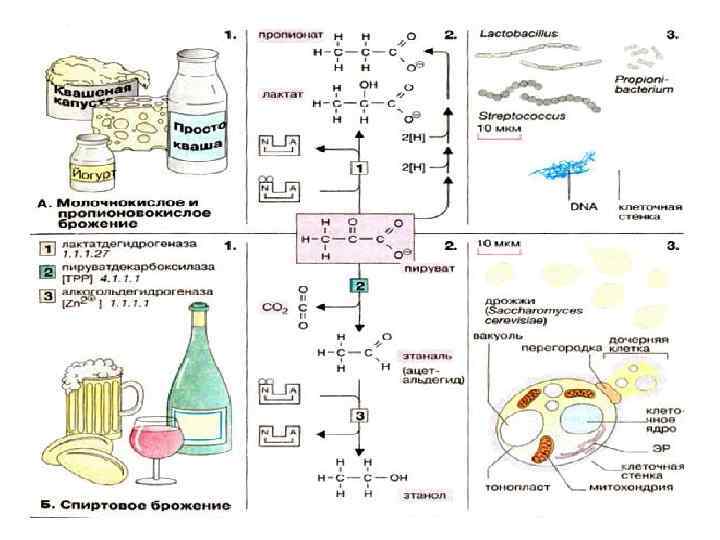
11. Генетическая связь дыхания и брожения «Дыхание и брожение – это два русла, по которым энергия изливается во все живые организмы» Энгельгард Л. Пастер обнаружил, что в растительных тканях наряду с выделением СО 2 идет спиртовое брожение Э. Ф. Пфлюгер показал наличие у лягушек интрамолекулярного дыхания (выделение CО 2 в бескислородной среде) и предположил, что оно является начальным этапом аэробного дыхания. Б. Пфефер и Э. Ф. Пфлюгер предложили уравнения механизма дыхания: С 6 Н 12 О 6 → 2 С 2 Н 5 ОН +2 СО 2 2 С 2 Н 5 ОН +6 О 2→ 4 СО 2 + 6 Н 2 О С 6 Н 12 О 6+6 О 2→ 6 СО 2 + 6 Н 2 О +О 2 глюкоза брожение дыхание

С. П. Костычев выдвинул положение о генетической связи процесов брожения и дыхания. При этом он опирался на следующие факты: 1. У высших растений найден весь набор ферментов, который катализирует отдельные этапы процесса брожения. 2. При временном попадании в условия анаэробиоза высшие растения определенное время существуют за счет энергии, выделяющейся в процессе брожения. 3. При добавлении к клеткам факультативных анаэробов (дрожжи) полусброженных сахаров интенсивность дыхания у них резко возрастает, следовательно, полусброженные продукты являются лучшим субстратом дыхания по сравнению с сахарами. +О 2 2 дыхание глюкоза Промежуточный продукт брожение -О 2

Сравнение дыхания и брожения Дыхание Брожение ферментативный распад органических соединений, сопровождаемый выделением энергии на простые неорганические на более простые органические аэробный анаэробный процесс окислительный степень окисленности не меняется акцептор электрона — кислород Больше энергии акцепторы электрона— орг. соединения Меньше энергии

Эффект Пастера – торможение брожения в присутствие кислорода при одновременном сокращении распада глюкозы.

Аэробная фаза 2 ПВК + 5 О 2 + 6 Н 2 О → 6 СО 2 + 5 Н 2 О Основные стадии: 1) окислительное декарбоксилирование пировиноградной кислоты; 2) цикл трикарбоновых кислот (цикл Кребса); 3) электронтранспортная цепь (ЭТЦ) Первые две стадии происходят в матриксе митохондрий, ЭТЦ локализована на внутренней мембране митохондрий.

Окислительное декарбоксилирование ПВК СН 3 СОСООН + НАД + Ко. А – SH → СН 3 СО– S– Ко. А + НАДН+H+ + СО 2 Мультиферментный комплекс пируватдекарбоксилаза (Мr 4, 0· 106) включает три фермента и пять коферментов (тиаминпирофосфат, липоевая кислота, коэнзим А – Ко. А- SH, ФАД и НАД)

Цикл Кребса Цикл трикарбоновых кислот • 1910 г. швед. химик Т. Тунберг показал, что в животных клетках содержатся ферменты, способные отнимать Н+ от некоторых органических кислот. • 1935 г. венгер. ученый А. Сент-Дьердьи установил, что добавление небольших количеств органических кислот (фумаровой, яблочной или янтарной) усиливает поглощение кислорода измельченными мышечными тканями. • 1937 г. англ. биохимик Г. Кребс (университет Шеффилда, Великобритания), пришел к выводу, что главным путем окисления углеводов являются циклические реакции, в которых происходит постепенное преобразование ряда органических кислот. Эти преобразования были названы циклом трикарбоновых кислот или циклом Кребса. В 1953 году Кребс был удостоен Нобелевской премии. • 1939 г. англ. Чибнелл доказал существование цикла у растений.

«Цикл Кребса объясняет два одновременно происходящих процесса: реакции распада, при которых высвобождается энергия, и синтетические процессы, при которых эта энергия расходуется» Эрик Хаммарстен Ханс Адольф Кребс (25. 08. 1900– 22. 11. 1981) «Наличие одного и того же механизма образования энергии у всех живых существ позволяет сделать еще два вывода. Во-первых, этот механизм возник на очень ранних этапах эволюции, и, во-вторых, жизнь в ее настоящем виде зародилась лишь однажды» . Х. Кребс, Нобелевская лекция
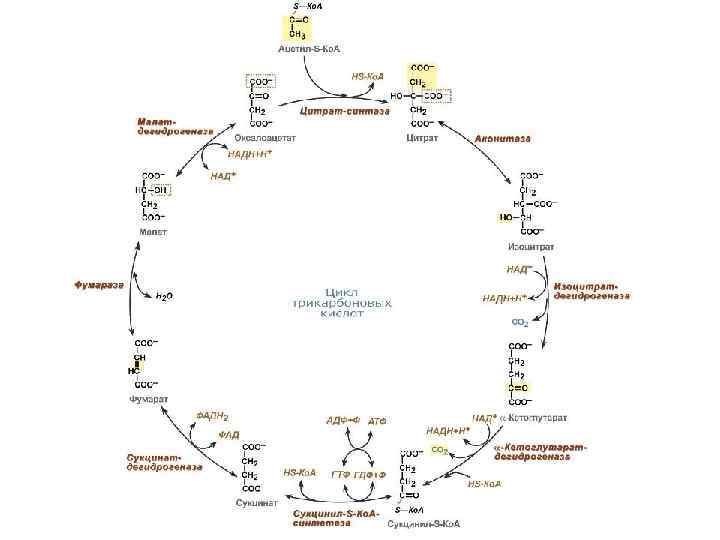
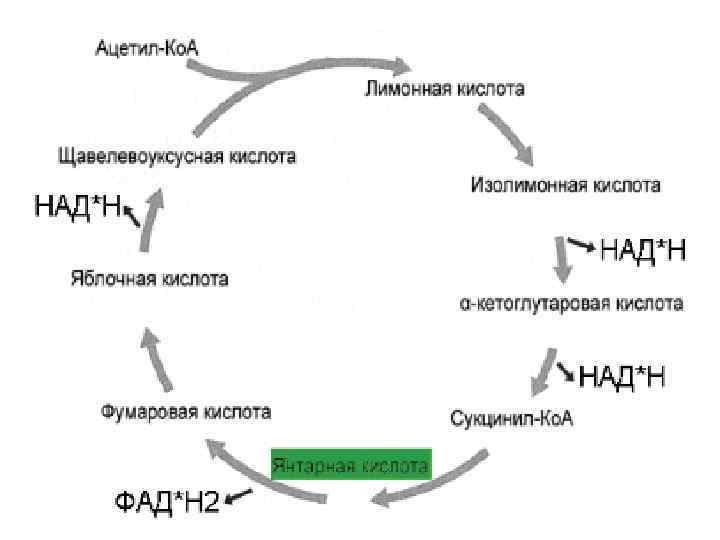

Значение Энергетическое: в цикле Кребса выделяется: 2 СО 2, 3 НАДН+H+ и 1 ФАД·Н 2, 1 АТФ Метаболическое: органические кислоты используются для синтеза аминокислот, жиров и углеводов. Конечный этап окисления углеводов, белков и жиров.

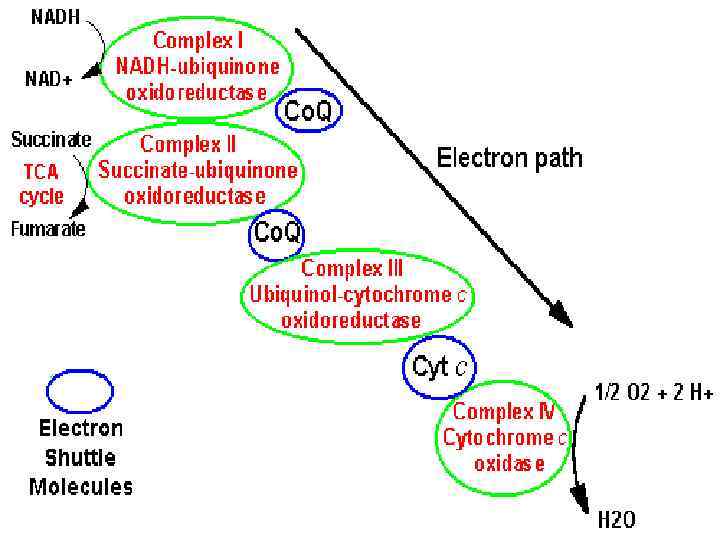
Электронтранспортная цепь локализована во внутренней мембране митохондрий. состоит из более, чем двух десятков переносчиков и значительного числа белков, сгруппированных в крупные ферментные комплексы. включает четыре погруженных в мембрану основных комплекса и два небольших по молекулярной массе лабильных компонента – кофермент Q и цитохром С.

Электронтранспортная цепь ЭТЦ служит для передачи электронов от восстановленных субстратов на кислород. С переносом электронов по ЭТЦ сопряжен синтез АТФ. Энергия высвобождается постепенно путем передачи протонов и электронов ряду переносчиков, связанных с белками и образующих ЭТЦ. Расположение переносчиков в ЭТЦ определяется величиной их окислительно-восстановительного потенциала (О/В). Движущей силой транспорта электронов в ЭТЦ является разность потенциалов. Электроны от НАДН (-0, 32 В) или ФАДН 2 передаются на О 2 (+0, 82 В), который восстанавливается, присоединяя Н+ и образуя Н 2 О: 4 Н + + 4 ē + О 2 → 2 Н 2 О
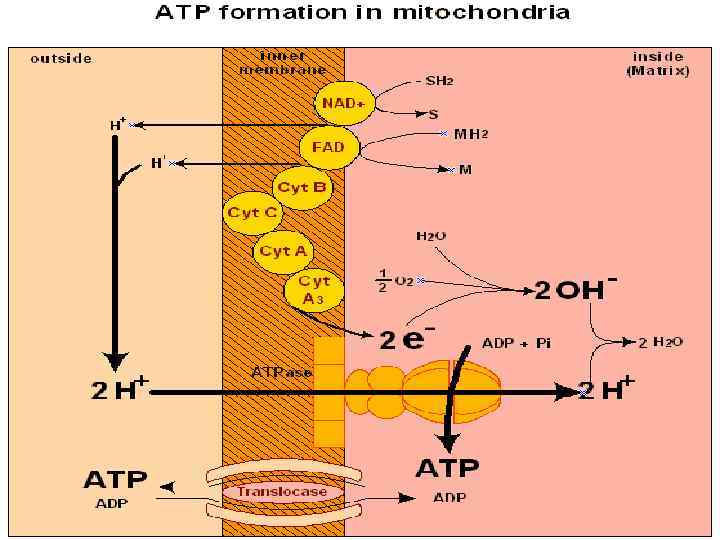
. Сопряжение обеспечивается внутренней митохондриальной мембраной. Происходит перекачивание")
Дыхание сопряжено с окислительным фосфорилированием (сопряженное фосфорилирование). Сопряжение обеспечивается внутренней митохондриальной мембраной. Происходит перекачивание протонов из матрикса и создается протонный градиент. Протонный потенциал - движущая сила синтеза АТФ. При переносе пары электронов от НАД на кислород на внешнюю сторону мембраны выделяется 6 (3 пары) протонов.
 поглощенному в процессе")
Коэффициент фосфорилирования — отношение количества связанного неорганического фосфора (АДФ + Фн→АТФ) поглощенному в процессе дыхания кислороду (Р/О) Коэффициент фосфорилирования — показатель сопряженности окисления и фосфорилирования Коэффициент Р/О может быть не более 3, т. к. перенос двух электронов к кислороду по дыхательной цепи сопровождается не более чем тремя фосфорилированиями. Зависит от условий и от состояния клеток: природы окисляемого вещества внешних условий (температура, обезвоживание и набухание митохондрий, разобщители). снижается при заболевании организмов.

Окислительное фосфорилирование - это накопление энергии окисления в АТФ при продвижении электрона по цепи переносчиков. Хемиосмотическая теория разработана в 1961 -1966 гг. П. Митчелл, англ. биохимик, Нобелевский лауреат (1978) Дыхание → протонный потенциал (∆μΗ+) → АТФ Сопряжение диффузии протонов с синтезом АТФ осуществляется АТФсинтазным комплексом.
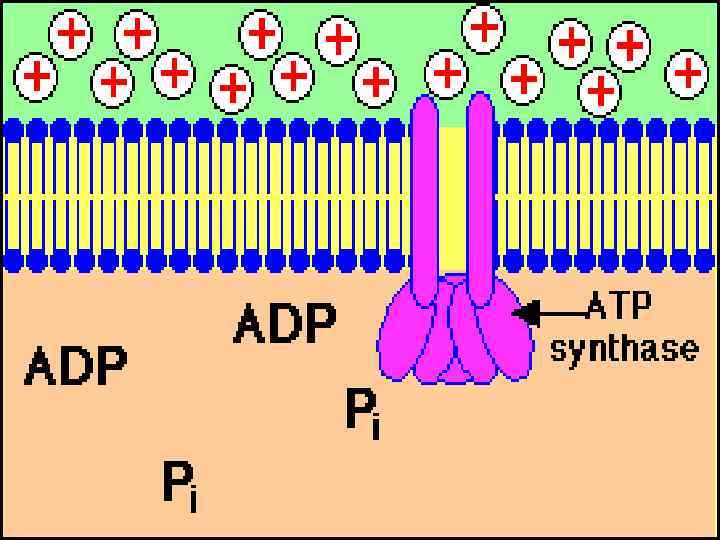

АТРсинтазный комплекс СF 0 – интегральный гидрофобный белок мембраны, протонный канал, 4 типа пептидов (a, b, b', c 12) СF 1 – гидрофильный белковый комплекс, 5 типов полипептидов: α, β, γ, δ, ε.
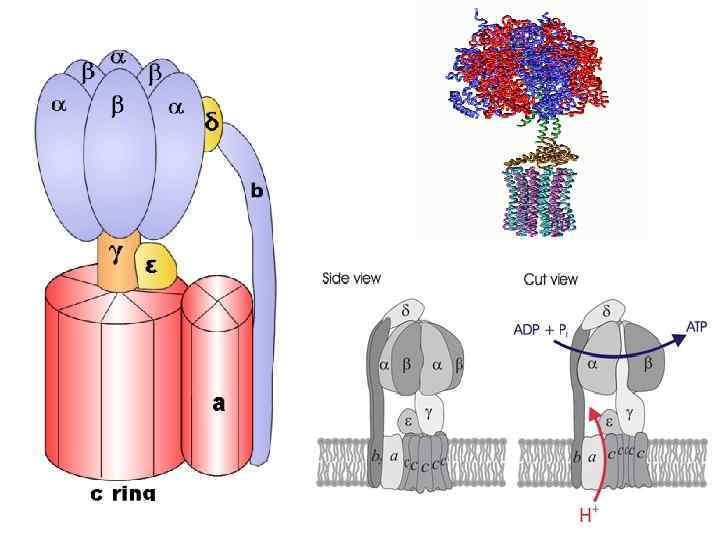

Модель АТФсинтазы P. Boyer, J. Walker Нобел. премия, 1997 Ротационный механизм работы ферментного комплекса: Энергия, освобождаемая при диффузии протонов через F 0 канал, используется для вращения γсубъединицы

Энергозависимые изменения состояния каталитических центров Три этапа синтеза АТФ : • Присоединение АДФ и Фн к активному центру фермента без затраты энергии. • Конформационные изменения фермента, синтез АТФ из АДФ и Фн. • Высвобождение АТР и возврат АТР-синтазного комплекса в исходное состояние.

Схема, иллюстрирующая вращение γ субъединицы АТФсинтазы


Энергетический баланс дыхания Этапы дыхания Генерация энерг. эквивалентов Энерг. выход дыхания Гликолиз 2 АТФ 2 НАДН 2 АТФ Окислительное декарбоксилирование ПВК 2 НАДН Цикл Кребса 2 АТФ 6 НАДН 2 ФАДН 2 ЭТЦ: окисление 10 НАДН окисление 2 ФАДН 2 2 АТФ 30 АТФ 4 АТФ 38 АТФ Х 30, 6 к. Дж = 1162, 8 к Дж - затрачено на образование 2824 к. Дж – выделяется при распаде глюкозы, следовательно КПД ≈ 40 %

Глиоксилатный цикл Впервые описан в 1957 г. у бактерий и плесневых грибов Г. Л. Корнбергом и Г. А. Кребсом. Характеристика: • функционирует в прорастающих семенах масличных растений и в других объектах, где запасные жиры превращаются в сахара (стареющие листья, при созревании пыльцы) • локализован в специализированных микротельцах — глиоксисомах. • отсутствуетв клетках животных, в вегетативных тканях растений и в созревающих семенах. • Перестает функционировать после того, как израсходован весь запас жира.
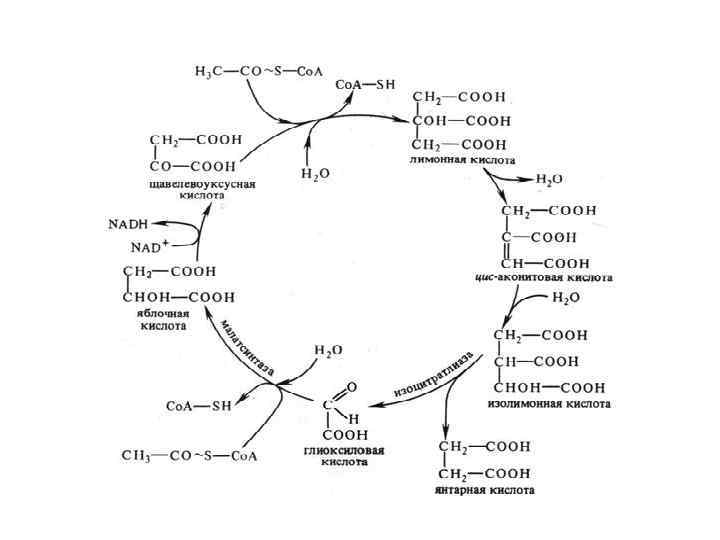

В глиоксилатном цикле по сравнению с циклом Кребса: • Участвует 2 молекулы ацетил-Со. А • Ацетил-Со. А используется для синтеза янтарной кислоты, которая может выходить из глиоксисом, превращаться в ЩУК и участвовать в глюконеогенезе (обращенном гликолизе) и других процессах биосинтеза. • Отсутствует декарбоксилирование • Участвует 2 специфических фермента: лиаза и малатсинтаза. • • Значение: Энергетическое – 1 НАДН (3 АТФ) С помощью цикла жиры превращаются в углеводы. Глиоксилатный цикл позволяет утилизировать запасные жиры, при распаде которых образуются молекулы ацетил-Со. А.
, не ингибируется цианидом. Характерен для семенных растений, грибов, водорослей.")
Альтернативный путь дыхания Цианидустойчивый (цианидрезистентный), не ингибируется цианидом. Характерен для семенных растений, грибов, водорослей. Цианидустойчивое дыхание: - связано с функционированием в дыхательной цепи альтернативной оксидазы. - энергия в основном не аккумулируется в АТФ, а рассеивается в виде тепла. - ингибируется салициловой кислотой - функцию альтернативной оксидазы в разных тканях выполняют 1— 3 белка, которые кодируются в ядерном геноме. - составляет 10 - 25 % (до 100 %) общего поглощения кислорода. - активируется при высоком содержании АТФ и ингибировании основной цепи транспорта электронов при дыхании.
 Глюкозо-6 -фосфат→ СО 2+НАДФ+ +рибулозо-5 -фосфат")
Пентозофосфатный путь -ПФП апотомический, или фосфоглюконатный Этапы: 1) Глюкозо-6 -фосфат→ СО 2+НАДФ+ +рибулозо-5 -фосфат 2) Регенерация глюкозо-6 -фосфата
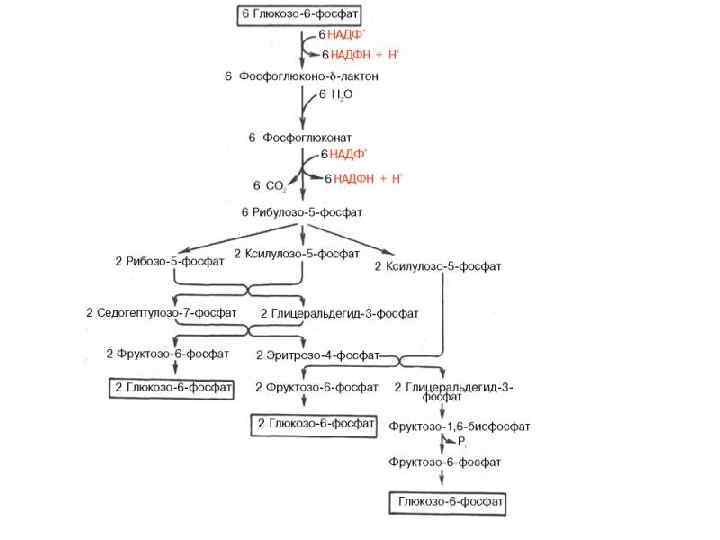
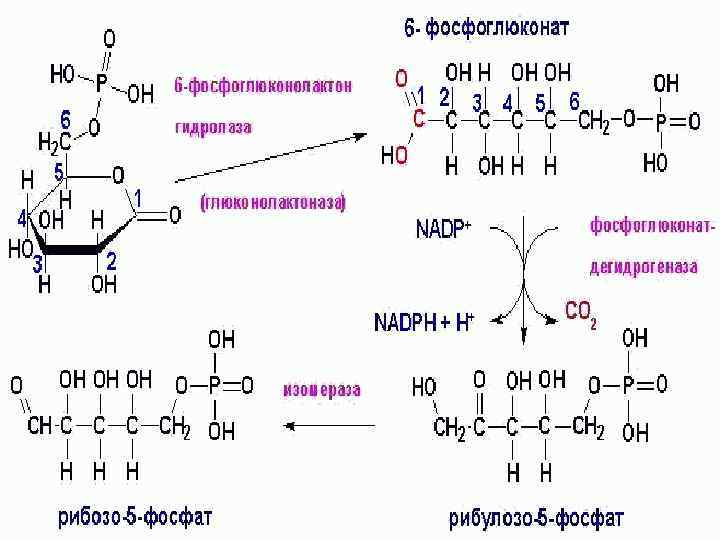

Значение • • • Энергетическое. Метаболическое, образуются: - НАДФН, который используется на синтетические процессы -углеводы (от С 3 до С 7), пентозы -рибулозо-1, 5 -дифосфат и НАДФН принимают участие в темновой фиксации СО 2. В хлоропластах функционирует в темноте, предотвращает резкое изменение концентрации НАДФН в отсутствие света. При н. у. ПФП составляет 10 -40%, зависит от типа ткани и состояния: – Преобладает в хлоропластах (ПФП > гликолиз) – Снижается в анаэробных условиях (гликолиз > ПФП), т. к. подавляется работа дегидрогеназ цикла. – При неблагоприятных условиях активность ПФП увеличивается : засуха, дефицит калия, инфекция, засоление, затенение, старение.
Лекция_15_17_дыхание_2013.ppt