Система гистосовместимости HLA.pptx
- Количество слайдов: 31



HLA Система гистосовместимости человека Выполнила: Бадалян Д. А. II ЛФ 12 группа
 – это группа генов и кодируемых ими антигенов клеточной поверхности, которые")
HLA (MHC) – это группа генов и кодируемых ими антигенов клеточной поверхности, которые играют важнейшую роль в распознавании чужеродного и развитии иммунного ответа. Антигены HLA представляют собой гликопротеиды, находящиеся на поверхности клеток и кодируемые группой тесно сцепленных генов 6 -й хромосомы.

Генетические локусы, ответственные за отторжение чужеродных тканей, образуют в хромосоме область, названную главным комплексом гистосовместимости (MHC). Высокий полиморфизм молекул MHC, а также способность каждой антигенпрезентирующей клетки (АПК) экспрессировать несколько разных молекул MHC обеспечивают возможность презентации T-клеткам множества самых различных антигенных пептидов. Следует отметить, что хотя молекулы MHC и называются обычно антигенами, они проявляют антигенность только в том случае, когда распознаются иммунной системой не собственного, а генетически иного организма, например, при аллотрансплантации органов.

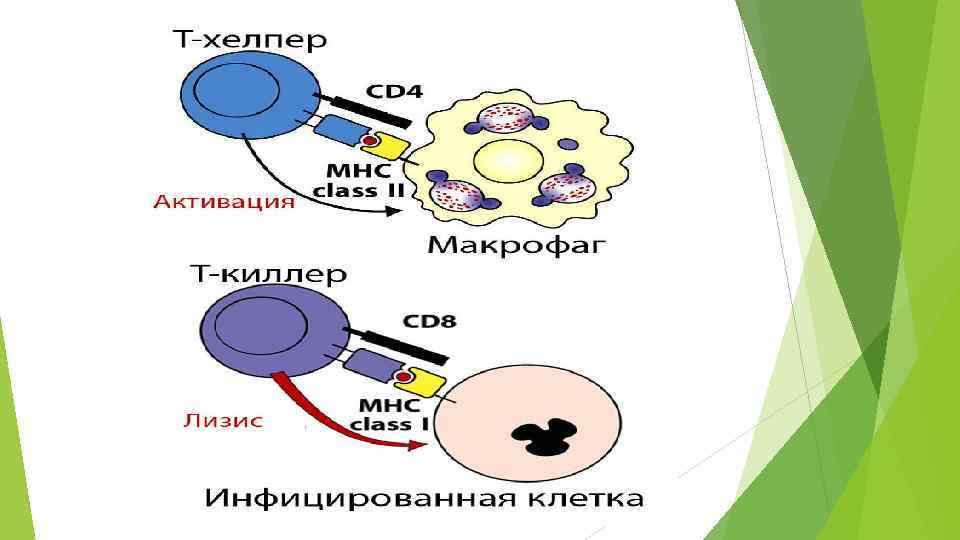
Классы антигенов HLA Антигены HLA класса I необходимы для распознавания трансформированных клеток цитотоксическими Т-лимфоцитами. Способны связывать пептиды из 8 -9 аминокислотных остатков Антигены HLA класса II - обеспечивают взаимодействия между Тлимфоцитами и макрофагами в процессе иммунного ответа. Тхелперы распознают чужеродный антиген лишь после его переработки макрофагами, соединения с антигенами HLA класса II и появления этого комплекса на поверхности макрофага. Способны связывать несколько более длинные пептиды, чем I класс.

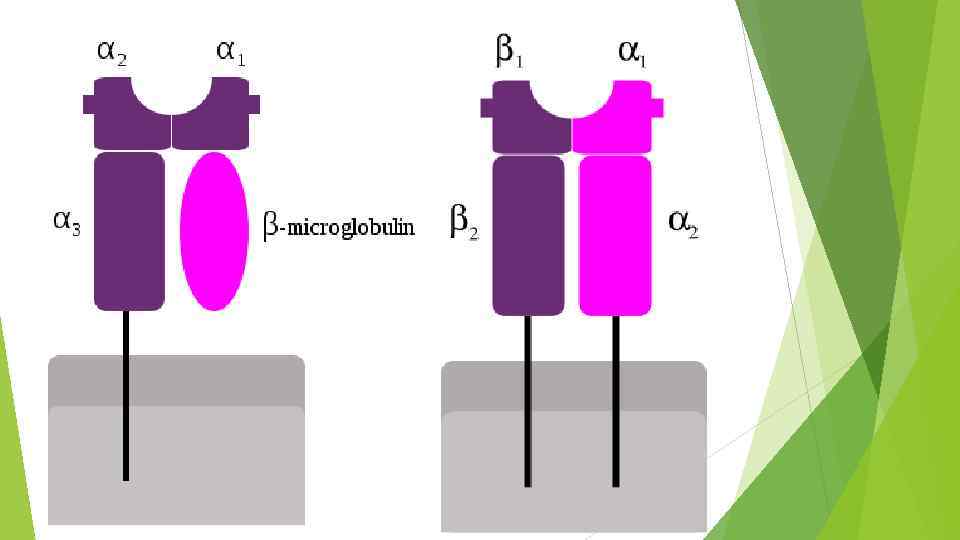
Строение MHC класса I Молекулы MHC класса I (аллельные варианты HLA : HLAA , HLA-B , HLA-С ) экспрессируются на клеточной поверхности и представляют собой гетеродимер, состоящий из одной тяжелой альфа-цепи (45 к. Да), нековалентно связанной с однодоменным бета 2 микроглобулином (12 к. Да). Тяжелая цепь состоит из внеклеточной части (образующей три домена : альфа 1 -, альфа 2 - и альфа 3 -домены), трансмембранного сегмента и цитоплазматического хвостового домена. Каждый внеклеточный домен содержит примерно 90 аминокислотных остатков.
 необходим для экспрессии всех")
Строение MHC класса I Бета 2 -микроглобулин (бета 2 -m) необходим для экспрессии всех молекул MHC класса I и имеет неизменную последовательность. По структуре этот белок соответствует C-домену иммуноглобулинов. Бета 2 микроглобулин способен также нековалентно взаимодействовать с неклассическими молекулами класса I , например, с продуктами генов CD 1. В зависимости от вида и гаплотипа внеклеточная часть тяжелых цепей MHC класса I в разной степени гликозилирована. Трансмембранный сегмент MHC I класса состоит из 25 преимущественно гидрофобных аминокислотных остатков и пронизывает липидный бислой, вероятнее всего, в альфаспиральной конформации.

Строение MHC класса I
 и представление")
MHC класса I Основное свойство молекул I класса - связывание пептидов (антигенов) и представление их в иммуногенной форме для Т-клеток - зависит от доменов альфа 1 и альфа 2. Эти домены имеют значительные альфа- спиральные участки, которые при взаимодействии между собой образуют удлиненную полость (щель), служащую местом связывания процессированного антигена. Образовавшийся комплекс антигена с альфа 1 - и альфа 2 -доменами и определяет его иммуногенность и возможность взаимодействовать с антигенраспознающими рецепторами Т-клеток. К классу I относятся антигены A , антигены AB и антигены AC. Антигены класса I присутствуют на поверхности всех ядросодержащих клеток и тромбоцитов.

Строение MHC класса II Молекулы MHC класса II являются гетеродимерами, построенными из нековалентно сцепленных тяжелой альфа- и легкой бета-цепей. Ряд фактов указывает на близкое сходство альфа- и бета-цепей по общему строению. Внеклеточная часть каждой из цепей свернута в два домена (альфа 1, альфа 2 и бета 1, бета 2, соответственно) и соединена коротким пептидом с трансмембранным сегментом (длиной примерно 30 аминокислотных остатков). Трансмембранный сегмент переходит в цитоплазматический домен, содержащий примерно 1015 остатков.

Строение MHC класса II

Отличия в строении MHC II Антигенсвязывающая область молекул MHC класса II формируется альфа-спиральными участками взаимодействующих цепей подобно молекулам I класса , но при одном существенном отличии: антигенсвязывающая полость молекул MHC класса II формируется не двумя доменами одной альфа-цепи, а двумя доменами разных цепей доменами альфа 1 и бета 1. Общее структурное сходство между двумя классами молекул MHC очевидно. Это - однотипность пространственной организации всей молекулы, количество доменов (четыре), конформационное строение антигенсвязывающего участка, близкие мол. веса. В структуре молекул II класса антигенсвязывающая полость открыта больше, чем у молекул I класса, поэтому в ней могут поместиться более длинные пептиды.
 класса II обеспечение взаимодействия между Т-лимфоцитами")
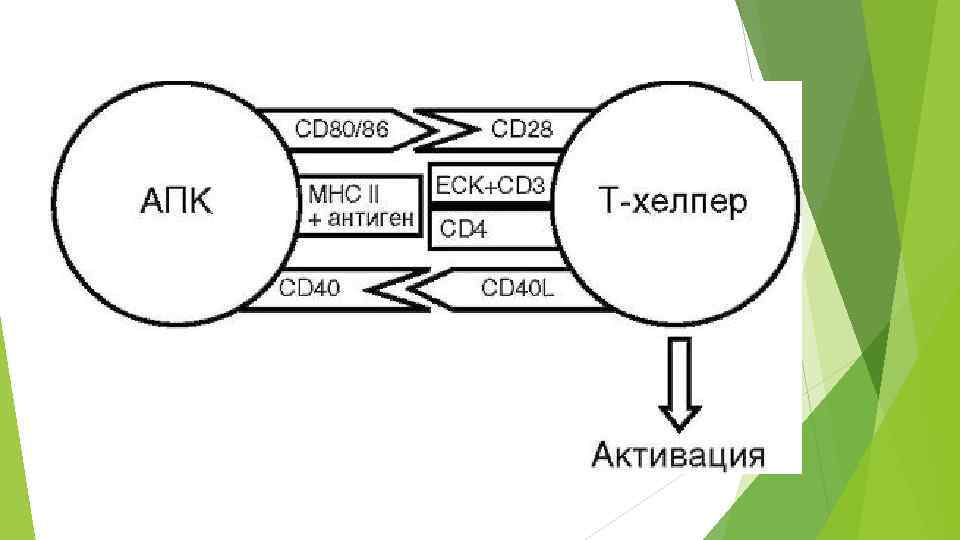
MHC класса II Важнейшая функция антигенов MHC (HLA) класса II обеспечение взаимодействия между Т-лимфоцитами и макрофагами в процессе иммунного ответа. Т-хелперы распознают чужеродный антиген лишь после его переработки макрофагами, соединения с антигенами HLA класса II и появления этого комплекса на поверхности макрофага. Антигены класса II присутствуют на поверхности В-лимфоцитов , активированных Т-лимфоцитов, моноцитов, макрофагов и дендритных клеток. Гены MHC класса II кодируют связанные с мембраной трансмембранные пептиды (гликопротеины). Молекулы антигенов гистосовместимости класса II ( DR , DP , DQ ).

Молекулы MHC класса II Каждая из цепей этих молекул состоит из двух Ig-подобных доменов, трансмембранного и цитоплазматического участков. Имеются некоторые различия между антигенами MHC класса II , кодируемыми локусами DR, DP и DQ. У продуктов генов DQ-локуса альфа-цепь является инвариантной, а аллельное разнообразие обусловливается бета-цепью, прежде всего бета 1 -доменом. В случае антигенов, кодируемых DP и DQ локусами, для обеих цепей обнаружены аллельные формы, однако и для них основной вклад вносится бета 1 -доменом. У антигенов MHC класса II антигенсвязывающий центр формируется альфа 1 - и бета 1 доменами, в то время как роль примембранных доменов сводится к усилению связывания АПС - T-хелперы за счет взаимодействия с CD 4 -антигеном. Продукты MHC класса II ассоциированы, главным образом, с Bлимфоцитами и макрофагами и служат распознаваемыми структурами для Tхелперов. Антигены MHC класса II - крайне полиморфные семейства молекул.

Молекулы MHC III, IV класса Гены класса III контролируют синтез белков, часть из которых принимает участие в иммунных процессах: один из компонентов комплемента , факторы некроза опухолей ( ФНОальфа и ФНО-бета ). Здесь же локализованы гены, контролирующие образование ряда ферментов синтеза стероидов. Гены MHC класса III, расположенные в пределах группы генов MHC или тесно сцепленные с ней, контролируют некоторые компоненты комплемента : C 4 и C 2 , а также фактор B , находящиеся скорее в плазме крови, чем на поверхности клеток. И в отличие от молекул MHC класса I и класса II не участвуют в контроле иммунного ответа. Термин MHC класса IV употребляется для описания некоторых локусов, сцепленных с MHC.

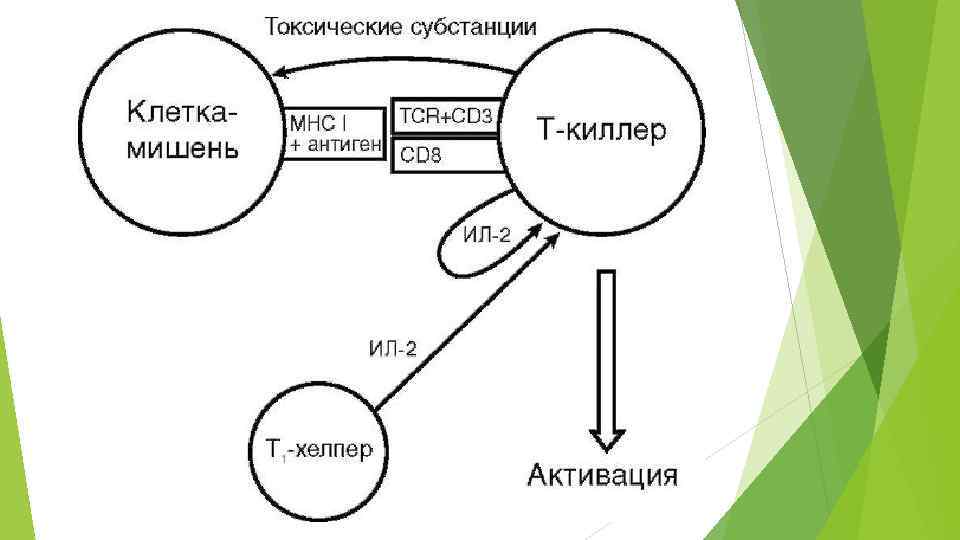
ПРОЦЕССИНГ И ПРЕЗЕНТАЦИЯ АНТИГЕНА Заключается в частичном изменении структуры антигена и связывании его с молекулой МНС класса II или I, что необходимо для развития полноценной иммунной реакции. Связывание антигена с молекулами MHC нужно для того, чтобы антиген смог взаимодействовать с рецептором для антигена на Тлимфоцитах, так как сам рецептор для антигена сопряжен с рецептором для молекул MHC. Соединение антигена с рецептором может осуществляться только при одновременном взаимодействии молекул MHC со своими рецепторами. Процессинг и представление антигенов осуществляется в антигенпредставляющих клетках ( макрофагах, клетках Лангерганса, В-лимфоцитах) и идет в несколько этапов.

А - 1 -й этап: эндоцитоз антигена, при этом антиген оказывается внутри клетки, заключенный в фагоцитарную вакуоль Б - 2 -й этап: фагоцитарная вакуоль связывается с лизосомой, лизосомальные ферменты частично разрушают или модифицируют антиген, видоизмененный антиген связывается с молекулой главного комплекса гистосовместимости (MHC) II или I класса внутри фагоцитарной вакуоли В - 3 -й этап: экзоцитоз видоизмененного антигена в комплексе с молекулой МНС на поверхность антигенпредставляющей клетки.

ПРОЦЕССИНГ И ПРЕЗЕНТАЦИЯ АНТИГЕНА

Функции MHC Способны вызывать отторжение трансплантата Принимают непосредственное участие в инициации иммунного ответа, контролируя молекулы, представляющие антиген в иммуногенной форме для его распознавания цитотоксическими T-клетками и хелперными Tклетками. В МНС локализованы гены, контролирующие синтез иммунорегуляторных и эффекторных молекул - цитокинов ФНО-альфа , ФНО-бета , а также некоторых компонентов комплемента. Клеточные маркеры, распознаваемые цитотоксическими Tлимфоцитами и T-хелперами в комплексе с антигеном. Молекулы MHC класса I могут входить в состав гормональных рецепторов. Описаны случаи ассоциации продуктов MHC с рецепторами глюкагона , эпидермального фактора роста и гаммаэндорфина.

Иммунологические свойства, связанные с MHC • Интенсивное отторжение трансплантатов тканей • Стимуляция образования антител • Стимуляция реакции в смешанной культуре лимфоцитов • Реакция "трансплантат против хозяина" • Клеточная реакция лимфолиза • Гены иммунного ответа (Ir) (англ. immune response) • Рестрикция иммунного ответа

Маркеры HLA Гены системы HLA, особенно локусы DR, многофункциональны. Они контролируют образование трансплантационных Аг, силу иммунного ответа, положительные и отрицательные ассоциации с различными болезнями. Таких генов более 30. Определённые HLA-Ar ассоциируются с некоторыми патологическими процессами. Около 10 болезней ассоциируются с областями А, В, С; 33 патологических состояния зависят от 8 HLA-DR Аг и т. д.

Нередки ситуации, когда несколько генов имеют «привязку» к одной болезни. D/DR-4, D/DR-2, D/DR-3 положительно связаны с инсулинозависимым сахарным диабетом. Аг HLA-B 27 в популяции здоровых лиц встречается с частотой 9%. У больных с хроническим анкилозирующим спондилитом (болезнь Бехтерева) — в 96%. Тот же Аг регистрируется при болезни Рейтера в 76— 80%, при псориазе и миеломной болезни — в 35— 40%. HLA-B 8 у здоровых людей представлен в 16%. У пациентов с поражением надпочечников (болезнь Аддисона) — в 80%, при подростковом ювенильном диабете — в 60%, герпетоидной форме дерматита — в 60%, хронических формах колита — в 60%, СКВ и хроническом гепатите — в 40%. При рассеянном склерозе обнаружено возрастание частоты встречаемости HLA-A 3 и HLA-B 17, при аллергии к пыльце растений — HLA-B 7, при тиреотоксикозе — HLA-A 2, инфекционном мононуклеозе — HLA-B 35. Даже проявление заболеваний может зависеть от Аг гистосовместимости. Например, тяжелое течение СКВ, при котором выраженыпоражения почек и аутоиммунные проявления, положительно коррелирует с Аг HLA-A 1 и HLA-B 8, более лёгкое — с HLAA 2 и HLA-B 7.

1. 2. 3. 4. Таким образом, человек, обладая тем или иным аллелем гена, имеет определённую степень риска заболеть данной болезнью или группой болезней. Их можно распределить на 4 категории. Иммунные расстройства: миастения гравис, системная красная волчанка, дефициты иммунных глобулинов. Болезни с аутоиммунным компонентом: инсулинозависимый сахарный диабет, тиреотоксикозы, ревматоидный артрит. Болезни неизвестной этиологии: рассеянный склероз, различные псориазы. Болезни без иммунного компонента: идиопатические гематохро-матозы, конгенитальная дисплазия.

Склонность человека к какому-либо патологическому процессу может быть первичной, когда ген предрасположенности к заболеванию одновременно служит HLA-Ar вторичной, когда есть ассоциация между геном предрасположенности и HLA-Ar. Возможен, также вариант, когда HLA-Ar-маркер ассоциирован или находится в неравновесном сцеплении с Аг, в свою очередь ассоциированным с геном предрасположенности. К сожалению, далеко не все HLA гены и Аг (особенно это касается класса II системы HLA) установлены, так что исследователи часто имеют дело с последним вариантом ассоциации.

При введении вакцин реакция разных людей неодинакова. На основании данных исследований сформулировано новое толкование иммунного статуса, который определяется суммарным эффектом активности генов, кодирующих клеточные и гуморальные факторы иммунитета и определяющих однотипную по силе иммунную реактивность к широкой группе Аг.

Тканевое распределение молекул I и II классов МНС человека Тип клеток В-клетки Тимоциты Макрофаги Гранулоциты Ретикулоциты Эритроциты Тромбоциты Фибробласты Эпителиальные клетки Эпидермальные клетки Печень Почка Сердечная мышца Скелетная мышца Мозг Плацента Сперматозоиды Яйцеклетки Трофобласт Бластоциты Эмбриональная ткань HLA комплекс человека Класс I + + + + (+) + +. (+). + Класс II + (+) +. + +. . . .

Участие молекул I и II классов МНС в некоторых иммунных реакциях Функциональная активность Класс II Интенсивность отторжения аллотрансплантата ++++ ++ Индукция образования антител ++++ Стимуляция СКЛ + ++++ Инициация РТПХ ++ ++++ Презентация антигена для цитотоксических Т- клеток ++++ - Презентация антигена для хелперных Т- клеток - ++++

Спасибо за внимание
Система гистосовместимости HLA.pptx