Гормони_p.ppt
- Количество слайдов: 96



Гормони

ПЛАН ЛЕКЦІЇ 1. Біологічна роль, хімічна природа та класифікація гормонів. 2. Ієрархія дії гормонів. 3. Сучасні уявлення про молекулярні механізми дії гормонів. 4. Гормони різних залоз внутрішньої секреції.


Гомеостатичний контроль рівня глюкози інсуліном та глюкагоном

A Brief Sequential Overview of the Regular Estrous Cycle • • Follicle stimulating hormone (FSH) and Luteinizing hormone (LH)

Залози внутрішньої секреції

Лептин – гормон жирової • Гормон лептин — тканини це білок, що складається зі 167 амінокислотних залишків загальною молекулярною масою 16 к. Да, член сімейства цитокінів, який утворюється у жирових клітинах і доносить до центрів апетиту в гіпоталамічних ділянках мозку інформацію про жировий обмін і масу тіла

Лептин – гормон жирової тканини Підвищений синтез лептину корелює із початком статевої зрілості. Лептин подає сигнал про достатню кількість жирової тканини для початку репродуктивної функції. При недостатній кількості лептину стимулюється синтез гонадотропіну, який може призупиняти виникнення менструації у жінок. Тобто лептин також може впливати на сексуальну функцію. Лептин є гормоном “антиожиріння” і належить до продуктів “генів економії”. Якщо розглянути його роль в еволюційному аспекті, коли наші предки їли нерегулярно, а проміжки між вживанням їжі були тривалими, то вона полягала в ефективному збереженні енергії протягом всього періоду вимушеного голодування, під час якого для виживання необхідні були великі запаси енергії.

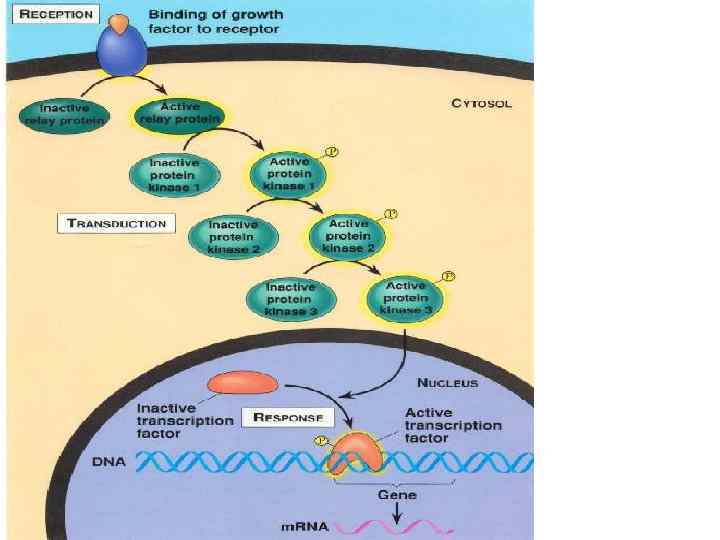
Нейрогуморальна регуляція Гормони діють за функціональною ієрархією. Нервові імпульси, що надходять у гіпоталамус, зумовлюють виділення специфічних гіпоталамічних гормонів, які потрапляють у гіпофіз і стимулюють (або пригнічують) вивільнення ним різних тропних гормонів. Гормони передньої частки гіпофізу, відповідно, стимулюють виділення гормонів іншими ендокринними залозами, що секретують гормони, які діють на специфічні тканини-мішені. На клітини-мішені гормони діють за участю білків-рецепторів. Взаємодія відбувається головно шляхом нековалентного зворотного зв’язування, унаслідок чого утворюються білоклігандні комплекси, що зумовлюють різні гормональні ефекти у клітині.

Ієрархія дії гормонів.

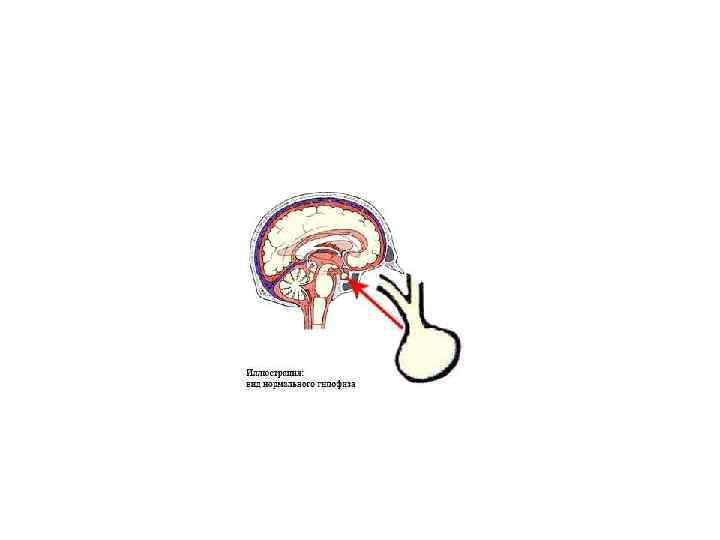
Гіпоталамус, гіпофіз, епіфіз

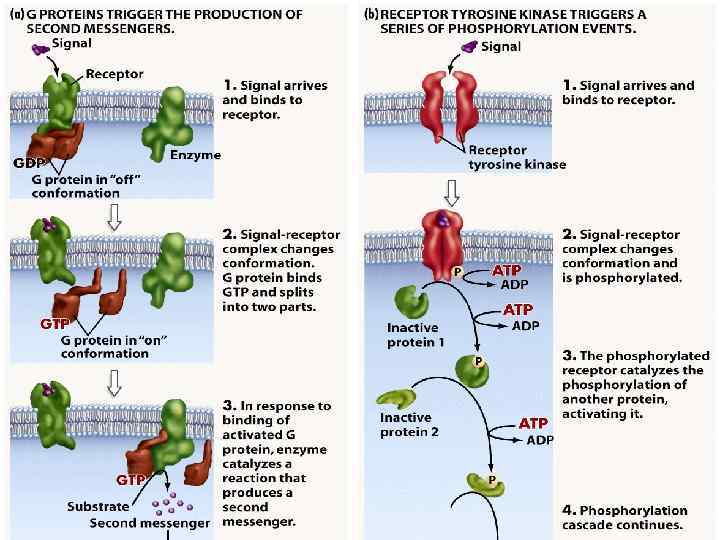
Механізм дії гормонів на клітинному рівні • За локалізацією рецепторних білків циторецепцію ормонів г поділяють на два типи: внутрішньоклітинну (цитозольну, або пряму) – за цим типом діють стероїдні й тиреоїдні гормони, і мембранну – таким способом ідбувається в рецепція білковопептидних гормонів і катехоламінів.

Механізм дії гормонів на клітинному рівні
, що передають сигнал від гормонального")
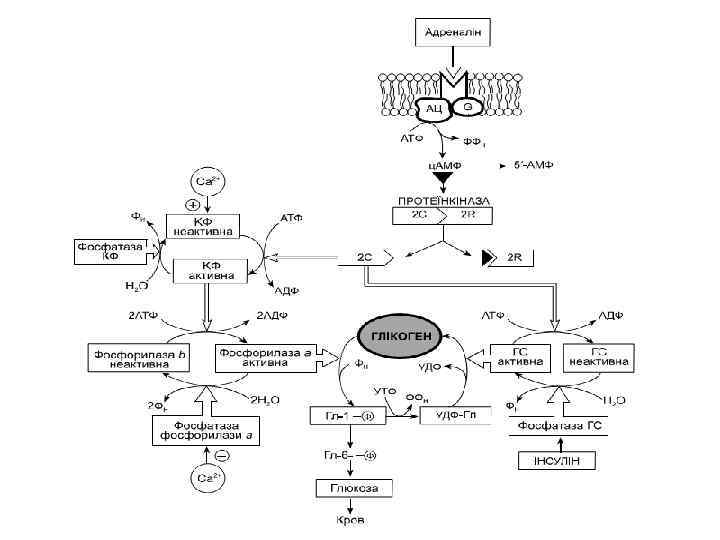
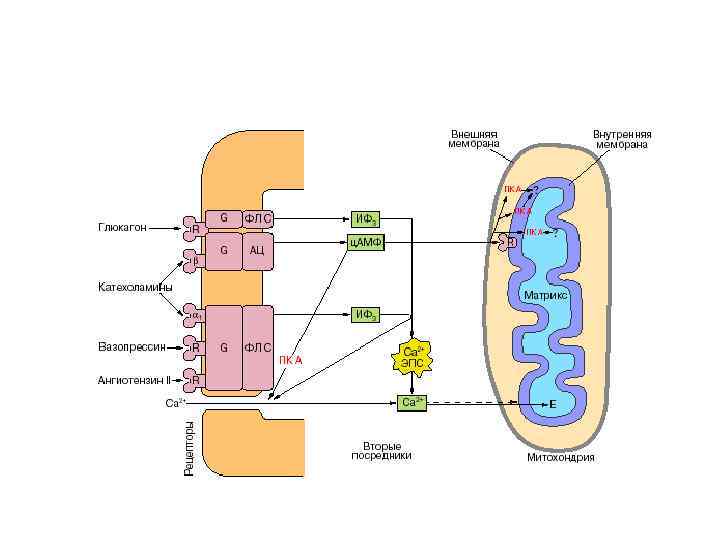
Дія гормонів може відбуватися через внутрішньоклітинні вторинні посередники (месенджери), що передають сигнал від гормонального рецептора на специфічну внутрішньоклітинну структуру чи фермент, які власне і є кінцевою мішенню дії гормону. Сьогодні відомі два головні внутрішньоклітинні медіатори дії гормонів: аденілат-циклазна система, що утворює ц. АМФ та „повільні” кальцієві канали, що визначають проникнення Са 2+ у клітину. Ці шляхи можуть діяти незалежно, та вони можуть бути і тісно пов’язані між собою. Можливе включення і гуанілатциклазної системи, однак це відбувається опосередковано, за участю Са 2+ і лише у деяких тканинах. Посередниками у здійсненні ефектів є ц. АМФ-залежні протеїнкінази, що впливають на активність багатьох ферментів і неферментних білків. Головний посередник у здійсненні ефектів іонів Кальцію – кальмодулін – основний кальційзв’язувальний білок. Чинником, який припиняє дію ц. АМФ-залежної протеїнкінази, є фосфодіестераза ц. АМФ, яка може бути активована різними шляхами.

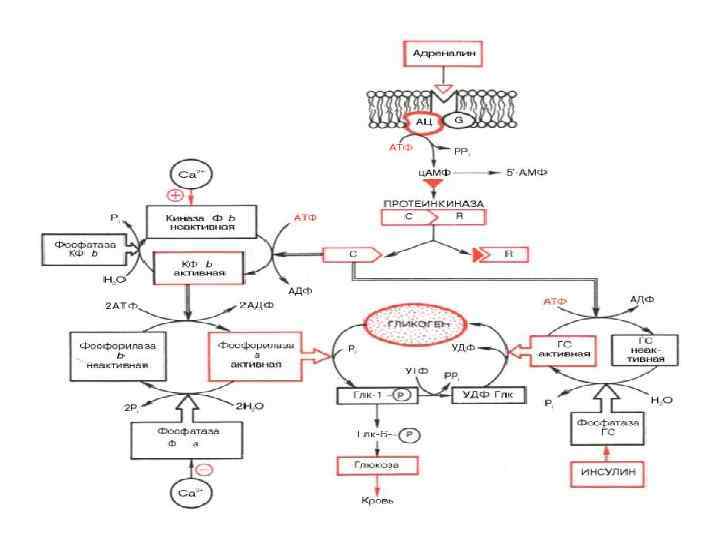
Аденілатциклазна система

Аденілатциклаза • Аденілатциклаза є інтегральним білком плазматичних мембран, її активний центр орієнтований у бік цитоплазми і каталізує реакцію синтезу ц. АМФ з АТФ:

Протеїнкіназа • Протеїнкіназа – це внутрішньоклітинний фермент, через який ц. АМФ реалізує свій ефект. Протеїнкіназа може існувати у двох формах. За відсутності ц. АМФ, протеїнкіназа представлена у вигляді тетрамерного комплексу, що складається з двох каталітичних (С 2) та двох регуляторних (R 2) субодиниць з мол. масами 49000 і 38000 дальтон відповідно. У цій формі фермент є неактивним. За присутності ц. АМФ протеїнкіназний комплекс зворотно дисоціює на димер R 2 та дві вільні каталітичні субодиниці С. Останні мають ферментативною активністю. Каталізуючи фосфорилювання білків і ферментів, вони здатні змінювати інтенсивність метаболічних процесів.
 і ГТФсв’язуючих білків.")
Трансмембранна передача інформації за участю аденілатциклази (АС) і ГТФсв’язуючих білків.

• Одним із найважливіших механізмів проведення гормонального сигналу в кальціймесенджерній системі є запуск клітинних реакцій (відповіді) шляхом активації специфічної Са 2+-кальмодулін-залежної протеїнкінази. Регуляторною субодиницею даного ферменту є білок кальмодулін (мол. маса 17 000). При підвищенні концентрації Са 2+ в клітині у відповідь на поступаючі сигнали, специфічна протеїнкіназа каталізує фосфорилювання багатьох внутрішньоклітинних ферментів – мішеней, регулюючи тим самим їхню активність. До складу кінази фосфорилази b (яка активується іонами Са 2+) входить кальмодулін як субодиниця. При підвищенні концентрації Кальцію звязування Са 2+ з кальмодуліном супроводжується його конформаційними змінами, і в такій Са 2+зв’язаній формі кальмодулін модулює активність багатьох внутрішньоклітинних білків (звідси його назва).

Інозитол-1, 4, 5 -трифосфат зв’язується зі специфічним рецептором на ендоплазматичному ретикулумі, сприяючи виходу іонів Са 2+ у цитозоль До внутрішньоклітинної системи месенджерів відносять також похідні фосфоліпідів мембран еукаріотичних клітин, зокрема фосфорильовані похідні фосфатидилінозитолу. Ці похідні вивільняються у відповідь на гормональний сигнал (наприклад, від вазопресину чи тиреотропіну) під дією специфічної мембранозв’язаної фосфоліпази С. У результаті послідовних реакцій утворюються два потенційні вторинні месенджери – діацилгліцерол та інозитол-1, 4, 5 -трифосфат.

• Біологічні ефекти цих вторинних месенджерів реалізуються по-різному. Дія діацилгліцеролу, як і вільних іонів Са 2+, опосередковується через мембранозв’язаний Са 2+-залежний фермент протеїнкіназу С, яка каталізує фосфорилювання внутрішньоклітинних ферментів, змінюючи їхню активність. Інозитол-1, 4, 5 -трифосфат зв’язується зі специфічним рецептором на ендоплазматичному ретикулумі, сприяючи виходові іонів Са 2+ у цитозоль.

Kожній із систем посередників гормонального ефекту відповідає певний клас протеїнкіназ. Активність протеїнкіназ типу А регулюється ц. АМФ, протеїнкінази G – ц. ГМФ; Са 2+-кальмодулінзалежні протеїнкінази перебувають під контролем внутрішньоклітинної концентрації іонів Кальцію, а протеїнкіназа типу С регулюється діацилгліцеролом у синергізмі з вільним Са 2+ і кислими фосфоліпідами. Підвищення рівня будь-якого вторинного месенджера призводить до активації відповідного класу протеїнкіназ і фосфорилювання їхніх білкових субстратів. У результаті змінюється не тільки активність, але й регуляторні та каталітичні властивості багатьох ферментних систем клітини: іонних каналів, внутрішньоклітинних структурних елементів і генетичного апарату.

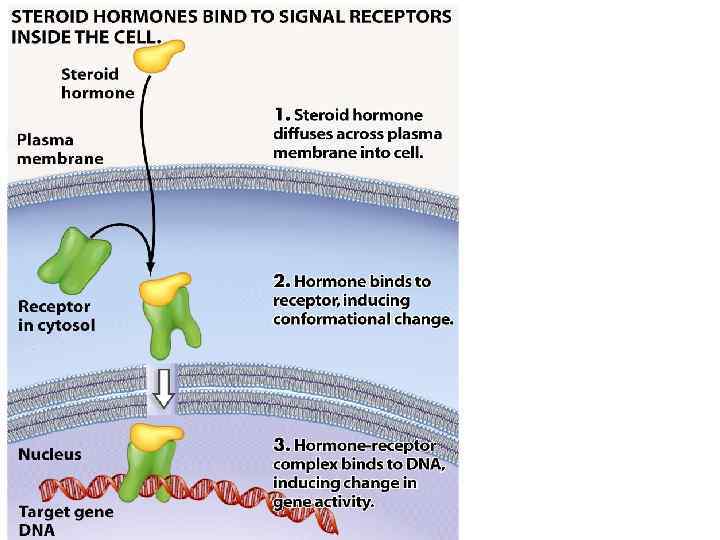
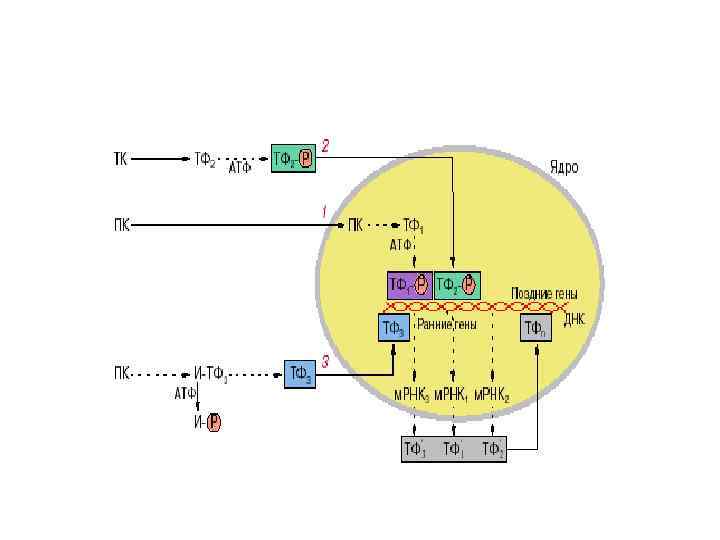
Механізм дії гормонів на клітинному рівні • • Ефект стероїдних гормонів реалізується через генетичний апарат шляхом зміни експресії генів. Гормон транспортується з білками крові, проникає у клітину через плазматичну мембрану (шляхом дифузії), пізніше через ядерну мембрану і зв’язується з внутрішньоядерним рецептором. Комплекс стероїд–білок зв’язується з регуляторною ділянкою ДНК (з так званими гормончутливими елементами), і таким чином сприяє транскрипції відповідних структурних генів, індукує синтез белка de novo та зумовлює зміни метаболізму клітини у відповідь на гормональний сигнал.

Механізм дії гормонів на клітинному рівні Основна відмінність молекулярних механізмів дії двох основних класів гормонів полягає в тому, що дія пептидних гормонів реалізується в основному шляхом посттрансляційних (постсинтетичних) модификацій білків у клітинах, тоді як стероїдні гормони (а також тиреоїдні гормони) є регуляторами експресії генів.

ПЛАН ЛЕКЦІЇ 1. Біологічна роль, хімічна природа та класифікація гормонів. 2. Ієрархія дії гормонів. 3. Сучасні уявлення про молекулярні механізми дії гормонів. 4. Гормони різних залоз внутрішньої секреції.

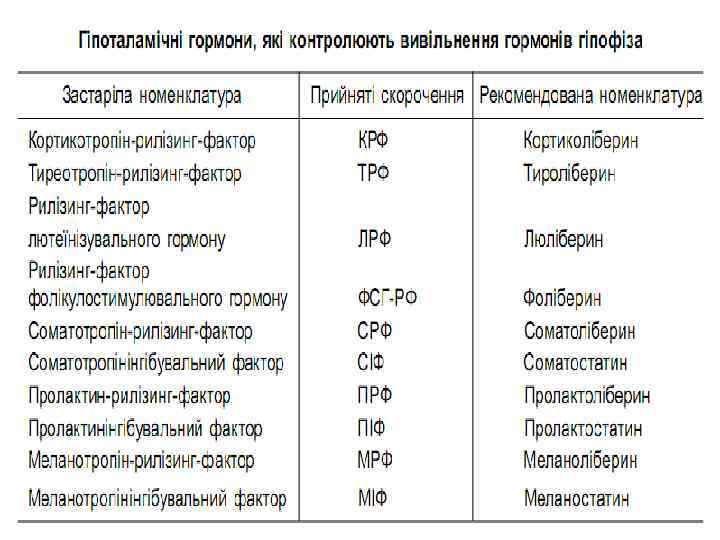
ГОРМОНИ ГІПОТАЛАМУСУ • Гіпоталамус є зоною головного мозку, яка регулює діяльність гіпофіза та перифе-ричних залоз. У гіпоталамусі координується взаємодія вищих відділів центральної нервової та ендокринної систем. Гормони гіпоталамуса, які сприяють виділенню пев-них гормонів гіпофіза, називають рилізинг-факторами (ліберинами), а ті, що гальмують виділення гіпофізарних гормонів, – статинами. • На сьогоднішній день у гіпоталамусі відкрито 7 стимуляторів (ліберини) і 3 інгібітори (статини) секреції гормонів гіпофіза, зокрема: кортиколіберин, тиреоліберин, люліберин, фоліліберин, соматоліберин, пролактоліберин, меланоліберин, соматостатин, пролактостатин і меланостатин

ГОРМОНИ ГІПОТАЛАМУСУ
 впливає на")
• Рилізинг-гормони. Тироліберин стимулює секрецію жиротропного та лютеїнізувального гормону; люліберин (гонадоліберин) впливає на продукування лютеїнізувального та фолікулостимулювального гормонів; соматоліберин стимулює синтез і секрецію соматотропіну, кортиколіберин – кортикотропіну, меланоліберин – меланіну. • Інгібувальні гормони (статини). Соматостатин інгібує виділення гормону росту, інсуліну, глюкагону. Крім гіпоталамуса, його синтезують панкреатичні острівці. Пролактостатин гальмує продукування пролактину, але виявляє гонадоліберинову активність; меланостатин гальмує секрецію меланотропіну, впливає на коркові та підкоркові зони головного мозку. • Вазопресин та окситоцин. Ці гормони утворюються в гіпоталамусі у великих ядрах: у супраоптичних – вазопресин, у паравентрикулярних – окситоцин, і транспортуються у задню частку гіпофіза. За структурою ці гормони є нанопептидами, що утворюють за допомогою дисульфідного мостика шестикомпонентну петлю, а боковий ланцюг містить три амінокислоти. • Головна функція вазопресину – вплив на діурез, стінки кровоносних судин, а окситоцину – на гладку мускулатуру матки, молочні залози, сім’явивідні протоки.

Вазопресин і окситоцин • • Гормони вазопресин і окситоцин синтезуються рибосомальним шляхом. Одночасно у гіпоталамусі синтезуються 3 білки: нейрофізин I, II та III, функція яких полягає в нековалентному зв’язуванні окситоцину і вазопресину та транспорті цих гормонів в нейросекреторні гранули гіпоталамусу. Далі у вигляді комплексів нейрофізин–гормон вони мігрують уздовж аксону і досягають задньої частки гіпофізу, де відкладаються про запас; після дисоціації комплексу вільний гормон секретуєтсья в кров. Нейрофізини також виділені в чистому вигляді, і досліджена первинна структура двох із них (92 з 97 амінокислотних залишків); це багаті цистеїном білки, що містять по сім дисульфідних зв’язків. Хімічна будова обох пептидів була розшифрована у класичних роботах В. дю Віньо, який першим виділив ці гормони із задньої частки гіпофізу і здійснив їхній хімічний синтез. Обидва гормони є нонапептидами
 • Тиреоліберин є трипептидом, що склалається з піроглутамінової (циклічної) кислоти, гістидину")
Тиреоліберин (Піро-Глу–Гіс–Про–NH 2) • Тиреоліберин є трипептидом, що склалається з піроглутамінової (циклічної) кислоти, гістидину та пролінаміду, з’єднаних пептидними зв’язками. На відміну від класичних пептидів він не містить вільних NH 2 - та СООН-груп у N- та С-кінцевих амінокислотах.

Гонадоліберин Піро-Глу–Гіс–Трп–Сер–Тир–Глі–Лей–Арг–Про–Глі-NН 2 • Гонадоліберин є декапептидом, що складається з 10 амінокислот. Кінцева С-амінокислота є гліцинамідом.
. Крім гіпоталамуса соматостатин продукується")
Соматостатин • Соматостатин є циклічним тетрадекапептидом (з 14 амінокислотних залишків). Крім гіпоталамуса соматостатин продукується нейронами центральної і периферичної нервових систем, а також синтезується в S -клітинах панкреатичних острівків Лангерганса у підшлунковій залозі та клітинах кишечника. Він виявляє широкий спектр біологічної дії; а саме , показано інгібуючу дію на синтез гормону росту в аденогіпофізі, а також пряму інгібуючу дію на біосинтез інсулину і глюкагону в β- и α-клітинах острівків Лангерганса

Соматоліберин • Соматоліберин недавно виділений з природних джерел. Він склвдвється з 44 амінокислотних залишків, і його послідовність повністю розкрита. • Біологічною активністю володіє синтетичний декапептид. Цей декапептид стимулює синтез і секрецію гормону росту гіпофізу соматотропіну. Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН.

Меланоліберин Н-Цис–Тир–Иле–Глн–Асн–Цис-ОН. • Меланоліберин за хімічною структурою є аналогічним до відкритого кільця гормону окситоцину (без трипептидного бокового ланцюга )
 представлений або трипептидом, або пентапептидом. Необхідно відзначити, що меланоліберин")
Меланостатин • Меланостатин (меланотропінінгібуючий фактор) представлений або трипептидом, або пентапептидом. Необхідно відзначити, що меланоліберин виявляє стимулюючу дію , а меланостатин, навпаки, інгібуючу дію на синтез та секрецію меланотропіну в передній частці гіпофізу. Піро-Глу–Лей–Глі-NН 2, або Піро-Глу–Гіс–Фен–Aрг–Глі–NН 2.
. •")
ГОРМОНИ ГІПОФІЗУ • У гіпофізі виділяють три частки: передню, середню і задню (нейрогіпофіз). • У передній частці синтезується адренокортикотропний гормон (кортикотропін), пролактин, лютропін (лютеїнізувальний гормон), фолітропін (фолікулостимулювальний), соматотропін. Кортикотропін, окрім гіпофіза, може синтезуватися ще в інших тканинах, зокрема у плаценті, легенях (у разі патології). • У середній гормон. • частці синтезується меланоцитостимулювальний Нейрогіпофіз запасає і секретує окситоцин і вазопресин.

ГОРМОНИ ГІПОФІЗУ
 • • • Меланотропины синтезируются и секретируются в кровь промежуточной")
Меланоцитстимулюючі гормони (МСГ, меланотропіни) • • • Меланотропины синтезируются и секретируются в кровь промежуточной долей гипофиза. Выделены и расшифрованы первичные структуры двух типов гормонов – α- и β-меланоцитстимулирующие гормоны (α-МСГ и β-МСГ). Оказалось, что у всех обследованных животных α-МСГ состоит из 13 остатков аминокислот, расположенных в одинаковой последовательности. В α-МСГ N-концевой серин ацетилирован, а С-концевая аминокислота представлена валинамидом. Состав и структура β-МСГ оказались более сложными. У большинства животных молекула β-МСГ состоит из 18 остатков аминокислот; кроме того, имеются видовые различия, касающиеся природы аминокислоты в положениях 2, 6 и 16 полипептидной цепи гормона. β-МСГ, выделенный из промежуточной доли гипофиза человека, оказался 22 -членным пептидом, удлиненным на 4 аминокислотных остатка с N-конца: Физиологическая роль меланотропинов заключается в стимулировании меланиногенеза у млекопитающих и увеличении количества пигментных клеток (меланоцитов) в кожных покровах земноводных. Возможно также влияние МСГ на окраску меха и секреторную функцию сальных желез у животных. СН 3 -СО-NH-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз–Про–Вал-СО-NН 2 Нила–Глу–Лиз–Асп–Глу–Гли–Про–Тир–Aрг–Мет–Глу–Гис–Фен– –Арг–Трп –Гли–Сер–Про–Лиз–Асп-ОН
 • • Н-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз– Про–Вал–Гли––Лиз–Aрг–Про–Вал–Лиз–Вал– Тир–Про–Асп–Ала–Гли–Глу––Асп–Глн–Сер–Ала–Глу– Ала–Фен–Про–Лей–Глу–Фен-ОН Еще в 1926 г.")
Адренокортикотропний гормон (АКТГ, кортикотропін) • • Н-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз– Про–Вал–Гли––Лиз–Aрг–Про–Вал–Лиз–Вал– Тир–Про–Асп–Ала–Гли–Глу––Асп–Глн–Сер–Ала–Глу– Ала–Фен–Про–Лей–Глу–Фен-ОН Еще в 1926 г. было установлено, что гипофиз оказывает стимулирующее влияние на надпочечники, повышая секрецию гормонов коркового вещества. Накопленные к настоящему времени данные свидетельствуют, что этим свойством наделен АКТГ, вырабатываемый базофильными клетками аденогипофиза. АКТГ, помимо основного действия – стимуляции синтеза и секреции гормонов коры надпочечников, обладает жиромобилизующей и меланоцитстимулирующей активностью. Молекула АКТГ у всех видов животных содержит 39 аминокислотных остатков. Первичная структура АКТГ свиньи и овцы была расшифрована еще в 1954 – 1955 гг. Строение АКТГ человека:
 • Оказалось, что в ткани мозга и в промежуточной доле")
Ліпотропні гормоні (ЛТГ, ліпотропіни) • Оказалось, что в ткани мозга и в промежуточной доле гипофиза синтезируются биологически активные пептиды, наделенные опиатоподобным действием. Приводим структуры некоторых из них: • Общим типом структуры для всех трех соединений является тетра-пептидная последовательность на N-конце. Доказано, что β-эндорфин (31 АМК) образуется путем протеолиза из более крупного гипофизарного гормона β-липотропина (91 АМК); последний вместе с АКТГ образуется из общего предшественника – прогормона, названного проопиокортином (является, таким образом, препрогормоном), имеющим молекулярную массу 29 к. Да и насчитывающим 134 аминокислотных остатка. Биосинтез и освобождение проопиокортина в передней доле гипофиза регулируется кортиколиберином гипоталамуса. В свою очередь из АКТГ и β-липо-тропина путем дальнейшего процессинга, в частности ограниченного протеолиза, образуются соответственно αи βмеланоцитстимулирующие гормоны (α- и β-МСГ).
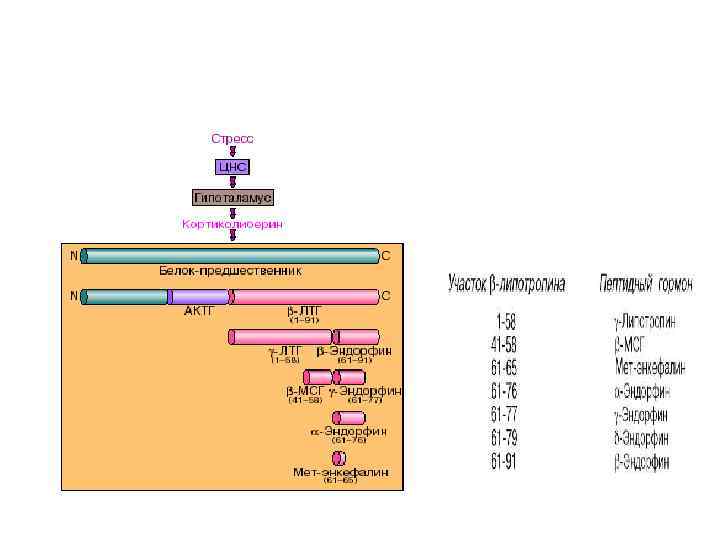
, точнее,")
Гормони паращитоподібних залоз • К гормонам белковой природы относится также паратиреоидный гормон (паратгормон), точнее, группа паратгормонов, различающихся последовательностью аминокислот. Они синтезируются паращитовидными железами. Еще в 1909 г. было показано, что удаление паращитовидных желез вызывает у животных тетанические судороги на фоне резкого падения концентрации кальция в плазм крови; введение солей кальция предотвращало гибель животных. Однако только в 1925 г. из паращитовидных желез был выделен активный экстракт, вызывающий гормональный эффект – повышение содержания кальция в крови. Чистый гормон был получен в 1970 г. из паращитовидных желез крупного рогатого скота; тогда же была определена его первичная структура. Выяснено, что паратгормон синтезируется в виде предшественника (115 аминокислотных остатков) пропарат-гормона, однако первичным продуктом гена оказался препропарат-гормон, содержащий дополнительно сигнальную последовательность из 25 аминокислотных остатков. Молекула паратгормона быка содержит 84 аминокислотных остатка и состоит из одной полипептидной цепи.

ГОРМОНИ ЩИТОПОДІБНОЇ ЗАЛОЗИ • В особых клетках – так называемых парафолликулярных клетках, или С-клетках щитовидной железы, синтезируется гормон пептидной природы, обеспечивающий постоянную концентрацию кальция в крови. Он получил название «кальцитонин» . Впервые на существование кальцитонина, обладающего способностью поддерживать постоянный уровень кальция в крови, указал в 1962 г. Д. Копп, который ошибочно считал, что этот гормон синтезируется паращитовидными железами. В настоящее время кальцитонин не только выделен в чистом виде из ткани щитовидной железы животных и человека, но и полностью раскрыта 32 -членная аминокислотная последовательность, подтвержденная химическим синтезом. Ниже приведена первичная структура кальцитонина, полученного из щитовидной железы человека: • Кальцитонин человека содержит дисульфидный мостик (между 1 -м и 7 -м аминокислотными остатками) и характеризуется N-концевым цистеином и С-концевым пролинамидом. Кальцитонины быка, овцы, свиньи и лососевых рыб мало отличаются друг от друга как по структуре и концевым аминокислотам, так и по гипокальциемической активности. Биологическое действие кальцитонина прямо противоположно эффекту паратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Таким образом, постоянство уровня кальция в крови человека и животных обеспечивается главным образом паратгормоном, кальцитриолом и кальцитонином, т. е. гормонами как щитовидной и паращитовидных желез, так и гормоном – производным витамина D 3. Это следует учитывать при хирургических лечебных манипуляциях на данных железах.

ГОРМОНИ ЩИТОПОДІБНОЇ ЗАЛОЗИ • Щитовидная железа состоит из множества особых полостей – фолликулов, заполненных вязким секретом – коллоидом. В состав коллоида входит особый йодсодержащий гликопротеин с высокой мол. массой – порядка 650000 (5000 аминокислотных остатков). Этот гликопротеин получил название йодтиреоглобулина. Он представляет собой запасную форму тироксина и трийодтиронина – основных гормонов фолликулярной части щитовидной железы.

Механізм дії тиреоїдних гормонів

ГОРМОНИ ПІДШЛУНКОВОЇ ЗАЛОЗИ Інсулін

Глюкагон Н–Гис–Сер–Глн–Гли–Тре–Фен–Тре–Сер–Асп–Тир–Сер–Лиз–Тир–Лей– –Асп– Сер–Aрг–Ала–Глн–Асп–Фен–Вал–Глн–Трп–Лей–Мет–Асн–Тре–ОН • Глюкагон впервые был обнаружен в коммерческих препаратах инсулина еще в 1923 г. , однако только в 1953 г. венгерский биохимик Ф. Штрауб получил этот гормон в гомогенном состоянии. Глюкагон синтезируется в основном в α -клетках панкреатических островков поджелудочной железы, а также в ряде клеток кишечника. По биологическому действию глюкагон, как и адреналин, относятся к гипергликемическим факторам, вызывает увеличение концентрации глюкозы в крови главным образом за счет распада гликогена в печени. Органами-мишенями для глюкагона являются печень, миокард, жировая ткань, но не скелетные мышцы. Биосинтез и секреция глюкагона контролируются главным образом концентрацией глюкозы по принципу обратной связи. Таким же свойством обладают аминокислоты и свободные жирные кислоты. На секрецию глюкагона оказывают влияние также инсулин и инсулиноподобные факторы роста.

ГОРМОНИ НАДНИРНИКІВ Гормони мозкового шару наднирників О способности экстрактов из надпочечников повышать кровяное давление было известно еще в XIX в. , однако только в 1901 г. Дж. Такамине и сотр. выделили из мозгового слоя надпочечников активное начало, идентифицированное с адреналином Это был первый гормон, полученный в чистом кристаллическом виде. Спустя более 40 лет, в 1946 г. , из мозгового вещества был выделен еще один гормон – норадреналин, который до этого был синтезирован химическим путем.

Гормоны коркового шару наднирників • К настоящему времени из коркового вещества надпочечников человека, свиньи и быка выделено около 50 различных соединений, которым дано общее название «кортикоиды» , или «кортикостероиды» . Общее число всех стероидов, которые синтезируются в надпочечниках многих животных, приближается к 100, однако биологической активностью наделены не все кортикостероиды.
. Жіночі")
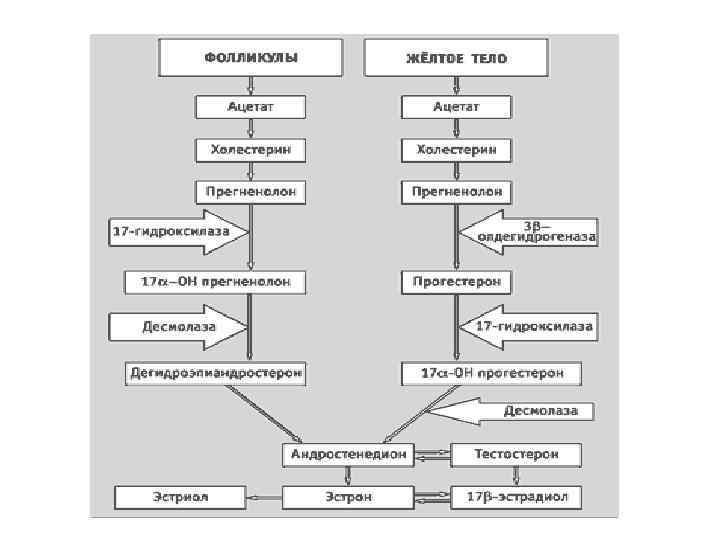
Статеві гормони • Статеві гормони утворюються в інтеростеціальних клітинах статевих залоз (сім’яники, яєчники). Жіночі статеві гормони – прогестини й естрогени – формуються у фолікулах яєчників, а чоловічі – андрогени – у клітинах Лейдіга. Крім того, ендокринним органом, що утворює гормони стероїдної природи, є плацента. • Жіночі статеві гормони є похідними естрану, який відрізняється від циклопентанпергідрофенантрену наявністю СН 3 – групи біля 13 -го Карбону. • Прогестерон і чоловічі статеві гормони є похідними андростану, який відрізняється від циклопентанпергідрофенантрену наявністю двох метильних груп у 10 -му та 13 -му положеннях.
 і")
Жіночі статеві гормони • До найважливіших жіночих статевих гормонів належать естрадіол, естрон (фолікулін) і естріол: Естрогени в нормі, головно, утворюються у внутрішній оболонці фолікулів, жовтому тілі, плаценті, менше – у сім’яниках, нейронах центральної нервової системи, а за умов патологій у великих кількостях їх виділяють надниркові залози.

Жіночі статеві гормони • Прогестерон – гормон жовтого тіла. • Його синтезують у нормі яєчники та плацента. Він регулює менструальний цикл, процес вагітності, є індуктором дозрівання овоцитів, стимулює імплантацію яйцеклітини тощо.

Чоловічі статеві гормони • Андрогени утворюються у клітинах сім’яників, але можуть також утворюватися і в яєчниках, плаценті, корі надниркових залоз. Основний андроген – тестостерон, але секретуються й інші – андростандіол, дегідроепіандростерон, андростендіон

Статеві гормони • Статеві гормони зумовлюють формування вторинних статевих ознак, а також реалізацію репродуктивної функції жіночих і чоловічих особин. • Естрогени впливають на біосинтез білка, нуклеїнових кислот, регулювання рівня Кальцію та Фосфору, циклічні явища в організмі самок. • Тестостерон, крім впливу на стимуляцію статевого апарату, виявляє потужний анаболічний ефект.

Механізм дії прогестерону на клітину-мішень

Steroids of the Adrenal Cortex

Gonadal Steroid Hormones

Synthesis of the major female sex hormones in the ovary. Synthesis of testosterone androstenedione from cholesterol occurs by the same pathways as indicated for synthesis of the male sex hormones.

Ейкозаноїди - похідні арахідонової кислоти Арахідонова кислота після вивільнення із фосфоліпідів біомембран под дією специфічних фосфоліпаз А (або С) в залежності від ферментативного шляху перетворення дає початок простаноїдам і лейкотрієнам за схемою:

Простагландини • Термін «простагландини» ввів У. Ейлер, який вперше показав, що у спермі людини і екстрактах із сім’янників барана містяться речовини, які виявляють виражену вазопресивну дію і викликають скорочення гладкої мускулатури матки. Припущення У. Ейлера, про те, що ці речовини є специфічним секретом передміхурової залози (prostate gland), не підтвердилося, оскільки, встановлено, що вони містяться у всіх органах і тканинах. Однак, запрпоновані ним терміни ( простатогландини, простагландини) в літературі зберігся.

Простагландини • Простагландини є предметом всебічних досліджень, з огляду на значне поширення у тканинах, виражену фармакологічну дію на різноманітні фізіологічні функції організму вплив на регуляцію гемодинаміки нирок, скоротливу функцію гладкої мускулатури, секреторну функцію шлунка, жировий, водно-сольовий обмін та ін. Існують думки про те, що простагландини не є „істинними" гормонами, хоча деякі автори вважають їх „локальними гормонами, або гормонами місцевої дії“. • Вони модулюють дію гормонів. Біологічні ефекти простагландинів, імовірно, опосередковуються через циклічні нуклеотиди.

Циклооксигеназний шлях перетворення арахідонової кислоти. • • Перший шлях перетворення арахідонової кислоти отримав назву циклооксигеназного шляху, оскільки перші стадії синтезу простагландинів каталізуються циклооксигеназою, точніше простагландинсинтазою (КФ 1. 14. 99. 1). Центральним хімічним процесом біосинтезу є включення кисню (двох молекул) у структуру арахідонової кислоти. Цей процес каталізується специфічними оксигеназами, які, окрім окиснення, каталізують циклізацію з утворенням проміжних продуктів – простагландинендопероксидів PG 2[H 2], які позначають PGG 2 і PGH 2; останні під впливом простагландинізомераз перетворюються в первинні простагландини. Розрізняють два класи первинних простагландинів: розчинні в ефірі простагландини PGE і розчинні у фосфатному буфері простагландини PGF. Кажен із класів ділиться на підкласи: PGE 1, PGE 2, PGF 1, PGF 2 і т. д. Простацикліни і тромбоксани синтезуються із вказаних проміжних продуктів за участі інших, відмінних від ізомераз ферментів.

Циклооксигеназний шлях перетворення арахідонової кислоти. R 1 та R 2 – бокові ланцюги, ідентичні для всіх простагландинів. Знаком мінус позначено блокуючу дію вказаних речовин.
, діють на гладку")
Простагландини • Первинні простагландини синтезуються у всіх клітинах (за винятком еритроцитів), діють на гладку мускулатуру травного тракту, репродуктивні та респіраторні тканини, на тонус судин, модулюють активність інших гормонів, автономно регулюють нервове збудження, запальні процеси (медіатори), швидкість ниркового кровотоку; їхня біологічна дія опосередковується шляхом регуляції синтезу ц. АМФ.
, синтезується в тканині")
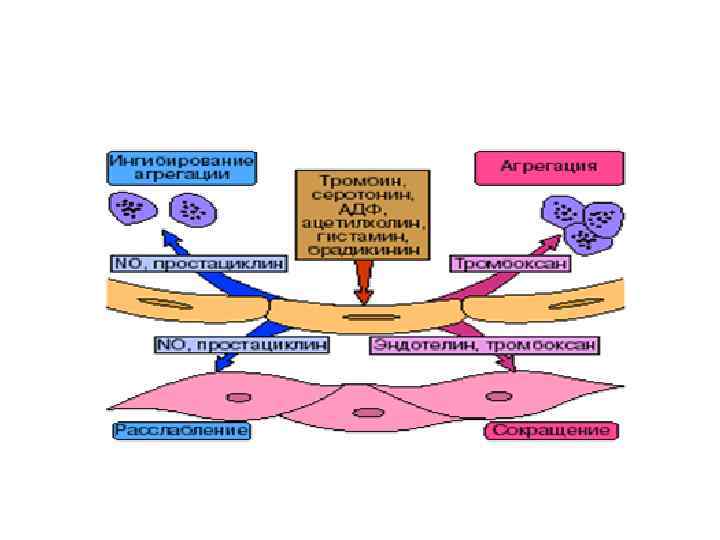
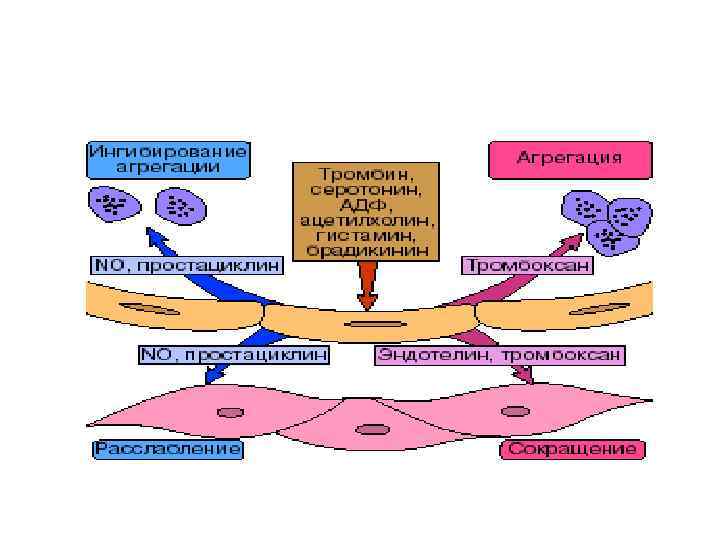
Тромбоксани • Тромбоксан А, зокрема тромбоксан А 2 (Тх. А 2), синтезується в тканині мозку, селезінки, легень, нирок, а також у тромбоцитах із PGH 2 під дією тромбоксансинтази. Решта тромбоксанів утворюються з Тх. А 2. Вони індукують агрегацію тромбоцитів, сприяючи тим самим тромбоутворенню, і, крім того, виявляють найбільш потужну судинозвужуючу дію поміж усіх простагландинів.
 синтезується переважно в ендотелії судин, серцевому м'язі, тканині матки")
Простациклін • Простациклін (PGI 2) синтезується переважно в ендотелії судин, серцевому м'язі, тканині матки і слизовій оболонці шлунка. Він, на противагу тромбоксанові, розслаблює гладком'язові волокна судин і викликає дезагрегацію тромбоцитів, сприяючи фібринолізу • Слід також вказати на важливе значення для фізіологічного статусу організму співвідношення у крові тромбоксани/простацикліни, а саме Tx. A 2/PGI 2. З'ясувалося, що у хворих, які мають схильність до тромбозів, є тенденція до зсуву балансу в бік агрегації; у хворих, які потерпають від уремії, навпаки, спостерігається дезагрегація тромбоцитів. Висунуто припущення про важливу роль балансу Tx. A 2/PGI 2 для регуляції функції тромбоцитів in vivo, серцево-судинного гомеостазу, тромботичної хвороби тощо.

Катаболізм простаноїдів • Початковою стадією катаболізму „класичних" простагландинів є стереоспецифічне окиснення ОН-групи біля 15 -го атома Карбону з утворенням відповідного 15 -кетопохідного. Фермент, який каталізує цю реакцію, - 15 -оксипростагландинде-гідрогеназа - відкритий у цитоплазмі, потребує НАД чи НАДФ. Тромбоксан інактивується in vivo або шляхом хімічного розщепления до тромбоксану В 2, або шляхом окислення дегідрогеназою чи редуктазою. Аналогічно PGI 2 (простациклін) швидко розпадаєтья до 6 -кето-PGFlα in vitro, a in vivo інактивується окисненням 15 окси-простагландиндегідрогеназою з утворенням 6, 15 -дикето. РGF 1 α.

Ліпооксигеназний шлях перетворення арахідонової кислоти • Другий шлях перетворення арахідонової кислоти - ліпооксигеназний шлях - відрізняється тим, що започатковує синтез іще одного класу біологічно активних сполук - лейкотрієнів. Характерна особливість структури лейкотрієнів полягає в тому, що вони не мають циклічної структури, хоча лейкотрієни, як і простаноїди, побудовані з 20 атомів Карбону. У структурі лейкотрієнів міститься чотири подвійні зв'язки, деякі з них утворюють пептидоліпідні комплекси з глутатіоном або з його складовими частинами (лейкотрієн D може перетворюватися в лейкотрієн Е, втрачаючи залишок гліцину).

Лейкотрієни • Основні біологічні ефекти лейкотрієнів пов'язані з запальними процесами, алергічними й імунними реакціями, анафілаксією та діяльністю гладких м'язів. • Зокрема, лейкотрієни сприяють скороченню гладкої мускулатури дихальних шляхів, травного тракту, регулюють тонус судин (виявляють судинозвужуючу дію) і стимулюють скорочення коронарних артерій. • Катаболічні шляхи лейкотрієнів до кінця не з'ясовані. • Таким чином, завдяки широкому розповсюдженню у тканинах, а також високій і всебічній біологічній активності простагландини (і простаноїди взагалі) та лейкотрієни широко застосовуються у медичній практиці як лікарські препарати.

Ліпооксигеназний шлях перетворення арахідонової кислоти R – акцептор залишку глутамінової кислоти. Знаком мінус позначена блокуюча дія вітаміну Е.


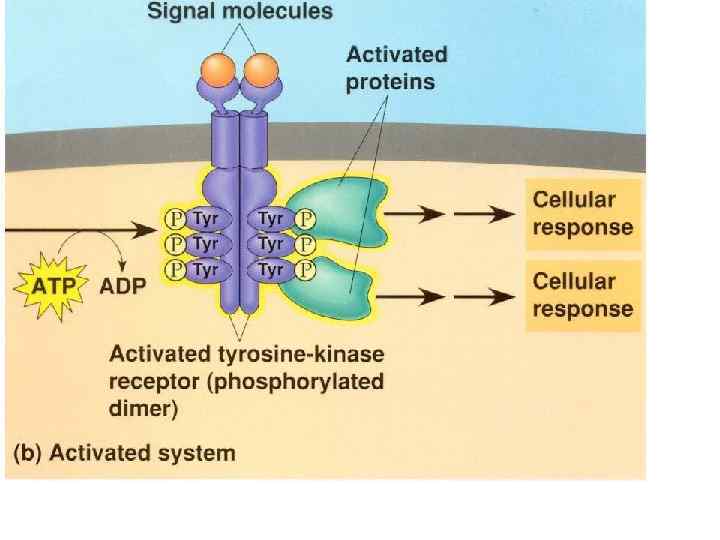
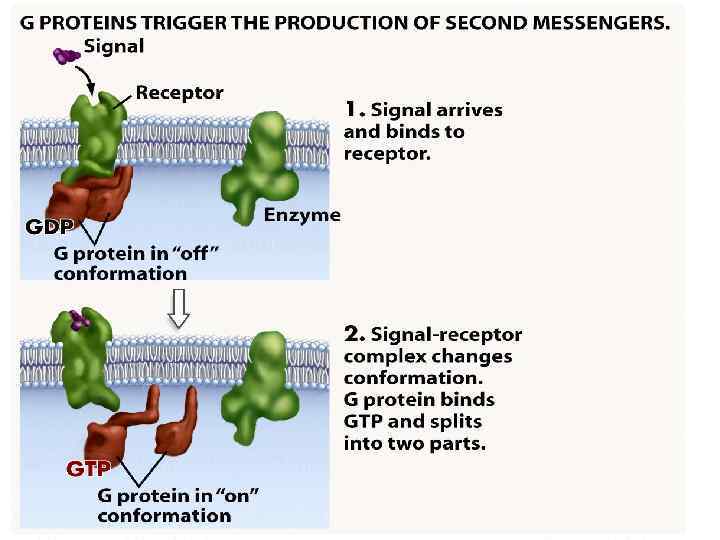
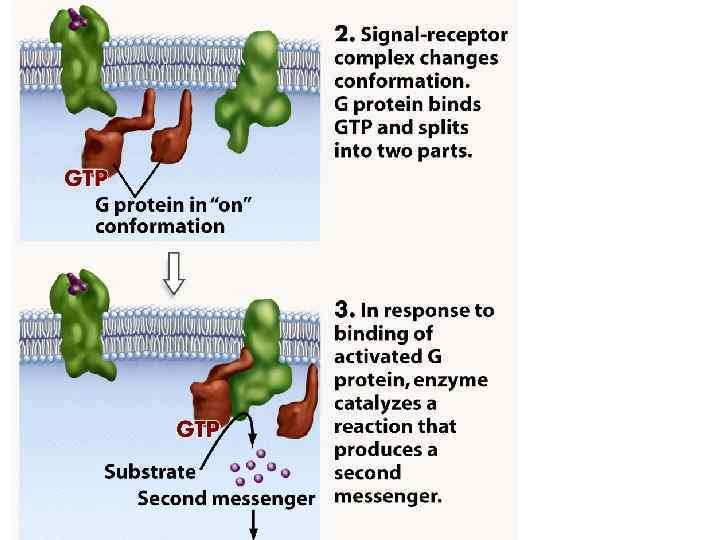
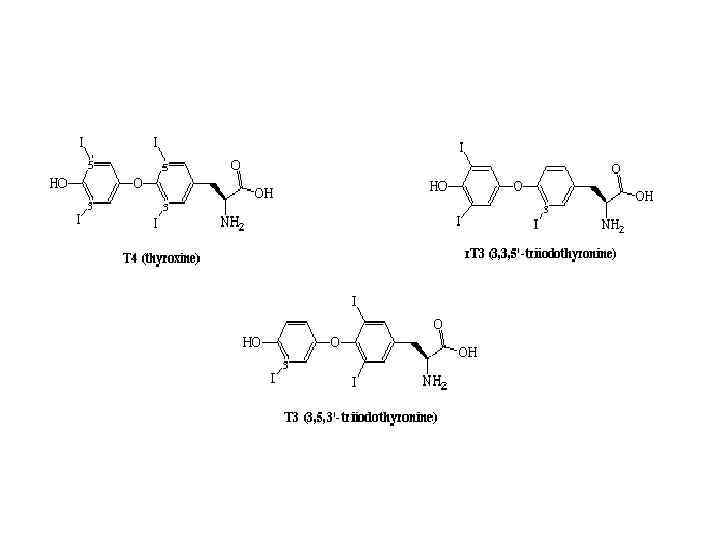

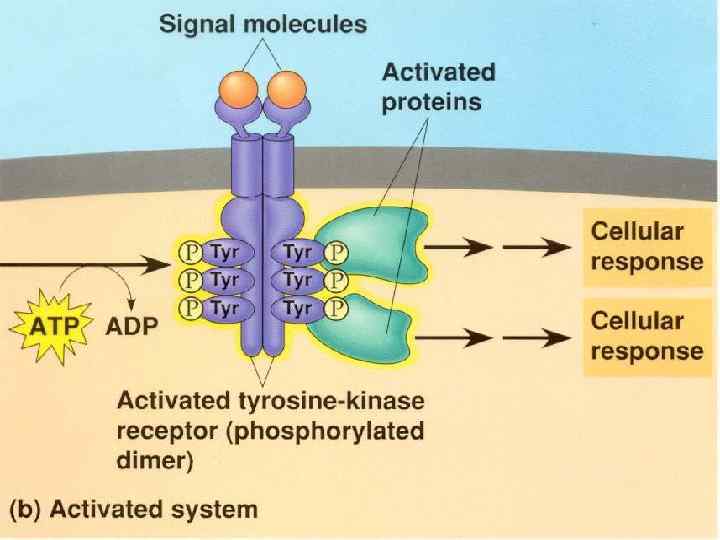
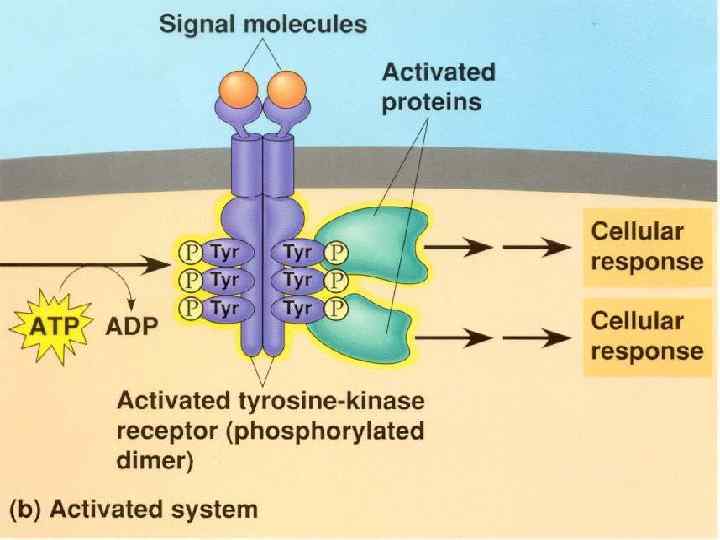
Figure 22 -31 The insulin receptor consists of two α chains located on the outer face of the plasma membrane and two β chains that traverse the membrane and protrude on the cytosolic face. Binding of insulin to the α chains triggers autophosphorylation of Tyr residues in the carboxyl-terminal domain of the β subunits, which allows the tyrosine kinase domain to catalyze phosphorylation of other target proteins.

Аденілатциклазна система

Нейрогуморальна регуляція

Механізм дії гормонів на клітинному рівні

Аденілатциклазна система

Аденілатциклазна система
Гормони_p.ppt