

13Ауксин Bio.ppt
- Количество слайдов: 71

 § Гормональная § Электрическая § Трофическая
§ Гормональная § Электрическая § Трофическая
 Гормоны растений – эндогенные регуляторы программ развития • • Критерии гормонов: Вещество вызывает специфический физиологический ответ у определенных клеток. Разобщено место синтеза и место действия, т. е. необходим транспорт сигнального вещества по растению (для растений - не строгий критерий) Вещество практически не играет роли в основном метаболизме клетки, используется только для сигнальных целей. Вещество должно действовать в низкой концентрации (начиная с 10 -5 моль/л). Особенности фитогормонов: Низкомолекулярные Регулируют целые программы развития В принципе могут синтезироваться любой клеткой растения Взаимное действие – «Переговоры гормонов» .
Гормоны растений – эндогенные регуляторы программ развития • • Критерии гормонов: Вещество вызывает специфический физиологический ответ у определенных клеток. Разобщено место синтеза и место действия, т. е. необходим транспорт сигнального вещества по растению (для растений - не строгий критерий) Вещество практически не играет роли в основном метаболизме клетки, используется только для сигнальных целей. Вещество должно действовать в низкой концентрации (начиная с 10 -5 моль/л). Особенности фитогормонов: Низкомолекулярные Регулируют целые программы развития В принципе могут синтезироваться любой клеткой растения Взаимное действие – «Переговоры гормонов» .
 Где находится светочувствительная часть растения? апекс зона изгиба Вывод: Из апекса в зону изгиба идет сигнал. Гипотеза: Сигнал – это вещество (ауксин)
Где находится светочувствительная часть растения? апекс зона изгиба Вывод: Из апекса в зону изгиба идет сигнал. Гипотеза: Сигнал – это вещество (ауксин)
 2. Электрический сигнал 3. Механический сигнал") Какова природа сигнала? Гипотезы: 1. Химический сигнал (ауксин) 2. Электрический сигнал 3. Механический сигнал 4. Светооптический сигнал апекс агар Вывод: Сигнал – это вещество (ауксин)
Какова природа сигнала? Гипотезы: 1. Химический сигнал (ауксин) 2. Электрический сигнал 3. Механический сигнал 4. Светооптический сигнал апекс агар Вывод: Сигнал – это вещество (ауксин)
 Многообразие акусинов Для табака было показано, что треть всей ИУК находится в хлоропластах, две трети – в цитозоле. Триптофан – предшественник ИУК – образуется в хлоропластах Индолилуксусная кислота ИУК наиглавнейший ауксин Индолилмасляная кислота (ИМК) – у горчицы и пшеницы, хлоро-индолилуксусная (горох), фенилуксусная (ФУК) кислоты – природные соединения, обладающие ауксиновой активностью. Присутствуют не у всех
Многообразие акусинов Для табака было показано, что треть всей ИУК находится в хлоропластах, две трети – в цитозоле. Триптофан – предшественник ИУК – образуется в хлоропластах Индолилуксусная кислота ИУК наиглавнейший ауксин Индолилмасляная кислота (ИМК) – у горчицы и пшеницы, хлоро-индолилуксусная (горох), фенилуксусная (ФУК) кислоты – природные соединения, обладающие ауксиновой активностью. Присутствуют не у всех
 Биосинтез и деградация акусинов Триптофан Нетриптофановый путь Индолилуксусная кислота Окисление: Депонирование: ИУК-оксидаза ИУК-гликозиды Комплексы с белками Полифенолоксидаза И аминокислотами
Биосинтез и деградация акусинов Триптофан Нетриптофановый путь Индолилуксусная кислота Окисление: Депонирование: ИУК-оксидаза ИУК-гликозиды Комплексы с белками Полифенолоксидаза И аминокислотами
 Синтез акусинов – много вариантов… Разные пути могут быть в разных тканях, на разных этапах онтогенеза… Триптофаннезависимый Из триптофана TAM и IPA – основные пути образования ИУК у растений IAN – у Brassicaceae, Poaceae, Muscaceae и ряда других… IAM – у бактерий
Синтез акусинов – много вариантов… Разные пути могут быть в разных тканях, на разных этапах онтогенеза… Триптофаннезависимый Из триптофана TAM и IPA – основные пути образования ИУК у растений IAN – у Brassicaceae, Poaceae, Muscaceae и ряда других… IAM – у бактерий
 Низкомолекулярные конъюгаты – с глюкозой, инозитолом, кислотами, -СН 3") Обратимая инактивация ауксинов (образование конъюгатов) Низкомолекулярные конъюгаты – с глюкозой, инозитолом, кислотами, -СН 3 Значительная часть ауксинов образует неактивные конъюгаты, которые хранятся в запасающих органах и семенах Высокомолекулярные С глюканами (7– 50 глюкозных остатков) С пептидами С гликопротеинами
Обратимая инактивация ауксинов (образование конъюгатов) Низкомолекулярные конъюгаты – с глюкозой, инозитолом, кислотами, -СН 3 Значительная часть ауксинов образует неактивные конъюгаты, которые хранятся в запасающих органах и семенах Высокомолекулярные С глюканами (7– 50 глюкозных остатков) С пептидами С гликопротеинами
 Катаболизм акусинов Катаболизм – декарбоксилирующий за счет работы пероксидазы, ИУК-оксидазы
Катаболизм акусинов Катаболизм – декарбоксилирующий за счет работы пероксидазы, ИУК-оксидазы
 Рецепторы ауксинов – экстраили внутриклеточные? 1. ABP 1 Около 22 к. Da. У кукурузы – 5 генов. На N-конце -сигнал для транспорта в ЭР (38 а-к), на С-конце – КDEL, на N- конце – сайт связывания с ИУК: His-Arg-His-Ser-Cys-Glu. Сайт гликозилирования (Asp 33 -Thr), До 90% белка - в ЭР. 2. Транспортеры ИУК в клетку (4 гена - AUX 1 и др. ) из клетки (8 генов – PIN 1, PIN 2 и др). 3. ABP 60 k. Da ? Гликозидаза цитокининовых конъюгатов. Взаимодействие фитогормонов? 4. ABP 24 -25 k. Da Глютатион-5 -трансфераза (GST). GST контролирует уровень глутатиона, влияет на уровень жасмонатов, перекиси водорода. Множественность рецепторов ИУК – особый гормон: специфичный полярный транспорт (от клетки к клетке), эндогенный ритм образования и транспорта.
Рецепторы ауксинов – экстраили внутриклеточные? 1. ABP 1 Около 22 к. Da. У кукурузы – 5 генов. На N-конце -сигнал для транспорта в ЭР (38 а-к), на С-конце – КDEL, на N- конце – сайт связывания с ИУК: His-Arg-His-Ser-Cys-Glu. Сайт гликозилирования (Asp 33 -Thr), До 90% белка - в ЭР. 2. Транспортеры ИУК в клетку (4 гена - AUX 1 и др. ) из клетки (8 генов – PIN 1, PIN 2 и др). 3. ABP 60 k. Da ? Гликозидаза цитокининовых конъюгатов. Взаимодействие фитогормонов? 4. ABP 24 -25 k. Da Глютатион-5 -трансфераза (GST). GST контролирует уровень глутатиона, влияет на уровень жасмонатов, перекиси водорода. Множественность рецепторов ИУК – особый гормон: специфичный полярный транспорт (от клетки к клетке), эндогенный ритм образования и транспорта.
 Рецептор ауксина АВР-1 Гидрофильный гликопротеид с молекулярной массой 22 к. Да, формирующий гомодимер. ABP-1 (Auxin-Binding Protein 1). На С-конце ABP-1 имеется специфическая последовательность, состоящая из 4 -х аминокислот: лиз асп глу лей (KDEL), характерная для белков эндоплазматического ретикулума. У кукурузы выявлено пять АВР-1 генов, у арабидопсиса только один. Однако установить реальную связь между АВР 1 и ауксин-зависимой активацией генов пока не удалось.
Рецептор ауксина АВР-1 Гидрофильный гликопротеид с молекулярной массой 22 к. Да, формирующий гомодимер. ABP-1 (Auxin-Binding Protein 1). На С-конце ABP-1 имеется специфическая последовательность, состоящая из 4 -х аминокислот: лиз асп глу лей (KDEL), характерная для белков эндоплазматического ретикулума. У кукурузы выявлено пять АВР-1 генов, у арабидопсиса только один. Однако установить реальную связь между АВР 1 и ауксин-зависимой активацией генов пока не удалось.
 Существует как минимум два рецептора аукси Один из них активирует убиквитинировани Рецептор ауксина TIR 1 (transport inhibitor response 1) имеет F-box мотив (около 50 аминокислот для белокбелковых взаимодействий) и LRR участок. Он является компонентом специфичного E 3 -убиквитин-лигазного комплекса SCFTIR 1. Когда ауксин связывается с SCFTIR 1 то он активирует его, в результате происходит убиквитинирование и протеолитическая деградация AUX/IAA репрессорных белков.
Существует как минимум два рецептора аукси Один из них активирует убиквитинировани Рецептор ауксина TIR 1 (transport inhibitor response 1) имеет F-box мотив (около 50 аминокислот для белокбелковых взаимодействий) и LRR участок. Он является компонентом специфичного E 3 -убиквитин-лигазного комплекса SCFTIR 1. Когда ауксин связывается с SCFTIR 1 то он активирует его, в результате происходит убиквитинирование и протеолитическая деградация AUX/IAA репрессорных белков.
 короткоживущих ядерных") Система трансдукции ауксинового сигнала: AUX/IAA - ARF. Aux/IAA – смейство (более 25) короткоживущих ядерных ауксин-индуцируемых белков. ARFs - Auxin-respons factor (ауксиновые факторы транскрипции). Более 20 белков. Aux/IAA – белки образуют разнообразные димеры как между собой, так и с ARF. Разные димеры регулируют транскрипцию генов с Aux. RE (Auxin-respons element - TGTCNC). Димеры ARF – активируют, Aux/IAA+ARF – тормозят. Ауксин одновременно активирует транскрипцию Aux/IAA и их распад путем убиквитирования и деградации в 26 S протеосоме.
Система трансдукции ауксинового сигнала: AUX/IAA - ARF. Aux/IAA – смейство (более 25) короткоживущих ядерных ауксин-индуцируемых белков. ARFs - Auxin-respons factor (ауксиновые факторы транскрипции). Более 20 белков. Aux/IAA – белки образуют разнообразные димеры как между собой, так и с ARF. Разные димеры регулируют транскрипцию генов с Aux. RE (Auxin-respons element - TGTCNC). Димеры ARF – активируют, Aux/IAA+ARF – тормозят. Ауксин одновременно активирует транскрипцию Aux/IAA и их распад путем убиквитирования и деградации в 26 S протеосоме.
 Работа внутриклеточного рецептора ауксина. А. При низких концентрациях ауксина, транскрипция аусинрегулируемых генов, активирующихся ARFбелками блокирована реперессорами транскрипции Aux/IAA. В. При высоких концентрациях ауксин связывается с белком TIR 1, или с другими AFB белками, которые входят в SCFTIR 1 комплекс (RBX 1 -Cullin-ASK 1 TIR 1) убиквитирования. Связывание ауксина стимулирует взаимодействие Aux/IAAs с SCFTIR 1 что ведет к убиквитированию Aux/IAAs белков, и их разрушению в 26 S протеасоме. Это снимает ингибирование ARFs и запускает транскрипцию
Работа внутриклеточного рецептора ауксина. А. При низких концентрациях ауксина, транскрипция аусинрегулируемых генов, активирующихся ARFбелками блокирована реперессорами транскрипции Aux/IAA. В. При высоких концентрациях ауксин связывается с белком TIR 1, или с другими AFB белками, которые входят в SCFTIR 1 комплекс (RBX 1 -Cullin-ASK 1 TIR 1) убиквитирования. Связывание ауксина стимулирует взаимодействие Aux/IAAs с SCFTIR 1 что ведет к убиквитированию Aux/IAAs белков, и их разрушению в 26 S протеасоме. Это снимает ингибирование ARFs и запускает транскрипцию
 Рецепция акусинового сигнала: ядерный рецептор
Рецепция акусинового сигнала: ядерный рецептор
 Репрессор Aux/IAA формирует гетеродимер с ARF, что блокирует экспрессию ауксин-зависимых генов. Белок F-бокса TIR 1 метит ауксиновый репрессор Aux/IAA для убиквитинирования и последующего протеолиза. После снятия репрессии гомодимеры или мономеры ARF обеспечивают трансляцию ИУК-регулируемых генов.
Репрессор Aux/IAA формирует гетеродимер с ARF, что блокирует экспрессию ауксин-зависимых генов. Белок F-бокса TIR 1 метит ауксиновый репрессор Aux/IAA для убиквитинирования и последующего протеолиза. После снятия репрессии гомодимеры или мономеры ARF обеспечивают трансляцию ИУК-регулируемых генов.
 и «глобального» (TIR 1)") Принципиальная схема работы двух рецепторов ауксинов – «быстрого» (АВР 1) и «глобального» (TIR 1)
Принципиальная схема работы двух рецепторов ауксинов – «быстрого» (АВР 1) и «глобального» (TIR 1)
 Активный выход PIN 1") Транспорт ауксинов AUX 1 Пассивный вход Кортикальный цитоскелет (актиновые филаменты) Активный выход PIN 1
Транспорт ауксинов AUX 1 Пассивный вход Кортикальный цитоскелет (актиновые филаменты) Активный выход PIN 1
 Транспорт ауксинов AUX 1 Активный выход PIN 1 Пассивный вход From D. Reinhardt et al. , 2003
Транспорт ауксинов AUX 1 Активный выход PIN 1 Пассивный вход From D. Reinhardt et al. , 2003
 Транспорт ауксинов AUX 1 в корне: колумелла, корневой чехлик и стела. . Полярный транспорт ауксинов. Вход ауксинов клетку – комбинация двух механизмов Пассивная диффузия в протонированной форме (IAAH) через любую фосфолипидную мембрану. Вторично активный транспорт в диссоциированной форме – IAA- - 2 H+ -симпортер. Переносчик ауксина в клетку – AUX 1. Похож на бактериальный переносчик аминокислот. Обнаружен в проводящих тканях листа и корневом апексе.
Транспорт ауксинов AUX 1 в корне: колумелла, корневой чехлик и стела. . Полярный транспорт ауксинов. Вход ауксинов клетку – комбинация двух механизмов Пассивная диффузия в протонированной форме (IAAH) через любую фосфолипидную мембрану. Вторично активный транспорт в диссоциированной форме – IAA- - 2 H+ -симпортер. Переносчик ауксина в клетку – AUX 1. Похож на бактериальный переносчик аминокислот. Обнаружен в проводящих тканях листа и корневом апексе.
 Транспорт ауксинов
Транспорт ауксинов
 Транспорт ауксинов Мутация pin 1
Транспорт ауксинов Мутация pin 1
 Транспорт ауксинов Выход ауксинов из клетки В цитозоле р. Н 7, 2, ауксин в диссоциированной форме, поэтому он концентрируется в клетке (так как IAA- плохо проходит через мембрану). Выход – семейство переносчиков PIN (название от pin-shaped inflorescences – булавковидное соцветие, по мутанту PIN 1) Они располагаются как и предсказано – по линии транспорта ауксинов. Разные белки – в разных тканях. Работа PIN связана с другими белками – регуляторными и транспортными. В частности, АТФ-зависимые переносчики, принадлежащие к семейству MDR/PGR белков (multidrug resistance/P-glycoprotein PGPs) -. Это интегральные белки – анионные транспортеры. У арабидопсиса – 21, у риса – 17 PGP. Три PGPs – это транспортеры ауксина. PIN и PGPs действуют синергично. ИУК, формирующийся в листьях, транспортируется также неполярно по флоэме. При этом скорость транспорта выше, чем полярного. Загрузка и разгрузка – переносчиками. При этом неполярный транспорт видимо важен для камбиального деления клеток, формирования каллозы в ситовидных трубках и формирования боковых корней. Он может вносить значительный вклад в поступление ауксина в корни. PIN 1 – вертикальный транспорт ИУК от побега к корню PIN 3 – возврат ауксина в сосудистую паренхиму
Транспорт ауксинов Выход ауксинов из клетки В цитозоле р. Н 7, 2, ауксин в диссоциированной форме, поэтому он концентрируется в клетке (так как IAA- плохо проходит через мембрану). Выход – семейство переносчиков PIN (название от pin-shaped inflorescences – булавковидное соцветие, по мутанту PIN 1) Они располагаются как и предсказано – по линии транспорта ауксинов. Разные белки – в разных тканях. Работа PIN связана с другими белками – регуляторными и транспортными. В частности, АТФ-зависимые переносчики, принадлежащие к семейству MDR/PGR белков (multidrug resistance/P-glycoprotein PGPs) -. Это интегральные белки – анионные транспортеры. У арабидопсиса – 21, у риса – 17 PGP. Три PGPs – это транспортеры ауксина. PIN и PGPs действуют синергично. ИУК, формирующийся в листьях, транспортируется также неполярно по флоэме. При этом скорость транспорта выше, чем полярного. Загрузка и разгрузка – переносчиками. При этом неполярный транспорт видимо важен для камбиального деления клеток, формирования каллозы в ситовидных трубках и формирования боковых корней. Он может вносить значительный вклад в поступление ауксина в корни. PIN 1 – вертикальный транспорт ИУК от побега к корню PIN 3 – возврат ауксина в сосудистую паренхиму
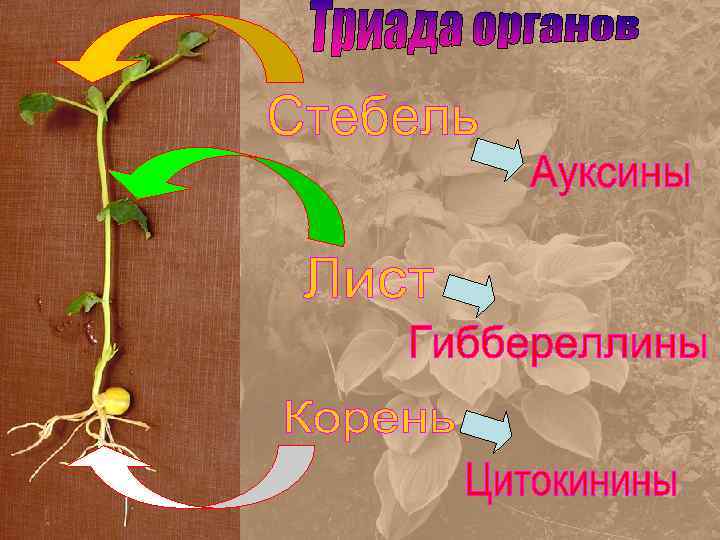
 Карта основных эффектов Аттрагирующий эффект Рост растяжением: Тропизмы – неравномерный рост Дифференцировка прокамбия и ксилемы (Аттрагирующий эффект) Апикальное доминирование (Конкуренция между главным побегом и боковыми) Деление клеток камбия и дифференцировка ксилемы (Аттрагирующий эффект) Ризогенез – образование боковых и придаточных корней (Аттрагирующий эффект)
Карта основных эффектов Аттрагирующий эффект Рост растяжением: Тропизмы – неравномерный рост Дифференцировка прокамбия и ксилемы (Аттрагирующий эффект) Апикальное доминирование (Конкуренция между главным побегом и боковыми) Деление клеток камбия и дифференцировка ксилемы (Аттрагирующий эффект) Ризогенез – образование боковых и придаточных корней (Аттрагирующий эффект)
 Роль ауксинов в закладке листа PIN 1 From D. Reinhardt et al. , 2003
Роль ауксинов в закладке листа PIN 1 From D. Reinhardt et al. , 2003
 Роль ауксинов в закладке листа У мутантов по гену pin 1 листья развиваются только в месте прикладывания блоков с ауксином From D. Reinhardt et al. , 2003
Роль ауксинов в закладке листа У мутантов по гену pin 1 листья развиваются только в месте прикладывания блоков с ауксином From D. Reinhardt et al. , 2003
 Роль ауксинов в закладке листа
Роль ауксинов в закладке листа
 Лирическое отступление
Лирическое отступление
 Полисахариды клеточной стенки построены всего из 11 сахаров
Полисахариды клеточной стенки построены всего из 11 сахаров
 Строение микрофибрилл целлюлозы
Строение микрофибрилл целлюлозы
 Строение целлюлозо-синтазы
Строение целлюлозо-синтазы
 Электронные фотографии КС с целлюлозо-синтазой
Электронные фотографии КС с целлюлозо-синтазой
 Ксилоглюканы (Xy. Gs) Фуко-Xy. Gs XXXG: XXFG (двудольные, некоммелиноидн. )") Сшивочные гликаны (cross-linking glycans) Ксилоглюканы (Xy. Gs) Фуко-Xy. Gs XXXG: XXFG (двудольные, некоммелиноидн. ) Гликаны со смешанной связью (злаки) Арабино-Xy. Gs AXGG, XAGG, AAGG Пасленовые, мята Нерегулярные Xy. Gs (коммелиноидные) Глюкуроноарабиноксиланы (GAXs) Коммелиноидные Ara: O-3, Glc. A: O-2 Некоммелин. Ara, Glc. A: O-2 Обозначения: G: Gl X: Gl-Xyl L: Gl-Xyl-Gal F: Gl-Xyl-Gal-Fuc A: Gl-Xyl-Ara
Сшивочные гликаны (cross-linking glycans) Ксилоглюканы (Xy. Gs) Фуко-Xy. Gs XXXG: XXFG (двудольные, некоммелиноидн. ) Гликаны со смешанной связью (злаки) Арабино-Xy. Gs AXGG, XAGG, AAGG Пасленовые, мята Нерегулярные Xy. Gs (коммелиноидные) Глюкуроноарабиноксиланы (GAXs) Коммелиноидные Ara: O-3, Glc. A: O-2 Некоммелин. Ara, Glc. A: O-2 Обозначения: G: Gl X: Gl-Xyl L: Gl-Xyl-Gal F: Gl-Xyl-Gal-Fuc A: Gl-Xyl-Ara
") Гемицеллюлозы: ксилоглюкан двудольных (фуко-галакто-Xy. Gs)
Гемицеллюлозы: ксилоглюкан двудольных (фуко-галакто-Xy. Gs)
") Гемицеллюлозы: ксилоглюкан двудольных (арабино-Xy. Gs пасленовых и мяты)
Гемицеллюлозы: ксилоглюкан двудольных (арабино-Xy. Gs пасленовых и мяты)
 Гемицеллюлозы: глюкуроно-арабиноксиланы двудольных и коммелиноидов
Гемицеллюлозы: глюкуроно-арабиноксиланы двудольных и коммелиноидов
 Гемицеллюлозы: глюкан злаковых
Гемицеллюлозы: глюкан злаковых
 Пектины Галактуронаны Гомогалактуронаны Ксилогалактуронаны Рамногалактуронаны II Рамногалактуронаны I
Пектины Галактуронаны Гомогалактуронаны Ксилогалактуронаны Рамногалактуронаны II Рамногалактуронаны I
") Пектины: галактоктуронаны (гомо- и ксило-галактуронаны)
Пектины: галактоктуронаны (гомо- и ксило-галактуронаны)
") Петкины: рамногалактуронаны II (к цепи гомогалактуронана присоединены различные боковые цепочки по разным гидроксилам )
Петкины: рамногалактуронаны II (к цепи гомогалактуронана присоединены различные боковые цепочки по разным гидроксилам )
 Пектины: рамногалактуронаны I гетерополимер: линейная цепь из чередующихся остатков Gal. A и Rha с различными боковыми фрагментами)
Пектины: рамногалактуронаны I гетерополимер: линейная цепь из чередующихся остатков Gal. A и Rha с различными боковыми фрагментами)
 Пектины: димер рамногалактуронана II (мономеры RGII 4200 k. Da связаны диэфирными связями остатками апиозы через бор)
Пектины: димер рамногалактуронана II (мономеры RGII 4200 k. Da связаны диэфирными связями остатками апиозы через бор)
 «Замковые зоны» пектиновой сети
«Замковые зоны» пектиновой сети
 Пектины: зоны «Ca 2+-застежек» и количество нейтральных боковых цепочек RGI регулируют размер пор клеточной стенки
Пектины: зоны «Ca 2+-застежек» и количество нейтральных боковых цепочек RGI регулируют размер пор клеточной стенки
") Структурные белки клеточной стенки: HGRPs, PRPs, GRPs (гидроксипролин-, пролин- и глицин- обогащенные)
Структурные белки клеточной стенки: HGRPs, PRPs, GRPs (гидроксипролин-, пролин- и глицин- обогащенные)
.") Структурные белки клеточной стенки: AGPs (арабино-галактановые белки - протеогликаны).
Структурные белки клеточной стенки: AGPs (арабино-галактановые белки - протеогликаны).
.") Структурные белки клеточной стенки: AGPs (арабино-галактановые белки - протеогликаны).
Структурные белки клеточной стенки: AGPs (арабино-галактановые белки - протеогликаны).
 и тип II (коммелиноиды)") Трехмерная модель двух типов клеточной стенки: тип I (двудольные) и тип II (коммелиноиды)
Трехмерная модель двух типов клеточной стенки: тип I (двудольные) и тип II (коммелиноиды)
![ИУК РЕЦЕПТОРЫ УВЕЛИЧЕНИЕ АКТИВАЦИЯ - АВР -1, TIR-1 [Са 2+]- зависимой В ЦИТОЗОЛЕ ПРОТЕИНКИНАЗЫ](https://present5.com/presentation/112200768_352222206/image-51.jpg "ИУК РЕЦЕПТОРЫ УВЕЛИЧЕНИЕ АКТИВАЦИЯ - АВР -1, TIR-1 [Са 2+]- зависимой В ЦИТОЗОЛЕ ПРОТЕИНКИНАЗЫ") ИУК РЕЦЕПТОРЫ УВЕЛИЧЕНИЕ АКТИВАЦИЯ - АВР -1, TIR-1 [Са 2+]- зависимой В ЦИТОЗОЛЕ ПРОТЕИНКИНАЗЫ ФОСФОРИЛИРОВАНИЕ Н+ АТФ азы НА ПЛАЗМАЛЕММЕ ПО ОСТАТКУ СЕРИНА СВЯЗЫВАНИЕ РЕГУЛЯТОРНОГО БЕЛКА АКТИВАЦИЯ Н+ АТФ азы ПОДКИСЛЕНИЕ КЛЕТОЧНОЙ СТЕНКИ РОСТ РАСТЯЖЕНИЕМ 14 -3 -3 И
ИУК РЕЦЕПТОРЫ УВЕЛИЧЕНИЕ АКТИВАЦИЯ - АВР -1, TIR-1 [Са 2+]- зависимой В ЦИТОЗОЛЕ ПРОТЕИНКИНАЗЫ ФОСФОРИЛИРОВАНИЕ Н+ АТФ азы НА ПЛАЗМАЛЕММЕ ПО ОСТАТКУ СЕРИНА СВЯЗЫВАНИЕ РЕГУЛЯТОРНОГО БЕЛКА АКТИВАЦИЯ Н+ АТФ азы ПОДКИСЛЕНИЕ КЛЕТОЧНОЙ СТЕНКИ РОСТ РАСТЯЖЕНИЕМ 14 -3 -3 И
 и экспансина в росте клеток растяжением") Возможное участие ХЕТ (ксилоглюкан-эндотрансгликозилазы) и экспансина в росте клеток растяжением
Возможное участие ХЕТ (ксилоглюкан-эндотрансгликозилазы) и экспансина в росте клеток растяжением
 Роль акусинов в развитии проводящей системы
Роль акусинов в развитии проводящей системы
 ФОТОТРОПИЗМ Апекс побега Зона изгиба Изгиб невозможен
ФОТОТРОПИЗМ Апекс побега Зона изгиба Изгиб невозможен
 ФОТОТРОПИЗМ СВЕТ
ФОТОТРОПИЗМ СВЕТ
 Апикальное доминирование
Апикальное доминирование
 Индолилуксусная кислота Индолилмасляная кислота Гетероауксин – промышленный препарат смеси индольных") Закладка новых корней (Ризогенез) Индолилуксусная кислота Индолилмасляная кислота Гетероауксин – промышленный препарат смеси индольных соединений с ауксиновой активностью
Закладка новых корней (Ризогенез) Индолилуксусная кислота Индолилмасляная кислота Гетероауксин – промышленный препарат смеси индольных соединений с ауксиновой активностью
 Стимуляция роста плодов
Стимуляция роста плодов
 Гербицидное действие ауксинов Индолилуксусная кислота 2, 4 -Дихлорфеноксиуксусная кислота контроль 2, 4 -Д
Гербицидное действие ауксинов Индолилуксусная кислота 2, 4 -Дихлорфеноксиуксусная кислота контроль 2, 4 -Д
 Гербицидное действие ауксинов Индолилуксусная кислота 2, 4 -Дихлорфеноксиуксусная кислота 2, 4, 5 -Трихлорфеноксиуксусная кислота Гербицид «Дикамба» Гербицид «Клопиралид»
Гербицидное действие ауксинов Индолилуксусная кислота 2, 4 -Дихлорфеноксиуксусная кислота 2, 4, 5 -Трихлорфеноксиуксусная кислота Гербицид «Дикамба» Гербицид «Клопиралид»
 Использование гербицидов Гербицид «Дикамба» Гербицид «Клопиралид»
Использование гербицидов Гербицид «Дикамба» Гербицид «Клопиралид»
 Доказать, что раздражителем служит сила тяжести, можно с помощью клиностата.
Доказать, что раздражителем служит сила тяжести, можно с помощью клиностата.
 Центрифужное ускорение влияет на растение также, как ускорение силы тяжести.
Центрифужное ускорение влияет на растение также, как ускорение силы тяжести.
 Микрофотография клеток корневого чехлика
Микрофотография клеток корневого чехлика
 В ответ на геоиндукцию в корне происходят следующие процессы: • рецепция силы тяжести (корневой чехлик) • трансдукция механического сигнала в физиологический и распространение его к зоне ответа (к EZ) • ответ на сигнал, включающий изгибание корня за счет неравномерного растяжения клеток на верхней и нижней стороне DEZ
В ответ на геоиндукцию в корне происходят следующие процессы: • рецепция силы тяжести (корневой чехлик) • трансдукция механического сигнала в физиологический и распространение его к зоне ответа (к EZ) • ответ на сигнал, включающий изгибание корня за счет неравномерного растяжения клеток на верхней и нижней стороне DEZ
 Кончик корня как место восприятия геотропического раздражения Корень Vicia faba укрепляют на центрифуге так, чтобы центрифужное ускорение воздействовало на кончик и основание корня в противоположных направлениях. Результат: корень изгибается в направлении, показанном стрелкой, т. е. Соответственно, геоиндукции, полученной кончиком.
Кончик корня как место восприятия геотропического раздражения Корень Vicia faba укрепляют на центрифуге так, чтобы центрифужное ускорение воздействовало на кончик и основание корня в противоположных направлениях. Результат: корень изгибается в направлении, показанном стрелкой, т. е. Соответственно, геоиндукции, полученной кончиком.
 С-конец Coiled-coil –домен связь с цитоскелетом N-конец Трансмембранный") Arg 1 -белок (Dna. Jlike protein) С-конец Coiled-coil –домен связь с цитоскелетом N-конец Трансмембранный домен J-домен действует как шаперон
Arg 1 -белок (Dna. Jlike protein) С-конец Coiled-coil –домен связь с цитоскелетом N-конец Трансмембранный домен J-домен действует как шаперон
 A. до геоиндукции B. после геоиндукции") Модель транспорта ауксина в кончике корня (“fountain” model) A. до геоиндукции B. после геоиндукции
Модель транспорта ауксина в кончике корня (“fountain” model) A. до геоиндукции B. после геоиндукции
 Перераспределение выносящих ауксин переносчиков под действием геоиндукции A. до геоиндукции B. после геоиндукции
Перераспределение выносящих ауксин переносчиков под действием геоиндукции A. до геоиндукции B. после геоиндукции
 Распределение ауксинов при гравитропизме.
Распределение ауксинов при гравитропизме.
 В ответ на геоиндукцию в корне происходят следующие процессы: • рецепция силы тяжести (корневой чехлик) • трансдукция механического сигнала в физиологический и распространение его к зоне ответа (к EZ) • ответ на сигнал, включающий изгибание корня за счет неравномерного растяжения клеток на верхней и нижней стороне DEZ
В ответ на геоиндукцию в корне происходят следующие процессы: • рецепция силы тяжести (корневой чехлик) • трансдукция механического сигнала в физиологический и распространение его к зоне ответа (к EZ) • ответ на сигнал, включающий изгибание корня за счет неравномерного растяжения клеток на верхней и нижней стороне DEZ