

Обзорная лекция заочники биологи1.ppt
- Количество слайдов: 24
 Гипотезы происхождения Metazoa
Гипотезы происхождения Metazoa
 Отличия клеток простейших от клеток многоклеточных организмов • Клетка одноклеточного организма экологически, этологически и физиологически является самостоятельным организмом. • Форма клеток в многоклеточном организме разнообразнее. • У одноклеточных организмов встречается дуализм ядер (у инфузории парамеции-туфельки есть макронуклеус и микронуклеус). • У одноклеточных может быть много ядер. • Клетки одноклеточных способны к инцистированию. • Клетки одноклеточных не дифференцированы. • Клетки простейших не специализированы. • Митоз у одноклеточных разнообразнее. • У простейших многообразнее жизненные циклы. • В клетках многоклеточных нет пульсирующих вакуолей. • В колониях простейших клетки не интегрированы.
Отличия клеток простейших от клеток многоклеточных организмов • Клетка одноклеточного организма экологически, этологически и физиологически является самостоятельным организмом. • Форма клеток в многоклеточном организме разнообразнее. • У одноклеточных организмов встречается дуализм ядер (у инфузории парамеции-туфельки есть макронуклеус и микронуклеус). • У одноклеточных может быть много ядер. • Клетки одноклеточных способны к инцистированию. • Клетки одноклеточных не дифференцированы. • Клетки простейших не специализированы. • Митоз у одноклеточных разнообразнее. • У простейших многообразнее жизненные циклы. • В клетках многоклеточных нет пульсирующих вакуолей. • В колониях простейших клетки не интегрированы.
 Основные группы теорий происхождения многоклеточности • Колониальные теориии • Симбиотические теории • Теория целлюляризации
Основные группы теорий происхождения многоклеточности • Колониальные теориии • Симбиотические теории • Теория целлюляризации
 Теория гастреи • Сформулирована Э. Геккелем в 1872. • Согласно этой теории предком многоклеточных была гастрея - многоклеточный двуслойный организм. Она произошла от колониальных протистов с шарообразными колониями. Процесс интеграции клеток в колонии сделал возможным разделение функций между клетками: передние клетки утрачивают жгутики и превращаются в фагоциты, сидящие во впячивании на переднем конце - образуется кишечник. Остальные клетки утрачивают пищеварительную функцию и становятся чисто двигательными. Рот гастреи находился на переднем конце, и пища "сама заплывала" в кишечник. Симметрия у гастреи была радиальной. При переходе к сидячему образу жизни ее потомки эволюционировали в губок и кишечнополостных, а при переходе к ползанию по дну - в плоских червей и всех остальных многоклеточных.
Теория гастреи • Сформулирована Э. Геккелем в 1872. • Согласно этой теории предком многоклеточных была гастрея - многоклеточный двуслойный организм. Она произошла от колониальных протистов с шарообразными колониями. Процесс интеграции клеток в колонии сделал возможным разделение функций между клетками: передние клетки утрачивают жгутики и превращаются в фагоциты, сидящие во впячивании на переднем конце - образуется кишечник. Остальные клетки утрачивают пищеварительную функцию и становятся чисто двигательными. Рот гастреи находился на переднем конце, и пища "сама заплывала" в кишечник. Симметрия у гастреи была радиальной. При переходе к сидячему образу жизни ее потомки эволюционировали в губок и кишечнополостных, а при переходе к ползанию по дну - в плоских червей и всех остальных многоклеточных.
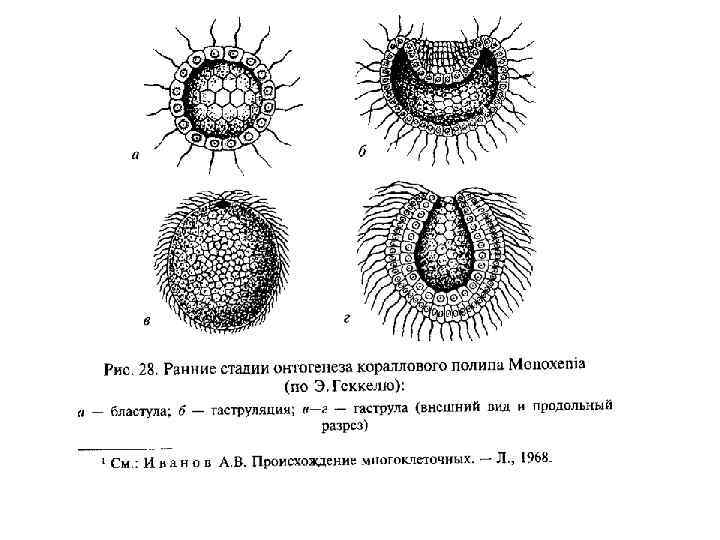
 Теория фагоцителлы • Разработана И. И. Мечниковым в 1879– 86. • У примитивных кишечнополостных гаструляция происходит не путем инвагинации (впячивания одного полюса однослойного зародыша — бластулы), а посредством миграции некоторых клеток из однослойной стенки тела внутрь. Там они образуют рыхлое скопление, позднее организующееся в виде стенок гастральной полости, которая прорывается наружу ротовым отверстием. • В сфероидной колонии простейших — жгутиконосцев клетки ее однослойной стенки, захватывавшие пищу, мигрировали для ее переваривания внутрь, в полость колонии. Эти клетки образовали рыхлое внутреннее скопление — фагоцитобласт, функцией которого стало обеспечение всего организма пищей, включая ее переваривание и распределение, тогда как поверхностный слой клеток — кинобласт — осуществлял функции защиты и движения организма. Эта гипотетическая стадия эволюции Metazoa была названа Мечниковым фагоцителлой (или паренхимеллой); ее строение соответствует таковому паренхимулы, личинки некоторых кишечнополостных и губок. • В дальнейшем у потомков фагоцителлы произошла эпителизация фагоцитобласта с образованием первичного кишечника и возникновением ротового отверстия в том месте, где происходила преимущественная миграция клеток внутрь. Это место, вероятно, соответствовало заднему по направлению движения полюсу тела, где при плавании возникают завихрения водяного потока, и поэтому условия наиболее благоприятны для захвата пищевых частиц.
Теория фагоцителлы • Разработана И. И. Мечниковым в 1879– 86. • У примитивных кишечнополостных гаструляция происходит не путем инвагинации (впячивания одного полюса однослойного зародыша — бластулы), а посредством миграции некоторых клеток из однослойной стенки тела внутрь. Там они образуют рыхлое скопление, позднее организующееся в виде стенок гастральной полости, которая прорывается наружу ротовым отверстием. • В сфероидной колонии простейших — жгутиконосцев клетки ее однослойной стенки, захватывавшие пищу, мигрировали для ее переваривания внутрь, в полость колонии. Эти клетки образовали рыхлое внутреннее скопление — фагоцитобласт, функцией которого стало обеспечение всего организма пищей, включая ее переваривание и распределение, тогда как поверхностный слой клеток — кинобласт — осуществлял функции защиты и движения организма. Эта гипотетическая стадия эволюции Metazoa была названа Мечниковым фагоцителлой (или паренхимеллой); ее строение соответствует таковому паренхимулы, личинки некоторых кишечнополостных и губок. • В дальнейшем у потомков фагоцителлы произошла эпителизация фагоцитобласта с образованием первичного кишечника и возникновением ротового отверстия в том месте, где происходила преимущественная миграция клеток внутрь. Это место, вероятно, соответствовало заднему по направлению движения полюсу тела, где при плавании возникают завихрения водяного потока, и поэтому условия наиболее благоприятны для захвата пищевых частиц.

 Теория синзооспоры • Разработана А. А. Захваткиным в 1949 году • Согласно данной теории многоклеточные произошли от колониальных протистов. У протистов встречаются клетки, сильно увеличенные за счет запасания питательных веществ - как яйцеклетка у животных. Часто такие клетки делятся несколько раз подряд - это похоже на дробление. Таким способом образуются у протистов одноклеточные мелкие расселительные стадии - зооспоры. У колониальных протистов зооспоры могут оставаться все вместе, образуя колонию - синзооспору. В процессе эволюции могла произойти неотения и утратиться взрослая сидячая стадия. Таким образом бластула - это синзооспора, семья зооспор. Отличия от теорий фагоцителлы и гастреи: • Считается, что никогда не существовало однослойного шарообразного предка. Об этом свидетельствует то, что у всех многоклеточных бластулы не питаются. Не питаются и образующиеся из них паренхимулы. Поэтому и у древних многоклеточных такие стадии не были взрослыми организмами - это были всегда только личинки. • Сидячий образ жизни примитивных взрослых многоклеточных. • Наиболее примитивной из расселительных личинок считается бластула. В паренхимулу она превращается, готовясь к превращению во взрослый организм. Этот метаморфоз происходит после перехода к сидячему образу жизни. У всех остальных многоклеточных взрослая сидячая стадия утратилась. У этих животных личинки стали взрослыми - произошла неотения.
Теория синзооспоры • Разработана А. А. Захваткиным в 1949 году • Согласно данной теории многоклеточные произошли от колониальных протистов. У протистов встречаются клетки, сильно увеличенные за счет запасания питательных веществ - как яйцеклетка у животных. Часто такие клетки делятся несколько раз подряд - это похоже на дробление. Таким способом образуются у протистов одноклеточные мелкие расселительные стадии - зооспоры. У колониальных протистов зооспоры могут оставаться все вместе, образуя колонию - синзооспору. В процессе эволюции могла произойти неотения и утратиться взрослая сидячая стадия. Таким образом бластула - это синзооспора, семья зооспор. Отличия от теорий фагоцителлы и гастреи: • Считается, что никогда не существовало однослойного шарообразного предка. Об этом свидетельствует то, что у всех многоклеточных бластулы не питаются. Не питаются и образующиеся из них паренхимулы. Поэтому и у древних многоклеточных такие стадии не были взрослыми организмами - это были всегда только личинки. • Сидячий образ жизни примитивных взрослых многоклеточных. • Наиболее примитивной из расселительных личинок считается бластула. В паренхимулу она превращается, готовясь к превращению во взрослый организм. Этот метаморфоз происходит после перехода к сидячему образу жизни. У всех остальных многоклеточных взрослая сидячая стадия утратилась. У этих животных личинки стали взрослыми - произошла неотения.
 Теория целлюляризации • Высказана впервые Иерингом в 1877 году, позднее пропагандировалась югославским зоологом Иованом Хаджи • Эта теория предполагает, что предками многоклеточных были сложно организованные протисты, такие как инфузории, а органы многоклеточных образовались в результате отделения мембранами органелл. Так, в соответствии с этой теорией кишечник образовался из глотки инфузории, выделительная система - из ее сократительных вакуолей, покровы - из периферического слоя цитоплазмы. Предполагается далее, что акт спаривания самца и самки у многоклеточных животных развился из конъюгации двух инфузорий, а процесс оплодотворения яйца у Metazoa - из процесса слияния двух половых ядер конъюгирующих особей, наконец, все тело инфузории приравнивается телу целого многоклеточного организма. Потомками инфузорий по мнению авторов гиппотезы являются бескишечные турбеллярии • Эта теория полностью не соответствует взглядам современной науки и является безусловно ошибочной. Эта теория на сегодняшнем уровне знаний имеет лишь исторический интерес.
Теория целлюляризации • Высказана впервые Иерингом в 1877 году, позднее пропагандировалась югославским зоологом Иованом Хаджи • Эта теория предполагает, что предками многоклеточных были сложно организованные протисты, такие как инфузории, а органы многоклеточных образовались в результате отделения мембранами органелл. Так, в соответствии с этой теорией кишечник образовался из глотки инфузории, выделительная система - из ее сократительных вакуолей, покровы - из периферического слоя цитоплазмы. Предполагается далее, что акт спаривания самца и самки у многоклеточных животных развился из конъюгации двух инфузорий, а процесс оплодотворения яйца у Metazoa - из процесса слияния двух половых ядер конъюгирующих особей, наконец, все тело инфузории приравнивается телу целого многоклеточного организма. Потомками инфузорий по мнению авторов гиппотезы являются бескишечные турбеллярии • Эта теория полностью не соответствует взглядам современной науки и является безусловно ошибочной. Эта теория на сегодняшнем уровне знаний имеет лишь исторический интерес.
 О предках многоклеточных • В рамках рассмотренных теорий предполагается, что скорее всего предками многоклеточных была группа воротничковых жгутиконосцев - хоанофлагеллят. В пользу этого говорит то, что для примитивных многоклеточных очень характерны воротничково - жгутиковые клетки, строение которых очень слабо отличается от строения хоанофлагеллят. Подходящий у воротничковых жгутиконосцев и способ питания. Все они гетеротрофы, питающиеся за счет фагоцитоза и пиноцитоза. Им свойственна колониальность. Среди колоний есть даже такие, у которых наружные клетки имеют воротнички и жгутики, а внутренние - амебоидные. Единственный недостаток хоанофлагеллят, как предков многоклеточных, состоит в том, что до сих пор у них достоверно известен только один способ размножения - деление пополам. Ничего похожего на половое размножение или на дробление у них нет. Тем не менее, многие ученые считают эту группу протистов наиболее вероятными предками всех Metazoa
О предках многоклеточных • В рамках рассмотренных теорий предполагается, что скорее всего предками многоклеточных была группа воротничковых жгутиконосцев - хоанофлагеллят. В пользу этого говорит то, что для примитивных многоклеточных очень характерны воротничково - жгутиковые клетки, строение которых очень слабо отличается от строения хоанофлагеллят. Подходящий у воротничковых жгутиконосцев и способ питания. Все они гетеротрофы, питающиеся за счет фагоцитоза и пиноцитоза. Им свойственна колониальность. Среди колоний есть даже такие, у которых наружные клетки имеют воротнички и жгутики, а внутренние - амебоидные. Единственный недостаток хоанофлагеллят, как предков многоклеточных, состоит в том, что до сих пор у них достоверно известен только один способ размножения - деление пополам. Ничего похожего на половое размножение или на дробление у них нет. Тем не менее, многие ученые считают эту группу протистов наиболее вероятными предками всех Metazoa
: А - Uroglenopsis americana, общий вид; Б - то") Монотомические колонии жгутиковых (из Захваткина): А - Uroglenopsis americana, общий вид; Б - то же, оптический разрез через часть колонии; В - то же, отдельная делящаяся особь; Г - Syncrypta vol vox; Д - Chrysosphaerella longispina; E - Synura uvella, общий вид колонии; Ж - то же, делящаяся особь; 3 - Sphaeroeca volvox, общий вид; И - один из членов колонии; К - одиночное воротничковое жгутиковое Соdosiga
Монотомические колонии жгутиковых (из Захваткина): А - Uroglenopsis americana, общий вид; Б - то же, оптический разрез через часть колонии; В - то же, отдельная делящаяся особь; Г - Syncrypta vol vox; Д - Chrysosphaerella longispina; E - Synura uvella, общий вид колонии; Ж - то же, делящаяся особь; 3 - Sphaeroeca volvox, общий вид; И - один из членов колонии; К - одиночное воротничковое жгутиковое Соdosiga
") Бесполое размножение беспозвоночных (основные типы бесполого размножения; адаптивное значение процесса)
Бесполое размножение беспозвоночных (основные типы бесполого размножения; адаптивное значение процесса)
 • Бесполое размножение широко распространено среди низших беспозвоночных (простейшие, губки, кишечнополостные, плоские черви, кольчатые черви и т. п. ), но не встречается среди высших беспозвоночных (членистоногие, моллюски). Для бесполого размножения характерно то, что начало новому организму дают соматические клетки, а не специализированные половые. • Очень распространено бесполое размножение у простейших. Осуществляется оно посредством деления клетки. Самый простой вариант – деление клетки пополам. Часто встречается так же множественное деление - шизогония, палинтомия. В этом случае из одной материнской клетки получается большое количество дочерних. • Распространено бесполое размножение и у многоклеточных. В этом случае обычно выделяют 2 его формы: фрагментация и почкование. При фрагментации тело животного распадается на несколько кусков, каждый из которых дорастает в последствии до целого организма. Такой способ типичен для плоских и некоторых кольчатых червей, иглокожих. • Почкование - тип бесполого размножения животных, при котором дочерние особи формируются из выростов тела материнского организма (почек). Почкование характерно для губок, кишечнополостных. Часто почкование не проходит до конца, молодые особи не отделяются и образуется колония.
• Бесполое размножение широко распространено среди низших беспозвоночных (простейшие, губки, кишечнополостные, плоские черви, кольчатые черви и т. п. ), но не встречается среди высших беспозвоночных (членистоногие, моллюски). Для бесполого размножения характерно то, что начало новому организму дают соматические клетки, а не специализированные половые. • Очень распространено бесполое размножение у простейших. Осуществляется оно посредством деления клетки. Самый простой вариант – деление клетки пополам. Часто встречается так же множественное деление - шизогония, палинтомия. В этом случае из одной материнской клетки получается большое количество дочерних. • Распространено бесполое размножение и у многоклеточных. В этом случае обычно выделяют 2 его формы: фрагментация и почкование. При фрагментации тело животного распадается на несколько кусков, каждый из которых дорастает в последствии до целого организма. Такой способ типичен для плоских и некоторых кольчатых червей, иглокожих. • Почкование - тип бесполого размножения животных, при котором дочерние особи формируются из выростов тела материнского организма (почек). Почкование характерно для губок, кишечнополостных. Часто почкование не проходит до конца, молодые особи не отделяются и образуется колония.
 Hydra oligactis при небольшом увеличении. А - с мужскими гонадами; Б - с женскими гонадами; В - почкующаяся гидра
Hydra oligactis при небольшом увеличении. А - с мужскими гонадами; Б - с женскими гонадами; В - почкующаяся гидра
 в состоянии деления - цепочка из 16 особей (из Граффа): 1") Турбеллярия Microstomum (Macrostomida) в состоянии деления - цепочка из 16 особей (из Граффа): 1 - передняя (материнская) особь, 2 - задняя (1 -я дочерняя) особь, 3, 4 - дочерние особи, отделившиеся от двух первых и частично уже сформированные, 5 - начальные стадии отделения новых дочерних особей, 6 - кишечник, 7 - глотка, 8 - глаза, 9 - границы между формирующимися особями
Турбеллярия Microstomum (Macrostomida) в состоянии деления - цепочка из 16 особей (из Граффа): 1 - передняя (материнская) особь, 2 - задняя (1 -я дочерняя) особь, 3, 4 - дочерние особи, отделившиеся от двух первых и частично уже сформированные, 5 - начальные стадии отделения новых дочерних особей, 6 - кишечник, 7 - глотка, 8 - глаза, 9 - границы между формирующимися особями
; Б и В -") Размножение полихет. А - бесполое размножение полихеты Autolytus (по Грассе); Б и В - получающиеся посредством концевого почкования половые особи (Б - самка и В - самец) тех же червей (по Свешникову)
Размножение полихет. А - бесполое размножение полихеты Autolytus (по Грассе); Б и В - получающиеся посредством концевого почкования половые особи (Б - самка и В - самец) тех же червей (по Свешникову)
 • Все особи, появившиеся в результате бесполого размножения генетически идентичны, поскольку рекомбинации генома не было. Бесполое размножение позволяет быстро и с меньшими затратами повысить численность вида, но при длительном размножении только бесполым путем возможно накопление негативных мутаций в соматических клетках.
• Все особи, появившиеся в результате бесполого размножения генетически идентичны, поскольку рекомбинации генома не было. Бесполое размножение позволяет быстро и с меньшими затратами повысить численность вида, но при длительном размножении только бесполым путем возможно накопление негативных мутаций в соматических клетках.
 Половое размножение беспозвоночных Понятие о чередовании поколений. Примеры.
Половое размножение беспозвоночных Понятие о чередовании поколений. Примеры.
 • Конъюгация. Своеобразная форма полового процесса, характерная для инфузорий. Во время конъюгации происходит обмен частью ядерного аппарата между конъюгирующими особями. Это позволяет каждой особи обновить свой ядерный аппарат и не допускает накопления негативных мутаций. Увеличения количества особей в результате конъюгации не происходит. После конъюгации начинается размножение бесполым путем. • Размножение посредством слияния гамет, каждая из которых имеет гаплоидный набор хромосом. В результате оплодотворения получается диплоидная зигота, дающая начало новому организму. По способу оплодотворения различают наружное оплодотворение, внутреннее оплодотворение и оплодотворение посредством сперматофора. В зависимости от размера гамет выделяют изогамию, анизогамию, овогамию. По половой принадлежности родительские организмы могут быть либо раздельнополы, либо гермафродитные, но даже в случае гермафродитных организмов самооплодотворение чаще всего невозможно. • Партеногенез. Размножение, когда развитие яйцеклетки начинается без оплодотворения. При этом либо яйцеклетка в результате усложнения мейоза становится диплоидной, либо развивается гаплоидный организм.
• Конъюгация. Своеобразная форма полового процесса, характерная для инфузорий. Во время конъюгации происходит обмен частью ядерного аппарата между конъюгирующими особями. Это позволяет каждой особи обновить свой ядерный аппарат и не допускает накопления негативных мутаций. Увеличения количества особей в результате конъюгации не происходит. После конъюгации начинается размножение бесполым путем. • Размножение посредством слияния гамет, каждая из которых имеет гаплоидный набор хромосом. В результате оплодотворения получается диплоидная зигота, дающая начало новому организму. По способу оплодотворения различают наружное оплодотворение, внутреннее оплодотворение и оплодотворение посредством сперматофора. В зависимости от размера гамет выделяют изогамию, анизогамию, овогамию. По половой принадлежности родительские организмы могут быть либо раздельнополы, либо гермафродитные, но даже в случае гермафродитных организмов самооплодотворение чаще всего невозможно. • Партеногенез. Размножение, когда развитие яйцеклетки начинается без оплодотворения. При этом либо яйцеклетка в результате усложнения мейоза становится диплоидной, либо развивается гаплоидный организм.
 Конъюгация у инфузорий 1 -соединение конъюгантов, 2 -4 деление микронуклеуса на четыре ядра, 5 -редукция трех ядер из четырех, 6 -деление оставшегося ядра на стационарное и мигрирующее и обмен мигрирующими ядрами, 7 -слияние ядер и образование синкариона, 8 -расхождение конъюгантов, 9 -реорганизация ядерного аппарата при делении клеток после конъюгации, Мамакронуклеус, Ми-микронуклеус
Конъюгация у инфузорий 1 -соединение конъюгантов, 2 -4 деление микронуклеуса на четыре ядра, 5 -редукция трех ядер из четырех, 6 -деление оставшегося ядра на стационарное и мигрирующее и обмен мигрирующими ядрами, 7 -слияние ядер и образование синкариона, 8 -расхождение конъюгантов, 9 -реорганизация ядерного аппарата при делении клеток после конъюгации, Мамакронуклеус, Ми-микронуклеус
 • Для многих беспозвоночных характерно чередование разных способов размножения. В таком случае говорят о чередовании поколений. Это может быть чередование бесполого и полового поколения, либо чередование гермафродитного и партеногенетического поколений. Крайне редко (у фораминифер) имеется чередование гаплоидного и диплоидного поколений.
• Для многих беспозвоночных характерно чередование разных способов размножения. В таком случае говорят о чередовании поколений. Это может быть чередование бесполого и полового поколения, либо чередование гермафродитного и партеногенетического поколений. Крайне редко (у фораминифер) имеется чередование гаплоидного и диплоидного поколений.
;") Жизненный цикл гидроида из рода Obelia: А — яйцо; Б — планула (личинка); В — колония полипов с одним видоизменённым полипом — бластостилем, на котором видны развивающиеся медузы; Г — отделившаяся медуза.
Жизненный цикл гидроида из рода Obelia: А — яйцо; Б — планула (личинка); В — колония полипов с одним видоизменённым полипом — бластостилем, на котором видны развивающиеся медузы; Г — отделившаяся медуза.
: 1 - окончательный хозяин (лягушка),") Схема типичного жизненного цикла дигенетического сосальщика (по Смиту, изменено): 1 - окончательный хозяин (лягушка), в котором паразитируют половозрелые сосальщики гермафродитного поколения (мариты), 2 - яйца сосальщика, выходящие из кишечника хозяина в воду, 3 - первый промежуточный хозяин (улитка), в котором паразитируют партеногенетические поколения, 4 - свободноплавающая личинка (церкария), внедряющаяся в тело второго промежуточного хозяина, 5 - второй промежуточный хозяин (водная личинка стрекозы), в полости тела которой инцистировалась метацеркария (6). Окончательный хозяин заражается паразитом, поедая личинку стрекозы с метацеркарией
Схема типичного жизненного цикла дигенетического сосальщика (по Смиту, изменено): 1 - окончательный хозяин (лягушка), в котором паразитируют половозрелые сосальщики гермафродитного поколения (мариты), 2 - яйца сосальщика, выходящие из кишечника хозяина в воду, 3 - первый промежуточный хозяин (улитка), в котором паразитируют партеногенетические поколения, 4 - свободноплавающая личинка (церкария), внедряющаяся в тело второго промежуточного хозяина, 5 - второй промежуточный хозяин (водная личинка стрекозы), в полости тела которой инцистировалась метацеркария (6). Окончательный хозяин заражается паразитом, поедая личинку стрекозы с метацеркарией
: 1 - одноядерный гамонт, 2 - гамонт") Цикл развития фораминиферы Myxotheca arenilega (пo Грелю): 1 - одноядерный гамонт, 2 - гамонт после образования ядер гамет, 3 - копуляция гамет, 4 - зигота, 5 - молодой агамонт, 6 - растущий агамонт, 7 - мейоз (момент редукции); 8 - образование агамет, 9 - молодая агамета (гамонт)
Цикл развития фораминиферы Myxotheca arenilega (пo Грелю): 1 - одноядерный гамонт, 2 - гамонт после образования ядер гамет, 3 - копуляция гамет, 4 - зигота, 5 - молодой агамонт, 6 - растущий агамонт, 7 - мейоз (момент редукции); 8 - образование агамет, 9 - молодая агамета (гамонт)