Биология_5_Гены (2)Мутации.pptx
- Количество слайдов: 31


 Мутации")
Гены (2) Мутации

Мутации: общие положения • • • Мутация – это, согласно "Bio. File. ru", внезапное наследственное изменение, вызванное резким структурным и функциональным изменением генетического материала. Генетический материал организован в иерархию структурно-функциональных единиц – от молекулярных сайтов внутри гена до целых хромосом и геномов. Соответственно существуют разные типы мутаций – от генных до геномных. Этот раздел посвящён в основном генным мутациям. Внезапные наследственные изменения фенотипа могут быть вызваны не только структурными изменениями генов, но и другими генетическими процессами. Мутации могут быть истинными или ложными. Фенотипические изменения сами по себе не дают представления о тех генетических процессах, которые их вызывают. На основании одних лишь прямых наблюдений трудно различать разные типы истинных и ложных мутаций. Существует также, как мы увидим в дальнейшем, внезапное изменение генетического материала, не вызывающее фенотипического эффекта. Здесь следует сразу же оговориться, что и внезапность, и случайность мутаций далеко не абсолютны. Согласно современным представлениям, мутации, приводящие в конечном счёте к эволюционным изменениям, в основном направленны и детерминированы факторами внешней среды.
 макроэволюционные преобразования связаны со")
Точечные мутации • • По мнению В. Н. Стегния (2013) макроэволюционные преобразования связаны со всеми типами мутаций, причём главным аспектом макроэволюционных изменений является их необратимость. В этом плане генные мутации, связанные с заменой нуклеотидов, являются обратимыми и относятся в основном к микроэволюционным событиям. Современная типология генных мутаций была хорошо разработана М. Д. Голубовским в 1981 г. Она включает облигатные компоненты (ОК) генома (структурные гены ядра и органелл цитоплазмы) и факультативные компоненты (ФК) (мобильные элементы, транспозоны, вирусы, плазмиды и др. ). Природные наследуемые изменения представлены двумя этапами. Сначала активируются ФК, а затем опосредованно изменяются генные локусы (ОК).

Классификация точечных мутаций • • • Точечная мутация это тип мутации в ДНК или РНК, для которого характерна замена одного азотистого основания другим. Термин также применяется и в отношении парных замен, инсерции или делеции одного или нескольких нуклеотидов. Точечные мутации, возникающие в некодирующей ДНК, обычно никак себя не проявляют. Точечный мутант – организм, в генотипе которого произошла точечная мутация. Точечные мутации классифицируют по воздействию, которое изменённый нуклеотид оказывает на триплет: нонсенс-мутация – мутация, в результате которой кодон теряет способность кодировать какую-либо аминокислоту и становится стоп-кодоном, что приводит к преждевременной терминации синтеза белка; миссенс-мутация – переключает кодон на кодирование другой аминокислоты; сеймсенс-мутация – кодон продолжает кодировать ту же аминокислоту. Точечные мутации замены оснований классифицируют на транзиции и трансверсии. Транзиция – одно пуриновое основание замещается на другое (аденин на гуанин или наоборот), либо происходит аналогичная замена пиримидиновых оснований (тимин с цитозином). Трансверсия – пуриновое основание замещается на пиримидиновое основание или наоборот. Транзиции происходят чаще, чем трансверсии.
 • • • Мутации замены оснований не всегда образуются сразу")
Классификация точечных мутаций (2) • • • Мутации замены оснований не всегда образуются сразу же после воздействия мутагена. Иногда они появляются после десятков циклов репликации. Это явление носит название задерживающихся мутаций. Нестабильность генома – главная причина образования злокачественных опухолей. Механизмы образования задерживающихся мутаций в настоящее время не известны. Точечные мутации сдвига рамки чтения гена классифицируют на делеции и инсерции: делеция – в молекуле ДНК выпадает один или несколько нуклеотидов; инсерция – в молекулу ДНК встраивается один или несколько нуклеотидов. Точечные мутации противопоставляются сложным мутациям, при которых один участок ДНК заменяется участком другой длины и иного нуклеотидного состава. Точечные мутации могут появляться напротив таких повреждений молекулы ДНК, которые способны останавливать синтез ДНК. Мутации этого типа образуются не всегда сразу же после воздействия мутагена. Иногда они появляются после десятков циклов репликации. Это явление носит название задерживающихся мутаций, например, при нестабильности генома – главной причине образования злокачественных опухолей.

Причины возникновения точечных мутаций • • • Точечные мутации могут возникать во время репликации ДНК. Они также могут возникать в результате действия мутагенов – к примеру, воздействия ультрафиолетового или рентгеновского излучения, высокой температуры или химических веществ. Мутации появляются при синтезе молекулы ДНК, содержащей повреждения, в процессах репликации ДНК, репарации ДНК или транскрипции. В настоящее время существует несколько подходов, используемых для объяснения природы и механизмов образования точечных мутаций. В рамках общепринятой модели считается, что единственная причина образования мутаций замены оснований – спорадические ошибки ДНК-полимераз. Генная, или точечная, мутация представляет собой изменение последовательности нуклеотидов в пределах одного гена, приводящее к изменению характера действия гена. Как правило, это молекулярное изменение в гене, которое вызывает фенотипический эффект. Допустим, что какой-то ген содержит в некоторой своей точке кодон, или триплет, ЦТТ, кодирующий одну из аминокислот полипептидной цепи – глутаминовую кислоту. В результате замены всего лишь одного нуклеотида кодон ЦТТ может превратиться в кодон ГТТ, Этот новый кодон обусловливает синтез уже не глутаминовой кислоты, а глутамина, так что в полипептидной цепи, синтезируемой под действием измененного гена, на месте глутаминовой кислоты окажется глутамин. Первоначальная и мутантная молекулы белка отличаются одна от другой, и вполне возможно, что это влечет за собой другие, вторичные, фенотипические различия.
 • • Стабильность генов на протяжении последовательных поколений клеток")
Причины возникновения точечных мутаций (2) • • Стабильность генов на протяжении последовательных поколений клеток и особей, а следовательно, и консервативность наследственности обусловлены точностью процесса копирования при репликации гена. Однако процесс копирования несовершенен. Время от времени при копировании возникают ошибки. Генные мутации можно рассматривать как такие ошибки копирования. Следует иметь в виду, что ошибки копирования, помимо их случайности, одновременно являются и необходимостью, так как служат одним из важных источников как генотипической, так и фенотипической изменчивости. По данным Е. Кунина нормой является 1 -10 ошибок на одну рамку считывания. Любой ген, входящий в состав генотипа, по-видимому, подвержен мутированию. Во всяком случае, в генах, контролирующих весьма разнообразные признаки, наблюдаются мутации. Например, у Drosophila melanogaster известны мутанты со слегка сморщенными крыльями, сильно укороченными крыльями или вообще бескрылые; мутанты с белыми или пурпурными глазами; мутанты с разнообразными изменениями щетинок и т. п. Известен ряд мутантных разновидностей смородинного томата (Lycopersicon pimpinellifolium), различающихся по форме листьев. Биохимические мутации, затрагивающие различные звенья метаболических процессов, хорошо известны у микроорганизмов и имеются, хотя и гораздо менее изучены, у высших организмов.

Частота возникновения мутаций • • Частота возникновения отдельных спонтанных мутаций выражается числом гамет одного поколения, несущих определенную мутацию по отношению к общему числу гамет. Частоты, точно определенные для некоторых видов растений, животных и микроорганизмов, оказываются весьма близкими по величине и составляют в среднем от 1 из 100000 или 1 из 10 000 мутантных гамет в определенном локусе. Частота мутаций неодинакова для разных генов. Общая частота мутаций, складывающаяся из частот мутаций отдельных генов, также оказывается близкой у разных организмов: от нескольких процентов (одноклеточные водоросли, низшие грибы, бактерии) до 25% (дрозофила) всех гамет одного поколения. Приведенные цифры касались частоты спонтанных мутаций. При воздействии облучением, определенными химическими веществами, температурой частота мутаций резко повышается и может достигать значительных величин. На частоту возникновения мутаций оказывает влияние физиологическое состояние организма. Например, в процессе хранения семян частота мутаций (особенно хромосомных) заметно увеличивается. Точно так же отмечено увеличение частоты мутаций в спермиях дрозофилы при хранении ее в семяприемниках самки. Есть данные о том, что частота мутирования одного и того же локуса у гетерозиготных особей выше, чем у гомозиготных. Спектр мутантных признаков, затрагиваемых мутациями, очень широкий. Нет признаков и свойств, которые в той или иной степени не затрагивались бы мутациями.
 • • Наследственной изменчивости (изменчивости под воздействие мутаций) подвержены все морфологические, физиологические,")
Мутации (продолжение) • • Наследственной изменчивости (изменчивости под воздействие мутаций) подвержены все морфологические, физиологические, биохимические, этологические и другие признаки и свойства. Эти вариации по средним значениям варьирующих признаков выражаются как в качественных различиях, так и количественно. Мутации могут происходить как в сторону увеличения, так и уменьшения выраженности определенного признака или свойства. Они могут быть выражены резко (вплоть до летальности) или представлены в виде незначительных отклонений от исходной формы ( «малые» мутации). В качестве примера «большой» мутации может быть упомянута мутация tetraptera у дрозофилы, которая привела к возникновению признака, характерного для другого отряда (четыре крыла вместо обычных для мух двух крыльев). Во многих работах показано, что мутации затрагивают такие существенные биологические признаки, как общая жизнеспособность, способность к скрещиванию, плодовитость, скорость роста и др. являются важнейшим свойством, определяющим значение мутаций в эволюции как элементарного эволюционного материала.

Генотипический контроль мутаций • • У Drosophila melanogaster известен ген, который вызывает высокую частоту мутаций других генов данного комплемента. Этот ген-мутатор обозначают символом Hi. У мух, гомозиготных по гену Hi, частота мутаций в 10 раз выше обычной; у гетерозигот по этому гену частота мутаций выше обычной в 2 -7 раз. Ген Hi индуцирует как видимые, так и летальные мутации во многих генах. Он вызывает также инверсии – один из типов хромосомных мутаций. Выше мы рассматривали генные мутации как случайные ошибки копирования, происходящие во время воспроизведения гена, и это, несомненно, само по себе верно. Однако, как показывают данные о действии генов-мутаторов, существует и другой аспект мутационного процесса. Возникновение новой мутационной изменчивости, которая имеет важное значение для долговременного успеха данного вида в эволюции, может быть не целиком предоставлено воле случая, а инициироваться генами-мутаторами. Частота возникновения мутаций у данного вида частично может быть одним из генотипически контролируемых компонентов всей его генетической системы.

Адаптивная ценность мутаций • • Большинство новых, полученных в эксперименте, мутантов характеризуется более низкой жизнеспособностью, чем нормальный или дикий тип. Снижение жизнеспособности может быть выражено в разной степени – от чуть заметного субвитального состояния до полулетальности и летальности. При оценке жизнеспособности мутантов Drosophiia melanogaster, возникших в результате мутаций в Х-хромосоме, 90% оказались менее жизнеспособными, чем нормальные мухи, а 10% были супервитальными, т. е. обладали повышенной жизнеспособностью. Среди 90% мух с пониженной жизнеспособностью наблюдается весь диапазон изменений от слабой субвитальности (45%) через промежуточные стадии понижения жизнеспособности до полулетальности (6%) и летальности (14%). Вообще адаптивная ценность новых мутантов обычно бывает понижена. Адаптивная ценность слагается из плодовитости и функциональной полезности морфологических признаков, а также физиологической жизнеспособности. Многие мутанты неплодовиты независимо от того, обладают они нормальной жизнеспособностью или нет. Морфологические макромутации обычно сопровождаются нарушениями функциональной эффективности. Из обширной выборки индуцированных мутантов ячменя (Hordeum vulgare) у 99% адаптивная ценность была понижена.
 • • • Этим наблюдениям легко дать объяснение. Все гены,")
Адаптивная ценность мутаций (2) • • • Этим наблюдениям легко дать объяснение. Все гены, входящие в состав генотипа нормальной, или дикой, формы на протяжении многих поколений подвергались естественному отбору; они прошли сквозь сито стабилизирующего отбора, в результате чего сохранились те из них, которые обладали максимальной адаптивной ценностью. Следует ожидать, что любые изменения в таких генах, почти наверняка, окажутся изменениями к худшему, подобно тому, как копанье наугад в часовом механизме скорее нарушит, чем усовершенствует его «функциональную эффективность» . Генные мутации нередко называют случайными изменениями в генах. В данном контексте определение «случайные» требует разъяснения. Мутационные изменения на самом деле могут не быть случайными на молекулярном уровне. Определенные изменения последовательности нуклеотидов могут возникать в цепи ДНК чаще, чем другие. Так называемая случайность мутационного процесса относится не к молекулярному строению, а к адаптивным свойствам мутантных генов. Спонтанные мутации случайны в том смысле, что они не направлены в сторону какого-либо нынешнего или будущего состояния приспособленности данного организма. В лабораторных и полевых опытах некоторые мутации могут оказаться и полезными, однако вряд ли это может относиться к диким организмам.

Роль и значение мутаций • • Относительное значение в эволюции малых мутаций и макромутаций Большинство эволюционных генетиков до настоящего времени подчеркивают важную роль в эволюции малых мутаций. Согласно же мнению меньшинства, к которому принадлежат Гольдшмидт и некоторые современные ученые, главную роль в эволюции играют макромутации. В прошлом, как и в настоящем, между сторонниками двух указанных противоположных точек зрения было и остаётся много споров, которые, с одной точки зрения, излишни, поскольку эти взгляды не исключают, а напротив, дополняют друга, в эволюции играют роль как малые мутации, так и макромутации. С другой, более современной точки зрения, малые мутации более характерны и существенны для внутрипопуляционной изменчивости, в то время как макромутации играли и, вероятно будут играть решающую роль в эволюционных процессах. К этим вопросам мы вернёмся в разделах, посвящённых теории эволюции. Вероятность сохранения новой мутации Каждый отдельный мутантный аллель при своем первом возникновении бывает редок. Насколько вероятно, что он сохранится в родительской популяции? Мы знаем, что частота нового мутантного аллеля, нe может увеличиваться за счет одного лишь процесса воспроизведения. Вероятность же чисто случайной элиминации мутантного аллеля, напротив, очень велика.

Давление мутаций • • • Роль мутационного процесса в эволюции можно определить следующим образом. Мутации служат основным источником новшеств в виде внутрипопуляционной изменчивости. Однако нормальный мутационный процесс обычно не представляет собой важной эволюционной силы в ходе крупных эволюционных сдвигов. Частота возникновения мутаций столь низка, что само по себе мутационное давление не может вызвать существенных изменений частот аллелей в популяциях. Кроме того, прямые мутации А→а со временем отсеиваются, полностью или частично, в результате обратных мутаций а→А. Приведенное выше утверждение касается нормы. В тех особых ситуациях, когда давление мутаций становится значительной силой, вполне могут возникнуть исключения. Во всей Британской Колумбии и на Аляске окраска цветков у Epilobium angustifolium по большей части одинакова; однако в одной популяции, растущей на вершине урансодержащих гор вблизи Большого Медвежьего озера (Канада), обнаружены многочисленные мутантные растения с аберрантными бледно-розовыми цветками. Высокая частота хромосомных мутаций наблюдалась у растений, выращенных из семян, которые находились в зоне извержения вулкана в Исландии. Так или иначе, мутационное давление независимо от его интенсивности не может быть направляющей силой эволюции. Огромное большинство мутаций, как мы видели, вредны и им суждено быть элиминированными из популяции. Ориентация, или направленность, процесса эволюции определяется другими эволюционными силами.

Геномика • • • Гено мика это раздел молекулярной генетики, посвящённый изучению генома и генов живых организмов. Геномика сформировалась как особое направление генетики в 1980 -1990 -х гг. вместе с возникновением первых проектов по секвенированию геномов некоторых организмов. Первым был полностью секвенирован геном бактериофага в 1977 году. Следующим этапным событием было секвенирование генома бактерии Haemophilus influenzae. После этого были полностью секвенированы геномы ещё нескольких видов, включая геном человека. Развитие этого направления стало возможно не только благодаря совершенствованию биохимических методик, но и благодаря появлению более мощной вычислительной техники, которая позволила работать с огромными массивами данных. Протяженность геномов у живых организмов подчас измеряется миллиардами пар оснований. Например, объём генома человека составляет порядка 3 млрд пар оснований. Самый крупный из известных (на начало 2010 года) геномов принадлежит одному из видов двоякодышащих рыб (примерно 110 млрд пар). Структурная геномика – содержание и организация геномной информации. Функциональная геномика это реализация информации, записанной в геноме, от гена к признаку. Сравнительная геномика (эволюционная) – сравнительные исследования содержания и организации геномов разных организмов.

Получение полных последовательностей геномов позволило пролить свет на степень различий между геномами разных живых организмов. Ниже в таблице представлены предварительные данные о сходстве геномов разных организмов с геномом человека. Сходство дано в процентах (отражает долю пар оснований, идентичных у двух сравниваемых видов). Вид Сходство в % Примечания и источники Человек 99, 9 Human Genome Project Однояйцевые близнецы 100 98, 4 Americans for Medical Progress; Jon Entine в San Francisco Examine 98, 7 Richard Mural из Celera Genomics, цитируется в MSNBC Шимпанзе Бонобо – карликовый шимпанзе Горилла 98, 38 То же, что и для шимпанзе. American Journal of Human Genetics, февраль 2001, 682, стр. 444 -456 Мышь 98 Americans for Medical Progress Собака 95 Americans for Medical Progress Круглый червь (нематода) Caenorhabditis elegans 74 Jon Entine в San Francisco Examiner Банан 50 Americans for Medical Progress Нарцисс 35 Steven Rose в The Guardian от 22 января 2004
 • В догеномную эру были установлены фундаментальные принципы молекулярной")
Сравнительная геномика (по Е. Кунину) • В догеномную эру были установлены фундаментальные принципы молекулярной эволюции и было сделано немало конкретных наблюдений, имеющих большое значение и повлиявших на основы эволюционной биологии. Но масштабные работы по расшифровке геномов, начавшиеся в середине 90 -х и стремительно развивавшиеся в новом тысячелетии, качественно изменили всю эволюционную биологию. Важность обширной базы данных геномных последовательностей, имеющих различную степень расхождения, очевидна. Этот материал дает исследователям возможность изучать механизмы и отдельные события эволюции с необходимой статистической точностью и выявлять даже самые малозаметные эволюционные подвижки. Как бы то ни было, в эволюционной биологии получение разнообразных и полных геномных последовательностей чрезвычайно важно далеко не только ради накопления количества данных. Действительно, лишь полностью расшифрованный геном (в отличие от, скажем, расшифрованного лишь на 95 процентов) дает исследователю объективное и непредвзятое представление о генном репертуаре той или иной формы жизни. Иными словами, исследователь может определить присутствие в организме тех или иных генов и, что одинаково важно, их отсутствие. Таким образом, сравнение полных геномов представляет собой единственный удовлетворяющий исследователя путь к реконструкции эволюции. Открывающаяся картина во многом отличается от всего, что можно было себе представить, оставаясь в рамках традиционной эволюционной биологии.

Текст из книги Е. Кунина • • Если мы действительно стремимся «понять» эволюцию, принципиально важно исследовать геномные образцы как вглубь (для этого необходимы геномные последовательности множества близкородственных представителей одного и того же таксона), так и вширь (для этой цели нужны последовательности как можно большего числа различных таксонов – в идеале всех таксонов). Ко времени написания этих строк, в последние дни 2010 года, собрание секвенированных геномов состояло из нескольких тысяч геномов вирусов, более чем тысячи геномов бактерий и архей, а также приблизительно сотни геномов эукариот. Ко времени издания этой книги геномная база данных почти удвоится, а благодаря новому поколению методов секвенирования в предстоящие годы ее темпы роста должны еще более ускориться. Несмотря на то, что не все основные таксоны должным образом охвачены, быстро пополняющееся собрание геномов все более отвечает потребностям исследований как в области микроэволюции, так и в области макроэволюции. Успехи традиционной геномики дополняют и стремительно накапливающиеся в последнее время, обширные по объему данные по метагеномике – а именно всеобъемлющее (или, по меньшей мере, обширное) секвенирование нуклеиновых кислот форм жизни из разнообразных сред обитания. Хотя применяемые в настоящее время в метагеномике подходы обычно не обеспечивают полную расшифровку геномов, они предоставляют бесценную, объективную информацию о разнообразии жизни в различных средах.

Поразительное разнообразие геномов • • Геном стал первым термином с окончанием «-ом» – и до сих пор является наиболее употребительным термином этой группы. Как это всегда бывает в биологии, определить, что же такое геном, нелегко. Говоря просто, геном – это генетическая информация конкретного организма во всей ее полноте. Существование стабильного ядра унаследованной генетической информации (а более конкретно, генов) вытекает из самого факта существования надежной наследственности, а в терминах более фундаментальных – из принципа подверженной ошибкам репликации (ПОР). Однако связь между «генетической информацией во всей ее полноте» и «стабильным ядром» не так уж проста. Стоит, к примеру, задать на первый взгляд невинный вопрос: «Что есть геном кишечной палочки Escherichia coli» – как тут же возникает целый ряд серьезных затруднений. А вопрос «Что такое геном человека? » вызывает свои, не менее сложные проблемы. Новая эра геномики наступила на исходе лета 1995 года. Тогда лаборатория Дж. Крейга Вентера опубликовала результаты секвенирования генома условно-патогенной бактерии гемофильного гриппа Haemophilus influenzae. В процессе расшифровки геномной последовательности Н. influenzae Вентер, Гамильтон Смит и их коллеги усовершенствовали так называемый «метод дробовика» . Этот подход грубого деления генома на короткие произвольные участки с расшифровкой их по частям и последующим восстановлением полной геномной последовательности быстро превратил секвенирование длинных нуклеотидных цепочек в рутинное дело. В течение года были расшифрованы геномы некоторых других бактерий, первый геном археи (Methanocaldococcus jannaschii) и первый геном эукариота (пекарские дрожжи Saccharomyces cerevisiae). К 1999 году установился стабильный экспоненциальный рост коллекции секвенированных геномов.
 • • В диапазоне от вирусов до животных геномы различаются")
Поразительное разнообразие геномов (2) • • В диапазоне от вирусов до животных геномы различаются по размеру на шесть порядков – от нескольких тысяч до нескольких миллиардов нуклеотидов; для клеточных организмов, исключая вирусы, ширина диапазона составляет четыре порядка. По количеству генов диапазон значительно уже и составляет всего около четырех порядков, от двух-трех генов у простейших вирусов до приблизительно 40 тысяч генов у некоторых животных. Если же исключить вирусы и паразитические (симбиотические) бактерии, диапазон по числу генов становится довольно узким, немногим более одного порядка. Кажется весьма удивительным, что млекопитающие или цветковые растения имеют всего примерно в десять раз больше (легко идентифицируемых) генов, чем какая-нибудь средняя свободно живущая бактерия, и лишь примерно в два раза больше, чем бактерия из разряда наиболее сложных. Грубо говоря, геномы могут быть разделены на два четко выделенных класса. 1. Геномы со строгим соответствием между размером генома и числом генов. К ним относятся геномы всех вирусов и прокариот, имеющие огромную плотность генов от 0, 5 до 2 генов на тысячу пар оснований и очень короткие участки между генами (10 -15 процентов длины генома и даже меньше), состоящие главным образом из регуляторных элементов, они почти полностью состоят из легко определяемых генов. Геномы большинства одноклеточных эукариот демонстрируют несколько меньшую зависимость между размером генома и числом генов, чем геномы вирусов и прокариот, тем не менее, они могут быть отнесены к этому же классу.
 • • 2. Геномы, у которых нет четкой взаимосвязи между")
Поразительное разнообразие геномов (3) • • 2. Геномы, у которых нет четкой взаимосвязи между размером генома и числом генов, в частности большие геномы многоклеточных и некоторых одноклеточных эукариот. Здесь в лучшем случае наблюдается слабая корреляция между общим размером генома и числом генов. Соответственно, доля генома, занимаемая межгенными участками (а также другими некодирующими последовательностями, такими как интроны), сильно варьирует. В некоторых наиболее сложных геномах, в частности у млекопитающих, основную часть генома составляют именно некодирующие последовательности. Вариабельность размеров генома и числа генов дополняется разнообразием в других измерениях – например, в физической организации и композиции нуклеотидов. При рассмотрении как вирусной, так и клеточной формы жизни геномы предстают во всевозможных формах нуклеиновых кислот. Все геномы клеточных организмов состоят из двухцепочечных ДНК, однако количество геномных сегментов (хромосом) и их размеры, форма (кольцевая или линейная), а также плоидность (число наборов) широко разнятся. Азбучная истина гласит, что прокариоты имеют гаплоидные, простые кольцевые хромосомы, в то время как у геномов эукариот, сильно различающихся по плоидности, гены распределены между множеством линейных хромосом. И хотя такие геномные формы, по-видимому, действительно доминируют, на самом деле разнообразие геномов выходит далеко за рамки такого простого дихотомического разделения. В частности, у многих прокариот имеется несколько хромосом, в отдельных случаях – линейных. Более того, вопреки распространенному заблуждению, у прокариот большинство клеток не гаплоидные, то есть они содержат несколько копий генома.

Древние гены составляют в геноме большинство и имеют отчетливую эволюционную судьбу • • Эрнст Майр , великий эволюционист XX века и один из основателей СТЭ, с уверенностью предсказывал исходя из больших фенотипических различий между организмами, что гены разных организмов, даже близкородственных, не будут иметь узнаваемого сходства. Ошибочность этого предсказания оказалась просто феерической, что само по себе делает его нетривиальным и ценным. Сравнение последовательностей даже в догеномный период выявило высокий консерватизм последовательностей у некоторых гомологичных белков и молекул некодирующих РНК по всему спектру жизни, от бактерий до млекопитающих. Более того, высокая степень сходства последовательностей существует у древних паралогов, которые, по-видимому, происходят от копий, ведущих свое происхождение от LUCA. Геномика позволяет перевести это общее понимание в количественное разбиение генов любого генома на классы эволюционной консервативности. Ключевое открытие сравнительной геномики состоит в том, что большинство генов в каждом геноме могут считаться высококонсервативными – они имеют легко обнаруживаемые гомологи в организмах, разделяемых сотнями миллионов лет эволюции (например, в случае генов человека, на уровне общего предка позвоночных (см. рис. ). Это открытие демонстрирует поразительную устойчивость последовательностей РНК и белков в процессе эволюции: типичное время исчезновения сходства последовательностей у гомологичных генов сравнимо со временем существования жизни на Земле. Помимо основополагающего значения, данный факт имеет огромные практические последствия: благодаря ему, прежде всего, сравнительная геномика становится крайне информативной и действенной.
. Распределение генов по эволюционному возрасту.")
(Левая диаграмма – аспергилус фумигатус, правая – человек ). Распределение генов по эволюционному возрасту. «Эволюционный возраст» соответствует самому старому таксономическому узлу, в котором могут быть определены гомологи для белка, производимого данным геном. В частности для человека древние всеобщие означает «гомологи, обнаруживаемые у прокариот» , древние зукариотические означает «гомологи обнаруживаемые у вне супергруппы униконтов» , царство и тип означает «гомологи, обнаруживаемые у животных вне класса млекопитающих» , а «класс и моложе» означает «вне класса млекопитающих гомологи не обнаружены» (из Кунин, 2014)

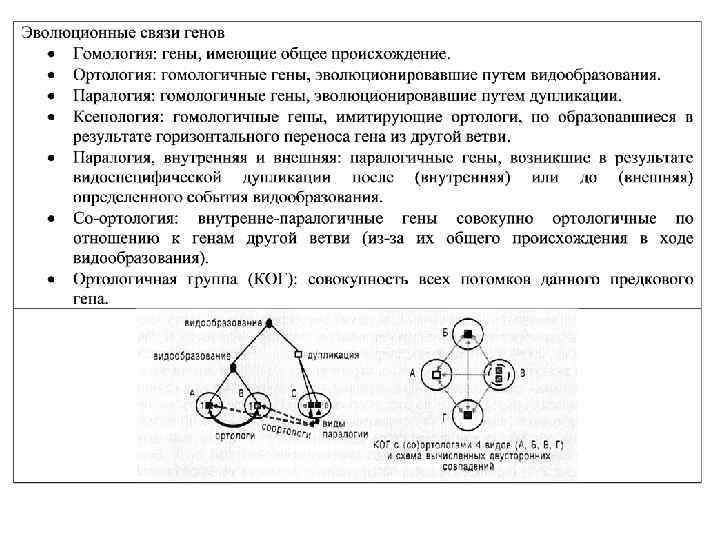
Структуру эволюционного процесса определяют не только консервативные последовательности. На протяжении чрезвычайно длительных эволюционных периодов не просто сохраняется сходство последовательностей РНК и белков, но и гены имеют свойство сохранять свою уникальность. Иными словами, большинство генов развиваются как ортологичные линии, с редкими случаями дупликации. Устойчивость ортологии генов становится очевидной благодаря простой процедуре, широко применяемой в сравнительной геномике и позволяющей эффективно выявлять ортологичные наборы генов. При этом ортологи обнаруживаются как «наилучшие совпадения при двунаправленном сравнении» (bidirectional best hits): все закодированные в геноме белковые последовательности сравниваются со всеми белками, закодированными в другом геноме, а затем процедура повторяется в обратном направлении. Пары генов, дающие наилучшие совпадения (те, которые демонстрируют наибольшее сходство последовательностей) при обоих направлениях сравнения, считаются возможными ортологами; нетрудно применить эту процедуру к нескольким видам путем совмещения треугольников двунаправленных совпадений, имеющих общую сторону (см. табл. 5. 2). Примечательно, что такой прямолинейный подход в большинстве случаев хорошо срабатывает: к примеру, порядка 70 процентов генов организмов, разделенных приблизительно 100 миллионами лет эволюции, таких как люди и мыши, легко идентифицируются как ортологи при помощи описанной процедуры.

Если применить простую модификацию этого алгоритма и включить дупликации генов, характерных для одной линии наследования (дупликации, образовавшиеся после расхождения сравниваемых видов), такой подход позволяет идентифицировать наборы ортологов (известных как кластеры ортологичных генов, КОГ) во многих геномах, в том числе столь удаленных друг от друга, как археи и бактерии – представители двух доменов прокариот. Более точные и мощные способы обнаружения ортологов требуют подробного анализа филогенетических деревьев; впрочем, результаты такого анализа обычно близки к тем, что дают более простые методы, основанные только на сравнении последовательностей. Разумеется, для части генов история дупликаций и потерь настолько сложна, что обнаружить КОГ трудно, поэтому они становятся нечеткими кластерами с неопределенной внутренней структурой. По счастью, этих «трудных» генов в каждом геноме относительно немного.

Размер генома и сложность организмов • • • Данный раздел представляет собой выдержки из книги В. Н. Стегния (2013). С-парадокс Положения Моргана о зависимости между числом генов и сложностью организмов было опубликовано в 1932 году. Позднее, после того как следующим поколением исследователей было установлено, что гены состоят из ДНК, стало возможным получить гораздо более ясное представление о природе генов, однако достигнутые успехи еще больше запутали вопрос о зависимости между числом генов и сложностью организма. В целом измерение количества ДНК, содержащегося в гаплоидном геноме (значение С) у большого числа самых разнообразных организмов, указывает на повышение содержания ДНК с увеличением сложности, однако величина генома варьирует так сильно, что у многих морфологически примитивных организмов геномы оказались значительно больше, чем у морфологически более продвинутых форм. Это явление, получило название парадокса значений С, или С-парадокса. Как выше было показано, у прокариот преобладают структурные гены (~95% от геномной ДНК), тогда как у эукариот их часть составляет ~5% от геномной ДНК.


Число клеточных типов. Любая схема опасна тем, что ее можно принять за своего рода лестницу живых существ, созданную в XX в. , и в известном смысле так оно и есть. Прямое сравнение относительной сложности форм, обладающих различной морфологической организацией, неизбежно будет субъективным. Есть, однако, два показателя сложности, которые могут служить приближенными мерами: это число типов клеток, различаемых у представителей данной группы организмов, и число терминов, используемых систематиками для их описания. Теоретическое оправдание использованию числа типов клеток дал С. Кауфман (S. Kauffman), выдвинувший положение о том, что число дифференцированных клеточных типов в организме зависит от числа стабильных состояний, создаваемых в результате регуляторных взаимодействий, возможных в пределах того или иного генома. Оценить число клеточных типов у сравнительно несложных организмов относительно просто. Так, у бактерий имеются клетки двух типов (вегетативные клетки и споры), у дрожжей – 3 -4 типа, у водорослей и грибов – примерно 5 типов, у губок – 11, у кишечнополостных – 14 -20, у растений – от 20 до 40, у кольчецов (например, кольчатых червей) – примерно 55. Для более высокоорганизованных животных получить такие оценки труднее; возможно, что оценка Кауфмана, считающего, что в организме человека содержатся клетки 100 разных типов, занижена на целый порядок.

Некоторые выводы • • До настоящего времени нет единого, всеми принятого, представления о том, что такое ген. Неоднозначность этого понятия проявляется во множестве аспектов. Например, что является объектом эволюции: гены (геномы) или организмы (фенотипы)? Есть ли обратная связь между геном и фенотипом, и ели есть, то на сколько она сильна и устойчива. Каковы механизмы возникновения задерживающихся мутаций? Насколько мутации случайны, а если не всегда случайны, то каков механизм их детерминированности? Какую роль в организации и функциях ДНК играют диминуция хроматина и Спарадокс. Перечисление вопросов подобного рода могло бы занять много времени. В заключение раздела «Ген» можно прийти к следующему выводу. По мере развития генетики и учения об онтогенезе понятие «ген» расширяется и теряет легко формулируемую определённость. Соответственно и понятие генетического контроля, как в онтогенезе, так и в филогенезе, существенно усложняется. Существование такого контроля не вызывает сомнений, однако содержание этого понятия включает и весьма сложные межгенные взаимовлияния, и регуляторную роль кодируемых генами белков (обратная связь), и влияние комплекса цитоплазматических генов, и горизонтальный перенос, и эпигенетические факторы, и вырожденность генетического кода, и, наконец, взаимосвязь со средой, как внутри организма, так вне его.
 • Определение гена, как последовательности нуклеотидов, кодирующей определённй белок, остаётся принципиально")
Некоторые выводы (2) • Определение гена, как последовательности нуклеотидов, кодирующей определённй белок, остаётся принципиально правильным, однако в подавляющем большинстве случаев, между информацией, закодированной в ДНК и её фенотипическим проявлением иногда лежит столь длинный и осложненный множеством превходящих обстоятельств путь, что считать ген однозначно определяющим конкретный фенотипический признак можно далеко не всегда, а точнее говоря, лишь в очень немногих случаях. В этом смысле Менделю, а, следовательно, и генетике, в своё время просто повезло.
Биология_5_Гены (2)Мутации.pptx