

Лекция4_генные мутации_хромосомные карты.ppt
- Количество слайдов: 62
 Генные мутации Характеристика генных мутаций Хромосомные карты Методы исследования генома человека
Генные мутации Характеристика генных мутаций Хромосомные карты Методы исследования генома человека
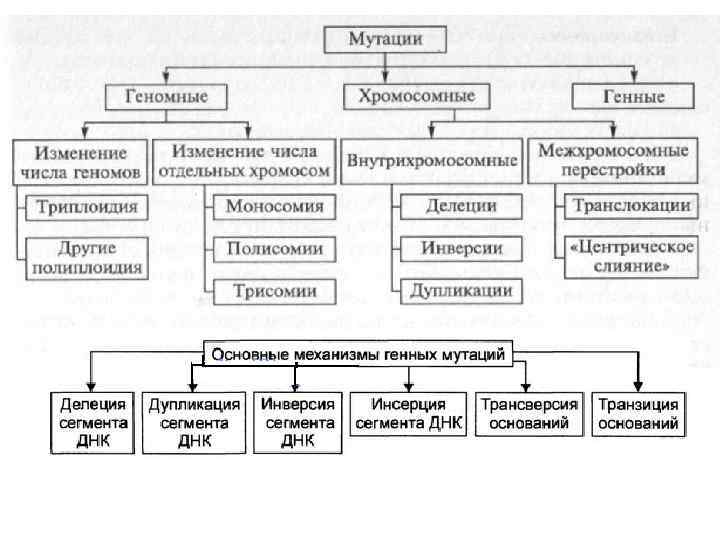


 мутации изменения числа и/или - это последовательности нуклеотидов в структуре ДНК (вставки,") Генные (точечные) мутации изменения числа и/или - это последовательности нуклеотидов в структуре ДНК (вставки, выпадения, перемещения, замещения нуклеотидов) в пределах отдельных генов, приводящие к изменению количества или качества соответствующих белковых продуктов. Выделяют несколько типов точечных мутаций: • Точечные мутации замены оснований • Точечные мутации сдвига рамки чтения • сложные мутации
Генные (точечные) мутации изменения числа и/или - это последовательности нуклеотидов в структуре ДНК (вставки, выпадения, перемещения, замещения нуклеотидов) в пределах отдельных генов, приводящие к изменению количества или качества соответствующих белковых продуктов. Выделяют несколько типов точечных мутаций: • Точечные мутации замены оснований • Точечные мутации сдвига рамки чтения • сложные мутации
 Точечные мутации замены оснований Транзиция мутация замены оснований, когда одно пуриновое основание замещается на другое пуриновое основание (аденин на гуанин или наоборот), либо пиримидиновое основание на другое пиримидиновое основание (тимин на цитозин или наоборот. Трансверсия мутация замены оснований, когда одно пуриновое основание замещается на пиримидиновое основание или наоборот).
Точечные мутации замены оснований Транзиция мутация замены оснований, когда одно пуриновое основание замещается на другое пуриновое основание (аденин на гуанин или наоборот), либо пиримидиновое основание на другое пиримидиновое основание (тимин на цитозин или наоборот. Трансверсия мутация замены оснований, когда одно пуриновое основание замещается на пиримидиновое основание или наоборот).
 Точечные мутации сдвига рамки чтения: Делеции Инсерция мутация сдвига рамки чтения, когда в молекуле ДНК выпадает один или несколько нуклеотидов мутация сдвига рамки чтения, когда в молекулу ДНК встраивается один или несколько нуклеотидов Сложные мутации Это такие изменения ДНК, когда один её участок заменяется участком другой длины и другого нуклеотидного состава.
Точечные мутации сдвига рамки чтения: Делеции Инсерция мутация сдвига рамки чтения, когда в молекуле ДНК выпадает один или несколько нуклеотидов мутация сдвига рамки чтения, когда в молекулу ДНК встраивается один или несколько нуклеотидов Сложные мутации Это такие изменения ДНК, когда один её участок заменяется участком другой длины и другого нуклеотидного состава.
, которые (от появляться напротив повреждений") Точечные мутации бывают: 1. мишенными мутациями слова «мишень» ), которые (от появляться напротив повреждений молекулы ДНК и способны останавливать синтез ДНК. Например, напротив циклобутановых пиримидиновых димеров. Циклобутановые пиримидиновые димерывызывают • мишенные мутации замены оснований • мишенные мутации сдвига рамки 2. немишенными мутациями которые образуются на , неповрежденных участках ДНК, часто в небольшой окрестности от фотодимеров. • немишенными мутациями замены оснований • немишенными мутациями сдвига рамки 3. задерживающиеся мутации , которые образуются не сразу после воздействия мутагена, а появляются иногда после десятков цик репликаций.
Точечные мутации бывают: 1. мишенными мутациями слова «мишень» ), которые (от появляться напротив повреждений молекулы ДНК и способны останавливать синтез ДНК. Например, напротив циклобутановых пиримидиновых димеров. Циклобутановые пиримидиновые димерывызывают • мишенные мутации замены оснований • мишенные мутации сдвига рамки 2. немишенными мутациями которые образуются на , неповрежденных участках ДНК, часто в небольшой окрестности от фотодимеров. • немишенными мутациями замены оснований • немишенными мутациями сдвига рамки 3. задерживающиеся мутации , которые образуются не сразу после воздействия мутагена, а появляются иногда после десятков цик репликаций.
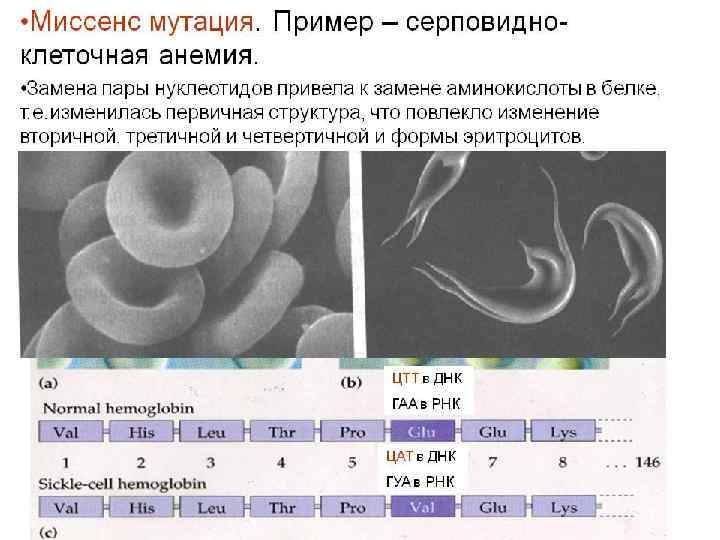
 нейтральные мутации (молчащая мутация) – сохранение смысла") Возможны четыре генетических последствия точечн мутаций: 1) нейтральные мутации (молчащая мутация) – сохранение смысла кодона, не имеет фенотипического выражения из-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) миссенс-мутация изменение смысла кодона, замена нуклеотида в кодирующей части гена, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи К примеру, миссенс-мутация 553 -м кодоне гена FAC, приводящая к замене в лейцина на пролин, делает продукт этого гена неспособным комплементировать функциональный дефект в клетках больных анемией. Фанкони.
Возможны четыре генетических последствия точечн мутаций: 1) нейтральные мутации (молчащая мутация) – сохранение смысла кодона, не имеет фенотипического выражения из-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) миссенс-мутация изменение смысла кодона, замена нуклеотида в кодирующей части гена, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи К примеру, миссенс-мутация 553 -м кодоне гена FAC, приводящая к замене в лейцина на пролин, делает продукт этого гена неспособным комплементировать функциональный дефект в клетках больных анемией. Фанкони.
 В генетическом коде имеются три бессмысленных кодона: амбер UAG — охр — UAA") 3) В генетическом коде имеются три бессмысленных кодона: амбер UAG — охр — UAA опал — UGA в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов — например амбер-мутация 4) обратная заменастоп-кодонана смысловой кодон). (
3) В генетическом коде имеются три бессмысленных кодона: амбер UAG — охр — UAA опал — UGA в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов — например амбер-мутация 4) обратная заменастоп-кодонана смысловой кодон). (

 По влиянию на экспрессию генов мутации разделяют на две категории: - мутации типа замен пар оснований -Мутации типа сдвига рамки считывания (frameshift). Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трём, что связано с триплетностью генетического кода. Первичную мутацию иногда называют прямой мутацией , а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны. Почковые мутации (спорты) — стойкие соматические мутации происходящие в клетках точек роста растений. Приводят к клоновой изменчивости. При вегетативном размножении сохраняются. Многие сорта культурных растений являются почковыми мутациями.
По влиянию на экспрессию генов мутации разделяют на две категории: - мутации типа замен пар оснований -Мутации типа сдвига рамки считывания (frameshift). Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трём, что связано с триплетностью генетического кода. Первичную мутацию иногда называют прямой мутацией , а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны. Почковые мутации (спорты) — стойкие соматические мутации происходящие в клетках точек роста растений. Приводят к клоновой изменчивости. При вегетативном размножении сохраняются. Многие сорта культурных растений являются почковыми мутациями.
 С точки зрения структурно-функциональной организации генов , происходящие внутри них замены, вставки, выпадения, перемещения нуклеотидов можно объединить в следующие группы: 1) мутации в регуляторных областях генов • мутации в промоторной части снижают уровень синтеза белкового продукта; (например, регуляторном элементе с последовательностью Pu. CCC и внутри ТАТА-бокса у гена р-глобина) • мутации в сайте полиаденилирования снижают уровень транскрипции (характерно для афроамериканцев, страдающих талассемией) Таким образом, мутации в регуляторных 5' и 3'-нетранслируемы областях генов вызывают количественные изменения соответствующих продуктов и проявляются фенотипически (клинически) в зависимости от порогового уровня белков, при котором их функция еще сохраняется;
С точки зрения структурно-функциональной организации генов , происходящие внутри них замены, вставки, выпадения, перемещения нуклеотидов можно объединить в следующие группы: 1) мутации в регуляторных областях генов • мутации в промоторной части снижают уровень синтеза белкового продукта; (например, регуляторном элементе с последовательностью Pu. CCC и внутри ТАТА-бокса у гена р-глобина) • мутации в сайте полиаденилирования снижают уровень транскрипции (характерно для афроамериканцев, страдающих талассемией) Таким образом, мутации в регуляторных 5' и 3'-нетранслируемы областях генов вызывают количественные изменения соответствующих продуктов и проявляются фенотипически (клинически) в зависимости от порогового уровня белков, при котором их функция еще сохраняется;
 мутации в кодирующих областях генов • мутации в экзонахмогут приводить к преждевременному окончанию") 2) мутации в кодирующих областях генов • мутации в экзонахмогут приводить к преждевременному окончанию белкового синтеза. (к примеру, в результате мутаций внутри экзона гемоглобина белковая цепь оказывается укороченной и не обладает активностью) • мутации в интронахспособны генерировать новые сайты сплайсинга, которые, конкурируя с нормальными (исходными), в итоге, заменяют их. Возникновение замен в гене гемоглобина, замедляющих сплайсинг, известно и для В 0 -, и для В+-талассемии. • мутации в сайтах сплайсинга(на стыках экзонов и нитронов), нарушают процессинг первичного РНК-транскрипта и приводят к трансляции бессмысленных белков: удлиненного при неправильном вырезании интронов либо укороченного при вырезании экзонов. Так, мутация сдвига рамки считывания в акцепторном участке сплайсинга гена ХРА приводит к полной инактивации белка и, как следствие, к развитию тяжелой формы пигментной ксеродермы. Замены нуклеотидов в кодирующих областях генов, не сопровождающиеся заменами аминокислот в силу вырожденност генетического кода, приводят к нейтральным мутациям , не оказывающим заметного влияния ни на функцию соответствующе белка, ни на его структуру.
2) мутации в кодирующих областях генов • мутации в экзонахмогут приводить к преждевременному окончанию белкового синтеза. (к примеру, в результате мутаций внутри экзона гемоглобина белковая цепь оказывается укороченной и не обладает активностью) • мутации в интронахспособны генерировать новые сайты сплайсинга, которые, конкурируя с нормальными (исходными), в итоге, заменяют их. Возникновение замен в гене гемоглобина, замедляющих сплайсинг, известно и для В 0 -, и для В+-талассемии. • мутации в сайтах сплайсинга(на стыках экзонов и нитронов), нарушают процессинг первичного РНК-транскрипта и приводят к трансляции бессмысленных белков: удлиненного при неправильном вырезании интронов либо укороченного при вырезании экзонов. Так, мутация сдвига рамки считывания в акцепторном участке сплайсинга гена ХРА приводит к полной инактивации белка и, как следствие, к развитию тяжелой формы пигментной ксеродермы. Замены нуклеотидов в кодирующих областях генов, не сопровождающиеся заменами аминокислот в силу вырожденност генетического кода, приводят к нейтральным мутациям , не оказывающим заметного влияния ни на функцию соответствующе белка, ни на его структуру.

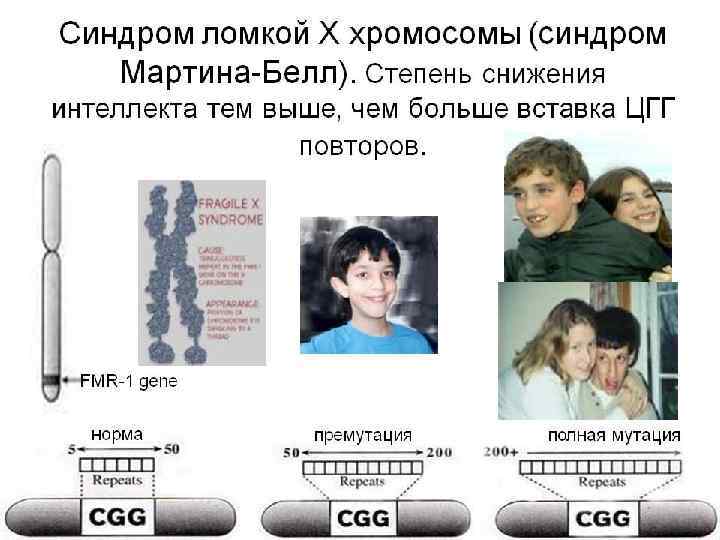
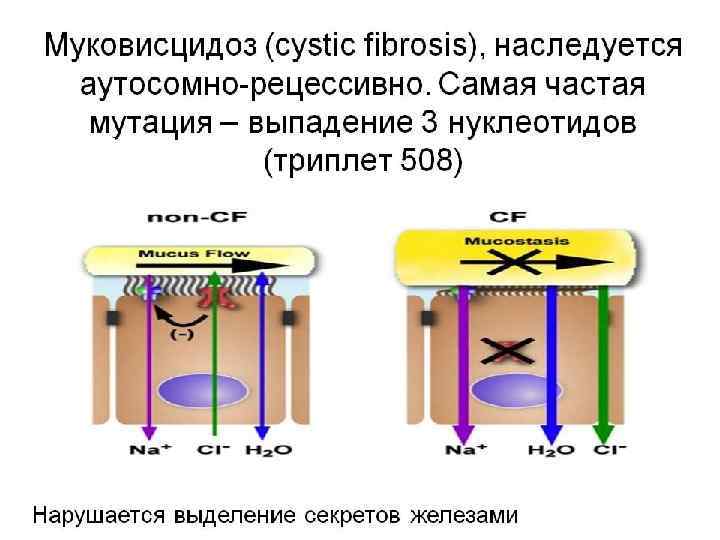

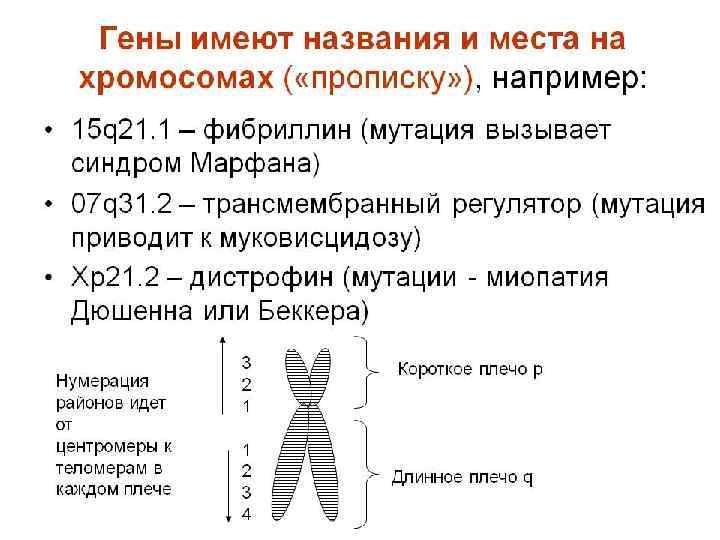
 ХРОМОСОМНЫЕ КАРТЫ Графическое изображение взаимного расположения и относительных расстояний между генами определенных хромосом находящихся в одной , группе сцепления. Графическое представление нормального человеческого кариотипа
ХРОМОСОМНЫЕ КАРТЫ Графическое изображение взаимного расположения и относительных расстояний между генами определенных хромосом находящихся в одной , группе сцепления. Графическое представление нормального человеческого кариотипа
 Впервые в 913 — 1915 годах 1 на возможность построения генетических карт хромосом указывают Морган Т. и его сотрудники. Они экспериментально показали, что основываясь на явлениях цепления с генов и кроссинговера можно построить генетические карты хромосом.
Впервые в 913 — 1915 годах 1 на возможность построения генетических карт хромосом указывают Морган Т. и его сотрудники. Они экспериментально показали, что основываясь на явлениях цепления с генов и кроссинговера можно построить генетические карты хромосом.
 • Морган представлял себе гены упорядоченными по длине хромосом, как бусинки в ожерелье • Экспериментальные данные привели его к идее создания генетических карт хромосом • Очевидно, что чем дальше находятся два гена друг от друга, тем больше вероятность разрыва нити, связывающей их, и получения новых сочетаний генов • Стало возможным определить относительное расстояние между генами в хромосоме путем простого расчета процента кроссинговера 25
• Морган представлял себе гены упорядоченными по длине хромосом, как бусинки в ожерелье • Экспериментальные данные привели его к идее создания генетических карт хромосом • Очевидно, что чем дальше находятся два гена друг от друга, тем больше вероятность разрыва нити, связывающей их, и получения новых сочетаний генов • Стало возможным определить относительное расстояние между генами в хромосоме путем простого расчета процента кроссинговера 25
 Возможность картирования основана на постоянстве процента кроссинговера между определенными генами, позволяет локализовать их. Единицей расстояния между генами служит 1 % кроссинговера; в честь Моргана эта единица называется морганида (М), или сантиморганида (с. М). 1 с. М (сентиморганида– единица = расстояния между генами, при которой вероятност кроссинговера 1%, соответствует примерно 1 мегабазе. Гаплоидный геном человека составляет примерно 3 300 000, т. е. 3300 с. М
Возможность картирования основана на постоянстве процента кроссинговера между определенными генами, позволяет локализовать их. Единицей расстояния между генами служит 1 % кроссинговера; в честь Моргана эта единица называется морганида (М), или сантиморганида (с. М). 1 с. М (сентиморганида– единица = расстояния между генами, при которой вероятност кроссинговера 1%, соответствует примерно 1 мегабазе. Гаплоидный геном человека составляет примерно 3 300 000, т. е. 3300 с. М
 карты –отмечено линейное расположение генов, установленное на основании частоты") Хромосомные карты • Генетические (теоретические) карты –отмечено линейное расположение генов, установленное на основании частоты обменов между гомологичными хромосомами • Цитологические (реальные) – составленные на основании цитологических исследований особей с хромосомными мутациями • Физические графическое представление порядка – следования физических маркеров (фрагментов молекулы ДНК), расстояние между которыми определяется в парах нуклеотидов (основаны на точном расстоянии в базах, кило-, мега- и гига базах). • Рестрикционные вид физической карты, на которой – указан порядок следования и расстояния между сайтами расщепления ДНК рестриктазами (обычно участок узнавания рестриктазы 4 -6 п. н. ).
Хромосомные карты • Генетические (теоретические) карты –отмечено линейное расположение генов, установленное на основании частоты обменов между гомологичными хромосомами • Цитологические (реальные) – составленные на основании цитологических исследований особей с хромосомными мутациями • Физические графическое представление порядка – следования физических маркеров (фрагментов молекулы ДНК), расстояние между которыми определяется в парах нуклеотидов (основаны на точном расстоянии в базах, кило-, мега- и гига базах). • Рестрикционные вид физической карты, на которой – указан порядок следования и расстояния между сайтами расщепления ДНК рестриктазами (обычно участок узнавания рестриктазы 4 -6 п. н. ).
 Последовательности, распознаваемы разнымирестриктазами • Eco. RI • Г ААТТЦ • ЦТТАА Г • Sma. I • ЦЦЦ ГГГ • ГГГ ЦЦЦ ДНК разрезают рестриктазами и подвергают электрофорезу. Рестрикционная карта - вид физической карты, на которой указаны расстояния между соседними сайтами расщепления ДНК определенной рестриктазой. Маркерами этой карты являются рестрикционные фрагменты/сайты рестрикции.
Последовательности, распознаваемы разнымирестриктазами • Eco. RI • Г ААТТЦ • ЦТТАА Г • Sma. I • ЦЦЦ ГГГ • ГГГ ЦЦЦ ДНК разрезают рестриктазами и подвергают электрофорезу. Рестрикционная карта - вид физической карты, на которой указаны расстояния между соседними сайтами расщепления ДНК определенной рестриктазой. Маркерами этой карты являются рестрикционные фрагменты/сайты рестрикции.
 • Анализ родословных") Методы создания хромосомных карт • Гибридологический (по результатам скрещиваний Т. Морган) • Анализ родословных (генеалогический) • Генетика соматических клеток* • ДНК-зондов*
Методы создания хромосомных карт • Гибридологический (по результатам скрещиваний Т. Морган) • Анализ родословных (генеалогический) • Генетика соматических клеток* • ДНК-зондов*
 Генетические и физические карты хромосом • Генетическое картирование основывается на использовании генетических методов для построения карт, указывающих позиции генов и других последовательностей в геноме. • Генетические методы включают гибридологические эксперименты или, в случае с людьми, генеалогический метод (анализ родословных) 30
Генетические и физические карты хромосом • Генетическое картирование основывается на использовании генетических методов для построения карт, указывающих позиции генов и других последовательностей в геноме. • Генетические методы включают гибридологические эксперименты или, в случае с людьми, генеалогический метод (анализ родословных) 30
 • Физическое картирование использует молекулярнобиологические методы для непосредственного исследования молекул ДНК и создания карт, обозначающих позиции определенных последовательностей, в том числе генов 31
• Физическое картирование использует молекулярнобиологические методы для непосредственного исследования молекул ДНК и создания карт, обозначающих позиции определенных последовательностей, в том числе генов 31
 Опорные точки карт хромосом – гены и ДНК-маркеры • Гены – часто используемые как маркеры - не идеальны. Одна из проблем (особенно для больших геномов позвоночных) - карты, основанные на генах, не очень детализованы • Необходимы другие типы маркеров • Опорные точки карт, не являющиеся генами, называются ДНК-маркерами • Основные типы ДНК-маркеров: – полиморфизм длины рестрикционных фрагментов (RFLPs) – полиморфизм длины простой последовательности (SSLPs) – однонуклеотидный полиморфизм (SNPs) 32
Опорные точки карт хромосом – гены и ДНК-маркеры • Гены – часто используемые как маркеры - не идеальны. Одна из проблем (особенно для больших геномов позвоночных) - карты, основанные на генах, не очень детализованы • Необходимы другие типы маркеров • Опорные точки карт, не являющиеся генами, называются ДНК-маркерами • Основные типы ДНК-маркеров: – полиморфизм длины рестрикционных фрагментов (RFLPs) – полиморфизм длины простой последовательности (SSLPs) – однонуклеотидный полиморфизм (SNPs) 32
 • А. Стертевант в 1913 г. составил первую генетическую карту локализации генов в Ххромосоме дрозофилы • Генетические карты уже разработаны для дрозофилы, мыши, нейроспоры; для высших растений: кукурузы, риса, ячменя и др. Альфред Стертевант (1891 -1970) 33
• А. Стертевант в 1913 г. составил первую генетическую карту локализации генов в Ххромосоме дрозофилы • Генетические карты уже разработаны для дрозофилы, мыши, нейроспоры; для высших растений: кукурузы, риса, ячменя и др. Альфред Стертевант (1891 -1970) 33
 Построение генетической карты на основании частот рекомбинации Пример показывает реальные эксперименты, . выполненные Артуром Стуртевантом на плодовой мушке. Все 4 гена находятся в Х-хромосоме плодовой мушки. Показаны частоты рекомбинации между генами и относительное взаиморасположение генов на карте 34
Построение генетической карты на основании частот рекомбинации Пример показывает реальные эксперименты, . выполненные Артуром Стуртевантом на плодовой мушке. Все 4 гена находятся в Х-хромосоме плодовой мушки. Показаны частоты рекомбинации между генами и относительное взаиморасположение генов на карте 34
 Генеалогический анализ в составлении генетических карт человека • Для человека невозможно заключение экспериментальных браков с целью создания генетических карт • Данные для расчета частот рекомбинации могут быть получены исследованием генотипов членов поколений существующих семей • Это значит, что доступными являются только ограниченные данные, и их интерпретация часто усложняется, т. к. браки людей редко приводят к необходимым «скрещиваниям» , а часто генотипы одного или большего числа членов семьи являются недоступными в связи со смертью или отказом от сотрудничества 35
Генеалогический анализ в составлении генетических карт человека • Для человека невозможно заключение экспериментальных браков с целью создания генетических карт • Данные для расчета частот рекомбинации могут быть получены исследованием генотипов членов поколений существующих семей • Это значит, что доступными являются только ограниченные данные, и их интерпретация часто усложняется, т. к. браки людей редко приводят к необходимым «скрещиваниям» , а часто генотипы одного или большего числа членов семьи являются недоступными в связи со смертью или отказом от сотрудничества 35
 Родословная показывает наследование генетической болезни в") • Пример анализа родословной человека • (A) Родословная показывает наследование генетической болезни в семье двух живых родителей и 6 детей, а также при наличии информации о родителях матери. Аллель болезни является доминантным по отношению к аллелю здоровья. Реальным является определение степени сцепления между геном болезни и микросателлитом М типированием аллелей для этого микросателлита (M 1, M 2, и т. п. ) у живых членов семьи. (B) Родословная может быть интерпретирована двумя разными путями: Гипотеза 1 дает низкую частоту рекомбинации и свидетельствует, что ген болезни сильно сцеплен с микросателлитом М; Гипотеза 2 подтверждает, что ген и микросателлит менее прочно сцеплены (C) Реконструкция генотипа микросателлита бабушки подтверждает справедливость Гипотезы 1 •
• Пример анализа родословной человека • (A) Родословная показывает наследование генетической болезни в семье двух живых родителей и 6 детей, а также при наличии информации о родителях матери. Аллель болезни является доминантным по отношению к аллелю здоровья. Реальным является определение степени сцепления между геном болезни и микросателлитом М типированием аллелей для этого микросателлита (M 1, M 2, и т. п. ) у живых членов семьи. (B) Родословная может быть интерпретирована двумя разными путями: Гипотеза 1 дает низкую частоту рекомбинации и свидетельствует, что ген болезни сильно сцеплен с микросателлитом М; Гипотеза 2 подтверждает, что ген и микросателлит менее прочно сцеплены (C) Реконструкция генотипа микросателлита бабушки подтверждает справедливость Гипотезы 1 •
 • На генетической карте хромосом обозначают: номер группы сцепления, сокращенные названия генов, расстояние между генами • С помощью генетических карт можно предвидеть характер наследования признаков, определяемых нанесенными генами: теоретически рассчитывать вероятность их расщепления у потомков Физические карты - более точны! 37
• На генетической карте хромосом обозначают: номер группы сцепления, сокращенные названия генов, расстояние между генами • С помощью генетических карт можно предвидеть характер наследования признаков, определяемых нанесенными генами: теоретически рассчитывать вероятность их расщепления у потомков Физические карты - более точны! 37
 Методы физического картирования хромосом Большое число методов физического картирования хромосом основано на 3 главных подходах: – Рестрикционное картирование устанавливающее , относительные позиции на молекуле ДНК последовательностей узнавания для рестрикционных эндонуклеаз – Флуоресцентная situгибридизация in (FISH), при которой расположение маркера картируется гибридизацией образца, который содержит маркер с интактными хромосомами – Картирование последовательностями маркерных сайто (STS-картирование) при котором позиции коротких , последовательностей картируются ПЦР и/или гибридизационным анализом фрагментов генома
Методы физического картирования хромосом Большое число методов физического картирования хромосом основано на 3 главных подходах: – Рестрикционное картирование устанавливающее , относительные позиции на молекуле ДНК последовательностей узнавания для рестрикционных эндонуклеаз – Флуоресцентная situгибридизация in (FISH), при которой расположение маркера картируется гибридизацией образца, который содержит маркер с интактными хромосомами – Картирование последовательностями маркерных сайто (STS-картирование) при котором позиции коротких , последовательностей картируются ПЦР и/или гибридизационным анализом фрагментов генома
 Методы картирования хромосом человека В связи с невозможностью картирования хромосом человека традиционными методами анализирующего скрещивания, приемлемыми для других организмов, с этой целью применяют другие методы Для этого используются три подхода: 1. технология гибридных соматических клеток, 2. активизированная флюоресценцией сортировка хромосом (FACS) – поточная цитометрия 3. и FISH-метод Эти методы обеспечивают грубое картирование, перед более детальным молекулярным анализом и секвенированием.
Методы картирования хромосом человека В связи с невозможностью картирования хромосом человека традиционными методами анализирующего скрещивания, приемлемыми для других организмов, с этой целью применяют другие методы Для этого используются три подхода: 1. технология гибридных соматических клеток, 2. активизированная флюоресценцией сортировка хромосом (FACS) – поточная цитометрия 3. и FISH-метод Эти методы обеспечивают грубое картирование, перед более детальным молекулярным анализом и секвенированием.
 1. Гибридизациясоматических клеток
1. Гибридизациясоматических клеток
 метод гибридизации соматических клеток грызунов и человека в культуре тканей Если изолировать из тела и смешать клетки мыши и человека в культуре, то в результате их слияния можно получить гибридные клетки, содержащие хромосомы одного или другого вида. Клетки мыши имеют 40 хромосом, а клетки человека - 46. Суммарное число хромосом гибридных клеток должно составлять 86, но обычно это не происходит и чаще всего гибридные клетки содержат от 40 до 50 хромосом
метод гибридизации соматических клеток грызунов и человека в культуре тканей Если изолировать из тела и смешать клетки мыши и человека в культуре, то в результате их слияния можно получить гибридные клетки, содержащие хромосомы одного или другого вида. Клетки мыши имеют 40 хромосом, а клетки человека - 46. Суммарное число хромосом гибридных клеток должно составлять 86, но обычно это не происходит и чаще всего гибридные клетки содержат от 40 до 50 хромосом
 • Пример показывает как стабильные «человек-мышь» гибридные соматические клетки могут получаться применением ПЭГ • По понятным причинам хромосомы человека избирательно утрачиваются первичным продуктом слияния • Утрата хромосом человека приводит к образованию большого разнообразия гибридных клеток по набору хромосом человека • Эти клетки могут быть клонированными для получения отдельных клеточных линий со специфическим набором хромосом человека • Идентификация хромосом человека может проводиться методами, основанными на ПЦР с применением хромосомспецифических маркеров 42
• Пример показывает как стабильные «человек-мышь» гибридные соматические клетки могут получаться применением ПЭГ • По понятным причинам хромосомы человека избирательно утрачиваются первичным продуктом слияния • Утрата хромосом человека приводит к образованию большого разнообразия гибридных клеток по набору хромосом человека • Эти клетки могут быть клонированными для получения отдельных клеточных линий со специфическим набором хромосом человека • Идентификация хромосом человека может проводиться методами, основанными на ПЦР с применением хромосомспецифических маркеров 42
 • После размножения интересующих клеток можно провести анализ ферментов , активность которых связана с наличием именно этой хромосомы • Использование методов дифференцированной окраски хромосом позволяет связать гены с определенными локусами хромосом, так как в гибридных клетках довольно частыми являются хромосомные разрывы, перестройки, присутствие не целых хромосом, а отдельных фрагментов
• После размножения интересующих клеток можно провести анализ ферментов , активность которых связана с наличием именно этой хромосомы • Использование методов дифференцированной окраски хромосом позволяет связать гены с определенными локусами хромосом, так как в гибридных клетках довольно частыми являются хромосомные разрывы, перестройки, присутствие не целых хромосом, а отдельных фрагментов
 cell , Проточнаяцитометрия") 2. Сортировка клеток с активированной флуоресценцией, флуоресцентный сортинг (fluorescence-activated sorting FACS) cell , Проточнаяцитометрия —метод проточной цитометрии, в котором мишени (клетки, индивидуальные хромосомы и т. п. ) мечены флуоресцентным красителем, возбуждающимся под воздействием лазерного луча.
2. Сортировка клеток с активированной флуоресценцией, флуоресцентный сортинг (fluorescence-activated sorting FACS) cell , Проточнаяцитометрия —метод проточной цитометрии, в котором мишени (клетки, индивидуальные хромосомы и т. п. ) мечены флуоресцентным красителем, возбуждающимся под воздействием лазерного луча.
 хромосом chromosomesorting ( ) — разделение хромосом по") Жидкостная цитометрия хромосом(Сортировка сортинг ( ) хромосом chromosomesorting ( ) — разделение хромосом по их размерам после окраски флуорохромами, основанное на методе проточной цитометрии. • Жидкостная цитометрия, обычно используемая для анализа и разделения клеток, в 1979 году группой исследователей (лаборатория в Калифорнии) была адаптирована для количественного анализа и сортировки. • Этот метод позволяет отобрать все, кроме четырех хромосом человека [9 -12] и получить информацию о вариабельности их размеров. • Жидкостной сортинг лежал в основе создания библиотеки клонов хромосом-специфичной ДНК, которая используется для построения физической карты генома.
Жидкостная цитометрия хромосом(Сортировка сортинг ( ) хромосом chromosomesorting ( ) — разделение хромосом по их размерам после окраски флуорохромами, основанное на методе проточной цитометрии. • Жидкостная цитометрия, обычно используемая для анализа и разделения клеток, в 1979 году группой исследователей (лаборатория в Калифорнии) была адаптирована для количественного анализа и сортировки. • Этот метод позволяет отобрать все, кроме четырех хромосом человека [9 -12] и получить информацию о вариабельности их размеров. • Жидкостной сортинг лежал в основе создания библиотеки клонов хромосом-специфичной ДНК, которая используется для построения физической карты генома.
 прибор-сортер критерием для сортировки , материала служат различия в эмиссии флуоресцентного сигнала.
прибор-сортер критерием для сортировки , материала служат различия в эмиссии флуоресцентного сигнала.
 Принцип метода Методика заключается в выявлении рассеяния света лазерного луча при прохождении через него клетки в струе жидкости, причём, степень световой дисперсии позволяет получить представление о размерах и структуре клетки. Кроме того, в ходе анализа учитывается уровень флуоресценции химических соединений, входящих в состав клетки (аутофлуоресценция) или внесённых в образец перед проведением проточной цитометрии.
Принцип метода Методика заключается в выявлении рассеяния света лазерного луча при прохождении через него клетки в струе жидкости, причём, степень световой дисперсии позволяет получить представление о размерах и структуре клетки. Кроме того, в ходе анализа учитывается уровень флуоресценции химических соединений, входящих в состав клетки (аутофлуоресценция) или внесённых в образец перед проведением проточной цитометрии.
 • В данное время для картирования генов хромосом человека применяются также другие методы: – Биохимические методы — сравнение аминокислотных последовательностей белков и нуклеотидной последовательности ДНК отдельных хромосом – Цитологические методы — сопоставление изменений морфологии хромосомного участка с характерным фенотипом, анализ «ломких» участков хромосом – Молекулярно-генетические методы и др. 48
• В данное время для картирования генов хромосом человека применяются также другие методы: – Биохимические методы — сравнение аминокислотных последовательностей белков и нуклеотидной последовательности ДНК отдельных хромосом – Цитологические методы — сопоставление изменений морфологии хромосомного участка с характерным фенотипом, анализ «ломких» участков хромосом – Молекулярно-генетические методы и др. 48
 • Благодаря картированию удалось установить локализацию многих генов в хромосомах человека • Гены располагаются в хромосомах довольно скученно, образуя группы, между которыми могут находиться большие незанятые участки (пустыни) • В различных хромосомах находится различное число генов • Наибольшее число генов - свыше 3 000 в наибольшей из аутосом - первой хромосоме • В Х-хромосоме их известно свыше 1400 49
• Благодаря картированию удалось установить локализацию многих генов в хромосомах человека • Гены располагаются в хромосомах довольно скученно, образуя группы, между которыми могут находиться большие незанятые участки (пустыни) • В различных хромосомах находится различное число генов • Наибольшее число генов - свыше 3 000 в наибольшей из аутосом - первой хромосоме • В Х-хромосоме их известно свыше 1400 49
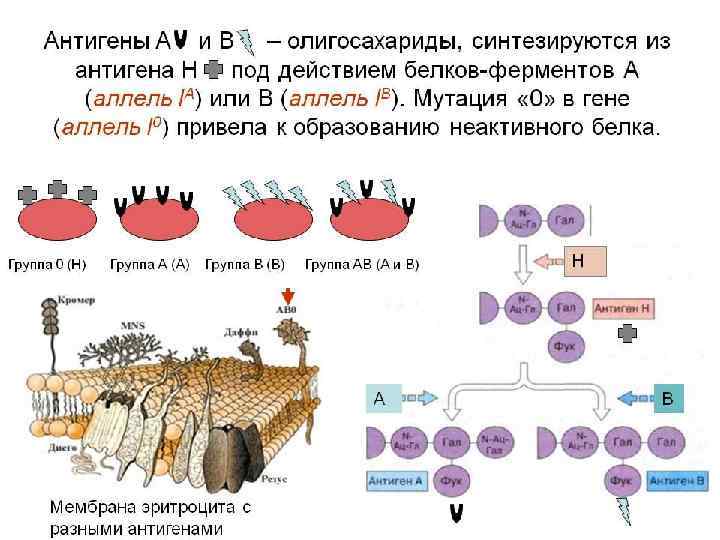
 • Аллели – группы крови по системе АВ 0, находятся в 9 -й хромосоме – группы крови по системе MN - во 2 -й – системы резус-фактора (Rh) - в 1 -й хромосоме – главного комплекса гистосовместимости – в 6 й хромосоме • Свыше 200 генов напрямую унаследованы нами от бактерий • Повторяющиеся последовательности ДНК, которые ранее считались лишними, могут оказаться «черным ящиком» эволюции и рассказать о предыдущих 800 млн. годах развития органического мира 50
• Аллели – группы крови по системе АВ 0, находятся в 9 -й хромосоме – группы крови по системе MN - во 2 -й – системы резус-фактора (Rh) - в 1 -й хромосоме – главного комплекса гистосовместимости – в 6 й хромосоме • Свыше 200 генов напрямую унаследованы нами от бактерий • Повторяющиеся последовательности ДНК, которые ранее считались лишними, могут оказаться «черным ящиком» эволюции и рассказать о предыдущих 800 млн. годах развития органического мира 50
 Хромосома 1 Хромосома 4
Хромосома 1 Хромосома 4
 Х-хромосома
Х-хромосома
 Примеры карт хромосом человека
Примеры карт хромосом человека
 Карта хромосомы 1, самой большой
Карта хромосомы 1, самой большой
 Карта хромосомы 9
Карта хромосомы 9
 Карта хромосомы 21 и митохондриального генома
Карта хромосомы 21 и митохондриального генома
 • Цитологические карты представляют собой фотографию или точный рисунок хромосомы, на котором обозначается последовательность Генетическая карта хромосомы расположения генов • Расположение генов на цитологических картах определяется с помощью транслокации • Транслокация– тип хромосомной перестройки • Во время транслокации происходит обмен негомологичными участками хромосом Цитологическая карта хромосомы • С помощью микроскопии изучают размеры, локализацию хромосомных перестроек 57
• Цитологические карты представляют собой фотографию или точный рисунок хромосомы, на котором обозначается последовательность Генетическая карта хромосомы расположения генов • Расположение генов на цитологических картах определяется с помощью транслокации • Транслокация– тип хромосомной перестройки • Во время транслокации происходит обмен негомологичными участками хромосом Цитологическая карта хромосомы • С помощью микроскопии изучают размеры, локализацию хромосомных перестроек 57
 • Дальнейшее картирование хромосом человека будет иметь практическое значение: станет возможным с помощью методов генной инженерии проводить профилактику и лечение многих наследственных болезней 58
• Дальнейшее картирование хромосом человека будет иметь практическое значение: станет возможным с помощью методов генной инженерии проводить профилактику и лечение многих наследственных болезней 58
 Генетические карты 7 -10 хромосом кукурузы Цифры по длине хромосом обозначают расстояние от конца хромосомы в морганидах; буквы - сокращенные названия признаков, определяемых соответствующими генами
Генетические карты 7 -10 хромосом кукурузы Цифры по длине хромосом обозначают расстояние от конца хромосомы в морганидах; буквы - сокращенные названия признаков, определяемых соответствующими генами
 Особенности построения генетических карт у прокариот Для построения генетических карт у прокариот используется явление конъюгации переноса генетического материала из одной – клетки в другую с помощью специальных кольцевых молекул ДНК (плазмид, в частности, с помощью F–плазмиды). Вероятность переноса определенного гена в клетку–реципиент зависит от его удаления от F–плазмидной ДНК, а точнее, от точки О, в которой начинается репликация F–плазмидной ДНК. Чем больше время конъюгации, тем выше вероятность переноса данного гена. Это дает возможность составить генетическую карту бактерий в минутах конъюгации. Например, у кишечной палочки ген thr (оперон из трех генов, контролирующих биосинтез треонина) находится в нулевой точке (то есть непосредственно рядом с F–плазмидной ДНК), ген lac переносится через 8 мин, ген rec. E – через 30 мин, ген arg. R – через 70 мин и т. д.
Особенности построения генетических карт у прокариот Для построения генетических карт у прокариот используется явление конъюгации переноса генетического материала из одной – клетки в другую с помощью специальных кольцевых молекул ДНК (плазмид, в частности, с помощью F–плазмиды). Вероятность переноса определенного гена в клетку–реципиент зависит от его удаления от F–плазмидной ДНК, а точнее, от точки О, в которой начинается репликация F–плазмидной ДНК. Чем больше время конъюгации, тем выше вероятность переноса данного гена. Это дает возможность составить генетическую карту бактерий в минутах конъюгации. Например, у кишечной палочки ген thr (оперон из трех генов, контролирующих биосинтез треонина) находится в нулевой точке (то есть непосредственно рядом с F–плазмидной ДНК), ген lac переносится через 8 мин, ген rec. E – через 30 мин, ген arg. R – через 70 мин и т. д.
 Цифры означают время (в") Генетическая карта хромосомы кишечной палочки ( Escherichia coli К 12) Цифры означают время (в мин), необходимое для переноса в клетку-реципиент генетических маркёров (характеризуют расстояние между генами), контролирующих биосинтез ряда аминокислот, а также устойчивость к стрептомицину и к фагу Т 6; Обозначения: ade - аденин; his - гистидин; try триптофан; gal - галактоза; lac - лактоза: pro - пролин; leu - лейцин; tre - треонин; met метионин; arg - аргинин; mt маннит; хуl - ксилоза; mal мальтоза; ser - серин; gly глицин; str и Т 6 устойчивость к стрептомицину или фагу T 6.
Генетическая карта хромосомы кишечной палочки ( Escherichia coli К 12) Цифры означают время (в мин), необходимое для переноса в клетку-реципиент генетических маркёров (характеризуют расстояние между генами), контролирующих биосинтез ряда аминокислот, а также устойчивость к стрептомицину и к фагу Т 6; Обозначения: ade - аденин; his - гистидин; try триптофан; gal - галактоза; lac - лактоза: pro - пролин; leu - лейцин; tre - треонин; met метионин; arg - аргинин; mt маннит; хуl - ксилоза; mal мальтоза; ser - серин; gly глицин; str и Т 6 устойчивость к стрептомицину или фагу T 6.
 Хромосомные карты
Хромосомные карты