

Лекц.6 Генетический аппарат клетки.pptx
- Количество слайдов: 174
 Генетический материал клетки. Уровни его организации
Генетический материал клетки. Уровни его организации
 Живые организмы существуют во времени и пространстве благодаря способности к самовоспроизведению. В результате самовоспроизведения про исходит смена поколений, сохраняющих черты организации предков. Эта особенность обеспечивается свойст вом живых систем передавать потомкам черты своей организации, характер обмена веществ и тип индивидуального развития в неизменном виде, которая называется наследственностью.
Живые организмы существуют во времени и пространстве благодаря способности к самовоспроизведению. В результате самовоспроизведения про исходит смена поколений, сохраняющих черты организации предков. Эта особенность обеспечивается свойст вом живых систем передавать потомкам черты своей организации, характер обмена веществ и тип индивидуального развития в неизменном виде, которая называется наследственностью.
 Но, жизнь живых существ была бы невоз можной в постоянно меняющихся условиях среды, если бы не их способность приобретать и сохранять некоторые изменения полезные в новых условиях. Свойство живых организмов приобретать и сохранять эти изменения называется изменчивостью.
Но, жизнь живых существ была бы невоз можной в постоянно меняющихся условиях среды, если бы не их способность приобретать и сохранять некоторые изменения полезные в новых условиях. Свойство живых организмов приобретать и сохранять эти изменения называется изменчивостью.
 Наследственность и изменчивость являются одними из главных свойств живой материи. Эти свойства обеспечиваются функционированием особого материаль ного субстрата – генетического аппарата.
Наследственность и изменчивость являются одними из главных свойств живой материи. Эти свойства обеспечиваются функционированием особого материаль ного субстрата – генетического аппарата.
 История изучения генетического аппарата .
История изучения генетического аппарата .
 чешским мо нахом,") Закономерности наследования признаков были установлены впервые Г. Менделем (1822 -1884) чешским мо нахом, человеком огромного интеллек туального развития; ученого и педагога, создателя классической генетики. 1865 г. , Г. Мендель. Экспериментально обосновал и сформулировал идею о наследственных задатках, их дискретности, специфичности и аллельном состоянии, наличии доминантных и рецессивных вариантов наследственных задатков (аллелей) или признаков, присутствии в половых клетках только одного аллеля (гаплоидность, чистота гамет) каждого наследственного задатка, а в соматических клетках - двух (диплоидность).
Закономерности наследования признаков были установлены впервые Г. Менделем (1822 -1884) чешским мо нахом, человеком огромного интеллек туального развития; ученого и педагога, создателя классической генетики. 1865 г. , Г. Мендель. Экспериментально обосновал и сформулировал идею о наследственных задатках, их дискретности, специфичности и аллельном состоянии, наличии доминантных и рецессивных вариантов наследственных задатков (аллелей) или признаков, присутствии в половых клетках только одного аллеля (гаплоидность, чистота гамет) каждого наследственного задатка, а в соматических клетках - двух (диплоидность).
 Наследственность человека как самостоятельный предмет исследования впервые выделил в 1865 г. английский ученый Ф. Гальтон (1822 -1911), которого считают одним из основателей генетики.
Наследственность человека как самостоятельный предмет исследования впервые выделил в 1865 г. английский ученый Ф. Гальтон (1822 -1911), которого считают одним из основателей генетики.
 Август Вейсман. , немецкий зоолог и теоретик эволюционного учения. Впервые высказал и доказал правильность мнения о том, что переход родительских свойств зависит от некоего материального вещества, оказывающего влияние на развитие потомства и это вещество заключено в хромосомах и хроматине Вейсман — автор умозрительных теорий наследственности и индивидуального развития, неверных в деталях, но в принципе предвосхитивших современные представления о дискретности носителей наследственной информации и их связи с хромосомами, а также концепции о роли наследст-венных задатков в индивидуальном развитии.
Август Вейсман. , немецкий зоолог и теоретик эволюционного учения. Впервые высказал и доказал правильность мнения о том, что переход родительских свойств зависит от некоего материального вещества, оказывающего влияние на развитие потомства и это вещество заключено в хромосомах и хроматине Вейсман — автор умозрительных теорий наследственности и индивидуального развития, неверных в деталях, но в принципе предвосхитивших современные представления о дискретности носителей наследственной информации и их связи с хромосомами, а также концепции о роли наследст-венных задатков в индивидуальном развитии.
 — немецкий анатом и гистолог. 1888 г.") Генрих Вильгельм Готфрид Вальдейер (1836 — 1921) — немецкий анатом и гистолог. 1888 г. , В. Вальдейер. Предложил термин «хромосома» для обозначения особых ядерных структур.
Генрих Вильгельм Готфрид Вальдейер (1836 — 1921) — немецкий анатом и гистолог. 1888 г. , В. Вальдейер. Предложил термин «хромосома» для обозначения особых ядерных структур.

 датский биолог, один из основоположников сов ременной генетики Назвал:") Иогансен Вильгельм Людвиг (1857 1927) датский биолог, один из основоположников сов ременной генетики Назвал: наследственные задатки Г. Менделя генами (от греч. genos род, происхождение), совокупность генов организ ма генотипом, проявление генов (совокупность признаков и свойств) организма фенотипом
Иогансен Вильгельм Людвиг (1857 1927) датский биолог, один из основоположников сов ременной генетики Назвал: наследственные задатки Г. Менделя генами (от греч. genos род, происхождение), совокупность генов организ ма генотипом, проявление генов (совокупность признаков и свойств) организма фенотипом
 — американский биолог, один из осново положников генетики, Лауреат") Томас Хант Морган 1866 1945) — американский биолог, один из осново положников генетики, Лауреат Но белевской премии «За открытия, связанные с ролью хромосом в нас ледственности» Вместе со своими учениками К. Бриджесом, А. Стертевантом и Г. Мёл лером сформулировал Хромосомную теорию наследственности. Обнаружив сцепленное наследование признаков, он предположил, что гены располагаются в хромосоме в тесной близости друг к другу, и составил «карты» , отражающие эту картину.
Томас Хант Морган 1866 1945) — американский биолог, один из осново положников генетики, Лауреат Но белевской премии «За открытия, связанные с ролью хромосом в нас ледственности» Вместе со своими учениками К. Бриджесом, А. Стертевантом и Г. Мёл лером сформулировал Хромосомную теорию наследственности. Обнаружив сцепленное наследование признаков, он предположил, что гены располагаются в хромосоме в тесной близости друг к другу, и составил «карты» , отражающие эту картину.

 У. Сеттон американский цитолог Т. Г. Бо вери немецкий биолог. Пришли к заключению и представили доказательства, что внутриклеточными носителями генетической про граммы являются хромосомы.
У. Сеттон американский цитолог Т. Г. Бо вери немецкий биолог. Пришли к заключению и представили доказательства, что внутриклеточными носителями генетической про граммы являются хромосомы.
 (1861, 1926) — английский биолог, один из осно вателей генетики, автор") Уильям Бэтсон (Бейтсон) (1861, 1926) — английский биолог, один из осно вателей генетики, автор термина «генетика» (1907). Ввел терми ны «генотип» и «фенотип» .
Уильям Бэтсон (Бейтсон) (1861, 1926) — английский биолог, один из осно вателей генетики, автор термина «генетика» (1907). Ввел терми ны «генотип» и «фенотип» .
 — русский биолог, основатель русс") Никола й Константи нович Кольцо в (1872 1940) — русский биолог, основатель русс кой советской школы экспери ментальной биологии. Первым разработал гипотезу молекуляр ного строения и матричной репродукции хромосом ( «наследственные молекулы» ), предвосхитившую главнейшие принци пиальные положения современной моле кулярной биологии и генетики. Первым из русских учёных привнёс физико хими ческий метод, вошедший в набор основ ных методов биологических исследова ний.
Никола й Константи нович Кольцо в (1872 1940) — русский биолог, основатель русс кой советской школы экспери ментальной биологии. Первым разработал гипотезу молекуляр ного строения и матричной репродукции хромосом ( «наследственные молекулы» ), предвосхитившую главнейшие принци пиальные положения современной моле кулярной биологии и генетики. Первым из русских учёных привнёс физико хими ческий метод, вошедший в набор основ ных методов биологических исследова ний.
 Освальд Теодор Эвери американский молекулярный биолог, иммунолог, медик. 1944 г. Установил химическую природу вещества наследствен ности ДНК. Один из соавторов эксперимента Эвери, К. Маклеода и М. Макка рти, который в 1944 году пока зал, что ДНК представляет со бой носитель генетической информации.
Освальд Теодор Эвери американский молекулярный биолог, иммунолог, медик. 1944 г. Установил химическую природу вещества наследствен ности ДНК. Один из соавторов эксперимента Эвери, К. Маклеода и М. Макка рти, который в 1944 году пока зал, что ДНК представляет со бой носитель генетической информации.
 Международные проект «Геном человека» был начат в 1988 г. Это один из самых трудоемких и дорогостоящих проектов в истории науки. Если в 1990 г. на него было потрачено около 60 млн долларов в целом, то в 1998 г. одно только правительство США израсходовало 253 млн долларов, а частные компании – и того больше. В проекте задействованы несколько тысяч ученых из бо лее чем 20 стран. С 1989 г. в нем участвует и Россия, где по проекту работает около 100 групп. Все хромосомы человека по делены между странами участницами, и России для исследо вания достались 3 , 13 и 19 я хромосомы. В 2000 году был выпущен рабочий черновик структуры ге нома, полный геном — в 2003 году, однако и сегодня допол нительный анализ некоторых участков ещё не закончен.
Международные проект «Геном человека» был начат в 1988 г. Это один из самых трудоемких и дорогостоящих проектов в истории науки. Если в 1990 г. на него было потрачено около 60 млн долларов в целом, то в 1998 г. одно только правительство США израсходовало 253 млн долларов, а частные компании – и того больше. В проекте задействованы несколько тысяч ученых из бо лее чем 20 стран. С 1989 г. в нем участвует и Россия, где по проекту работает около 100 групп. Все хромосомы человека по делены между странами участницами, и России для исследо вания достались 3 , 13 и 19 я хромосомы. В 2000 году был выпущен рабочий черновик структуры ге нома, полный геном — в 2003 году, однако и сегодня допол нительный анализ некоторых участков ещё не закончен.
 1955 -1966 гг. , Х. Г. Корана, М. У. Ниренберг. Расшифровали кодовую систему записи генетической информации в молекулах нуклеиновых кислот генетический код. 1960 г. (Денвер); 1963 г. (Лондон); 1966 г. (Чикаго). 1971 г. (Париж). Классификации и номенклатура кариотипа человека; 1961 г. , Ф. Жакоб, Ж. Л. Моно. Сформулировали гипотезу о оперонной регуляции активности генов прокариот. 1970 -е годы, Р. Д. Робертс и Ф. А. Шарп. Предоставили доказательства ин-трон-экзонной организации генов эукариот.
1955 -1966 гг. , Х. Г. Корана, М. У. Ниренберг. Расшифровали кодовую систему записи генетической информации в молекулах нуклеиновых кислот генетический код. 1960 г. (Денвер); 1963 г. (Лондон); 1966 г. (Чикаго). 1971 г. (Париж). Классификации и номенклатура кариотипа человека; 1961 г. , Ф. Жакоб, Ж. Л. Моно. Сформулировали гипотезу о оперонной регуляции активности генов прокариот. 1970 -е годы, Р. Д. Робертс и Ф. А. Шарп. Предоставили доказательства ин-трон-экзонной организации генов эукариот.
 Генетический аппарат - это совокупность компонентов клет ки человека, обеспечивающих хра нение, передачу и реализацию гене тической информации. Генетическим аппаратом клетки является молекула ДНК
Генетический аппарат - это совокупность компонентов клет ки человека, обеспечивающих хра нение, передачу и реализацию гене тической информации. Генетическим аппаратом клетки является молекула ДНК
 Основная функция генетического аппарата - контроль наследственности и изменчивости
Основная функция генетического аппарата - контроль наследственности и изменчивости
 Хранение генетической информации осуществляют следующие структуры эукариотической клетки: ядерная ДНК – 46 линейных молекул ДНК, упакованных в форме хроматина или хромосом в ядре; ü 50% материнского и 50% отцовского происхождения; ü содержит около 30 000 пар кодирующих белки генов, лока лизованных в 23 парах молекул ДНК (что составляет 25% ядерного генома); ü 22 пары аутосом (одинаковых у мужчин и женщин) и пара половых хромосом, отличающихся у мужчин и женщин (XY и XX); ü соматические клетки имеют идентичный генетический материал; митохондриальная ДНК – от одной до нескольких сотен кольцевых молекул ДНК; ü 100% материнского происхождения; ü содержит 13 структурных генов;
Хранение генетической информации осуществляют следующие структуры эукариотической клетки: ядерная ДНК – 46 линейных молекул ДНК, упакованных в форме хроматина или хромосом в ядре; ü 50% материнского и 50% отцовского происхождения; ü содержит около 30 000 пар кодирующих белки генов, лока лизованных в 23 парах молекул ДНК (что составляет 25% ядерного генома); ü 22 пары аутосом (одинаковых у мужчин и женщин) и пара половых хромосом, отличающихся у мужчин и женщин (XY и XX); ü соматические клетки имеют идентичный генетический материал; митохондриальная ДНК – от одной до нескольких сотен кольцевых молекул ДНК; ü 100% материнского происхождения; ü содержит 13 структурных генов;
 Передача генетической информации от клетки к клетке осуществляется во время митоза. Передачу генетической информации осу ществляют следующие компоненты: Ø реплиосома – комплекс белков, обеспечива ющих репликацию; Ø белковые факторы, необходимые для репа рации; Ø центриоли, участвующие в образовании ве ретена деления и ответственные за точное и равное распределение хромосом в митозе;
Передача генетической информации от клетки к клетке осуществляется во время митоза. Передачу генетической информации осу ществляют следующие компоненты: Ø реплиосома – комплекс белков, обеспечива ющих репликацию; Ø белковые факторы, необходимые для репа рации; Ø центриоли, участвующие в образовании ве ретена деления и ответственные за точное и равное распределение хромосом в митозе;
 Реализация генетической информации, закодирован ной в структурных генах, протекает в несколько этапов с учас тием: ü аппарата транскрипции – белковых факторов, распознающих ген мишень и обеспечивающих синтез m. РНК; ü аппарата трансляции – рибосом, являющихся местом трансляции и синтеза полипептида; ü молекул m. РНК – переводчиков генетического кода; ü белковых факторов – регуляторов трансляции и созревания белков. Реализация генетической информации: - на молекулярном уровне осуществляется в процессе синтеза определенного белка; на клеточном уровне проявляется в выполнении функций данного белка в клетке, образованием определенной клеточной структуры/метабо лической цепи/сигнального пути; на уровне организма –развитием определенного фенотипа.
Реализация генетической информации, закодирован ной в структурных генах, протекает в несколько этапов с учас тием: ü аппарата транскрипции – белковых факторов, распознающих ген мишень и обеспечивающих синтез m. РНК; ü аппарата трансляции – рибосом, являющихся местом трансляции и синтеза полипептида; ü молекул m. РНК – переводчиков генетического кода; ü белковых факторов – регуляторов трансляции и созревания белков. Реализация генетической информации: - на молекулярном уровне осуществляется в процессе синтеза определенного белка; на клеточном уровне проявляется в выполнении функций данного белка в клетке, образованием определенной клеточной структуры/метабо лической цепи/сигнального пути; на уровне организма –развитием определенного фенотипа.
 Американские генетики, авторы гипотезы «один ген — один фермент» . Э дуард Те йтем Бидл, Джордж Уэлс В 1941 1944 годах американские ученые Дж. У. Бидл и Э. Л. Тейтам сформулировали свой знаменитый принцип: «один ген — один фер мент — один признак (симптом)» .
Американские генетики, авторы гипотезы «один ген — один фермент» . Э дуард Те йтем Бидл, Джордж Уэлс В 1941 1944 годах американские ученые Дж. У. Бидл и Э. Л. Тейтам сформулировали свой знаменитый принцип: «один ген — один фер мент — один признак (симптом)» .
 Различают следующие уровни структурно-функциональной организации наследственного материала: генный, хромосомный, геномный. Элементарной структурой ГЕННОГО уровня организации служит ГЕН.
Различают следующие уровни структурно-функциональной организации наследственного материала: генный, хромосомный, геномный. Элементарной структурой ГЕННОГО уровня организации служит ГЕН.
 Гены клеток эукариот распределены по хромосомам, образуя ХРОМОСОМНЫЙ уровень организации наследственного материала
Гены клеток эукариот распределены по хромосомам, образуя ХРОМОСОМНЫЙ уровень организации наследственного материала
 Геномный уровень организациинаследственного аппарата объясняет взаимодействие генов как вод-ной , так и в разных хромосомах.
Геномный уровень организациинаследственного аппарата объясняет взаимодействие генов как вод-ной , так и в разных хромосомах.
 Генетический аппарат клетки. Генный уровень организации генетического аппарата
Генетический аппарат клетки. Генный уровень организации генетического аппарата
 Ген – это: ü элементарная единица функционирования генного уровня организации генетического аппарата ü единица генетической инфор мации
Ген – это: ü элементарная единица функционирования генного уровня организации генетического аппарата ü единица генетической инфор мации
 Физическое определени Ген – это участок ДНК, на кото- ром закодирована информация о структуре РНК или полипептидной цепи. Ген занимает определенное положение в хромосоме или геноме и контролирует выполнение определенной функции в организме.
Физическое определени Ген – это участок ДНК, на кото- ром закодирована информация о структуре РНК или полипептидной цепи. Ген занимает определенное положение в хромосоме или геноме и контролирует выполнение определенной функции в организме.
 Функциональное определение Ген - это информационная единица, которая обеспечивает синтез РНК; кодируя последовательность аминокислот в белках, несет информацию о признаках данного организма.
Функциональное определение Ген - это информационная единица, которая обеспечивает синтез РНК; кодируя последовательность аминокислот в белках, несет информацию о признаках данного организма.
 Признак – это свойство по которому один организм отличается от другого Свойство белка, определяемое последовательностью аминокислот, является элементарным или простым признаком.
Признак – это свойство по которому один организм отличается от другого Свойство белка, определяемое последовательностью аминокислот, является элементарным или простым признаком.
 В 1941 году Э. Тэйтум и Дж. Бидл обнаружили, что у гриба нейрооспоры мутация одного гена приводит к утрате у гриба способности синтезировать какую-либо необходимую для роста аминокислоту, витамин или др. ростовой фактор. Дальнейшие эксперименты показали, что под действием различных мутаций каждый раз происходит выключение только одной какой-либо цепи биохимических реакций
В 1941 году Э. Тэйтум и Дж. Бидл обнаружили, что у гриба нейрооспоры мутация одного гена приводит к утрате у гриба способности синтезировать какую-либо необходимую для роста аминокислоту, витамин или др. ростовой фактор. Дальнейшие эксперименты показали, что под действием различных мутаций каждый раз происходит выключение только одной какой-либо цепи биохимических реакций
 Дж. Бидл и Э. Тэйтум сформулировали гипотезу «один ген - один фермент» : На основании исследования гемоглобина В. Ингрэм (1957) предложил уточнить формулировку гипотезы «один ген – один фермент» в виде «один ген – одна полипептидная цепь» .
Дж. Бидл и Э. Тэйтум сформулировали гипотезу «один ген - один фермент» : На основании исследования гемоглобина В. Ингрэм (1957) предложил уточнить формулировку гипотезы «один ген – один фермент» в виде «один ген – одна полипептидная цепь» .
 Мозаичное строение генов эукариот. Тонкая структура гена Р. Робетсон и Ф. Шарп установили факт существования расщепленных генов, то есть, генов, состоящих из кодирующих и некодирующих последовательности нуклеотидов. Они были названы, экзонами и интронами. Экзоны – это кодирующие последовательности нук леотидов, а интроны – некодирующие последовательности. Интрон экзонная структура гена характерна для генов эукариот.
Мозаичное строение генов эукариот. Тонкая структура гена Р. Робетсон и Ф. Шарп установили факт существования расщепленных генов, то есть, генов, состоящих из кодирующих и некодирующих последовательности нуклеотидов. Они были названы, экзонами и интронами. Экзоны – это кодирующие последовательности нук леотидов, а интроны – некодирующие последовательности. Интрон экзонная структура гена характерна для генов эукариот.
 Альтернативный сплайсинг - процесс, позволяющий одному гену производить несколько м. РНК и, соответственно, белков.
Альтернативный сплайсинг - процесс, позволяющий одному гену производить несколько м. РНК и, соответственно, белков.
 В изучении природы гена особенно большое значение имели работы С. Бензера и ряда других исследователей, проведенные на бактериофагах и других объектах. В результате этих работ было доказано, что ген дискретен. Ген может быть разделен кроссинговером на множество частей. Дискретная организация генов была установлена и у эукариот.
В изучении природы гена особенно большое значение имели работы С. Бензера и ряда других исследователей, проведенные на бактериофагах и других объектах. В результате этих работ было доказано, что ген дискретен. Ген может быть разделен кроссинговером на множество частей. Дискретная организация генов была установлена и у эукариот.
 Дискретность гена заключается в наличии субъединиц: МУТОН - элементарная единица мутации. РЕКОН - единица рекомбинации. ЦИСТРОН - единица функции. САЙТ - это структурная единица гена. Равен 1 паре нуклеотидов. Является минимальным размером мутона, рекона и цистрона. КОДОН - функциональная единица гена.
Дискретность гена заключается в наличии субъединиц: МУТОН - элементарная единица мутации. РЕКОН - единица рекомбинации. ЦИСТРОН - единица функции. САЙТ - это структурная единица гена. Равен 1 паре нуклеотидов. Является минимальным размером мутона, рекона и цистрона. КОДОН - функциональная единица гена.
 С. Бензер ввел три новых понятия: рекон, мутон, цистрон. Считалось, кроссинговер происходит между генами и, таким образом, ген - это элементарная единица генетической рекомбинации. Однако доказано, что рекомбинации происходят и внутри гена. Наименьшая единица рекомбинации названа реконом. Ранее считали ген единицей мутации. Однако было обнаружено, что даже изменения отдельных участков внутри сложного гена приводят к изменению его функции. Мельчайшая единица, способная к изменению, была названа мутоном.
С. Бензер ввел три новых понятия: рекон, мутон, цистрон. Считалось, кроссинговер происходит между генами и, таким образом, ген - это элементарная единица генетической рекомбинации. Однако доказано, что рекомбинации происходят и внутри гена. Наименьшая единица рекомбинации названа реконом. Ранее считали ген единицей мутации. Однако было обнаружено, что даже изменения отдельных участков внутри сложного гена приводят к изменению его функции. Мельчайшая единица, способная к изменению, была названа мутоном.
 Наименьшая часть гена, рекомбинирующаяся в процессе кроссинговера, называется рекон и она является элементарной единицей рекомбинации. Рекомбинация генов - это процесс образования новых сочетаний генов в процессе кроссинговера Внутригенный кроссинговер с неравноценным обменом наследственного материала : I – неравноценный кроссинговер между гомологичными хромосомами; II – выпадение 3 и 4 фрагментов; III – удвоение 3 и 4 фрагментов
Наименьшая часть гена, рекомбинирующаяся в процессе кроссинговера, называется рекон и она является элементарной единицей рекомбинации. Рекомбинация генов - это процесс образования новых сочетаний генов в процессе кроссинговера Внутригенный кроссинговер с неравноценным обменом наследственного материала : I – неравноценный кроссинговер между гомологичными хромосомами; II – выпадение 3 и 4 фрагментов; III – удвоение 3 и 4 фрагментов
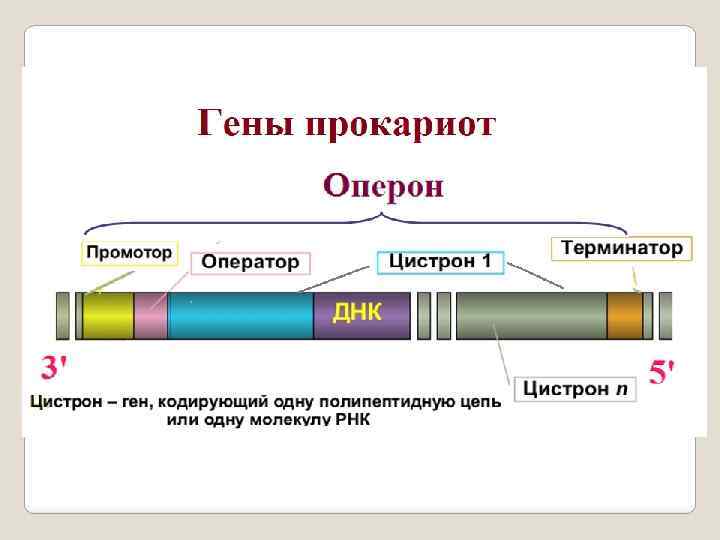
 Ген как функциональную единицу предложено называть цистроном. Цистро н — у эукариот синоним термина "ген", обозначающего участок ДНК, ответственный за синтез определённого белка. Для эукариот термин «цистрон» не применяется. У прокариот гены, выполняющие сходные метаболические функции, часто объединены в опероны и называются цистронами (полицистронный механизм регуляции активности генов). Размеры рекона и мутона могут равняться одной или нескольким парам нуклеотидов, цистрона — сотням и тысячам нуклеотидов, но наименьшая величина рекона и мутона близка к величине одного или нескольких нуклеотидов. Цистрон же содержит тысячу и более нуклеотидов.
Ген как функциональную единицу предложено называть цистроном. Цистро н — у эукариот синоним термина "ген", обозначающего участок ДНК, ответственный за синтез определённого белка. Для эукариот термин «цистрон» не применяется. У прокариот гены, выполняющие сходные метаболические функции, часто объединены в опероны и называются цистронами (полицистронный механизм регуляции активности генов). Размеры рекона и мутона могут равняться одной или нескольким парам нуклеотидов, цистрона — сотням и тысячам нуклеотидов, но наименьшая величина рекона и мутона близка к величине одного или нескольких нуклеотидов. Цистрон же содержит тысячу и более нуклеотидов.
, то его ген состоит из нескольких") Цистрон Если белок состоит из нескольких субъединиц (гемоглобин), то его ген состоит из нескольких цистронов
Цистрон Если белок состоит из нескольких субъединиц (гемоглобин), то его ген состоит из нескольких цистронов

 В настоящее время ген определяют как единицу наследственной информации, занимающую определенное положение в геноме и контролирующую выполнение определенной функции в организме. С использованием современных биохимических и молекулярно-биологических методов к настоящему времени уже расшифрована первичная структура многих генов, выяснены основные черты их строения и особенности функционирования.
В настоящее время ген определяют как единицу наследственной информации, занимающую определенное положение в геноме и контролирующую выполнение определенной функции в организме. С использованием современных биохимических и молекулярно-биологических методов к настоящему времени уже расшифрована первичная структура многих генов, выяснены основные черты их строения и особенности функционирования.
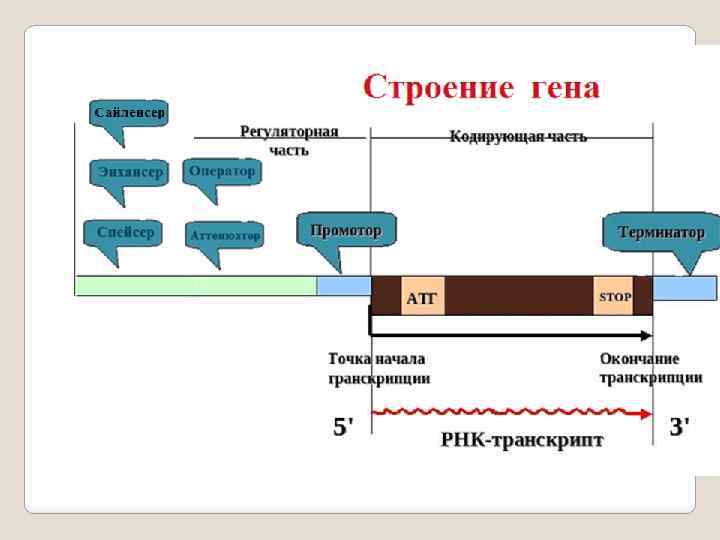
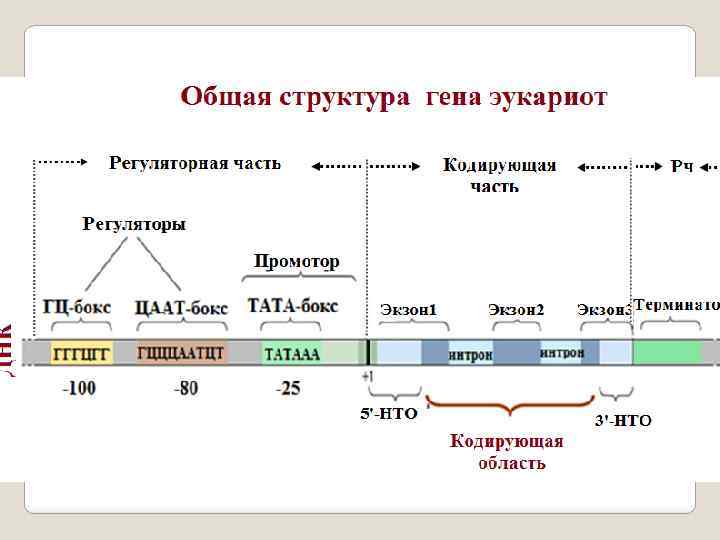
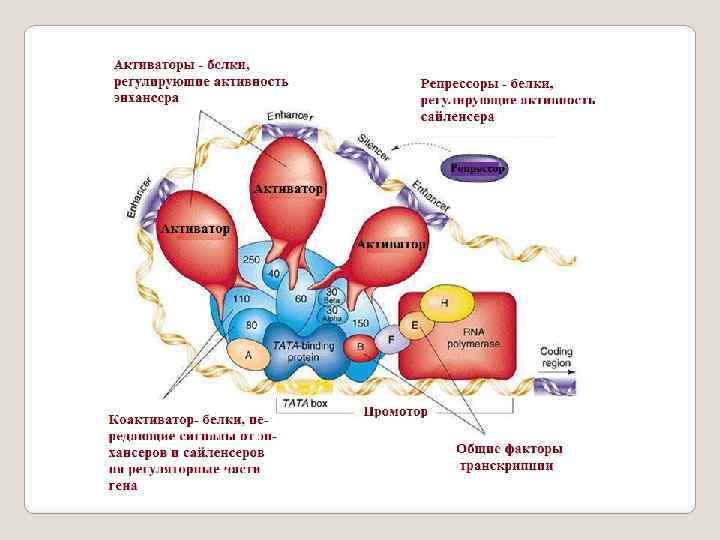
 Регуляторная часть гена Промотор – это последовательность нуклеотидов ДНК, узнаваемая РНКполимеразой; стартовая площадка для начала транскрипции. Оператор- регуляторный участок гена (оперона), с которым специфически связывается репрессор, предотвращая тем самым начало транскрипции. Энхансер — регуляторный участок ДНК, усиливающий скорость транскрипции с ближайшего к нему промотора. Сайленсер - регуляторный участок ДНК, снижающий скорость транскри -пции с ближайшего к нему промотора. Аттенюатор - (attenuator, лат. attenuatio — уменьшение, сокращение) - нуклеотидная последовательность ДНК, расположенная перед опероном, которая участвует в регуляции его экспрессии путем осуществления преждевременной терминации транскрипции; Спейсер - в ДНК или РНК — некодирующая последовательность нуклео -тидов между генами; Терминатор - последовательность нуклеотидов ДНК, узнаваемая РНКполимеразой как сигнал к прекращению синтеза молекулы РНК и диссоциации транскрипционного комплекса.
Регуляторная часть гена Промотор – это последовательность нуклеотидов ДНК, узнаваемая РНКполимеразой; стартовая площадка для начала транскрипции. Оператор- регуляторный участок гена (оперона), с которым специфически связывается репрессор, предотвращая тем самым начало транскрипции. Энхансер — регуляторный участок ДНК, усиливающий скорость транскрипции с ближайшего к нему промотора. Сайленсер - регуляторный участок ДНК, снижающий скорость транскри -пции с ближайшего к нему промотора. Аттенюатор - (attenuator, лат. attenuatio — уменьшение, сокращение) - нуклеотидная последовательность ДНК, расположенная перед опероном, которая участвует в регуляции его экспрессии путем осуществления преждевременной терминации транскрипции; Спейсер - в ДНК или РНК — некодирующая последовательность нуклео -тидов между генами; Терминатор - последовательность нуклеотидов ДНК, узнаваемая РНКполимеразой как сигнал к прекращению синтеза молекулы РНК и диссоциации транскрипционного комплекса.
 Для прокариот характерно: ü гены находятся в цитоплазме, ü гены независимые : структурный ген бактерии контролирует одну ферментативную реакцию, ü оперонная система организации генов. Гены у бактеориофагов и вирусов в основном схожы с бактериями, но более усложнена и сопряжена с геномом хозяев: у фагов и вирусов обнаружено перекрывание генов, у вирусов имется экзон интронная структуры генов. гены отделены друг от друга спейсерами, однако длина спейсеров.
Для прокариот характерно: ü гены находятся в цитоплазме, ü гены независимые : структурный ген бактерии контролирует одну ферментативную реакцию, ü оперонная система организации генов. Гены у бактеориофагов и вирусов в основном схожы с бактериями, но более усложнена и сопряжена с геномом хозяев: у фагов и вирусов обнаружено перекрывание генов, у вирусов имется экзон интронная структуры генов. гены отделены друг от друга спейсерами, однако длина спейсеров.
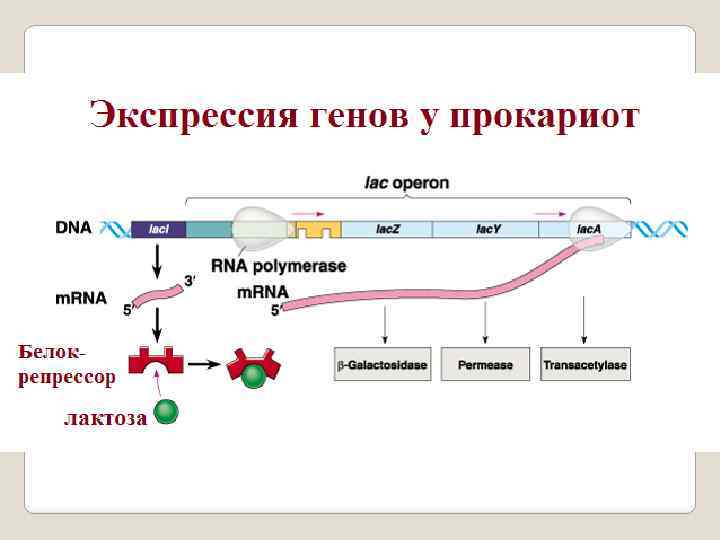
 Эукариотические гены: ü эукариотическая ДНК расположена в ядре, ü имеют прерывистое мозаичное строение: кодирующие последовательности (экзоны) перемежаются с некодирую щими (интронами), ü собраны в кластеры изофункциональных генов, ü отделены друг от друга спейсерами ü не все гены эукариот содержат интроны: гены гистоно вых белков, гены интерферонов, гены митохондрий млекопитающих и человека, ü ген может кодировать различные РНК продукты путем изменения инициирующих и терминирующих кодонов, а также с помощью альтернативного сплайсинга,
Эукариотические гены: ü эукариотическая ДНК расположена в ядре, ü имеют прерывистое мозаичное строение: кодирующие последовательности (экзоны) перемежаются с некодирую щими (интронами), ü собраны в кластеры изофункциональных генов, ü отделены друг от друга спейсерами ü не все гены эукариот содержат интроны: гены гистоно вых белков, гены интерферонов, гены митохондрий млекопитающих и человека, ü ген может кодировать различные РНК продукты путем изменения инициирующих и терминирующих кодонов, а также с помощью альтернативного сплайсинга,
 Интроны всегда имеют на 5'-конце пару последовательностей СТ, а на 3'-конце — AG.
Интроны всегда имеют на 5'-конце пару последовательностей СТ, а на 3'-конце — AG.
 Кластер генов – это группа различных генов с родственными функция ми, локализованных в определённых участках хромосом. В состав кластера входят активно функционирующие гены и псевдогены. Часто кластеры являются семейством генов, произошедшим от некоего гена-предка. Иногда гены, образующие кластеры называют мультигенными семействами.
Кластер генов – это группа различных генов с родственными функция ми, локализованных в определённых участках хромосом. В состав кластера входят активно функционирующие гены и псевдогены. Часто кластеры являются семейством генов, произошедшим от некоего гена-предка. Иногда гены, образующие кластеры называют мультигенными семействами.
;") Кластер генов гемоглобина гемоглобин А - основной гемоглобин взрослого организма, формула - (2α, 2β); гемоглобин А 2 - гемоглобин взрослого человека, но содержится в организме в меньшей концентрации ( 2% общего гемоглобина), формула - (2α, 2δ); эмбриональный гемоглобин, ; формула – (2ζ, 2ε); гемоглобин F – фетальный гемоглобин. Замещает эмбриональный гемоглобин на 6 месяце развития плода; формула - (2α, 2γ);
Кластер генов гемоглобина гемоглобин А - основной гемоглобин взрослого организма, формула - (2α, 2β); гемоглобин А 2 - гемоглобин взрослого человека, но содержится в организме в меньшей концентрации ( 2% общего гемоглобина), формула - (2α, 2δ); эмбриональный гемоглобин, ; формула – (2ζ, 2ε); гемоглобин F – фетальный гемоглобин. Замещает эмбриональный гемоглобин на 6 месяце развития плода; формула - (2α, 2γ);
 Кластеров генов р. РНК эукариот
Кластеров генов р. РНК эукариот
 Кластеров гистоновых генов Гистоны — обширный класс ядерных белков, выполняю щих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в регуляции таких ядерных процессов, как репликация, репарация и транскрипция. Существует пять различных типов гистонов H 1, H 2 A, H 2 B, H 3, H 4.
Кластеров гистоновых генов Гистоны — обширный класс ядерных белков, выполняю щих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в регуляции таких ядерных процессов, как репликация, репарация и транскрипция. Существует пять различных типов гистонов H 1, H 2 A, H 2 B, H 3, H 4.
 Мозг (более 3000) Глаз (более") Распределение генов человека по их функциям Кожа (около 700) Мозг (более 3000) Глаз (более 500) Лёгкие (около 2000) Сердце (более 1200) Кишечник (около 1500) Печень (более 2000) Матка (около 2000) Семенники (около 400) Белая клетка крови Эритроцит (8) (более 2000)
Распределение генов человека по их функциям Кожа (около 700) Мозг (более 3000) Глаз (более 500) Лёгкие (около 2000) Сердце (более 1200) Кишечник (около 1500) Печень (более 2000) Матка (около 2000) Семенники (около 400) Белая клетка крови Эритроцит (8) (более 2000)
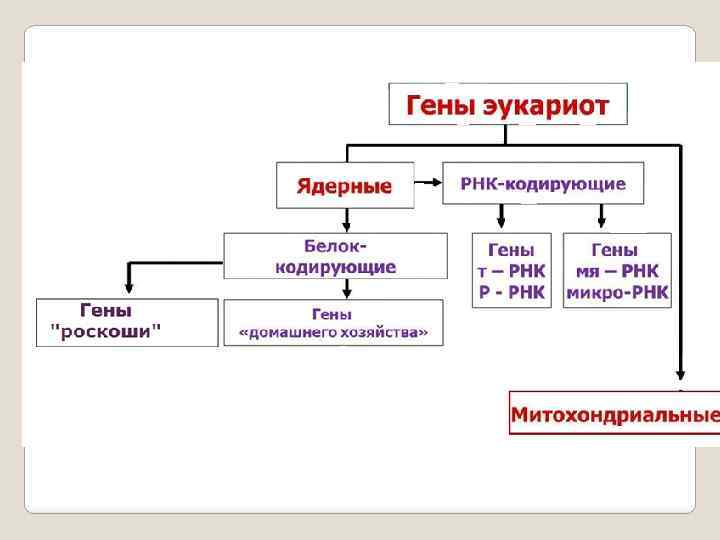
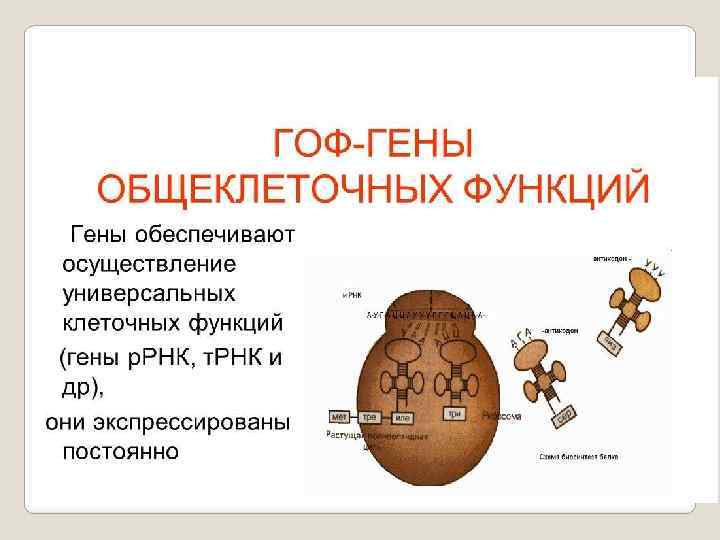
,") Классификация генов Существует несколько классификаций генов. 1. Конститутивные гены или гены общеклеточных функций (ГОФ-гены), гены домашнего хозяйства 2. Гены «роскоши» или гены специализированных функций (ГСФ-гены)
Классификация генов Существует несколько классификаций генов. 1. Конститутивные гены или гены общеклеточных функций (ГОФ-гены), гены домашнего хозяйства 2. Гены «роскоши» или гены специализированных функций (ГСФ-гены)
 постоянно") Конститутивные гены или гены общеклеточных функций (их ещё называют гены «домашнего хозяйства) постоянно находятся в активном состоянии. Их активность не зависит от состояния внешней среды (организма), т. е. практически не регулируется. Эти гены кодируют белки ферменты, которые при нимают участие в жизненно важных для клетки мета болических процессах. Например, таких как гликолиз, цепь передачи элек тронов, синтез ДНК, аминокислот и т. д. В сущности, эти гены полностью обеспечивают жизнедеятельность клетки.
Конститутивные гены или гены общеклеточных функций (их ещё называют гены «домашнего хозяйства) постоянно находятся в активном состоянии. Их активность не зависит от состояния внешней среды (организма), т. е. практически не регулируется. Эти гены кодируют белки ферменты, которые при нимают участие в жизненно важных для клетки мета болических процессах. Например, таких как гликолиз, цепь передачи элек тронов, синтез ДНК, аминокислот и т. д. В сущности, эти гены полностью обеспечивают жизнедеятельность клетки.
 Гены «роскоши» ГСФ гены контролируют строго спе циализированные, специфические функции клетки. На организменном уровне имеются собственные системы жизнеобеспечения, развития, размножения, дыхания и т. д. Поэтому любая клетка организма должна поддерживать не толь ко свои жизненные потенции (которые обеспечивают ГОФ ге ны), но и принимать участие в жизнедеятельности всего ор ганизма. Последним и занимаются специализированные гены – гены «роскоши» . Эти гены контролируют белки, которые обеспечи вают функционирование физиологических систем организма – его защитных свойств, процессов дыхания, выделе ния, кровоснабжения, пищеварения и т. д. ). К таким генам относятся гены, контролирующие синтез ге моглобина, иммуноглобулина и др. В отличии от генов «домаш него хозяйства» «гены роскоши» находятся под жёстким конт ролем организма и имеют сложный аппарат регуляции.
Гены «роскоши» ГСФ гены контролируют строго спе циализированные, специфические функции клетки. На организменном уровне имеются собственные системы жизнеобеспечения, развития, размножения, дыхания и т. д. Поэтому любая клетка организма должна поддерживать не толь ко свои жизненные потенции (которые обеспечивают ГОФ ге ны), но и принимать участие в жизнедеятельности всего ор ганизма. Последним и занимаются специализированные гены – гены «роскоши» . Эти гены контролируют белки, которые обеспечи вают функционирование физиологических систем организма – его защитных свойств, процессов дыхания, выделе ния, кровоснабжения, пищеварения и т. д. ). К таким генам относятся гены, контролирующие синтез ге моглобина, иммуноглобулина и др. В отличии от генов «домаш него хозяйства» «гены роскоши» находятся под жёстким конт ролем организма и имеют сложный аппарат регуляции.

 Классификация генов 1. Структурные гены -детерминируют синтез спе цифических продуктов 2. Гены-регуляторы – это гены, которые стимули руют или запрещают соединение РНК полимера зы (фермента, катализирующего транскрипцию) с геном 3. Гены модуляторы усиливают или ослабляют действие структурных генов, изменяя их функциональную активность.
Классификация генов 1. Структурные гены -детерминируют синтез спе цифических продуктов 2. Гены-регуляторы – это гены, которые стимули руют или запрещают соединение РНК полимера зы (фермента, катализирующего транскрипцию) с геном 3. Гены модуляторы усиливают или ослабляют действие структурных генов, изменяя их функциональную активность.
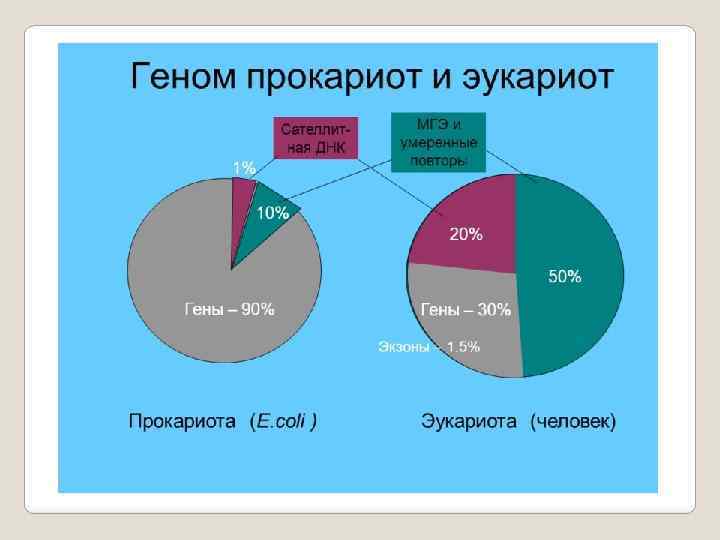
 Структурные гены Гены, которые транскрибирующие м. РНК и несущие информацию о последовательности аминокислот в полипеп тиде. Таких генов около 30 тысяч. Длина всей ДНК в клетке человека при мерно 1, 5 метра, ДНК всех генов в ней занимает всего 3 10 %. Многие из них уникальные. Однако есть гены имеющие копии. Как правило, число копий не превышает двух. Гены, с которых транскрибируется т. РНК. Эти гены не несут информацию о структуре белка. Их функция зак лючается в синтезе достаточного количества т. РНК способных обеспечить транспорт аминокислот в рибосомы для синтеза белка. Гены, с которых транскрибируются р. РНК. Эти гены, также как и предыдущие, не кодируют структуру полипептида, а синтезируют несколько разновидностей РНК.
Структурные гены Гены, которые транскрибирующие м. РНК и несущие информацию о последовательности аминокислот в полипеп тиде. Таких генов около 30 тысяч. Длина всей ДНК в клетке человека при мерно 1, 5 метра, ДНК всех генов в ней занимает всего 3 10 %. Многие из них уникальные. Однако есть гены имеющие копии. Как правило, число копий не превышает двух. Гены, с которых транскрибируется т. РНК. Эти гены не несут информацию о структуре белка. Их функция зак лючается в синтезе достаточного количества т. РНК способных обеспечить транспорт аминокислот в рибосомы для синтеза белка. Гены, с которых транскрибируются р. РНК. Эти гены, также как и предыдущие, не кодируют структуру полипептида, а синтезируют несколько разновидностей РНК.
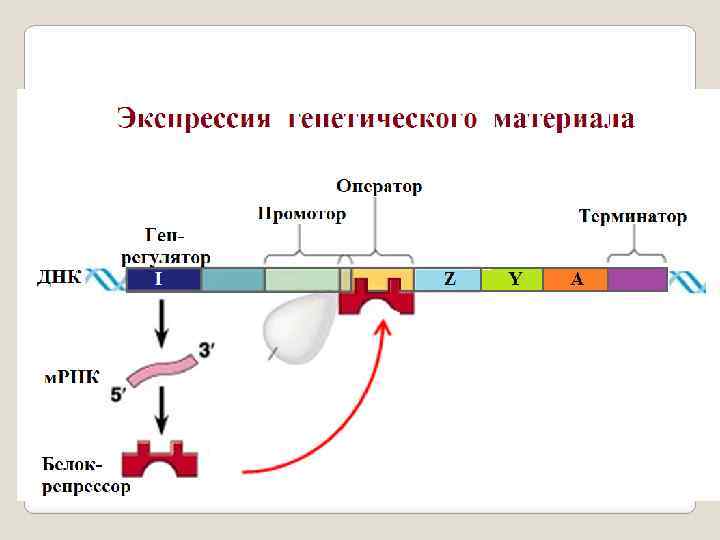
 Регуляторные гены. Эти гены регулируют активность структурных генов. Все известные регуляторные гены делятся на два типа: гены, с которых транскрибируются регуляторные РНК. Они не принимают непосредственного участия в синтезе белка, а регулируют от дельные стороны этого процесса (транскрипцию, процессинг и т. д. ). Так, например, малые ядерные РНК (мя. РНК); гены, которые несут информацию о структуре регуляторного белка. На них транскрибируется и. РНК. Этим они похожи на структурные гены. Однако, есть одно существенное отличие – на этих генах кодируется инфо рмация о регуляторном белке, который принимает участие в регуляции активности различных генетических процессов (транскрипции, трансляции, репликации, репарации и т. д. ) протекающих в клетке. Эти белки способны взаимодействовать с регуляторными областями ДНК (например с операто ром) или связываться с РНК или ДНК полимеразой. Белки носят различ ные название, например факторы транскрипции, трансляции, терминации и др. В отличии от этого и. РНК транскрибируемая на структурном гене конт ролирует синтез белка, который является участником клеточного метабо лизма выступая в роли фермента, строительного белка, белка переносчика и т. д. , но никак не белка регулятора.
Регуляторные гены. Эти гены регулируют активность структурных генов. Все известные регуляторные гены делятся на два типа: гены, с которых транскрибируются регуляторные РНК. Они не принимают непосредственного участия в синтезе белка, а регулируют от дельные стороны этого процесса (транскрипцию, процессинг и т. д. ). Так, например, малые ядерные РНК (мя. РНК); гены, которые несут информацию о структуре регуляторного белка. На них транскрибируется и. РНК. Этим они похожи на структурные гены. Однако, есть одно существенное отличие – на этих генах кодируется инфо рмация о регуляторном белке, который принимает участие в регуляции активности различных генетических процессов (транскрипции, трансляции, репликации, репарации и т. д. ) протекающих в клетке. Эти белки способны взаимодействовать с регуляторными областями ДНК (например с операто ром) или связываться с РНК или ДНК полимеразой. Белки носят различ ные название, например факторы транскрипции, трансляции, терминации и др. В отличии от этого и. РНК транскрибируемая на структурном гене конт ролирует синтез белка, который является участником клеточного метабо лизма выступая в роли фермента, строительного белка, белка переносчика и т. д. , но никак не белка регулятора.
; 2. Интенсификаторы – могут") . Гены модулято 1. Ингибиторы или супрессоры (эписта тичные гены); 2. Интенсификаторы – могут повышать способность структурных генов к мута циям; 3. Модификаторы – могут влиять на струк турные гены по типу комлементарности.
. Гены модулято 1. Ингибиторы или супрессоры (эписта тичные гены); 2. Интенсификаторы – могут повышать способность структурных генов к мута циям; 3. Модификаторы – могут влиять на струк турные гены по типу комлементарности.
 В зависимости от функции первичного продукта гены делятся на: гены ферментов, модуляторов белковой функции, рецепто ров, транскрипционных факторов, белков внутриклеточного и внеклеточного матрикса, трансмембранных переносчиков, структур ионных каналов, молекул клеточных сигналов, гор монов белковой природы, иммуноглобулинов. Количественное распределение генов, участвующих в основных процессах клеток человека, следующее: 22% сос тавляют гены, контролирующие синтез РНК и белков; 12% гены клеточного деления, 12% клеточные сигналы, 12% защита клетки, 17% обмен веществ, 8% клеточные структ уры, 17% функция неизвестна.
В зависимости от функции первичного продукта гены делятся на: гены ферментов, модуляторов белковой функции, рецепто ров, транскрипционных факторов, белков внутриклеточного и внеклеточного матрикса, трансмембранных переносчиков, структур ионных каналов, молекул клеточных сигналов, гор монов белковой природы, иммуноглобулинов. Количественное распределение генов, участвующих в основных процессах клеток человека, следующее: 22% сос тавляют гены, контролирующие синтез РНК и белков; 12% гены клеточного деления, 12% клеточные сигналы, 12% защита клетки, 17% обмен веществ, 8% клеточные структ уры, 17% функция неизвестна.
") Наряду с приведенной ранее функциональной классификацией генов существуют и другие разновидности генов: Мобильные (прыгающие) гены – транспозоны, это участки ДНК организмов, способные к передвижению (транспозиции) и размно жению в пределах генома. Эти гены, не имеют постоянной локали зации в хромосоме и в пределах хромосомного набора клетки. Перемещения генов влияют на их экспрессию – ранее не активные гены могут активизироваться, и наоборот. Ген-кандидат это ген, измененный продукт которого может стать причиной определенного наследственного заболевания. Псевдоген – это ген сходный по нуклеотидной последовательности с известным геном, но не выполняющий такую же функцию либо из за потери промотора, либо несущий мутацию, которая препятствует его экспрессии.
Наряду с приведенной ранее функциональной классификацией генов существуют и другие разновидности генов: Мобильные (прыгающие) гены – транспозоны, это участки ДНК организмов, способные к передвижению (транспозиции) и размно жению в пределах генома. Эти гены, не имеют постоянной локали зации в хромосоме и в пределах хромосомного набора клетки. Перемещения генов влияют на их экспрессию – ранее не активные гены могут активизироваться, и наоборот. Ген-кандидат это ген, измененный продукт которого может стать причиной определенного наследственного заболевания. Псевдоген – это ген сходный по нуклеотидной последовательности с известным геном, но не выполняющий такую же функцию либо из за потери промотора, либо несущий мутацию, которая препятствует его экспрессии.
 Гены, от которых зависит развитие альтер нативных признаков называются аллельными. Аллельные гены расположены в одинако вых локусах гомологичных хромосом и имеют 2 состояния доминантное и рецессивное.
Гены, от которых зависит развитие альтер нативных признаков называются аллельными. Аллельные гены расположены в одинако вых локусах гомологичных хромосом и имеют 2 состояния доминантное и рецессивное.
 Современная теория гена. 1. Ген занимает определенный локус в хромосоме. 2. Ген - часть молекулы ДНК; число нуклеотидов в гене неодинаково. 3. Внутри гена может происходить рекомбинация и мутация. 4. 5. Существуют структурные и функциональные гены. 6. Структурные гены контролируют синтез полипептидов (аминокислотных, т-РНК, р-РНК) и белков. 7. 8. Функциональные гены контролируют деятельность структурных генов. 9. 10. Расположение триплетов в генах структурных колинеарно последовательности аминокислот в полипептиде. 11. Генотип, будучи дискретным, функционирует как единое целое.
Современная теория гена. 1. Ген занимает определенный локус в хромосоме. 2. Ген - часть молекулы ДНК; число нуклеотидов в гене неодинаково. 3. Внутри гена может происходить рекомбинация и мутация. 4. 5. Существуют структурные и функциональные гены. 6. Структурные гены контролируют синтез полипептидов (аминокислотных, т-РНК, р-РНК) и белков. 7. 8. Функциональные гены контролируют деятельность структурных генов. 9. 10. Расположение триплетов в генах структурных колинеарно последовательности аминокислот в полипептиде. 11. Генотип, будучи дискретным, функционирует как единое целое.
 Свойства гена: 1. Ген хранит и передает информацию. 2. Ген способен к изменению. генетической информации (мутации). 3. Ген способен к репарации и ее передаче от поколения к поколению. 4. Ген способен к реализации - синтезу белка. 5. Генетический материал обладает устойчивостью.
Свойства гена: 1. Ген хранит и передает информацию. 2. Ген способен к изменению. генетической информации (мутации). 3. Ген способен к репарации и ее передаче от поколения к поколению. 4. Ген способен к реализации - синтезу белка. 5. Генетический материал обладает устойчивостью.
 Свойства генов: 1. Дискретность действия – каждый ген действует как самостоятельная единица наследственности. 2. Стабильность – при отсутствии мутаций ген передается в ряду поко лений в неизмененном виде. 3. Специфичность действия – каждый ген влияет на развитие своего опре деленного признака. 4. Плейотропность – способность одного гена обеспечивать развитие одно временно нескольких признаков (синдром Марфана). 5. Множественный аллелизм присутствие в виде двух у диплоидных и большего числа аллелей у полиплоидных организмов. 6. Лабильность – способность многократно мутировать 7. Пенетрантность – частота проявления гена в фенотипе 8. Дозированность гена действие гена дозировано, при изменении числа доз гена в организменяется признак (например, при болезни Дауна происходит увеличение до трех доз генов 21 хромосомы)
Свойства генов: 1. Дискретность действия – каждый ген действует как самостоятельная единица наследственности. 2. Стабильность – при отсутствии мутаций ген передается в ряду поко лений в неизмененном виде. 3. Специфичность действия – каждый ген влияет на развитие своего опре деленного признака. 4. Плейотропность – способность одного гена обеспечивать развитие одно временно нескольких признаков (синдром Марфана). 5. Множественный аллелизм присутствие в виде двух у диплоидных и большего числа аллелей у полиплоидных организмов. 6. Лабильность – способность многократно мутировать 7. Пенетрантность – частота проявления гена в фенотипе 8. Дозированность гена действие гена дозировано, при изменении числа доз гена в организменяется признак (например, при болезни Дауна происходит увеличение до трех доз генов 21 хромосомы)
 можно") Новая концепция гена. Благодаря альтернативному сплайсингу из одной молекулы первичного транскрипта (пре-м. РНК) можно получить несколько различных вариантов молекул зрелых м. РНК с разным набором экзонов. В связи с этим можно говорить о новой концепции гена: один ген – много РНК – много полипептидов и сформулировать новое определение гена. Ген - это участок молекулы ДНК, кодирующий синтез одной или нескольких функциональных молекул РНК.
Новая концепция гена. Благодаря альтернативному сплайсингу из одной молекулы первичного транскрипта (пре-м. РНК) можно получить несколько различных вариантов молекул зрелых м. РНК с разным набором экзонов. В связи с этим можно говорить о новой концепции гена: один ген – много РНК – много полипептидов и сформулировать новое определение гена. Ген - это участок молекулы ДНК, кодирующий синтез одной или нескольких функциональных молекул РНК.
 Генетический материал клетки. Хромосомный уровень организации генетического материала
Генетический материал клетки. Хромосомный уровень организации генетического материала
 Хромосомный уровень организации генетического материала Хромосомы - это нуклеопротеидные тела, в которых хранится, передается потомству и реализуется наследственная информация. Хромосомы клеток эукариот, имеют разную морфологию в митозе и интерфазе. ДНК (40%), белки ( около 60%), РНК (4%), липиды, углеводы, ионы металлов (менее 1%) Роль компонентов хромосом заключается в «разрешении» или «запре щении» считывания информации с молекулы ДНК.
Хромосомный уровень организации генетического материала Хромосомы - это нуклеопротеидные тела, в которых хранится, передается потомству и реализуется наследственная информация. Хромосомы клеток эукариот, имеют разную морфологию в митозе и интерфазе. ДНК (40%), белки ( около 60%), РНК (4%), липиды, углеводы, ионы металлов (менее 1%) Роль компонентов хромосом заключается в «разрешении» или «запре щении» считывания информации с молекулы ДНК.
 ромосомы вирусов прокариот и леточных органелл эукариот Одна молекула двуспиральной ДНК; ее на бактериальной хромосомой Для большинства бактерий и плазмид обычна суперспирализированная форма. У микроорганизмов ряда видов встречаются ДНК в линейной форме .
ромосомы вирусов прокариот и леточных органелл эукариот Одна молекула двуспиральной ДНК; ее на бактериальной хромосомой Для большинства бактерий и плазмид обычна суперспирализированная форма. У микроорганизмов ряда видов встречаются ДНК в линейной форме .
 ДНК бактерий кольцевая двухцепочная, упакована в структуру, которая называется нуклеоид. ДНК. Белки HU, INF, H 1, HLP, H, принимают участие в компактизации ДНК
ДНК бактерий кольцевая двухцепочная, упакована в структуру, которая называется нуклеоид. ДНК. Белки HU, INF, H 1, HLP, H, принимают участие в компактизации ДНК

 Роль компонентов хромосом заключается в «разрешении» или «запрещении» считывания информации с молекулы ДНК, поэтому хромосомы могут находится в состояниях: генетически активном (деспирализованном) с осуществлением процессов репликации и транскрипции генетически неактивном (спирализованном), осуществляющим перенос информации из родительских клеток в дочерние в процессе митоза На разных этапах клеточного цикла молекулы ДНК упакованы в нуклеопротеиновые структуры – хроматин. В фазе деления ядра выявляется в виде компактных структур, называемых метафазной хромосомой. В интерфазе он распределен равномерно по всему объему ядра и не выявляется обычными микроскопическими методами и называется интерфазной хромосомой
Роль компонентов хромосом заключается в «разрешении» или «запрещении» считывания информации с молекулы ДНК, поэтому хромосомы могут находится в состояниях: генетически активном (деспирализованном) с осуществлением процессов репликации и транскрипции генетически неактивном (спирализованном), осуществляющим перенос информации из родительских клеток в дочерние в процессе митоза На разных этапах клеточного цикла молекулы ДНК упакованы в нуклеопротеиновые структуры – хроматин. В фазе деления ядра выявляется в виде компактных структур, называемых метафазной хромосомой. В интерфазе он распределен равномерно по всему объему ядра и не выявляется обычными микроскопическими методами и называется интерфазной хромосомой
 ХРОМАТИН - ХРОМОСОМЫ • Хроматин – это частично упакованный генетический материал в интерфазы; • Хромосома – максимально упакованный генетический материал в митозе. • Химический состав: • 30% ДНК + 40% гистоны + 25% негистоновые белки + 5% РНК
ХРОМАТИН - ХРОМОСОМЫ • Хроматин – это частично упакованный генетический материал в интерфазы; • Хромосома – максимально упакованный генетический материал в митозе. • Химический состав: • 30% ДНК + 40% гистоны + 25% негистоновые белки + 5% РНК
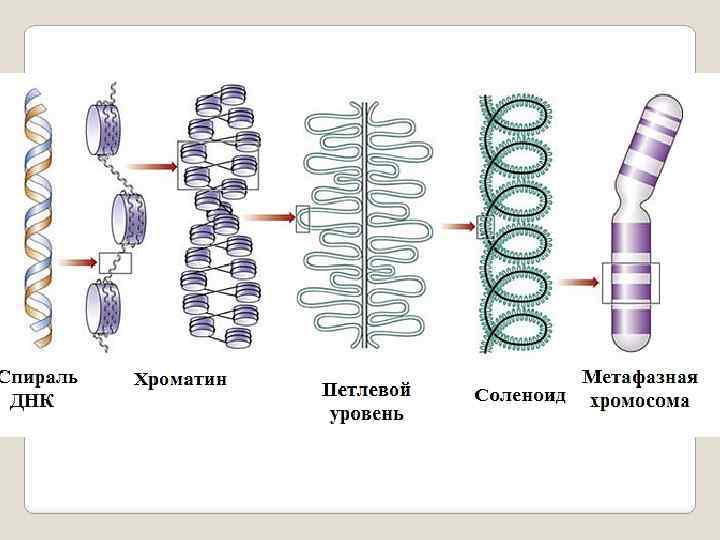
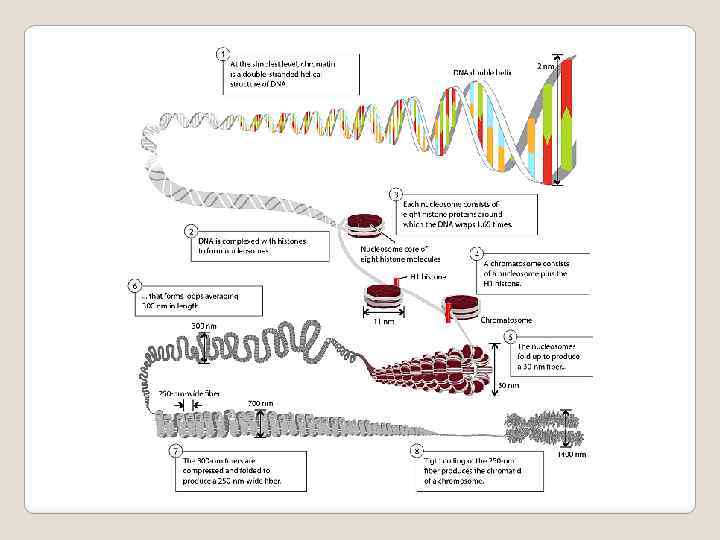
: нуклеосомная нить, хроматиновая фибрилла,") В процессе клеточного цикла хроматин претерпевает несколько уровней спирализации (компактизации): нуклеосомная нить, хроматиновая фибрилла, хромомеры, хромонемы, хроматиды. В ядрах клетки человека суммарная длина ДНК составляет почти 2 метра. Диаметр ядра - 5 -10 микрон, то есть в миллион раз меньше длины ДНК Самая маленькая хромосома человека - 22 я, составляет около 1, 4 см в длину и содержит 4, 6 ∙ 107 пар оснований. Для размещения молекулы ДНК на метафазной пластинке в процессе митоза, уменьшения риска спутывания или разрыва ДНК, необходимо упаковать ее в более короткие пучки – хромосомы. К концу профазы митоза 22 я хромосома укорачивается до 2 мкм, то есть в 7000 раз. Для достижения такого уровня компактизации и одно -временно сохранения эффективности основных генети - ческих процессов, требующих локальной распаковки, структура метафазной хромосомы должна пройти неско- лько уровней компактизации, что является основой складчато-петлевой модели структуры хромосомы
В процессе клеточного цикла хроматин претерпевает несколько уровней спирализации (компактизации): нуклеосомная нить, хроматиновая фибрилла, хромомеры, хромонемы, хроматиды. В ядрах клетки человека суммарная длина ДНК составляет почти 2 метра. Диаметр ядра - 5 -10 микрон, то есть в миллион раз меньше длины ДНК Самая маленькая хромосома человека - 22 я, составляет около 1, 4 см в длину и содержит 4, 6 ∙ 107 пар оснований. Для размещения молекулы ДНК на метафазной пластинке в процессе митоза, уменьшения риска спутывания или разрыва ДНК, необходимо упаковать ее в более короткие пучки – хромосомы. К концу профазы митоза 22 я хромосома укорачивается до 2 мкм, то есть в 7000 раз. Для достижения такого уровня компактизации и одно -временно сохранения эффективности основных генети - ческих процессов, требующих локальной распаковки, структура метафазной хромосомы должна пройти неско- лько уровней компактизации, что является основой складчато-петлевой модели структуры хромосомы
 Уровни компактизации молекулы ДНК в процессе клеточного цикла
Уровни компактизации молекулы ДНК в процессе клеточного цикла
 Уровни компактизации ДНК I уровень Нуклеосомный • Полинуклеосомная нить диаметром 11 нм хроматин • Степень конденсации – x 6 ; • Механизм образования: взаимодействие ДНК с гистоновыми основными белками • Транскрипция возможна.
Уровни компактизации ДНК I уровень Нуклеосомный • Полинуклеосомная нить диаметром 11 нм хроматин • Степень конденсации – x 6 ; • Механизм образования: взаимодействие ДНК с гистоновыми основными белками • Транскрипция возможна.
 субъединица ХРОМАТИНА, состоящая из ДНК и набора из четырех пар") Нуклеосомный уровень Нуклеосома (nucleosome) субъединица ХРОМАТИНА, состоящая из ДНК и набора из четырех пар гистоновых белков Н 2 А, Н 2 В, Н 3 и Н 4 и одной моле кулы гистона Н 1. Гистон Н 1 связывается с линкерной ДНК (участки ДНК между нуклеосомами). Участок ДНК накрученный на кор состав ляет 150 н. п. ; кор вместе с участком ДНК называется нуклеосома. Молекула ДНК, делая вокруг цилиндра около двух оборотов. Участок ДНК накрученный на кор составляет 150 н. п. ; кор вместе с участком ДНК называется нуклеосома.
Нуклеосомный уровень Нуклеосома (nucleosome) субъединица ХРОМАТИНА, состоящая из ДНК и набора из четырех пар гистоновых белков Н 2 А, Н 2 В, Н 3 и Н 4 и одной моле кулы гистона Н 1. Гистон Н 1 связывается с линкерной ДНК (участки ДНК между нуклеосомами). Участок ДНК накрученный на кор состав ляет 150 н. п. ; кор вместе с участком ДНК называется нуклеосома. Молекула ДНК, делая вокруг цилиндра около двух оборотов. Участок ДНК накрученный на кор составляет 150 н. п. ; кор вместе с участком ДНК называется нуклеосома.
 Нуклеосомный уровень Образуется при взаимодействии ДНК и гистонов в составе хроматина. Хроматин представляет собой нить, с нанизанными на нее бусинами, где каждая бусина является нуклеосомой. Электронная микрофотография нити хроматина с нуклеосомами из клеток ядер цыпленка
Нуклеосомный уровень Образуется при взаимодействии ДНК и гистонов в составе хроматина. Хроматин представляет собой нить, с нанизанными на нее бусинами, где каждая бусина является нуклеосомой. Электронная микрофотография нити хроматина с нуклеосомами из клеток ядер цыпленка
 II уровень – нуклеомерный или соленоидный • Нить хроматина диаметром 30 нм; • Степень конденсации – x 40 • Механизм образования: фосфорилирование H 1, сверхспирализация полинуклеосомной нити – по 6 нуклеосом в витке; • Блокирование транскрипции ДНК.
II уровень – нуклеомерный или соленоидный • Нить хроматина диаметром 30 нм; • Степень конденсации – x 40 • Механизм образования: фосфорилирование H 1, сверхспирализация полинуклеосомной нити – по 6 нуклеосом в витке; • Блокирование транскрипции ДНК.
 Нуклеомерный уровень Дальнейшая компактизация ДНК в соста ве хроматина свя зана с образованием нуклеосомных комплексов нуклеомеров. Этот уровень компактизации обеспечива ется гистоном Н 1, который будучи связа нным с линкерной ДНК, связывается также с соседними нуклеосомами и, изменяя свою конформацию, сближает их. Образуется компактная хроматиновая фибрилла построенная либо по типу соле ноида (спиральный тип укладки), либо по нуклеомерному типу (4 12 нуклеосом образуют глобулу нуклеомер). Такая структура называется элементарной хроматиновой фибриллой или нуклеомером; имеет диаметр 30 нм и длину 1, 2 мм. Уровень компактизации составляет 40 раз. На один нуклеомер приходится 1600 н. п.
Нуклеомерный уровень Дальнейшая компактизация ДНК в соста ве хроматина свя зана с образованием нуклеосомных комплексов нуклеомеров. Этот уровень компактизации обеспечива ется гистоном Н 1, который будучи связа нным с линкерной ДНК, связывается также с соседними нуклеосомами и, изменяя свою конформацию, сближает их. Образуется компактная хроматиновая фибрилла построенная либо по типу соле ноида (спиральный тип укладки), либо по нуклеомерному типу (4 12 нуклеосом образуют глобулу нуклеомер). Такая структура называется элементарной хроматиновой фибриллой или нуклеомером; имеет диаметр 30 нм и длину 1, 2 мм. Уровень компактизации составляет 40 раз. На один нуклеомер приходится 1600 н. п.
 II уровень – нуклеомерный или соленоидный
II уровень – нуклеомерный или соленоидный
 Третий уровень — петельный Петли • Петли большие и малые, диффузные или компактные, диаметр фибриллы– 300 нм; • Механизм образования: прикрепление нити хроматина 30 нм к белковой оси – скаффолду; взаимодействие SAR – скаффолд • Транскрипция генов – заблокирована Укорочение фибриллы на этом уровне происходит в среднем 25 раз, а на всех 3 уровнях в 1000 1500 раз. Такую петельную структуру называют интерфазной хромонемой.
Третий уровень — петельный Петли • Петли большие и малые, диффузные или компактные, диаметр фибриллы– 300 нм; • Механизм образования: прикрепление нити хроматина 30 нм к белковой оси – скаффолду; взаимодействие SAR – скаффолд • Транскрипция генов – заблокирована Укорочение фибриллы на этом уровне происходит в среднем 25 раз, а на всех 3 уровнях в 1000 1500 раз. Такую петельную структуру называют интерфазной хромонемой.
 Третий уровень — петлево Соленоидная хроматиновая фибрилла складывается в петли разной длины. В образовании петель участвуют негистоновые белки ядерного матрикса, которые способны узнавать специфические последовательности ДНК (SAR- последовательности), отдаленные друг от друга на рассто янии в несколько тысяч пар нуклеотидов. Эти белки образуют остов (scaffold), который связывается с SAR-последовательностями ДНК (scaffold attachment regions) и, сближая их, образует петли. Каждая петля содержит от 20 до 80 тыс. пар нуклеотидов, что может соответ ствовать одному или нескольким генам и называется петлевой домен. Средняя длина петли (300 400 нм) сходна у различных организмов (дрозофила и человек) и включает примерно 50 тысяч оснований. В интерфазном ядре такие петли связаны с сетчатым белковым образованием, расположенным на внутренней стороне ядерной оболочки и называемым сетчатым ядерным матриксом, ламиной или ядерной пластинкой. Белки ядерного матрикса ламины соединяются с последовательностями ДНК, которые называются МАR-последовательностями (matrix attachment regions). В результате такой спирализации хроматиновая фибрилла диаметром 30 нм преобразуется в структуру диаметром 300 нм, называемую интерфазной хромонемой.
Третий уровень — петлево Соленоидная хроматиновая фибрилла складывается в петли разной длины. В образовании петель участвуют негистоновые белки ядерного матрикса, которые способны узнавать специфические последовательности ДНК (SAR- последовательности), отдаленные друг от друга на рассто янии в несколько тысяч пар нуклеотидов. Эти белки образуют остов (scaffold), который связывается с SAR-последовательностями ДНК (scaffold attachment regions) и, сближая их, образует петли. Каждая петля содержит от 20 до 80 тыс. пар нуклеотидов, что может соответ ствовать одному или нескольким генам и называется петлевой домен. Средняя длина петли (300 400 нм) сходна у различных организмов (дрозофила и человек) и включает примерно 50 тысяч оснований. В интерфазном ядре такие петли связаны с сетчатым белковым образованием, расположенным на внутренней стороне ядерной оболочки и называемым сетчатым ядерным матриксом, ламиной или ядерной пластинкой. Белки ядерного матрикса ламины соединяются с последовательностями ДНК, которые называются МАR-последовательностями (matrix attachment regions). В результате такой спирализации хроматиновая фибрилла диаметром 30 нм преобразуется в структуру диаметром 300 нм, называемую интерфазной хромонемой.
 Третий уровень — петельный Схема ядерной мембраны
Третий уровень — петельный Схема ядерной мембраны
 етий уровень — петельный
етий уровень — петельный
 етий уровень — петельный
етий уровень — петельный
 Третий уровень — петельный
Третий уровень — петельный
 Четвертый уровень – хромонемный или доменный Отдельные участки интерфазной хромонемы подвергаются даль нейшей компактизации : соседние петли объединяются в струк турные блоки в виде розеток , петли которых, сильно спирали зуясь, образуют хромомеры, небольшие, сильно окрашивающиеся компактные тельца (0, 1 0, 20 мкм), различающиеся по форме и раз меру. Этот уровень спирализации ведёт к образованию хроматиды с диаметром 600 -700 нм.
Четвертый уровень – хромонемный или доменный Отдельные участки интерфазной хромонемы подвергаются даль нейшей компактизации : соседние петли объединяются в струк турные блоки в виде розеток , петли которых, сильно спирали зуясь, образуют хромомеры, небольшие, сильно окрашивающиеся компактные тельца (0, 1 0, 20 мкм), различающиеся по форме и раз меру. Этот уровень спирализации ведёт к образованию хроматиды с диаметром 600 -700 нм.
 Четвертый уровень – хромонемный или доменный Петлевые домены и хромомеры расположены неравно мерно. Неодинаковая степень спирализации разных учас тков хромосом имеет большое значение; в зависимости от степени спирализации выделяют эухроматин и гетерохроматин. Число и рисунок хромомер у данного организма на данной стадии клеточного цикла одинаковы и постоянны. Хромоме ры — утолщён ные, плотно спирализован ные участки нитей ДНК (хромонем), из которых состоит хромосома Хромомеры хромосомы XII тритона
Четвертый уровень – хромонемный или доменный Петлевые домены и хромомеры расположены неравно мерно. Неодинаковая степень спирализации разных учас тков хромосом имеет большое значение; в зависимости от степени спирализации выделяют эухроматин и гетерохроматин. Число и рисунок хромомер у данного организма на данной стадии клеточного цикла одинаковы и постоянны. Хромоме ры — утолщён ные, плотно спирализован ные участки нитей ДНК (хромонем), из которых состоит хромосома Хромомеры хромосомы XII тритона
 V уровень – метафазная хромо На хромосомном уровне происходит дальнейшая компактизация хромомеров вокруг осевого каркаса негистоновых белков. Эта супер спирализация сопровождается фосфорилированием в клетке всех мо лекул H 1. Наблюдается в делящихся клетках: в них хромосомы нас только плотны, что становятся видны в световой микроскоп как от дельные образования метафазная хромосома. В неделящейся клет ке хромосомы деспирализованы (деконденсированы), и называются хроматином. В результате метафазную хромосому можно изобразить в виде плотно уложенных соленоидных петель, свернутых в тугую спираль. Типичная хромосома человека может содержать до 2600 петель. Тол щина такой структуры достигает 1400 нм (две хрома тиды), а молекула ДНК при этом укорачивается в 104 раз, т. е. с 2 см растянутой ДНК до 2 мкм. Таким образом, процесс образования хромосом – сложное структурно морфологическое преобразование, в системе «молекула ДНК → хроматин (ДНП) → нуклеосома → хромонема → хромосома» .
V уровень – метафазная хромо На хромосомном уровне происходит дальнейшая компактизация хромомеров вокруг осевого каркаса негистоновых белков. Эта супер спирализация сопровождается фосфорилированием в клетке всех мо лекул H 1. Наблюдается в делящихся клетках: в них хромосомы нас только плотны, что становятся видны в световой микроскоп как от дельные образования метафазная хромосома. В неделящейся клет ке хромосомы деспирализованы (деконденсированы), и называются хроматином. В результате метафазную хромосому можно изобразить в виде плотно уложенных соленоидных петель, свернутых в тугую спираль. Типичная хромосома человека может содержать до 2600 петель. Тол щина такой структуры достигает 1400 нм (две хрома тиды), а молекула ДНК при этом укорачивается в 104 раз, т. е. с 2 см растянутой ДНК до 2 мкм. Таким образом, процесс образования хромосом – сложное структурно морфологическое преобразование, в системе «молекула ДНК → хроматин (ДНП) → нуклеосома → хромонема → хромосома» .
 Перед началом деления ядра происходит удвоение хромонем, то есть их репликация и самовоспроизведение. При удвоении хромосомного аппарата обе сестринские хромонемы становятся хроматидами и укладываются спирально или петлеобразно вместе, образуя метафазную хромосому. В этом случае упакованная хромосомная нить достигает 700 нм в ширину.
Перед началом деления ядра происходит удвоение хромонем, то есть их репликация и самовоспроизведение. При удвоении хромосомного аппарата обе сестринские хромонемы становятся хроматидами и укладываются спирально или петлеобразно вместе, образуя метафазную хромосому. В этом случае упакованная хромосомная нить достигает 700 нм в ширину.
: Б – схема метафазной хромосомы: В") . А – фото метафазной хромосомы (флюоресцирующий микроскоп): Б – схема метафазной хромосомы: В – схема соединения двух хроматид в районе центромеры: 4 а - участок соединения двух хроматид белком когезином(6), тоже в увеличенном виде
. А – фото метафазной хромосомы (флюоресцирующий микроскоп): Б – схема метафазной хромосомы: В – схема соединения двух хроматид в районе центромеры: 4 а - участок соединения двух хроматид белком когезином(6), тоже в увеличенном виде
 Эухроматин, активный хроматин — участки хроматина, деспирализованные, активно транскрибируемые в интерфазе и неактивные в митозе. Эухроматин содержит большинство структурных генов организма. Гетерохромати н — учас тки хроматина, находя щиеся в течение клеточ ного цикла в конденси рованном (компактном) состоянии. Особенностью гетеро хроматиновой ДНК является крайне низкая тран скрибируемость.
Эухроматин, активный хроматин — участки хроматина, деспирализованные, активно транскрибируемые в интерфазе и неактивные в митозе. Эухроматин содержит большинство структурных генов организма. Гетерохромати н — учас тки хроматина, находя щиеся в течение клеточ ного цикла в конденси рованном (компактном) состоянии. Особенностью гетеро хроматиновой ДНК является крайне низкая тран скрибируемость.
 Гетерохроматин – это высококомпактизированный и "молчащий" хро матин. Конститутивный и факультативный хроматин. У первом виде гены . организма экспрессируются лишь изредка в клетках любого типа; второй репрессирован в некоторых клетках в ходе специфического клеточного цикла или на специфической стадии развития. локализован преимущест венно в участках центромер, теломер и ядрышковом организаторе. Эухроматин — участки хроматина, деконденси рующиеся в конце митоза и слабо окрашивающи еся в интерфазных ядрах. В Э. расположено подав ляющее большинство активно транскрибируемых генов; содержит относительно большее количество негистоновых белков в сравнении с гетерохромати ном. Эухроматин характеризуется меньшей по срав нению с гетерохроматином компактизацией ДНК, и в нем главным образом локализуются активно экспрессирующиеся гены. Эухроматин (светлые участки) и гетерохроматин (темные участки) в кариотипе дрозофилы
Гетерохроматин – это высококомпактизированный и "молчащий" хро матин. Конститутивный и факультативный хроматин. У первом виде гены . организма экспрессируются лишь изредка в клетках любого типа; второй репрессирован в некоторых клетках в ходе специфического клеточного цикла или на специфической стадии развития. локализован преимущест венно в участках центромер, теломер и ядрышковом организаторе. Эухроматин — участки хроматина, деконденси рующиеся в конце митоза и слабо окрашивающи еся в интерфазных ядрах. В Э. расположено подав ляющее большинство активно транскрибируемых генов; содержит относительно большее количество негистоновых белков в сравнении с гетерохромати ном. Эухроматин характеризуется меньшей по срав нению с гетерохроматином компактизацией ДНК, и в нем главным образом локализуются активно экспрессирующиеся гены. Эухроматин (светлые участки) и гетерохроматин (темные участки) в кариотипе дрозофилы
 Морфология метафазных хромосом 1 - центромера и кинетохор Играет роль в движении хромосом к полюсам деления и точном распределении дочерних хроматид по дочерним клеткам в процессе митоза и мейоза. 3 - теломеры 2 - вторичные перетяжки, которые отделяют участки коротких плеч, называемые саттелиты (спутники) (4), состоящие из конститутивного - защищают концы хромосом от действия нуклеаз; - предотвращают слипание концов хромосом; - обеспечивают репликацию всей ДНК; - контролируют процессы старения клеток и многоклеточного организма; - регулируют фиксацию хроматина к ядерной мембране в интерфазе, - обеспечивают правильную конъ- Комплекс, содержащий участок ДНК центромеры, ДНК-связывающие белки, РНК, тубулин. Служит: 1 - центром прикрепления микротрубочек веретена деления; 2 - выравнивания хромосом в метафазной пластинке на экваторе деления клетки в метафазе; 3 - участвует в процессе расхождения хромосом во время анафазы
Морфология метафазных хромосом 1 - центромера и кинетохор Играет роль в движении хромосом к полюсам деления и точном распределении дочерних хроматид по дочерним клеткам в процессе митоза и мейоза. 3 - теломеры 2 - вторичные перетяжки, которые отделяют участки коротких плеч, называемые саттелиты (спутники) (4), состоящие из конститутивного - защищают концы хромосом от действия нуклеаз; - предотвращают слипание концов хромосом; - обеспечивают репликацию всей ДНК; - контролируют процессы старения клеток и многоклеточного организма; - регулируют фиксацию хроматина к ядерной мембране в интерфазе, - обеспечивают правильную конъ- Комплекс, содержащий участок ДНК центромеры, ДНК-связывающие белки, РНК, тубулин. Служит: 1 - центром прикрепления микротрубочек веретена деления; 2 - выравнивания хромосом в метафазной пластинке на экваторе деления клетки в метафазе; 3 - участвует в процессе расхождения хромосом во время анафазы
 От места положения центромеры различают хромосомы:
От места положения центромеры различают хромосомы:
 Набор хромосом диплоидной клетки называется кариотип 22 пары хромосом идентичны у мужчин и женщин и называются аутосомами Нормальный кариотип соматической клетки человека представлен 23 парами хромосом одна пара называется половыми хромо -сомами: у мужчин представлена Х и Y хромосомами, у женщин – двумя Х-хро -мосомами. Кариот ип Во всех парах хромосом одна получена от отца, другая – от матери. В половых клетках (сперматозоидах и яйцеклетках) содержится гаплоидный набор хромосом, то есть, 23 хромосомы. Сперматозоиды делятся на два типа: Хсперматозоиды (содержат Х-хромосому) и Y -сперматозоиды (содержат Y-хромосому). Яйцеклетки содержат только Х-хромо В S-периоде интерфазы набор хромосом соматической клет -ки обозначается как 2 n 4 c. n – число хромосом в гаплоидном наборе, с – количество ДНК Диплоидный набор хромосом обозначается 2 n 2 с, гаплоидный - 1 n 1 с.
Набор хромосом диплоидной клетки называется кариотип 22 пары хромосом идентичны у мужчин и женщин и называются аутосомами Нормальный кариотип соматической клетки человека представлен 23 парами хромосом одна пара называется половыми хромо -сомами: у мужчин представлена Х и Y хромосомами, у женщин – двумя Х-хро -мосомами. Кариот ип Во всех парах хромосом одна получена от отца, другая – от матери. В половых клетках (сперматозоидах и яйцеклетках) содержится гаплоидный набор хромосом, то есть, 23 хромосомы. Сперматозоиды делятся на два типа: Хсперматозоиды (содержат Х-хромосому) и Y -сперматозоиды (содержат Y-хромосому). Яйцеклетки содержат только Х-хромо В S-периоде интерфазы набор хромосом соматической клет -ки обозначается как 2 n 4 c. n – число хромосом в гаплоидном наборе, с – количество ДНК Диплоидный набор хромосом обозначается 2 n 2 с, гаплоидный - 1 n 1 с.
 диплоидного набора") Кариоти п — совокупность признаков (число, размеры, форма и т. д. ) диплоидного набора хромосом, присущая соматическим клеткам дан ного биологического вида (видовой кариотип). В 1960 году на конференции в американском городе Денвере была создана первая Международная система цитогенетической номенклатуры хромосом человека, обеспечившая международную стандартизацию исследований хромосом еще на начальных этапах становления цитогенетики человека. В основу Денверской классификации хромосом была положена их морфологическая характеристика: размер, форма и положение первичной перетяжки - центромеры. Согласно данной номенклатуре хромосомы нумеруются от 1 до 23 по мере убывания их длины: с 1 по 22 - аутосомы, а 23 пара- половые хромосомы. Самые крупные хромосомы человека, имеющие первые номера, в среднем 5 раз длиннее самых мелких - 21 и 22 хромосом.
Кариоти п — совокупность признаков (число, размеры, форма и т. д. ) диплоидного набора хромосом, присущая соматическим клеткам дан ного биологического вида (видовой кариотип). В 1960 году на конференции в американском городе Денвере была создана первая Международная система цитогенетической номенклатуры хромосом человека, обеспечившая международную стандартизацию исследований хромосом еще на начальных этапах становления цитогенетики человека. В основу Денверской классификации хромосом была положена их морфологическая характеристика: размер, форма и положение первичной перетяжки - центромеры. Согласно данной номенклатуре хромосомы нумеруются от 1 до 23 по мере убывания их длины: с 1 по 22 - аутосомы, а 23 пара- половые хромосомы. Самые крупные хромосомы человека, имеющие первые номера, в среднем 5 раз длиннее самых мелких - 21 и 22 хромосом.
 Хромосомный набор или кариотип человека включает 46 хромосом - 22 пары аутосом и 1 пару половых хромосом (XX- у лиц женского пола и XY - мужского). В кариотипе человека были выделены 7 групп аутосом и половые хромосомы X и Y. Хромосомы расположены по мере убывания их размеров. Группам аутосом были присвоены буквенные обозначения : А (1– 3), B (4– 5), C(6– 12), D (13– 15), E (16– 18), F (19– 20), G (21– 22). Х-хромосома относится к С-группе, Y – к G-группе. В скобках указаны номера хромосом каждой группы. Каждая пара хромосом идентифицируется по размеру и положению центромеры (метацентрические, субметацентрические, акроцентрические).
Хромосомный набор или кариотип человека включает 46 хромосом - 22 пары аутосом и 1 пару половых хромосом (XX- у лиц женского пола и XY - мужского). В кариотипе человека были выделены 7 групп аутосом и половые хромосомы X и Y. Хромосомы расположены по мере убывания их размеров. Группам аутосом были присвоены буквенные обозначения : А (1– 3), B (4– 5), C(6– 12), D (13– 15), E (16– 18), F (19– 20), G (21– 22). Х-хромосома относится к С-группе, Y – к G-группе. В скобках указаны номера хромосом каждой группы. Каждая пара хромосом идентифицируется по размеру и положению центромеры (метацентрические, субметацентрические, акроцентрические).
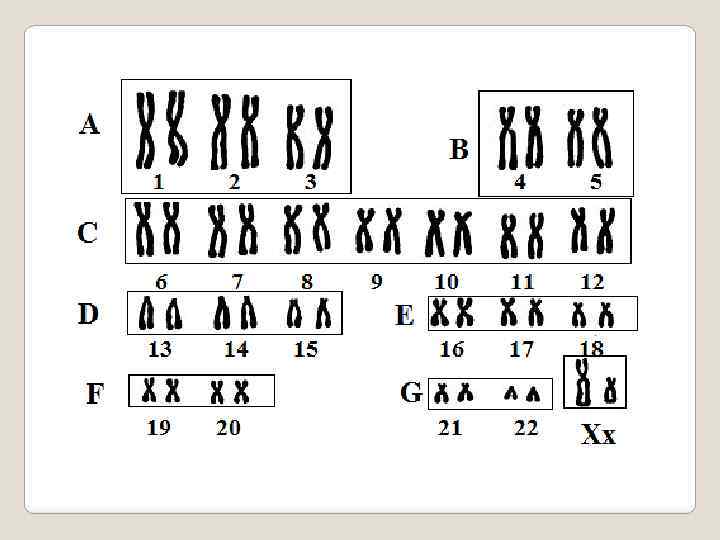
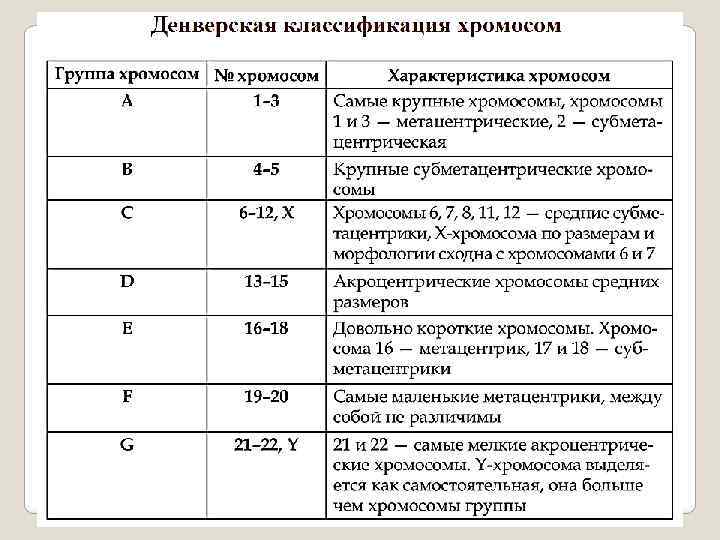
 Денверовская классификация кариотипа
Денверовская классификация кариотипа
 Парижская классификация. В 1971 году в Париже на IV международном конгрессе по генетике человека была согласована единая система идентификации хромосом человека, учитывавшая дифференцировку хромосом по длине при окрашивании. Каждая хромосома набора человека при дифференциальной окраске характеризуется уникальным для нее сочетанием темно окрашенных сегментов или полос, чередующихся с неокрашенными участками или светлыми сегментами.
Парижская классификация. В 1971 году в Париже на IV международном конгрессе по генетике человека была согласована единая система идентификации хромосом человека, учитывавшая дифференцировку хромосом по длине при окрашивании. Каждая хромосома набора человека при дифференциальной окраске характеризуется уникальным для нее сочетанием темно окрашенных сегментов или полос, чередующихся с неокрашенными участками или светлыми сегментами.
 Хромосома делится на плечи: короткое – р, 1 -я хромосома человека длинное – q. Плечи делятся на районы (регионы), районы сегменты. Нумерация идет от центромеры к теломерам последовательности : [номер хромосомы]- [плечо] [номер района]-[номер-сегмента]. Если при окраш вании сегментов выделяются дополнительные се менты их обозначают цифрами через точку. Например: 5 p 15. 2. - 5 -я хромосома коротко плечо 1 район 5 сегмент 2 подсегмент. Дополнительное деление субсегментов на другие сегменты например субсегмента 2 в 5 p 15. 2, соответственно обозначается как 5 p 15. 21; 5 р15. 22 и т. д. Сама центромера обозначается сочетанием цифр 1 и 0, т. е. часть центромеры в пределах короткого плеча обозначается как - р10, а часть, включающая длинное плечо -q 10. Полосы называются хромосомными маркерами.
Хромосома делится на плечи: короткое – р, 1 -я хромосома человека длинное – q. Плечи делятся на районы (регионы), районы сегменты. Нумерация идет от центромеры к теломерам последовательности : [номер хромосомы]- [плечо] [номер района]-[номер-сегмента]. Если при окраш вании сегментов выделяются дополнительные се менты их обозначают цифрами через точку. Например: 5 p 15. 2. - 5 -я хромосома коротко плечо 1 район 5 сегмент 2 подсегмент. Дополнительное деление субсегментов на другие сегменты например субсегмента 2 в 5 p 15. 2, соответственно обозначается как 5 p 15. 21; 5 р15. 22 и т. д. Сама центромера обозначается сочетанием цифр 1 и 0, т. е. часть центромеры в пределах короткого плеча обозначается как - р10, а часть, включающая длинное плечо -q 10. Полосы называются хромосомными маркерами.
 Схематическое изображение G-сегментов хромосом челове -ка и система их обозначения согласно решениям Парижской конференции
Схематическое изображение G-сегментов хромосом челове -ка и система их обозначения согласно решениям Парижской конференции
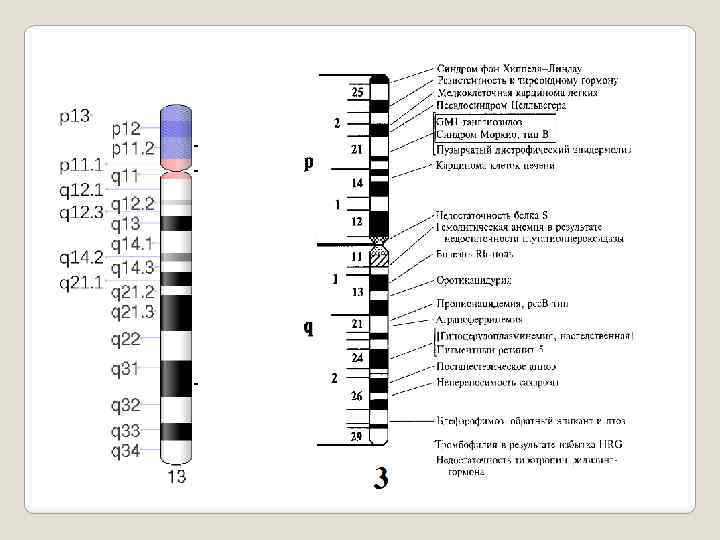

 Флуоресце нтная гибридиза ция in situ, или метод FISH — цитогенетический метод, который применяют для детекции и определения положения специфичес кой последовательности ДНК на метафазных хромосомах или в интерфазных яд рах in situ. Метод FISH используют в преимплантационной, пренатальной и по стнатальной генетической диагностике, в диагностике онкологических заболева ний, в ретроспективной биологической дозиметрии.
Флуоресце нтная гибридиза ция in situ, или метод FISH — цитогенетический метод, который применяют для детекции и определения положения специфичес кой последовательности ДНК на метафазных хромосомах или в интерфазных яд рах in situ. Метод FISH используют в преимплантационной, пренатальной и по стнатальной генетической диагностике, в диагностике онкологических заболева ний, в ретроспективной биологической дозиметрии.
 Генетический материал клетки Геномный уровень организации
Генетический материал клетки Геномный уровень организации
 между ними: спейсерные") Отделы ДНК Молекула ДНК состоит из генов и спейсерных участков (спейсерами) между ними: спейсерные участки ДНК, выполняющие структурную функцию участки ДНК, с которыми связы- ваются гистоновые белки нуклео- сомной цепи в хроматине, ДНКсвязывающиеся белки и др. , участ- вующие в правильной укладке нуклеосомной цепи в участки ДНК, высшие структуры участвующие в при- хроматина. креплении хромосом к аппарату центриолей; фиксации хроматина в ядерном матриксе (MAR- и SAR- спейсерные участки ДНК, которые служат специфи -ческими локусами связывания: спейсерные участки ДНК, служащие сигналами окончания транскрипции Аттенюаторы, терминаторы Промоторы Операторы Сайленсеры Энхансеры
Отделы ДНК Молекула ДНК состоит из генов и спейсерных участков (спейсерами) между ними: спейсерные участки ДНК, выполняющие структурную функцию участки ДНК, с которыми связы- ваются гистоновые белки нуклео- сомной цепи в хроматине, ДНКсвязывающиеся белки и др. , участ- вующие в правильной укладке нуклеосомной цепи в участки ДНК, высшие структуры участвующие в при- хроматина. креплении хромосом к аппарату центриолей; фиксации хроматина в ядерном матриксе (MAR- и SAR- спейсерные участки ДНК, которые служат специфи -ческими локусами связывания: спейсерные участки ДНК, служащие сигналами окончания транскрипции Аттенюаторы, терминаторы Промоторы Операторы Сайленсеры Энхансеры
 Геном – это количество ДНК в гаплоидном наборе хромосом; полный состав ДНК, содержа щий весь объем информации, необходимой для развития и существования организма; совокупность всех генов и меж генных участков ядерной и внеядерной ДНК клетки. Общие принципы построения генома и его структурно-функциональную организацию изучает раздел генетики – геномика. Различают следующие разделы геномики: структурная, функциональная, сравнительная, эволюционная и медицинская.
Геном – это количество ДНК в гаплоидном наборе хромосом; полный состав ДНК, содержа щий весь объем информации, необходимой для развития и существования организма; совокупность всех генов и меж генных участков ядерной и внеядерной ДНК клетки. Общие принципы построения генома и его структурно-функциональную организацию изучает раздел генетики – геномика. Различают следующие разделы геномики: структурная, функциональная, сравнительная, эволюционная и медицинская.
 Варианты геномов ГЕНОМ ПЛАЗМИДНЫЙ ПЛАСТИДНЫЙ . МИТОХОНДРИАЛЬНЫЙ ЯДЕРНЫЙ
Варианты геномов ГЕНОМ ПЛАЗМИДНЫЙ ПЛАСТИДНЫЙ . МИТОХОНДРИАЛЬНЫЙ ЯДЕРНЫЙ
 или РНК (РНКвирусы) и состоят из одной или") Геном вируса представлен молекулой ДНК (ДНКвирусы) или РНК (РНКвирусы) и состоят из одной или нескольких молекул нук -леиновых кислот. Вирусная частица, состоящая из одной или нескольких нук леиновых молекул называет- ся вирион. Он может быть представлен одно- и двуцепочными ДНК или РНК, кольцевой или линейной формы. Геном вирусов Вирион заключен в белковую оболочку – капсид, которая может содержать липиды и угле -воды. Вирусный геном относитель мал; кодирует белки капсида белки, участвующие в реплик - ции и экспрессии вирусного генома. Масса ДНК вирусов составляе 1∙ 106 - 200∙ 106; РНК - 1∙ 106 15∙ 106 дальтона.
Геном вируса представлен молекулой ДНК (ДНКвирусы) или РНК (РНКвирусы) и состоят из одной или нескольких молекул нук -леиновых кислот. Вирусная частица, состоящая из одной или нескольких нук леиновых молекул называет- ся вирион. Он может быть представлен одно- и двуцепочными ДНК или РНК, кольцевой или линейной формы. Геном вирусов Вирион заключен в белковую оболочку – капсид, которая может содержать липиды и угле -воды. Вирусный геном относитель мал; кодирует белки капсида белки, участвующие в реплик - ции и экспрессии вирусного генома. Масса ДНК вирусов составляе 1∙ 106 - 200∙ 106; РНК - 1∙ 106 15∙ 106 дальтона.
 Структура генома некоторых вирусов
Структура генома некоторых вирусов
 до 250 (вирус оспы). Структура") Геномы вирусов могут содержать от 4 генов (фаг Qβ) до 250 (вирус оспы). Структура генома фага φХ 174 Геномы могут быть цельными (состоять из одной молекулы нуклеиновой кислоты), либо сегментированными и состоять из нескольких молекул. Геном фага φХ 174 состоит из 9 генов и содержит 5500 нуклеотидов. В геноме фага имеются пере -крывающиеся гены (гены, которые могут быть частью двух разных генов или некоторая последо -вательность ДНК может быть составной частью двух разных генов) и «гены внутри генов» . Установлено, что ген В расположен внутри гена А, ген Е – вну -три гена Д. Подобная картина перекрывания генов было зафиксировано для фага λ и опухолевого ви -руса SV 40 и объясняла существование несоответ ствия между небольшим размером генома и коли -чеством белков, которые синтезируется генами это -го генома.
Геномы вирусов могут содержать от 4 генов (фаг Qβ) до 250 (вирус оспы). Структура генома фага φХ 174 Геномы могут быть цельными (состоять из одной молекулы нуклеиновой кислоты), либо сегментированными и состоять из нескольких молекул. Геном фага φХ 174 состоит из 9 генов и содержит 5500 нуклеотидов. В геноме фага имеются пере -крывающиеся гены (гены, которые могут быть частью двух разных генов или некоторая последо -вательность ДНК может быть составной частью двух разных генов) и «гены внутри генов» . Установлено, что ген В расположен внутри гена А, ген Е – вну -три гена Д. Подобная картина перекрывания генов было зафиксировано для фага λ и опухолевого ви -руса SV 40 и объясняла существование несоответ ствия между небольшим размером генома и коли -чеством белков, которые синтезируется генами это -го генома.
 имеет сферическую форму, ее диаметр") Геном вируса иммунодефицита человека ВИЧ-1. Вирусная частица (вирион) имеет сферическую форму, ее диаметр равен 100 нм Геном вируса составляет молекула РНК, она окружена двойным слоем липидов, пронизанных гликопротеина- ми, содержащими две субъединицы: gp 41, пронизыва- ющая мембрану и gp 120, находящаяся снаружи. Сердцевина вируса образована белками p 17 и p 24. В сердцевине вируса находится молекулы обратной тран- скриптазы, которая превращает РНК вируса в линейную молекулу двуцепочной молекулы ДНК в процессе обра- тной транскрипции в клетках Т-лимфоцитов тела хозя -ина. Образовавшаяся молекула ДНК вируса ВИЧ-1 прони- кает в ядро Т-лимфоцита, на концах линейной молеку- лы ДНК находятся особые последовательности И 3, R и U 5. Эти последовательности взаимодействуют друг с другом, образуя кольцевую ДНК с одним или двумя ДКП. Затем происходит встраивание вирусной ДНК в любое место клеточной ДНК хозяина, при участии бел -ка вируса интегразы. Геном вируса ВИЧ-1 содержит 9 генов, которые синтезируют 14 белковых продуктов. Ген gag кодирует
Геном вируса иммунодефицита человека ВИЧ-1. Вирусная частица (вирион) имеет сферическую форму, ее диаметр равен 100 нм Геном вируса составляет молекула РНК, она окружена двойным слоем липидов, пронизанных гликопротеина- ми, содержащими две субъединицы: gp 41, пронизыва- ющая мембрану и gp 120, находящаяся снаружи. Сердцевина вируса образована белками p 17 и p 24. В сердцевине вируса находится молекулы обратной тран- скриптазы, которая превращает РНК вируса в линейную молекулу двуцепочной молекулы ДНК в процессе обра- тной транскрипции в клетках Т-лимфоцитов тела хозя -ина. Образовавшаяся молекула ДНК вируса ВИЧ-1 прони- кает в ядро Т-лимфоцита, на концах линейной молеку- лы ДНК находятся особые последовательности И 3, R и U 5. Эти последовательности взаимодействуют друг с другом, образуя кольцевую ДНК с одним или двумя ДКП. Затем происходит встраивание вирусной ДНК в любое место клеточной ДНК хозяина, при участии бел -ка вируса интегразы. Геном вируса ВИЧ-1 содержит 9 генов, которые синтезируют 14 белковых продуктов. Ген gag кодирует
 Геном прокариот построен компактно и не отделен от цитоплазмы мембраной Размер генома у прокариот различается: он может варьировать от 580 тыс. н. п. до 9500 тыс. н. п. Геном бактерий почти целиком состоит из генов и последовательностей, регулирующих их активность. Геном бактерий обладает некоторыми осо- бенностями, обеспечивающими адапта - цию к определенным условиям их обита -ния. Это суперспирализация ДНК, обога - щенность генома ГЦ–парами, а также наличие генов, кодирующих белки, обеспе- чивающие приспособление микро Геном прокариот построен компактно и не отделен от цитоплазмы мембраной Молекула ДНК может быть кольцевой или линейной, а у некоторые бактерий имеются обе формы ДНК Гены прокариот не имеют интронов. Функционально связанные гены организованы в опероны Helicobacter pylori обитают в кислой среде желудка и их белки содержат вдвое больше аминокислотных остатков аргинина и лизи -на, чем у других бактерий. Кроме того, имеет- ся не менее пяти систем адгезии для прикре -пления этих бактерий к эпителию желудка, 14 генов системы
Геном прокариот построен компактно и не отделен от цитоплазмы мембраной Размер генома у прокариот различается: он может варьировать от 580 тыс. н. п. до 9500 тыс. н. п. Геном бактерий почти целиком состоит из генов и последовательностей, регулирующих их активность. Геном бактерий обладает некоторыми осо- бенностями, обеспечивающими адапта - цию к определенным условиям их обита -ния. Это суперспирализация ДНК, обога - щенность генома ГЦ–парами, а также наличие генов, кодирующих белки, обеспе- чивающие приспособление микро Геном прокариот построен компактно и не отделен от цитоплазмы мембраной Молекула ДНК может быть кольцевой или линейной, а у некоторые бактерий имеются обе формы ДНК Гены прокариот не имеют интронов. Функционально связанные гены организованы в опероны Helicobacter pylori обитают в кислой среде желудка и их белки содержат вдвое больше аминокислотных остатков аргинина и лизи -на, чем у других бактерий. Кроме того, имеет- ся не менее пяти систем адгезии для прикре -пления этих бактерий к эпителию желудка, 14 генов системы
 Количество генов 2.") Размеры геномов прокариот Организмы АРХЕИ Archaeoglodus fuigidis Размер генома (мегабазы) Количество генов 2. 17 2493 Methanococcus jannaschii 1. 66 1813 Thermoplasma acidophilum 1. 56 1509 ЭУБАКТЕРИИ 4. 64 4397 Bacillus sudtilis 4. 21 4212 Haemophilus influenzae Aquifex aeolicus 1. 83 1. 55 1791 1552 Pikettsia prowazekii 1. 11 834 Mycoplasma pheumoniae 0. 82 710 Mycoplasma genitalium 0. 58 503 Echerichia coli
Размеры геномов прокариот Организмы АРХЕИ Archaeoglodus fuigidis Размер генома (мегабазы) Количество генов 2. 17 2493 Methanococcus jannaschii 1. 66 1813 Thermoplasma acidophilum 1. 56 1509 ЭУБАКТЕРИИ 4. 64 4397 Bacillus sudtilis 4. 21 4212 Haemophilus influenzae Aquifex aeolicus 1. 83 1. 55 1791 1552 Pikettsia prowazekii 1. 11 834 Mycoplasma pheumoniae 0. 82 710 Mycoplasma genitalium 0. 58 503 Echerichia coli
 Геном бактерий практически не имеет перекрывающихся генов У бактерии около 1% генома является неко -дирующей и она представлена мобильными генетическими элементами, которые могут перемещаться по геному (см. геном эукариот). Мобильные (подвижные) гене – тические элементы бактерий – это IS-эле менты и транспозоны. Для бактерий характерно явление горизонтального переноса генов (перенос генов из генома одной бактерии в геном другой). Он происходит как между родственными, так и не родственными бактериями. Гены, которые появляется в результате такого перено- са называется ксенологами. Отмечен горизонтальный перенос генов от бактерий к эукариотом (около 30 генов). IS-элементы (инсерционные элементы или вставки) - это участки ДНК, способные переме- щается по геному как целое. Они содержат гены, обеспечивающие их перемещение – транспозицию При встраивании IS-элемента происходит: 1 - дупликация участков ДНК, находя- щихся на их концах; 2 - инверсия в смежных с IS -элемента- ми участках; 3 - взаимодействие между F-плазми - дой и бактериальной хромосомой. Перемещается не сам IS-элемент, а его копия, которая встраивается в новый локус
Геном бактерий практически не имеет перекрывающихся генов У бактерии около 1% генома является неко -дирующей и она представлена мобильными генетическими элементами, которые могут перемещаться по геному (см. геном эукариот). Мобильные (подвижные) гене – тические элементы бактерий – это IS-эле менты и транспозоны. Для бактерий характерно явление горизонтального переноса генов (перенос генов из генома одной бактерии в геном другой). Он происходит как между родственными, так и не родственными бактериями. Гены, которые появляется в результате такого перено- са называется ксенологами. Отмечен горизонтальный перенос генов от бактерий к эукариотом (около 30 генов). IS-элементы (инсерционные элементы или вставки) - это участки ДНК, способные переме- щается по геному как целое. Они содержат гены, обеспечивающие их перемещение – транспозицию При встраивании IS-элемента происходит: 1 - дупликация участков ДНК, находя- щихся на их концах; 2 - инверсия в смежных с IS -элемента- ми участках; 3 - взаимодействие между F-плазми - дой и бактериальной хромосомой. Перемещается не сам IS-элемент, а его копия, которая встраивается в новый локус
 – это такие же, как IS-элемен- ты, участки ДНК, но") Геном бактерий Транспозоны (Тn-элементы) – это такие же, как IS-элемен- ты, участки ДНК, но содержат гены, не имеющие отноше ния к транспозиции. Например, гены устойчивости к антибиотикам, гены токсинов и др. Структура транспозона: а- простой транспозон Тn 3 с тремя генами; б - сложный транспозон Тn 10 с двумя ISэлементами на левом и правом плече. Транспозоны : v придают бактериям устойчивость к различным антибиотикам, ядам, тяжелым металлам; v приводят к инактивации гена, в который они встраиваются; v повышают частоту мутаций, v активизируют ранее молчащий ген, v переносят гены метаболизма, которые позволяют использовать необычные источники углерода,
Геном бактерий Транспозоны (Тn-элементы) – это такие же, как IS-элемен- ты, участки ДНК, но содержат гены, не имеющие отноше ния к транспозиции. Например, гены устойчивости к антибиотикам, гены токсинов и др. Структура транспозона: а- простой транспозон Тn 3 с тремя генами; б - сложный транспозон Тn 10 с двумя ISэлементами на левом и правом плече. Транспозоны : v придают бактериям устойчивость к различным антибиотикам, ядам, тяжелым металлам; v приводят к инактивации гена, в который они встраиваются; v повышают частоту мутаций, v активизируют ранее молчащий ген, v переносят гены метаболизма, которые позволяют использовать необычные источники углерода,
 Геном бактерий Плазмиды – это структуры бак териальной клетки, способные реплицироваться самостоятельно от хромосом ДНК. Как правило, плазмиды встреча ются у бактерий и представляют собой двухцепочечные кольцевые молекулы, но изредка плазмиды встречаются также у эукариот. Эти внехромосомные элементы являются кольцевыми двуцепочными ДНК, несущими гены, которые будут необходимы в определенный момент жизни клетки.
Геном бактерий Плазмиды – это структуры бак териальной клетки, способные реплицироваться самостоятельно от хромосом ДНК. Как правило, плазмиды встреча ются у бактерий и представляют собой двухцепочечные кольцевые молекулы, но изредка плазмиды встречаются также у эукариот. Эти внехромосомные элементы являются кольцевыми двуцепочными ДНК, несущими гены, которые будут необходимы в определенный момент жизни клетки.
 Попадание плазмиды в клетку может осуществляться двумя путями: Трансмиссивные плазмиды Нетрансмиссивные плазмиды при непосредственном контак те клетки хозяина с другой клеткой в процессе конъюга ции вводятся в клетку с другой трансмиссивной плазмидой путём трансдукции плазмиды
Попадание плазмиды в клетку может осуществляться двумя путями: Трансмиссивные плазмиды Нетрансмиссивные плазмиды при непосредственном контак те клетки хозяина с другой клеткой в процессе конъюга ции вводятся в клетку с другой трансмиссивной плазмидой путём трансдукции плазмиды
 Размер плазмид варьирует от 1 до свыше 1000 тысяч пар оснований. Количество идентичных плазмид в пределах одной клетки изменяется от одной до тысяч в зависимости от дополнительных обстоятельств. Плазмиды можно считать видом мобильных генетических элементов, поскольку они часто передаются при конъюгации — механизме гори зонтального переноса генов.
Размер плазмид варьирует от 1 до свыше 1000 тысяч пар оснований. Количество идентичных плазмид в пределах одной клетки изменяется от одной до тысяч в зависимости от дополнительных обстоятельств. Плазмиды можно считать видом мобильных генетических элементов, поскольку они часто передаются при конъюгации — механизме гори зонтального переноса генов.
 R-плазмида, или R-фактор – фактор резистентности. Представляет собой кольцевую двуспиральную молекулу ДНК. В ней заключены гены, синтезирующие ферменты, расщепляю щие антибиотики; гены, тормозящие перенос антибиотиков через мембрану; гены, контролирующие перенос свойств резистентности в клетку реципиент; гены, определяющие устойчивость к конкретному антибиотику. R фактор передаётся при трансдукции и обычном делении клетки, при конъюгации бактерий. С действием R плазмид часто бывает связан тот факт, что некоторые бактериальные заболевания с трудом поддаются лечению при помощи известных на данный момент антибиотиков.
R-плазмида, или R-фактор – фактор резистентности. Представляет собой кольцевую двуспиральную молекулу ДНК. В ней заключены гены, синтезирующие ферменты, расщепляю щие антибиотики; гены, тормозящие перенос антибиотиков через мембрану; гены, контролирующие перенос свойств резистентности в клетку реципиент; гены, определяющие устойчивость к конкретному антибиотику. R фактор передаётся при трансдукции и обычном делении клетки, при конъюгации бактерий. С действием R плазмид часто бывает связан тот факт, что некоторые бактериальные заболевания с трудом поддаются лечению при помощи известных на данный момент антибиотиков.
 определяют синтез особых белков —") Col-плазмиды Col плазмиды (от англ. colicinogeny — колициноген ность) определяют синтез особых белков — колицинов, подавляющий рост и размножение других бактерий, но безвредных для бактерий, их выделяющих. Этот феномен был открыт в 1925 г. A. Gratia у штамма E. coli. Установлено, что E. coli синтезируют колицины разли чных типов: A, B, C, D и т. д. Колицины адсорбируются на клетках, лишённых Col фактора, и, не проникая внутрь, нарушают их метаболизм, вызывая гибель клетки.
Col-плазмиды Col плазмиды (от англ. colicinogeny — колициноген ность) определяют синтез особых белков — колицинов, подавляющий рост и размножение других бактерий, но безвредных для бактерий, их выделяющих. Этот феномен был открыт в 1925 г. A. Gratia у штамма E. coli. Установлено, что E. coli синтезируют колицины разли чных типов: A, B, C, D и т. д. Колицины адсорбируются на клетках, лишённых Col фактора, и, не проникая внутрь, нарушают их метаболизм, вызывая гибель клетки.
 F-плазмида Наиболее известной конъюгативной плазмидой является F плазмида, или F фактор. F плазмида является эписомой длиной около 100 тыс. пар оснований. F плазмида, как и все конъюгативные плазми ды, кодирует белки, противодействующие прикреп лению пилей (ворсинок бактерий) других бактерий к клеточной стенке данной. Контролирует синтез секс пилей (половые волоски); конъюгацию и перенос генов в клетку реципиент.
F-плазмида Наиболее известной конъюгативной плазмидой является F плазмида, или F фактор. F плазмида является эписомой длиной около 100 тыс. пар оснований. F плазмида, как и все конъюгативные плазми ды, кодирует белки, противодействующие прикреп лению пилей (ворсинок бактерий) других бактерий к клеточной стенке данной. Контролирует синтез секс пилей (половые волоски); конъюгацию и перенос генов в клетку реципиент.
 Эписомы – это плазмиды, которые могут существовать и реплицироваться как автономно, так и в составе хромосомной ДНК Встраиваясь в неё, становятся её частью и реплицируются вместе. Эписомы, перемещаясь из одной клетки в другую, могут придавать клеткам бактерий хозяев: v инфекционность или устойчивость к антибиотикам; v определяют болезнетворность бактерий; v обеспечивают почвенным бактерия способность усваивать углерод из необычных источников (нефте- продукты); v способность синтезировать бактериоцин, гемолецин и др.
Эписомы – это плазмиды, которые могут существовать и реплицироваться как автономно, так и в составе хромосомной ДНК Встраиваясь в неё, становятся её частью и реплицируются вместе. Эписомы, перемещаясь из одной клетки в другую, могут придавать клеткам бактерий хозяев: v инфекционность или устойчивость к антибиотикам; v определяют болезнетворность бактерий; v обеспечивают почвенным бактерия способность усваивать углерод из необычных источников (нефте- продукты); v способность синтезировать бактериоцин, гемолецин и др.
 Функции плазмид : ü перенос генетического материала при конъюгации — F-плазмида; ü плазмиды бактериоциногенности контролируют синтез белков, летальных для других бактерий — Col-плазмиды; ü синтез гемолизинов — Hly-плазмиды (являются конъюгативными); ü устойчивость к тяжёлым металлам; ü устойчивость к антибиотикам (R-плазмиды); ü синтез энтеротоксинов — Ent-плазмиды; ü устойчивость к УФ-излучению; ü синтез антигенов, обеспечивающих адгезию бактерий на клетках в организме человека и животных — плазмиды антигенов колонизации; ü система рестрикции-модификации; ü расщепление камфоры (плазмида САМ), ксилола (плазмида XYL), салицилата (плазмида SAL)
Функции плазмид : ü перенос генетического материала при конъюгации — F-плазмида; ü плазмиды бактериоциногенности контролируют синтез белков, летальных для других бактерий — Col-плазмиды; ü синтез гемолизинов — Hly-плазмиды (являются конъюгативными); ü устойчивость к тяжёлым металлам; ü устойчивость к антибиотикам (R-плазмиды); ü синтез энтеротоксинов — Ent-плазмиды; ü устойчивость к УФ-излучению; ü синтез антигенов, обеспечивающих адгезию бактерий на клетках в организме человека и животных — плазмиды антигенов колонизации; ü система рестрикции-модификации; ü расщепление камфоры (плазмида САМ), ксилола (плазмида XYL), салицилата (плазмида SAL)
 Геном прокариот Структура типичной R-плазм На рисунке изображена типичная плазмида. 50% генов отвечают за конъюгацию и репликацию. Вторая половина генов, контролирует устойчи -вость к тетрациклину, антибиотикам (гены tetr, camr, ranr и др. ). Перемещение плазмид обеспе – чивают и Тn-элементы , которые способны перемещаться не только между плазмидами, но и по всему геному
Геном прокариот Структура типичной R-плазм На рисунке изображена типичная плазмида. 50% генов отвечают за конъюгацию и репликацию. Вторая половина генов, контролирует устойчи -вость к тетрациклину, антибиотикам (гены tetr, camr, ranr и др. ). Перемещение плазмид обеспе – чивают и Тn-элементы , которые способны перемещаться не только между плазмидами, но и по всему геному
 м эукариот Количество ДНК в геноме клетки обозначается латинской буквой С, измеряется в пико - граммах (пг) и выражается в па- рах нуклеотидов (п. н. ), дальто - нах (Да) или в мегабазах (мб). 1 пг = расстоянию 0, 965∙ 109 нп = 6, 1∙ 1011 Да. 1 мб = 1 млн. оснований. Геном эукариот представлен: 1 - линейными молекулами ДНК, которые распределены по хромосомам и расположенны в ядре (ядерная ДНК); 2 - кольцевыми молекулами ДНК митохондрий и пластид (внехромосом ная ДНК). Геном эукариот – это суммарная ДНК гаплоид- - ного набора хромосом и внехромосомных элементов в клетках организма.
м эукариот Количество ДНК в геноме клетки обозначается латинской буквой С, измеряется в пико - граммах (пг) и выражается в па- рах нуклеотидов (п. н. ), дальто - нах (Да) или в мегабазах (мб). 1 пг = расстоянию 0, 965∙ 109 нп = 6, 1∙ 1011 Да. 1 мб = 1 млн. оснований. Геном эукариот представлен: 1 - линейными молекулами ДНК, которые распределены по хромосомам и расположенны в ядре (ядерная ДНК); 2 - кольцевыми молекулами ДНК митохондрий и пластид (внехромосом ная ДНК). Геном эукариот – это суммарная ДНК гаплоид- - ного набора хромосом и внехромосомных элементов в клетках организма.
 Геном эукариот характеризуется сложностью, плотностью генов, наличием интронов. Геном эукариот Сложность ДНК характеризуется общей длиной (в н. п. ) разных последовательнос- тей ДНК, определяющих ее избыточность. Размер генома человека составляет 3, 2∙ 109 н. п. По результатам выполнения проекта «Геном человека» (2001 г. ) 1 -2% генома составляют кодирующие экзоны, 24% - интроны и 75% - межгенные промежутки. Только 1 -2% генома несет информа цию о структуре белков. Следовательно, большая часть генома составляет избыточную, не кодирующую последовательность ДНК. Этот феномен получил название «парадокс С» .
Геном эукариот характеризуется сложностью, плотностью генов, наличием интронов. Геном эукариот Сложность ДНК характеризуется общей длиной (в н. п. ) разных последовательнос- тей ДНК, определяющих ее избыточность. Размер генома человека составляет 3, 2∙ 109 н. п. По результатам выполнения проекта «Геном человека» (2001 г. ) 1 -2% генома составляют кодирующие экзоны, 24% - интроны и 75% - межгенные промежутки. Только 1 -2% генома несет информа цию о структуре белков. Следовательно, большая часть генома составляет избыточную, не кодирующую последовательность ДНК. Этот феномен получил название «парадокс С» .
 Количество и плотность генов в 5 отсеквенированных эукариотических геномах Организм Количество отсеквенирован ных оснований (тнп) Дрожжи 12068 (S. cereviseae ) Круглый червь 97000 (C. elegans) Плодовая мушка 116000 (D. melanogas ter) Растение 115000 (A. thaliana) Человек 2693000 (H. sapiens) Степень законченност и, % 93 количество предсказанн ых генов 5885 Количеств о генов на 106 основан ий 483 99 19099 197 64 13601 117 92 84 25498 31780 221 12
Количество и плотность генов в 5 отсеквенированных эукариотических геномах Организм Количество отсеквенирован ных оснований (тнп) Дрожжи 12068 (S. cereviseae ) Круглый червь 97000 (C. elegans) Плодовая мушка 116000 (D. melanogas ter) Растение 115000 (A. thaliana) Человек 2693000 (H. sapiens) Степень законченност и, % 93 количество предсказанн ых генов 5885 Количеств о генов на 106 основан ий 483 99 19099 197 64 13601 117 92 84 25498 31780 221 12
 Последовательнос Различают следующие последовательности ДНК: уникальные и повторяющиеся. Уникальные последовательности представлены в ДНК в одном экземпляре; у человека они состав ляют 75% генома. К уникальным последовательнос тям относятся многие струк-турные гены. Общее число генов у эукариот по данным раз ных авторов различно. Уникальными последователь ностями являются экзоны и интроны генов и некоторые межгенные последовательно-сти.
Последовательнос Различают следующие последовательности ДНК: уникальные и повторяющиеся. Уникальные последовательности представлены в ДНК в одном экземпляре; у человека они состав ляют 75% генома. К уникальным последовательнос тям относятся многие струк-турные гены. Общее число генов у эукариот по данным раз ных авторов различно. Уникальными последователь ностями являются экзоны и интроны генов и некоторые межгенные последовательно-сти.
 Если последовательности ДНК присутствуют в геноме в виде нескольких копий, то их называют повторяющимися или повторами. Повторы делятся по распол нию в геном и по частоте речаемости. II По расположению в геноме повторы бывают: тандемные, диспергированные (то есть, разбросанные по геному), и обращенные. Основные классы повторов в геноме человека: А – диспергирован ные, Б тандемные, В – обращенные (стрелками обозначены повторы, прямыми линиями – основная часть ДНК)
Если последовательности ДНК присутствуют в геноме в виде нескольких копий, то их называют повторяющимися или повторами. Повторы делятся по распол нию в геном и по частоте речаемости. II По расположению в геноме повторы бывают: тандемные, диспергированные (то есть, разбросанные по геному), и обращенные. Основные классы повторов в геноме человека: А – диспергирован ные, Б тандемные, В – обращенные (стрелками обозначены повторы, прямыми линиями – основная часть ДНК)
 Обращенные повторы или палиндромы – это участки ДНК, которые читаются одинаково слева направо и нао борот. Палиндромы принимают участие в образовании шпилек при транскрипции ДНК; могут являтся местами связывания со специфическими белками, участвующими в регуляции репликации и транскрипции.
Обращенные повторы или палиндромы – это участки ДНК, которые читаются одинаково слева направо и нао борот. Палиндромы принимают участие в образовании шпилек при транскрипции ДНК; могут являтся местами связывания со специфическими белками, участвующими в регуляции репликации и транскрипции.
 Простые тандемные повторы (са При центрифугировании ДНК в растворе хло - ристого цезия происходит разделение ДНК по плотности на основную и сателиттную Сателлитными ДНК называются минорный комп - лекс ДНК, отделяющийся от основной ДНК при центрифугировании в растворе Основная масса ДНК имеет плотность равную 42% содержания ГЦ-пар нуклеотидов или 1, 693. Часть ДНК, имеющая иную плотность (а, следовательно, иное содержание нуклеотидных пар) получила название дополнительной, минорной или сателлитной ДНК (сат. ДНК) (от лат. satellitisспутник). Сателлитные ДНК состоят из коротких (чаще 1 -20 нуклеотидов) последовательностей (базовые после- довательности), идентичных или почти идентичных. Базовые последовательности расположены тан – демно (то есть, следуют друг за другом «голова к хвосту» ), образуя длинные блоки, протяженностью до десятков тысяч нуклеотидов. Например, у плодовой мушки дрозофилы имеются простые базовые после- довательности - ААТАГ-, -ААТАТ, - ААТАЦ-, которые повторяется «п» количество раз, где п = 10, 20 и более тысяч раз.
Простые тандемные повторы (са При центрифугировании ДНК в растворе хло - ристого цезия происходит разделение ДНК по плотности на основную и сателиттную Сателлитными ДНК называются минорный комп - лекс ДНК, отделяющийся от основной ДНК при центрифугировании в растворе Основная масса ДНК имеет плотность равную 42% содержания ГЦ-пар нуклеотидов или 1, 693. Часть ДНК, имеющая иную плотность (а, следовательно, иное содержание нуклеотидных пар) получила название дополнительной, минорной или сателлитной ДНК (сат. ДНК) (от лат. satellitisспутник). Сателлитные ДНК состоят из коротких (чаще 1 -20 нуклеотидов) последовательностей (базовые после- довательности), идентичных или почти идентичных. Базовые последовательности расположены тан – демно (то есть, следуют друг за другом «голова к хвосту» ), образуя длинные блоки, протяженностью до десятков тысяч нуклеотидов. Например, у плодовой мушки дрозофилы имеются простые базовые после- довательности - ААТАГ-, -ААТАТ, - ААТАЦ-, которые повторяется «п» количество раз, где п = 10, 20 и более тысяч раз.
 Типичная саттелитная Д. Сателлиты характеризуется следующими свойствами: 1. быстрая и точная реассоциация (восстановление структуры ДНК после дена турации ДНК) в процессе ренатурации ДНК; 2. тандемное расположение базовых последовательностей, объединенных в блоки 3. количество копии в геноме свыше 10 «степеней» ; 4. простая первичная структура; 5. гомогенность (идентичность) базовых последовательностей; 6. локализация в прицентромерном гетерохроматине и теломерах ; 7. ограниченная репликация;
Типичная саттелитная Д. Сателлиты характеризуется следующими свойствами: 1. быстрая и точная реассоциация (восстановление структуры ДНК после дена турации ДНК) в процессе ренатурации ДНК; 2. тандемное расположение базовых последовательностей, объединенных в блоки 3. количество копии в геноме свыше 10 «степеней» ; 4. простая первичная структура; 5. гомогенность (идентичность) базовых последовательностей; 6. локализация в прицентромерном гетерохроматине и теломерах ; 7. ограниченная репликация;
 У человека выделяют следующие виды сателлитных повторов: 1. классические сателлиты короткие прямые повторы. Выделяют три вида классических сателлитов: сателлит 1: базовая последовательность состоит из 42 н. п. и обнаружена в 3 4, 13 15, 21 хромосомах; сателлит 2: базовая последовательность равна 5 н. п. и обнаружен в хромосомах 1, 2, 10, 16; сателлит 3: базовая последовательность равна 5 или 10 н. п. и находится в 1, 5, 9 10, 13 15, 17, 21 22 хромосомах; 2. альфа-сателлитные повторы: базовая последовательность равна 171 н. п. ; обнаружена во всех хромосомах человека; имеют блоки длиной до 4 млн. пар нуклеотидов и расположены в прицентромерной, центромерной области; 3. бета-сателлитные повторы: базовая последовательность 68 н. п. ; находятся в 1, 3, 9, 13 15, 21 22 и У хромосомах; 4. гамма-сателлитные повторы: базовая последовательность имеет длину в 220 н. п. и находятся в хромосомах 8 и Х; 5. сателлитные повторы с базовая последовательностью в 48 н. п. в хромосомах 13 15, 21 22 и У; 6. Sn 5 -сателлитные повторы хромосом 2, 13 15, 020 22 с неустанов ленной базовой последовательностью
У человека выделяют следующие виды сателлитных повторов: 1. классические сателлиты короткие прямые повторы. Выделяют три вида классических сателлитов: сателлит 1: базовая последовательность состоит из 42 н. п. и обнаружена в 3 4, 13 15, 21 хромосомах; сателлит 2: базовая последовательность равна 5 н. п. и обнаружен в хромосомах 1, 2, 10, 16; сателлит 3: базовая последовательность равна 5 или 10 н. п. и находится в 1, 5, 9 10, 13 15, 17, 21 22 хромосомах; 2. альфа-сателлитные повторы: базовая последовательность равна 171 н. п. ; обнаружена во всех хромосомах человека; имеют блоки длиной до 4 млн. пар нуклеотидов и расположены в прицентромерной, центромерной области; 3. бета-сателлитные повторы: базовая последовательность 68 н. п. ; находятся в 1, 3, 9, 13 15, 21 22 и У хромосомах; 4. гамма-сателлитные повторы: базовая последовательность имеет длину в 220 н. п. и находятся в хромосомах 8 и Х; 5. сателлитные повторы с базовая последовательностью в 48 н. п. в хромосомах 13 15, 21 22 и У; 6. Sn 5 -сателлитные повторы хромосом 2, 13 15, 020 22 с неустанов ленной базовой последовательностью
 от") Нетипичные сателлиты Мини-сателлиты – это повторы со средней длиной повторяющегося элемента (базовой последовательности) от 6 до 100 н. п. , обога - щенным АТ- и ГЦ-парами, длиной кластера 0, 2 -20 тыс. н. п. Число их копий варьирует от несколький сотен до тысяч на геном. Мини-сателлитами являются также повторы, тандемно расположенные в теломерной области. Основной элемент этих повторов у человека – высококонсервативные последовательности ГГТАТТ. Эти последовательности и играют роль в сохранении целостности хромосом. Мегасателлиты – повторы с повторяющейся базовой последовательностью в 2, 5 -5 тыс. н. п. и числом копий от 50 до 400 присутст - вуют лишь в 4, 8, 19 и Х хромосомах.
Нетипичные сателлиты Мини-сателлиты – это повторы со средней длиной повторяющегося элемента (базовой последовательности) от 6 до 100 н. п. , обога - щенным АТ- и ГЦ-парами, длиной кластера 0, 2 -20 тыс. н. п. Число их копий варьирует от несколький сотен до тысяч на геном. Мини-сателлитами являются также повторы, тандемно расположенные в теломерной области. Основной элемент этих повторов у человека – высококонсервативные последовательности ГГТАТТ. Эти последовательности и играют роль в сохранении целостности хромосом. Мегасателлиты – повторы с повторяющейся базовой последовательностью в 2, 5 -5 тыс. н. п. и числом копий от 50 до 400 присутст - вуют лишь в 4, 8, 19 и Х хромосомах.
 Микросателлиты относятся к повторам с длиной повторяющегося элемента 1 -4 нуклеотида, тандемно и равномерно, расположенные по всему геному. Наиболее известный микросателлит человека - динуклеотид (АЦ)п, где n (число повторов) равен 5 -50. Кластер этого микросателлита расположен через каждые 30 тыс. н. п. Кроме динуклеотида АЦ - известны микросателлиты с повторяющимися три- и тетрануклеотидными мотивами АТ, ГЦ, ААЦ, ЦТГ, ГАЦА. Если микросателлит состоит только из одного повторяющего нуклеотида (АААААА…. . А - на одной нити ДНК и ТТТТТТ…. . Т - на другой), то он называется гомополимерной нуклеотидной последовательностью. Блоки микросателлитов равны 20 -60 тыс. н. п.
Микросателлиты относятся к повторам с длиной повторяющегося элемента 1 -4 нуклеотида, тандемно и равномерно, расположенные по всему геному. Наиболее известный микросателлит человека - динуклеотид (АЦ)п, где n (число повторов) равен 5 -50. Кластер этого микросателлита расположен через каждые 30 тыс. н. п. Кроме динуклеотида АЦ - известны микросателлиты с повторяющимися три- и тетрануклеотидными мотивами АТ, ГЦ, ААЦ, ЦТГ, ГАЦА. Если микросателлит состоит только из одного повторяющего нуклеотида (АААААА…. . А - на одной нити ДНК и ТТТТТТ…. . Т - на другой), то он называется гомополимерной нуклеотидной последовательностью. Блоки микросателлитов равны 20 -60 тыс. н. п.
 Микро- и минисателлиты очень изменчивы, то есть полиморфны. Изменяются как число нуклеотидов в базовой последовательности, так количество копий. Их распределение и число в геноме каждого конкрет ного человека настолько индивидуально, что лежит в основе метода судебно –медицинской экспертизы, служащего для идентификации личности называе мого ДНК-фингерпринтингом или методом «отпечатков пальцев» .
Микро- и минисателлиты очень изменчивы, то есть полиморфны. Изменяются как число нуклеотидов в базовой последовательности, так количество копий. Их распределение и число в геноме каждого конкрет ного человека настолько индивидуально, что лежит в основе метода судебно –медицинской экспертизы, служащего для идентификации личности называе мого ДНК-фингерпринтингом или методом «отпечатков пальцев» .
 по всему геному. Они встречаются") Диспергированные повторы – это повто -ры, которые рассеяны (диспергированы) по всему геному. Они встречаются во всех областях хромосом; обнаруживают -ся внутри генов в интронах, по краям генов, в межгенных участках, вне генной ДНК и составляют половину генома человека. Эти типы повторов объединены в группу мобильных (подвижных) элементов геномов, Они способны определять структуру ге -нома и перемещаться в нем как внутри одной хромосомы, так и между хромо- сомами. Процесс перемещения назы -вается транспозиция Диспергированные повторы Различают следующие типы таких повторов: 1. длинные диспергированные повторы или Line-повторы. Их длина составляет 6, 4 тыс н. п. , число копий 8, 5 х 105 раз (21% всего генома). 2. короткие диспергированные повторы или Sine –повторы, с длиной менее 500 н. п. и числом копий 1, 5 х 106 (13% генома); 3. повторы протяженность 5 -8 тыс. н. п. , ограниченные с флангов дополнительными повторами, которые называются LТR (long terminal repeats) или ДКП (длинные концевые повторы). 4. ДНК- транспозоны – повторы длиной от 80 -3000 нп. числом копий 3 х 105 Перемещение осуществляется двумя путями: (3% генома). 1 - путем вырезания элемента из одного места и встраивания его в другое при участии фермента транспозазы и вспомогательных белков; 2 - путем образования копии элемента, которая внедряется в новое место, а родительская копия остается на месте. Подвижные генетические элементы образую- щиеся первым путем называется
Диспергированные повторы – это повто -ры, которые рассеяны (диспергированы) по всему геному. Они встречаются во всех областях хромосом; обнаруживают -ся внутри генов в интронах, по краям генов, в межгенных участках, вне генной ДНК и составляют половину генома человека. Эти типы повторов объединены в группу мобильных (подвижных) элементов геномов, Они способны определять структуру ге -нома и перемещаться в нем как внутри одной хромосомы, так и между хромо- сомами. Процесс перемещения назы -вается транспозиция Диспергированные повторы Различают следующие типы таких повторов: 1. длинные диспергированные повторы или Line-повторы. Их длина составляет 6, 4 тыс н. п. , число копий 8, 5 х 105 раз (21% всего генома). 2. короткие диспергированные повторы или Sine –повторы, с длиной менее 500 н. п. и числом копий 1, 5 х 106 (13% генома); 3. повторы протяженность 5 -8 тыс. н. п. , ограниченные с флангов дополнительными повторами, которые называются LТR (long terminal repeats) или ДКП (длинные концевые повторы). 4. ДНК- транспозоны – повторы длиной от 80 -3000 нп. числом копий 3 х 105 Перемещение осуществляется двумя путями: (3% генома). 1 - путем вырезания элемента из одного места и встраивания его в другое при участии фермента транспозазы и вспомогательных белков; 2 - путем образования копии элемента, которая внедряется в новое место, а родительская копия остается на месте. Подвижные генетические элементы образую- щиеся первым путем называется
 Подвижные генетические элементы, встраиваясь в гены или в соседние с ними области: 1 - изменяют активность этих генов; 2 - вызывают мутации; 3 - могут принимать участие в перестройке хромосом, приводящей в результате к рекомбинации, инвер сии, транслокации, делеции и т. д. ; 4 - могут обусловливать поли -морфизмы внутри вида; 5 - обеспечивают горизонтальны перенос генов; 6 - способствуют сохранению структурной целостности хромосом. Подвижные генетические элементы
Подвижные генетические элементы, встраиваясь в гены или в соседние с ними области: 1 - изменяют активность этих генов; 2 - вызывают мутации; 3 - могут принимать участие в перестройке хромосом, приводящей в результате к рекомбинации, инвер сии, транслокации, делеции и т. д. ; 4 - могут обусловливать поли -морфизмы внутри вида; 5 - обеспечивают горизонтальны перенос генов; 6 - способствуют сохранению структурной целостности хромосом. Подвижные генетические элементы
 Транспозоны - это подвижные генетические элементы, ограниченные по флангам инвер тированными (направленными друг к другу) повторами и перемещающиеся по геному при участи фермента транслоказы, который коди- уется самим транспозоном. Более 50 генов человека произошли из транспо зонов(имму- ноглобуллины). Транспозоны входят во все типы диспергированных повторов. Структура транспозона: а- простой транспозон Тn 3 с тремя генами; б - сложный транспозон Тn 10 с двумя ISэлементами на левом и правом плече. ДНК-транспозоны способны кодировать фермент транспозазу, который обеспечи -вает их передвижение по геному. Наиболее изученные ДНКтранспозоны – это Р-элемент, Асэлемент, Ту-элемент. Транспозоны могут выполнять: 1 - роль регуляторов генов, являясь энхансерами или сайленсерами; 2 - функцию терминаторов транскрип- ции; 3 - участвуют в процессе репликации ДНК, упаковке хромосом, фиксации хро - мосом к ядерному матриксу.
Транспозоны - это подвижные генетические элементы, ограниченные по флангам инвер тированными (направленными друг к другу) повторами и перемещающиеся по геному при участи фермента транслоказы, который коди- уется самим транспозоном. Более 50 генов человека произошли из транспо зонов(имму- ноглобуллины). Транспозоны входят во все типы диспергированных повторов. Структура транспозона: а- простой транспозон Тn 3 с тремя генами; б - сложный транспозон Тn 10 с двумя ISэлементами на левом и правом плече. ДНК-транспозоны способны кодировать фермент транспозазу, который обеспечи -вает их передвижение по геному. Наиболее изученные ДНКтранспозоны – это Р-элемент, Асэлемент, Ту-элемент. Транспозоны могут выполнять: 1 - роль регуляторов генов, являясь энхансерами или сайленсерами; 2 - функцию терминаторов транскрип- ции; 3 - участвуют в процессе репликации ДНК, упаковке хромосом, фиксации хро - мосом к ядерному матриксу.
 Ретротранспозоны – это подвижные генетические элементы, которые перемещаются по геному, используя механизм обратной транскрипции. Ретротранспозоны Структура ретротранспозона
Ретротранспозоны – это подвижные генетические элементы, которые перемещаются по геному, используя механизм обратной транскрипции. Ретротранспозоны Структура ретротранспозона
") Ретротранспозоны SINE-повторы или короткие повторы име по краям короткие прямые повторы и поли ) последовательность на 3′ конце. Самый изве ный из изученных SINE повторов у человека – о семейство Аlu- повторов. Повторы этого семейства имеют длину 2000 н. п. , находятся как внутри гена, так между ми. Количество копий равно 1 млн и состав ют 5% от всей ДНК. Они обнаруживается в геноме повсеместно: в тронах, в сателлитной ДНК, между генами. пируя самих себя Аlu повторы перемещаются геному, внедряясь в экзон или интрон и: меняют регуляцию экспрессии этого гена. , способны модулировать репликацию ДНК, сплайсинг и некоторые другие процессы. участвуют в клеточном ответе на стрессовые воздействия, подавляя трансляцию РНК. LTR – последовательности (300 -400 н. п. ) принимают участие в процессе «тасова -ния экзонов» , то есть, способны переме- щать экзоны из одних генов в другие, обеспечивая образование новых генов. LINЕ-повторы также относятся к ретротран- спозонам, не имеющим LТR -последователь -ности. У человека имеется только одно семейство этих повторов – L 1 -повторы. Они кодируют два белка-фермента, которые и обеспечивают их перемещение в геноме по механизму обратной транскрипции, характерному для ретротранспозонов. В процессе обратной транскрипции L 1 -копий могут возникать псевдогены (нефункциональные гены с прямым повторами по краям длиной 10 -20 н. п. , гомологичные известным структурным геном) и безинтронные нормально функциониорующие гены.
Ретротранспозоны SINE-повторы или короткие повторы име по краям короткие прямые повторы и поли ) последовательность на 3′ конце. Самый изве ный из изученных SINE повторов у человека – о семейство Аlu- повторов. Повторы этого семейства имеют длину 2000 н. п. , находятся как внутри гена, так между ми. Количество копий равно 1 млн и состав ют 5% от всей ДНК. Они обнаруживается в геноме повсеместно: в тронах, в сателлитной ДНК, между генами. пируя самих себя Аlu повторы перемещаются геному, внедряясь в экзон или интрон и: меняют регуляцию экспрессии этого гена. , способны модулировать репликацию ДНК, сплайсинг и некоторые другие процессы. участвуют в клеточном ответе на стрессовые воздействия, подавляя трансляцию РНК. LTR – последовательности (300 -400 н. п. ) принимают участие в процессе «тасова -ния экзонов» , то есть, способны переме- щать экзоны из одних генов в другие, обеспечивая образование новых генов. LINЕ-повторы также относятся к ретротран- спозонам, не имеющим LТR -последователь -ности. У человека имеется только одно семейство этих повторов – L 1 -повторы. Они кодируют два белка-фермента, которые и обеспечивают их перемещение в геноме по механизму обратной транскрипции, характерному для ретротранспозонов. В процессе обратной транскрипции L 1 -копий могут возникать псевдогены (нефункциональные гены с прямым повторами по краям длиной 10 -20 н. п. , гомологичные известным структурным геном) и безинтронные нормально функциониорующие гены.
 ДНК-повторы генома человека
ДНК-повторы генома человека
 15% генома человека. К высокоповторяющимся последовательностям относится сателлитная ДНК; частота встречаемости таких повторов равна 105 копий и выше. Они функционально и структурно обособлены от всего генома. По частоте встречаемости в геноме все повторы делятся на: ü высокоповторяющиеся или высокочастотные ü умеренноповторяющие ся или среднечастотные
15% генома человека. К высокоповторяющимся последовательностям относится сателлитная ДНК; частота встречаемости таких повторов равна 105 копий и выше. Они функционально и структурно обособлены от всего генома. По частоте встречаемости в геноме все повторы делятся на: ü высокоповторяющиеся или высокочастотные ü умеренноповторяющие ся или среднечастотные
 Умеренно повторяющиеся последовательности эти последовательности, встречаются в геноме в десятках или сотнях копий и составляют: 1 семейства генов, расположен ных тандемно : гены гистонов, т. РНК и р. РНК. Они обеспечивают осуществление жизненно важных общеклеточных функций и назы ваются гены «домашнего хозяйства» ; 2 гены, кодирующие гомологичные белки со сходными функциями, которые могут быть рассеяны по геному или обра зовывать кластеры. Это гены актина, ту булина, глобина, иммуногло булина, гены теплового шока и др. Они выполняют в клетке как общеклеточные, так и специал изированные функции. 4 тандемные повторы или диспергиро ванные последовательности с неизвест ными функциями. Один из примеров таких последовательностей ДНК последовате льность VNTR. Эти последовательности со держат повторяющейся элемент из 14 100 нуклеотидов, который может содержат от 2 до 100 повторов. Они могут находится вну три гена или между ними. 3 различные подвижные (мобильные) генетические элементы. Они составляют от 10 20% генома и разбро саны (диспергированы) по всему геному;
Умеренно повторяющиеся последовательности эти последовательности, встречаются в геноме в десятках или сотнях копий и составляют: 1 семейства генов, расположен ных тандемно : гены гистонов, т. РНК и р. РНК. Они обеспечивают осуществление жизненно важных общеклеточных функций и назы ваются гены «домашнего хозяйства» ; 2 гены, кодирующие гомологичные белки со сходными функциями, которые могут быть рассеяны по геному или обра зовывать кластеры. Это гены актина, ту булина, глобина, иммуногло булина, гены теплового шока и др. Они выполняют в клетке как общеклеточные, так и специал изированные функции. 4 тандемные повторы или диспергиро ванные последовательности с неизвест ными функциями. Один из примеров таких последовательностей ДНК последовате льность VNTR. Эти последовательности со держат повторяющейся элемент из 14 100 нуклеотидов, который может содержат от 2 до 100 повторов. Они могут находится вну три гена или между ними. 3 различные подвижные (мобильные) генетические элементы. Они составляют от 10 20% генома и разбро саны (диспергированы) по всему геному;
 Структурные классы ДНК человека
Структурные классы ДНК человека
, которая") Геном человека Генома человека был исследован международной программой «Геном человека» (Human Genome Project), которая начала работу в 1990 году. Первый отчет по проекту «Геном человека» был сделан в 2000 году, окончательный - в 2003 году.
Геном человека Генома человека был исследован международной программой «Геном человека» (Human Genome Project), которая начала работу в 1990 году. Первый отчет по проекту «Геном человека» был сделан в 2000 году, окончательный - в 2003 году.
 Геном человека Общее количество ДНК в 46 хромосомах в: соматической клетке человека имеет длину 2 м и 6, 4∙ 109 пар оснований; в гаплоидной – 3, 2∙ 109 пар оснований. Основное количество ДНК локализовано в хромо сомах (99, 5%), то есть, составляет ядерную ДНК. человека ДНК митохондрий (0, 5%) и отдельных кольцевых ДНК в ядре и цитоплазме – это внехромосомная ДНК.
Геном человека Общее количество ДНК в 46 хромосомах в: соматической клетке человека имеет длину 2 м и 6, 4∙ 109 пар оснований; в гаплоидной – 3, 2∙ 109 пар оснований. Основное количество ДНК локализовано в хромо сомах (99, 5%), то есть, составляет ядерную ДНК. человека ДНК митохондрий (0, 5%) и отдельных кольцевых ДНК в ядре и цитоплазме – это внехромосомная ДНК.
 Геном человека 25% ДНК – это повторяющиеся пос ледовательности, которые встреча ются в геноме в различной частоте копий Менее 2% гены, кодирую щие белки у человека 20% последовательности кодирую щие РНК, 13% интроны и межген ные участки. Более 50% составляют не кодирую щие последовательности.
Геном человека 25% ДНК – это повторяющиеся пос ледовательности, которые встреча ются в геноме в различной частоте копий Менее 2% гены, кодирую щие белки у человека 20% последовательности кодирую щие РНК, 13% интроны и межген ные участки. Более 50% составляют не кодирую щие последовательности.
 4 класса элементов ДНК, возникших из транс позонов:") Геном челов Повторяющиеся последовательности включают: 1) 4 класса элементов ДНК, возникших из транс позонов: последовательности SINЕ, LТR-транспо- зоны (ретровирусподобные) и ДНК-транспозоны; 2) повторы, состоящие из 2 -3 нуклеотидов; 3) дупликации больших сегментов ДНК из разных частей генома. Основная их часть возникла из обратнотранск ри бируемой ДНК, и представляют собой паразитические са мопроизводящиеся элементы ДНК, имеющие свои промото ры. Повторы могут быть местом, где создаются гены с новы ми функциями и происходит перестройка генов. У человека все паразитические повторы ДНК кажутся старыми; нет доказательств их эволюции.
Геном челов Повторяющиеся последовательности включают: 1) 4 класса элементов ДНК, возникших из транс позонов: последовательности SINЕ, LТR-транспо- зоны (ретровирусподобные) и ДНК-транспозоны; 2) повторы, состоящие из 2 -3 нуклеотидов; 3) дупликации больших сегментов ДНК из разных частей генома. Основная их часть возникла из обратнотранск ри бируемой ДНК, и представляют собой паразитические са мопроизводящиеся элементы ДНК, имеющие свои промото ры. Повторы могут быть местом, где создаются гены с новы ми функциями и происходит перестройка генов. У человека все паразитические повторы ДНК кажутся старыми; нет доказательств их эволюции.
 Для генома человека уста Ø Плотность генов в геноме человека ниже, чем у других видов: на 106 н. п. у дрожжей приходится 483 гена, у дрозофилы – 117, у человека – 7 генов, в 16 хромосоме – 16, в среднем на геном – 12 генов Для генов человека характерна боль шая протяженность интронов, 35% ге нов считываются с разных рамок, 45% РНК подвержены альтернативному сплайсингу. У человека гены отличаются большей сложностью: большим числом доменов и комбинациями доменов. Типичный ген человека включает примерно 28 тыс. п. н. , имеет 8 экзонов и кодирует 447 аминокислот. Самый большой ген – ген мышечного белка дистро фина (2, 4 х106 п. н. ). Для белка титина установлена после довательность из 27 тыс н. п. и 234 экзонов. Это самое большое количество экзонов геноме человека.
Для генома человека уста Ø Плотность генов в геноме человека ниже, чем у других видов: на 106 н. п. у дрожжей приходится 483 гена, у дрозофилы – 117, у человека – 7 генов, в 16 хромосоме – 16, в среднем на геном – 12 генов Для генов человека характерна боль шая протяженность интронов, 35% ге нов считываются с разных рамок, 45% РНК подвержены альтернативному сплайсингу. У человека гены отличаются большей сложностью: большим числом доменов и комбинациями доменов. Типичный ген человека включает примерно 28 тыс. п. н. , имеет 8 экзонов и кодирует 447 аминокислот. Самый большой ген – ген мышечного белка дистро фина (2, 4 х106 п. н. ). Для белка титина установлена после довательность из 27 тыс н. п. и 234 экзонов. Это самое большое количество экзонов геноме человека.
 Гены распределены по 24 хромосом. Расположение генов на хромосоме чередуется: участки с плотной локализацией (богатые генами) разделены участками бедными генами ( «генные пустыни» ). Наибольшую плотность имеет хромосома 19, наименьшую 13 и У-хромосома. Различается плотность и в разных частях хромосомы: в 21 хромосоме в длинном плече плотность в прокси мальном отделе составляет 1 ген на 304 т. п. н, а в дистальном – 1 на 64 т. н. п. ; в других участках длинного плеча генов практически нет Сотни генов вероятно получены человеком в результате горизонтальный передачи от бактерий. 6500 генов установлены экспе риментально (по функции продукта, наличию мутаций, тканеспеци фичности и т. д. ). Хромосомная локализация выявлена для около 20 тыс. генов (из них 740 генов РНК). Идентифицированы гены прак тически всех наиболее часто встречаемых (320) и сравнительно ред ких (около 170) наследственных заболеваний, 130 онкогенов. Общее количество генов обеспечивающих развитие наследствен ных болезней составляет 1400.
Гены распределены по 24 хромосом. Расположение генов на хромосоме чередуется: участки с плотной локализацией (богатые генами) разделены участками бедными генами ( «генные пустыни» ). Наибольшую плотность имеет хромосома 19, наименьшую 13 и У-хромосома. Различается плотность и в разных частях хромосомы: в 21 хромосоме в длинном плече плотность в прокси мальном отделе составляет 1 ген на 304 т. п. н, а в дистальном – 1 на 64 т. н. п. ; в других участках длинного плеча генов практически нет Сотни генов вероятно получены человеком в результате горизонтальный передачи от бактерий. 6500 генов установлены экспе риментально (по функции продукта, наличию мутаций, тканеспеци фичности и т. д. ). Хромосомная локализация выявлена для около 20 тыс. генов (из них 740 генов РНК). Идентифицированы гены прак тически всех наиболее часто встречаемых (320) и сравнительно ред ких (около 170) наследственных заболеваний, 130 онкогенов. Общее количество генов обеспечивающих развитие наследствен ных болезней составляет 1400.
 генома человека") Предварительный список предполагаемых функций 26588 генов (60%) генома человека
Предварительный список предполагаемых функций 26588 генов (60%) генома человека
 ведениями о геноме человека аряду с сиквенсом являются енетические и физические арты хромосом. Генетические карты – это схемы, отра жающие порядок расположения всех генов человека, особенно генов наследственных болезней, регуляторных элементов и генет ческих маркеров, а также относительных расстояний между ними на хромосоме и других генетических элементов. Метод построения генетических карт называется генетическим картированием Генетическое расстояние между генами измеряется в сантиморганидах (с. М); одн с. М – это такое минимальное расстояние между генами, при котором кроссинговер возможен у 1% гамет и соответствует час тоте рекомбинаций между двумя локусами равной 1%. Длина генома человека равна примерно 3000 -3500 с. М. бэнды
ведениями о геноме человека аряду с сиквенсом являются енетические и физические арты хромосом. Генетические карты – это схемы, отра жающие порядок расположения всех генов человека, особенно генов наследственных болезней, регуляторных элементов и генет ческих маркеров, а также относительных расстояний между ними на хромосоме и других генетических элементов. Метод построения генетических карт называется генетическим картированием Генетическое расстояние между генами измеряется в сантиморганидах (с. М); одн с. М – это такое минимальное расстояние между генами, при котором кроссинговер возможен у 1% гамет и соответствует час тоте рекомбинаций между двумя локусами равной 1%. Длина генома человека равна примерно 3000 -3500 с. М. бэнды
 Физические карты – это схемы хромосом, отражающие физическое расстояние между генами выражаемое в па рах нуклеотидов. На хромосомах после дифференциаль ного окрашивания выявляются поперечные диски (бэн ды). Размер небольшого диска равен приблизительно 1 млн н. п. на физических картах. Физическая карта фрагмента линейной ДНК 5500 п
Физические карты – это схемы хромосом, отражающие физическое расстояние между генами выражаемое в па рах нуклеотидов. На хромосомах после дифференциаль ного окрашивания выявляются поперечные диски (бэн ды). Размер небольшого диска равен приблизительно 1 млн н. п. на физических картах. Физическая карта фрагмента линейной ДНК 5500 п
 Карты генов наследст венных болезней, их расположение в геноме (гены, мутации которых приведут к возникнове нию болезни) называют ся патологической анатомией генома человека. Карта X-хромосомы человека
Карты генов наследст венных болезней, их расположение в геноме (гены, мутации которых приведут к возникнове нию болезни) называют ся патологической анатомией генома человека. Карта X-хромосомы человека