Lection 10-2.ppt
- Количество слайдов: 50



Генетические программы, реализуемые в симбиогенезе • 1. Судьба бактериальной клетки в клубеньковом симбиозе или процесс инфицирования. • 2. Индукция органогенеза • 3. Микоризная инфекция • 4. Системная регуляция.

bac. A hsn nif, fix nod. ABC nod. D exo, ndv chitinases, gluconases RH 42, 44 flavonoids Nmd (sym 12, sym 21, sym 39) ENOD 2, 5, 12, 40 ENOD 3, 14 Lb, GS, GOGAT, AAT, PEPC, MDH Растение Преинфекция Aut (nod 3, nod 4, sym 28, sym 29) Бактерия cmp Генетическое разделение стадий морфогенеза корневого клубенька

Последовательное функционирование Sym-13 и Sym-31, двух генов, влияющих на развитие симбиосом в корневых клубеньках гороха (Pisum sativum L. )
 Sparkle E 135 f sym 13")
Cоздание линии RBT, несущей две симбиотические мутации (Fix-) Sparkle E 135 f sym 13 Sprint-2 Fixsym 31 Sym 13 sym 13 Sym 31 sym 31 1. 2. 3. 4. 9 Sym 13 - Sym 31 3 sym 13 Sym 31 3 Sym 13 - sym 31 1 sym 13 sym 31 1. Только Fix+ Fb 2. и 3. Fix+ или Fix 4. Только Fix- Fix+ F 1 Fix+ F 2 Fix. Fb 1. Только Fix+ 2. и 3. Fix+ или Fix 4. Только Fix- Растение № 14 из поколения F 2, которое проявило Fix- фенотип в обоих Fb, было использовано для закладки линии RBT (sym 13 sym 31 )

Два основных типа клубенька: Аи. Б– недерминированны е Cи Dдетерминированны е

Получение линий L. japonicus, несущих несколько симбиотических мутаций в одном генотипе (Madsen et al. , 2010). Мутация nfr 1 -1 nfr 5 -2 snf 1 symrk-3 Тип мутации SNP, Q 493 stop вставка ретротранспозона LORE-2 SNP, T 265 I вставка ретротранспозона LORE-2 Детекция аллельного состояния CAPS-маркер, секвенирование аллельспецифичная ПЦР d. CAPS-маркер, секвенирование аллельспецифичная ПЦР Обозначение гомозиготы по мутации aa bb cc dd nfr 1, nfr 5 – мутации предотвращают образование ИН и Клуб. snf-1 - замена треонина в ССа. МК в 265 позиции вызывает спонтанное клубенькообразование symrk – LRR рецепторная киназа , мутация INF- NOD-

Получение линий L. japonicus, несущих несколько симбиотических мутаций в одном генотипе (Madsen et al. , 2010) (продолжение). aa. BB x AAbb aabbcc. DD x AABBCCdd Aa. Bb A-B- A-bb aa. B- aabb Aa. Bb. Cc. Dd 1 / 16 A-B-C-D- A-B-C-dd A-B-cc. D- A-B-ccdd aabb. CC x AABBcc A-bb-C-D- A-bb. C-dd A-bbcc. D- A-bbccdd aa. B-C-D- aa. B-C-dd aa. B-cc. D- aa. B-ccdd Aa. Bb. Cc aabb-C-D- aabb. C-dd aabbcc. D- aabbccdd A-B-C- A-B-cc A-bb. C- A-bbcc aa. B-C- aa. B-cc aabb. C- aabbcc 1 / 64 1 / 256

Распространение сигнала 1 -недерминированные клуб. 2 -детерминированные 1 2

Cпонтанное клубенькообразование symrk snf 1 мутантов а- хлорозные растения б - нормальные растения с - разнообразие клубеньков d, e – M. loti: lac. Z f – инфеционная нить g-дикий тип клубенька h, I – частично и полностью заполненный клуб. j, k – ТЭМ инф. нитей
 symrk-3 snf")
|Инфекция спонтанных клубеньковых примордий у двойных мутантов. ( a – f ) symrk-3 snf 1. а. «Пустые клубеньки» . ( b , c ) Встречаются отдельные ИН с бактериями. d. Старые полностью инокулированные клубеньки ( e , f )Закрытые дезориентированные ИН ( g ) Iинфекционные нити nup 85 -1 snf 1 root hair. ( h ) Полностью инфиц. nup 85 -1 snf 1 клубеньки. ( i ) ИН у nup 133 -3 snf 1 напротив спонт. клуб. . ( j ) Полностью инфицированныеnup 133 -3 snf 1 клубеньки ( k ) ИН L. Japonicus дикого типа. ( l ) Молодые клуб. дикого типаnodule.

Программа судьба бактериальной клетки Nod-factor M. loti Органогенез Symrk Nup-133 Nup-85

Программа судьба бактериальной клетки nap 1 pir-1 cerberus Nod-factor M. loti Органогенез Symrk nup-133 nup-85

Морфология и структура спонтанных клубеньков А-Структура спонтанного клубенька у snf 1 мутанта Б- nfr 1 snf 1 C-nfr 5 -snf 1 D-har 1 -3 snf 1 E - har 1 -3 snf 1 + nod. A F - har 1 -3 snf 1 + nod. C H-snf (GFP) I-nup 85 -1 snf 1

Морфология и структура спонтанных клубеньков J- nup 85 -1 snf 1 K – то же L-nfr 1 -1 nfr 5 -2 snf 1 M-то же N- nfr 1 -1 nfr 5 -2 symrk -3 snf 1 O - то же P- nfr 1 -1 nfr 5 -2 symrk -3 snf 1 Q-то же R, S, T – то же Двойная стр. – межкл. Карман Стрелка –ИТ из кармана
 nfr 1 -1 snf")
Инфекция спонтанных клубеньков с нод-фактором и без ( a ) nfr 1 -1 snf 1 - двойной мутант ( b ) nfr 5 -2 snf 1 - двойной мутант Частично инфицированные клуб ( c ) nfr 1 -1 snf 1 + nod. A ( d ) nfr 1 -1 nfr 5 -2 snf 1+nod. C (e) nfr 1 -1 snf 1 + nod. A

Инфекция клубеньков с нод-фактором и без e - nfr 1 -1 snf 1 nod. A f- nfr 1 -1 nfr 5 -2 snf 1+ nod. C G- то же большего увелич. СТРЕЛКА – ВНОВЬ ДЕЛЯЩИЕСЯ ИНФ. КЛУТКИ ДВ. СТРЕЛКА-БАКТ. ВНУТРИ КЛЕТОК

Механизм внутриклеточной инфекции А –дикий тип ИН. B - nfr 1 -1 nfr 5 -2 symrk-3 Snf 1 – ИН C-symrk-3 snf 1 ИН D-E symrk-3 snf 1 ТЭМ G- nfr 1 -1 nfr 5 -2 symrk-3 snf 1 – проникновение через разрыв

Механизм внутриклеточной инфекции H - nfr 1 -1 nfr 5 -2 snf 1 раст. инокул. Nod-C мутантом. Точка проникновения. I - nfr 1 -1 nfr 5 -2 snf 1 Nod. C бактерии в клубеньках J – одиночная клетка nfr 1 -1 nfr 5 -2 snf 1 растений см бакт. Мутантами, проникшими через разрывы. K – запертые ИН L-увеличенные инфекционные нити

Программа судьба бактериальной клетки Nap 1 цитоскелет pir-1 - цитоскелет Cerberus- ubiquitin E 3 ligase Nod-factor M. loti nfr-1 nfr-5 Органогенез Symrk nup-133 nup-85

Мутации гороха, нарушающие развитие симбиосом Itn sym 33 Idd sym 40 sym 41 Bad sym 31 sym 32

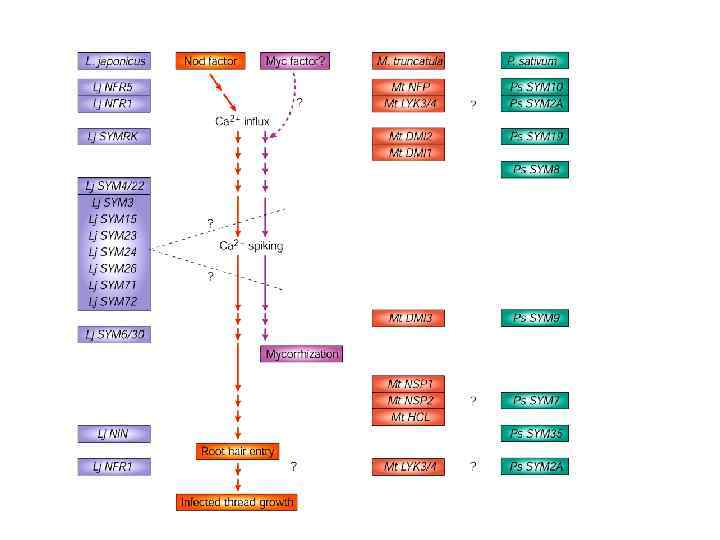
Путь передачи сигнала при развитии кубеньков у Lotus Обратить внимание: разное положение одинак. Генов в разных программах
 в процессе развития клубенька Sym 2")
Функционирования симбиотических генов гороха (Pisum sativum L. ) в процессе развития клубенька Sym 2 Sym 36 Sym 37 Sym 38 X X Sym 8 Sym 9 Sym 10 Sym 19 Sym 30 Sym 33 X X Sym 41 X XX X Sym 7 Sym 14 Sym 35 Sym 16 Sym 34 XX X
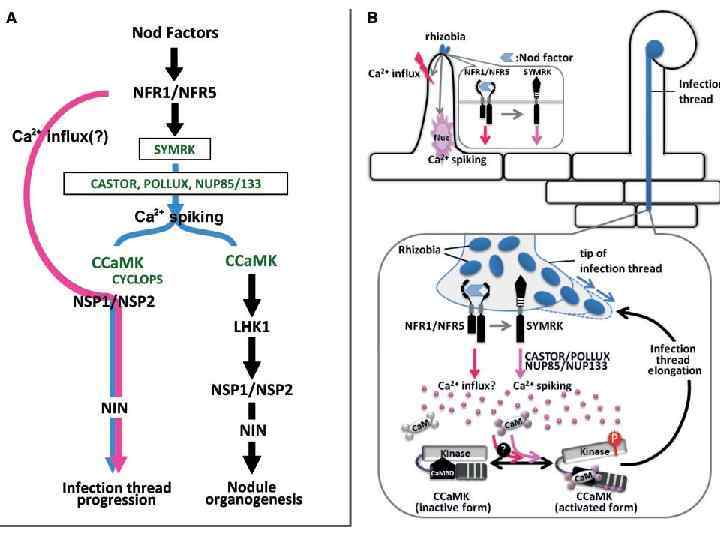

Путь передачи сигнала при развитии кубеньков у Lotus Обратить внимание: разное положение одинак. Генов в разных программах
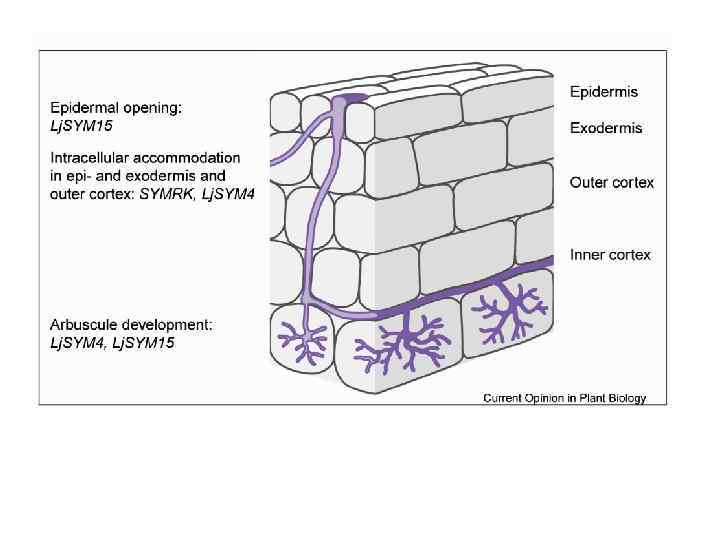

Signal exchange between arbuscular mycorrhizal fungus and host-plant at earliest stages of interaction A diffusible Myc factor from arbuscular mycorrhizal fungus induces symbiosis-specific Mt. ENOD 11 expression in roots of Medicago truncatula Kosuta S. et al. (2003) Plant Physiol. 131(3): 952– 962. Myc factor Branching factors flavonoids, sesquiterpenes a, b, g – induction of reporter fusion p. Mt. ENOD 11 -gus. A in Ri T-DNA-transformed roots of M. truncatula separated from fungus Gigaspora rosea by cellophane membrane. Myc factor – fungal lipochitooligosaccharide symbiotic Sesquiterpene strigolactone 5 -deoxy-strigol с and h signal (general structure). – roots cultivated without АМ fungus. isolated from Lotus japonicus roots. Akiyama K. et al. (2005) Nature. 435(9): 824 -827. a, scale bar, 0. 2 cm; b, с, scale bar, 2 cm; g, h, scale bar, 150 mkm. Maillet F. et al. (2011) Nature. Vol. 469. P. 58– 64.

Increase of cell size and nuclear volume in M. truncatula nodules Longitudinal nodule section I II III Blue staining: Nuclear DNA stained with DAPI Green staining: Microtubules are marked with MAP 4 -GFP

Plant development relies on constant coordination/interplay of the mitotic cell cycle and differentiation programs Cell cycle Differentiation Cell/tissue specificity Meristem Cell cycle exit Division arrest Expression of cell/tissue specific genes ORGAN Plant organs develop from meristems and their differentiation necessitates division arrest

How do eukaryotic cells grow? In eukaryotes, a positive correlation exists between ploidy levels and cell sizes within the same cell types. Polyploid cells are generated via endoreduplication cycles where the genome is duplicated without mitosis. Repeated endocycles lead to the formation 4 C, 8 C, 16 C, 32 C, … from 2 C cells. S ENDOREDUPLICATION G 2 G 1 -S-G 2 G 1 M MITOTIC CYCLE G 1 -S-G 2 -M
 G 2")
The cell cycle is regulated by consecutive actions of cyclin-dependent kinases (CDKs) G 2 S G 1 cyclins Mitotic cyclins (unstable proteins) G 1 M IT O SI S CDKs

Premature degradation of mitotic cyclins results in the inactivation of mitotic cyclin-dependent kinases G 2 S G 1 cyclins G 1 M IT O SI S CDKs Premature degradation of mitotic cyclins abolishes G 2 -M transition/mitosis

The ubiquitin-26 S proteasome pathway Activation Uba/E 1 Conjugation/Ligation Degradation Target protein Ubc/E 2 E 3 Polyubiquitination requires the E 1 (ubiquitin activating) E 2 (ubiquitin conjugating) E 3 (ubiquitin ligase) enzyme activities Polyubiquitinylated proteins are degraded by the 26 S proteasome : ubiquitin E 3 : ubiquitin ligase (substrate selection, polyubiquitinylation)

CCS 52: Cell Cycle Switch, 52 k. Da protein an activator of the Anaphase Promoting Complex E 3 ubiquitin ligase VII I VI II V III IV PEST P P P DRFIPSR AYTTLLRTALFG C-box I II IV V VI VII P WD 40 repeats CSM In plants, two classes of the Ccs 52 proteins exist: Ccs 52 A and Ccs 52 B (distinct functions and expression patterns (Tarayre et al. , 2004 Plant Cell). (Mt Ccs 52 A in nodules, Mt Ccs 52 B in roots) IR

CMSE - INRA Gene responses during early root interactions in Medicago truncatula Arbuscular mycorrhizal fungi Pseudomonas fluorescens plant genes fungal genes plant genes Comparative transcriptomics of root interactions with mycorrhizal fungi and beneficial bacteria MPMI, Melida 2005

CMSE - INRA Signal-related gene responses in nonrhizobial beneficial root interactions Vivienne Gianinazzi-Pearson Arbuscular mycorrhizal fungi PGPR MPMI, Melida 2005 Unité Mixte de Recherche Plante-Microbe-Environnement Centre de Microbiologie du Sol et de l’Environnement INRA-DIJON, F

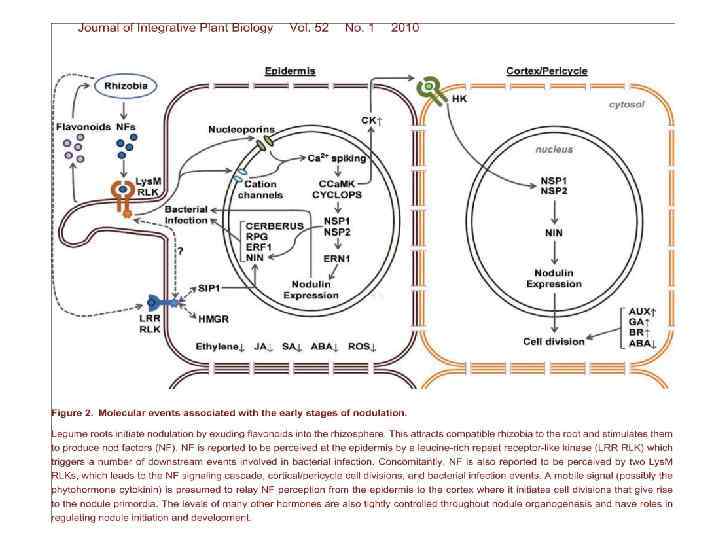
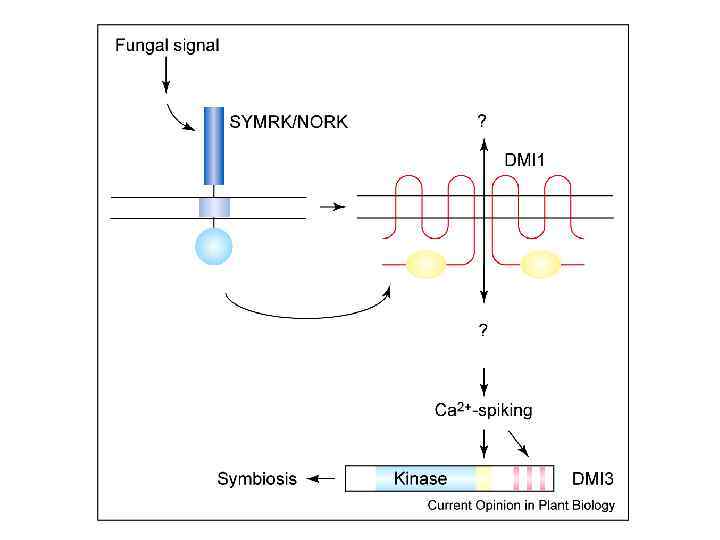
CMSE - INRA Appressorium-induced plant and fungal genes Medicago truncatula Jemalong Myc + J 5 Myc - dmi 3 -1(Mtsym 13) CCa. MPK Myc factors + Nod factors BEG 141 BEG 12 Glomus mosseae G. intraradices app/cm root arb 6 Ané et al. 2004 Endre et al. 2002 Mitra et al. 2004 Lévy et al. 2004 mycorrhiza MPMI, Melida 2005 4 DMI 1 (cation channel) 2 DMI 2 (LRR receptor-like kinase) DMI 3 (CCa. MPK) nodulation 0 24 ap 48 72 96 120 144 hai

CMSE - INRA P spore germination hyphal branching extraradical mycelium appressoria P C arbuscules MPMI, Melida 2005

CMSE - INRA Plant gene expression in compatible and incompatible M. truncatula genotypes Myc+ J 5 G. mosseae Myc- (CCa. MPK) annexin Ca 2+ dpdt lipid binding protein MAPK nitrate reductase Sn ribonucleoprotein eukaryotic IF 4 A eukaryotic IF 4 F 40 S RP S 3 60 S RP L 7 60 S RP L 18 a elongation factor 1 - fungal signals (Myc factors) expression ratio i/ni 5 MPMI, Melida 2005 0 -3 Ca 2+/calmodulin-dependent gene activation Weidmann et al. MPMI, 2004
 CMSE - INRA Gm. Mt.")
Appressorium-activated plant genes in G. mosseae-M. truncatula interactions (SSH) CMSE - INRA Gm. Mt. Es c. DNA library = inoculated-non inoculated roots (120 hai) plant genes 29 up-regulated (29 novel for mycorrhiza) annexin Ca 2+ dpdt lipid binding protein MAP-kinase nitrate reductase MPMI, Melida 2005 small nuclear ribonucleoprotein eukaryotic initiation factor 4 A eukaryotic initiation factor (ISO) 4 F 40 S ribosomal protein S 3 60 S ribosomal protein L 7 60 S ribosomal protein L 22 elongation factor 1 Weidmann et al. MPMI, 2004

Эксудаты корней изменяют способ прикрепления бактерий R. leguminosarum bv. viciae к поверхности (слайд предоставлен Алланом Доуни, Центр Джона Иннеса, Великобритания ) В присутствии корневых выделений Без корневых выделений (не флавоноиды и не результат экспрессии nod-генов бактерий) Активным компонентом эксудатов является метаболит небольшого размера

CMSE - INRA Plant gene responses to beneficial rhizosphere bacteria and comparative transcriptomics Mycorrhizal fungus Glomus mosseae Rhizobacteria Pseudomonas fluorescens N 2 -fixing bacteria Sinorhizobium meliloti MPMI, Melida 2005

CMSE - INRA P. fluorescens C 7 R 12/M. truncatula ni i rhizosphere intercellular intracellular Root colonisation Mutant (pseu+) dmi 3(Mtsym 13) Root fresh weight (mg) Wild-type Mt 80 80 70 70 60 60 50 40 30 10 4 6 8 dai MPMI, Melida 2005 Non inoculated 30 20 20 0 Inoculated 50 40 14 21 10 0 4 6 8 14 21 dai Sanchez et al. Plant Physiol. , 2005

CMSE - INRA Up-regulated plant genes in early P. fluorescens/ M. truncatula root interactions (SSH) Mt. Pf. Es c. DNA library = inoculated-non inoculated roots 58 plant genes no homology + unknown function primary metabolism signal transduction protein synthesis and processing cell division cell wall défense / stress miscellaneous gene expression and RNA metabolism MPMI, Melida 2005 biotic/abiotic stimuli membrane transport Sanchez et al. Plant Physiol. , 2005
 inactivation on root responses")
CMSE - INRA Influence of DMI 3 (Mt. SYM 13) inactivation on root responses to P. fluorescens C 7 R 12 plant genotype Myc - mutant Wild-type J 5 dai: receptor kinase ABC transporter b. ZIP transcription factor casein kinase II GH 3 small GTP binding protein protease inhibitor 14. 3. 3 4 6 8 14 21 (CCa. MPK) 4 6 8 14 21 expression ratio i/ni +3 1 -3 leucine zipper TFIIS TFIIB PR 10 Bacterial signal perception affected in dmi 3(Mtsym 13) Ca 2+/calmodulin-related signalling pathway MPMI, Melida 2005 Sanchez et al. Plant Physiol. , 2005

CMSE - INRA M. truncatula root interactions with beneficial microorganisms : comparative transcriptomics Early stage interactions Gm (Gm + Pf SSH) Pf Sm Gm Myc(CCa. MPK) signal transduction, transcription MPMI, Melida 2005 RLPK 14. 3. 3 casein kinase II s. GTP-binding protease inh nitrate red annexin CLB 1 MAPK Sn. RNP beta ZIP leucine zipper TFIIS 40 S S 3 60 S L 7 60 S L 18 a putative EF EIF-4 a EIF-4 f EF-1 a GH 3 ABC transp Gm: G. mosseae Pf: P. fluorescens Sm: S. meliloti expression ratio i/ni 4 0 -3 Sanchez et al. Plant Physiol. , 2005 Gianinazzi-Pearson et al. in press
 Gm Pf Sm glut synth PR")
CMSE - INRA Late stage interactions (Gm SSH) Gm Pf Sm glut synth PR 10 protease inh germin-like Mt. Gm. Ls 136 Mt. Gm. Ls 291 myb transf nodulin 26 PWIP GST Gm: G. mosseae Pf: P. fluorescens Sm: S. meliloti expression ratio i/ni 4 0 -3 Sanchez et al. New Phytol. , 2004 Shared plant cell programmes in non-rhizobial beneficial interactions CCa. MPK-dependent gene activation in M. truncatula Common pathway(s) in the perception of G. mosseae and P. fluorescens signals, different from S. meliloti MPMI, Melida 2005

Legume genes controlling interactions with beneficial soil microbes Arbuscular mycorrhizal fungi Nodule bacteria Beneficial rhizosphere bacteria
Lection 10-2.ppt