

ГАМЕТОГЕНЕЗ 2013.ppt
- Количество слайдов: 93
 ГАМЕТОГЕНЕЗ
ГАМЕТОГЕНЕЗ
 ГАМЕТОГЕНЕЗ – це процес утворення і дозрівання статевих клітин в статевих залозах: жіночих статевих клітин - яйцеклітин (овогенегез) і чоловічих статевих клітин – сперматозоїдів (сперматогенез). Мейоз – ділення статевих клітин, результатом якого є утворення гамет з гаплоїдним набором хромосом. Мейоз складається з двох послідовних поділів, під час яких з однієї клітини з диплоїдним набором хромосом утворюються чотири гаплоїдні клітини.
ГАМЕТОГЕНЕЗ – це процес утворення і дозрівання статевих клітин в статевих залозах: жіночих статевих клітин - яйцеклітин (овогенегез) і чоловічих статевих клітин – сперматозоїдів (сперматогенез). Мейоз – ділення статевих клітин, результатом якого є утворення гамет з гаплоїдним набором хромосом. Мейоз складається з двох послідовних поділів, під час яких з однієї клітини з диплоїдним набором хромосом утворюються чотири гаплоїдні клітини.
 Перший мейотичний поділ – редукційний Лептотена - спіралізація інтерфазних редуплікованих хромосом Кожна хромосома складається з двох сестринських хроматид, проте, вони настільки зближені, що кожна хромосома здається одинарною. По всій довжині можна побачити потовщення - хромомери. Кожна хромосома обома кінцями прикріплена до ядерної мембрани. Підкреслимо, що в лептотені хромосоми подвоєні, а отже, на цій стадії наявна диплоїдна кількість (2 n) подвоєних сестринських хроматид (їхня загальна кількість, 4 n, є результатом редуплікації в S-періоді).
Перший мейотичний поділ – редукційний Лептотена - спіралізація інтерфазних редуплікованих хромосом Кожна хромосома складається з двох сестринських хроматид, проте, вони настільки зближені, що кожна хромосома здається одинарною. По всій довжині можна побачити потовщення - хромомери. Кожна хромосома обома кінцями прикріплена до ядерної мембрани. Підкреслимо, що в лептотені хромосоми подвоєні, а отже, на цій стадії наявна диплоїдна кількість (2 n) подвоєних сестринських хроматид (їхня загальна кількість, 4 n, є результатом редуплікації в S-періоді).
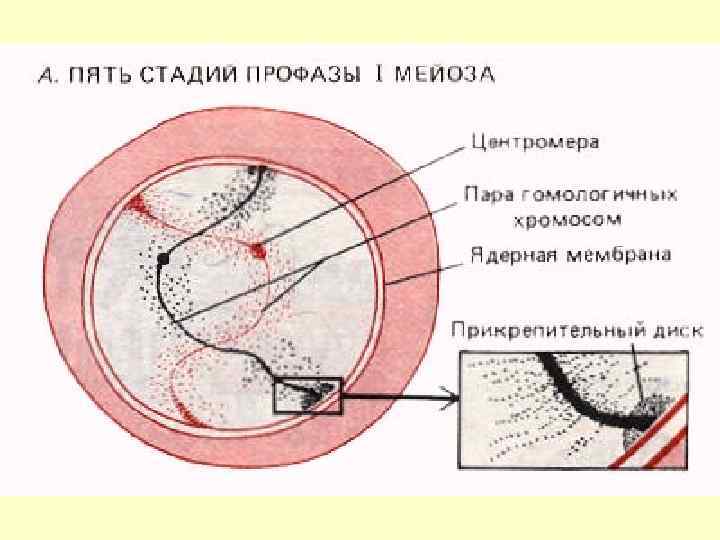
 Зиготена- кон’югація гомологічних хромосом. Утворення синаптонемального комплексу. Синтез специфічної ДНК. Внаслідок попарної кон’югації хромосомгомологів утворюються хромосомні асоціації – біваленти. До того ж кожна гомологічна хромосома пари складається з двох тісно наближених сестринських хроматид, тому кожну пару хромосом називають «тетрада» . Кількість бівалентів, що утворюються внаслідок попарної кон’югації гомологічних хромосом у два рази менша, за вихідну кількість хромосом.
Зиготена- кон’югація гомологічних хромосом. Утворення синаптонемального комплексу. Синтез специфічної ДНК. Внаслідок попарної кон’югації хромосомгомологів утворюються хромосомні асоціації – біваленти. До того ж кожна гомологічна хромосома пари складається з двох тісно наближених сестринських хроматид, тому кожну пару хромосом називають «тетрада» . Кількість бівалентів, що утворюються внаслідок попарної кон’югації гомологічних хромосом у два рази менша, за вихідну кількість хромосом.
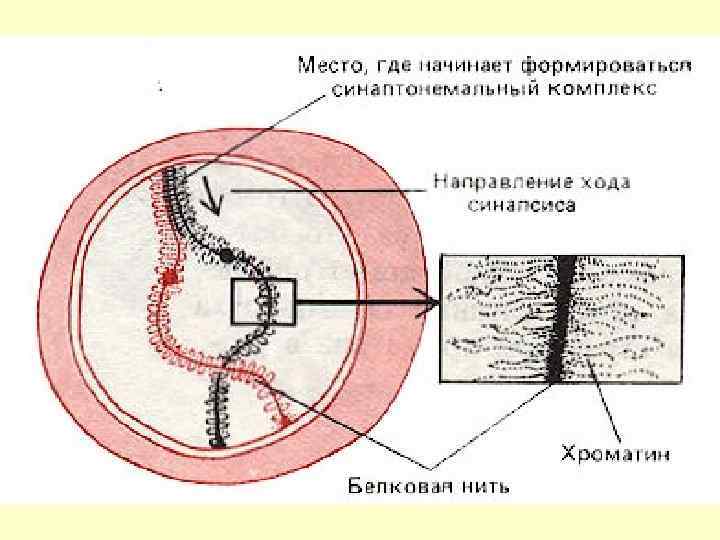
. Останні утворюються з") Cинаптонемальний комплекс субмікроскопічна структура, що складається зі щільних поздовжніх тяжів (cor). Останні утворюються з ниток на кожній гомологічній хромосомі (зауважимо, що вони ніколи не утворюються на мітотичних хромосомах). Між тяжами гомологічних хромосом, що зблизились, виникають поперечні волокна, й зближення через деякий час припиняється, що співпадає з завершенням формування синаптонемального комплексу.
Cинаптонемальний комплекс субмікроскопічна структура, що складається зі щільних поздовжніх тяжів (cor). Останні утворюються з ниток на кожній гомологічній хромосомі (зауважимо, що вони ніколи не утворюються на мітотичних хромосомах). Між тяжами гомологічних хромосом, що зблизились, виникають поперечні волокна, й зближення через деякий час припиняється, що співпадає з завершенням формування синаптонемального комплексу.
 - кросинговер. Синтез невеликої к-ті ДНК для відновлення") Пахітена – (стадія товстих ниток) - кросинговер. Синтез невеликої к-ті ДНК для відновлення молекул ДНК. Ампліфікація рибосомних генів і утворення хромосом типу “лампових щіток”. Хромосоми продовжують спіралізуватись. Максимально ущільнююється всі структури синаптонемального комплексу. В результаті утворюється одна вкорочена потовщена мейотична хромосома, яка складається з чотирьох хроматид. Число таких товстих пахітенних хромосом гаплоїдне (1 n), проте вони складаються з двох об’єднаних гомологів, кожний з яких має по дві сестринські хроматиди. Отже, і на цій стадії кількість ДНК – 4 с, а число хроматид – 4 n. .
Пахітена – (стадія товстих ниток) - кросинговер. Синтез невеликої к-ті ДНК для відновлення молекул ДНК. Ампліфікація рибосомних генів і утворення хромосом типу “лампових щіток”. Хромосоми продовжують спіралізуватись. Максимально ущільнююється всі структури синаптонемального комплексу. В результаті утворюється одна вкорочена потовщена мейотична хромосома, яка складається з чотирьох хроматид. Число таких товстих пахітенних хромосом гаплоїдне (1 n), проте вони складаються з двох об’єднаних гомологів, кожний з яких має по дві сестринські хроматиди. Отже, і на цій стадії кількість ДНК – 4 с, а число хроматид – 4 n. .
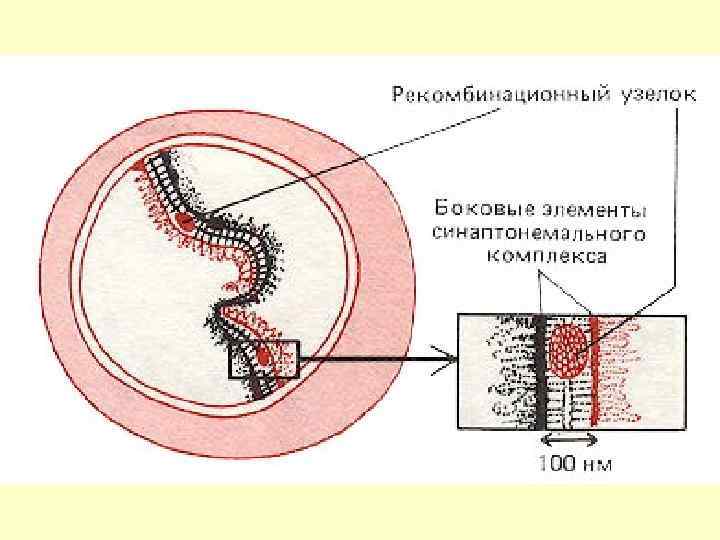
 У пахітені в повздовжній щілині синаптонемального комплексу з’являються рекомбінаційні вузлики, які беруть участь у перехресті певних ділянок гомологічних хромосом, забезпечуючи їхній обмін. Це явище отримало назву кросинговер.
У пахітені в повздовжній щілині синаптонемального комплексу з’являються рекомбінаційні вузлики, які беруть участь у перехресті певних ділянок гомологічних хромосом, забезпечуючи їхній обмін. Це явище отримало назву кросинговер.
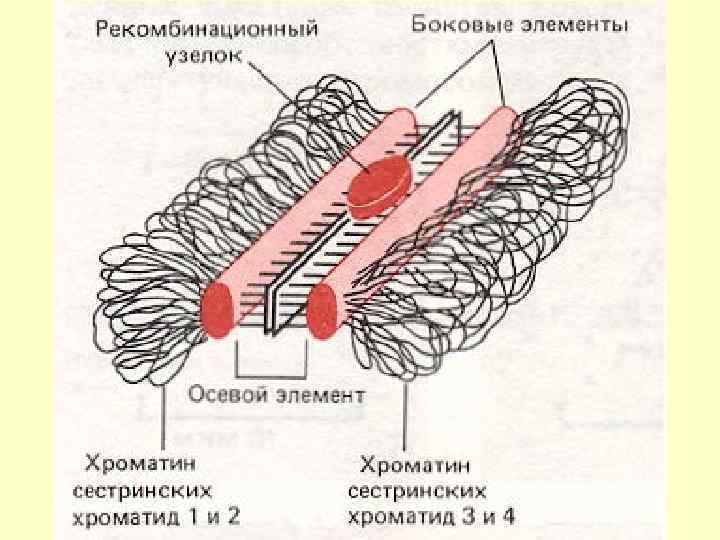
 У пахітені перехрести ще практично не видимі, тоді як пізніше вони будуть проявлятися у вигляді Х-подібних фігур, що дістали назву хіазми. Важливо підкреслити, що в пахітені відбувається синтез невеликої кількості ДНК (1% від геному), яка відрізняється тим, що містить повторювальні послідовності нуклеотидів. Однак цей синтез є репаративним (відбувається відновлення втраченої кількості ДНК).
У пахітені перехрести ще практично не видимі, тоді як пізніше вони будуть проявлятися у вигляді Х-подібних фігур, що дістали назву хіазми. Важливо підкреслити, що в пахітені відбувається синтез невеликої кількості ДНК (1% від геному), яка відрізняється тим, що містить повторювальні послідовності нуклеотидів. Однак цей синтез є репаративним (відбувається відновлення втраченої кількості ДНК).

 розходження гомологів. Синтез РНК Цілісність синаптонемального комплексу порушується,") Диплотена – (стадія розходження ниток) розходження гомологів. Синтез РНК Цілісність синаптонемального комплексу порушується, він розпадається, що дозволяє двом гомологічним хромосомам бівалента дещо відійти одна від одної. Однак, хромосоми все ще зв’язані однією або декількома хіазмами, тобто місцями, де відбувся кросинговер.
Диплотена – (стадія розходження ниток) розходження гомологів. Синтез РНК Цілісність синаптонемального комплексу порушується, він розпадається, що дозволяє двом гомологічним хромосомам бівалента дещо відійти одна від одної. Однак, хромосоми все ще зв’язані однією або декількома хіазмами, тобто місцями, де відбувся кросинговер.
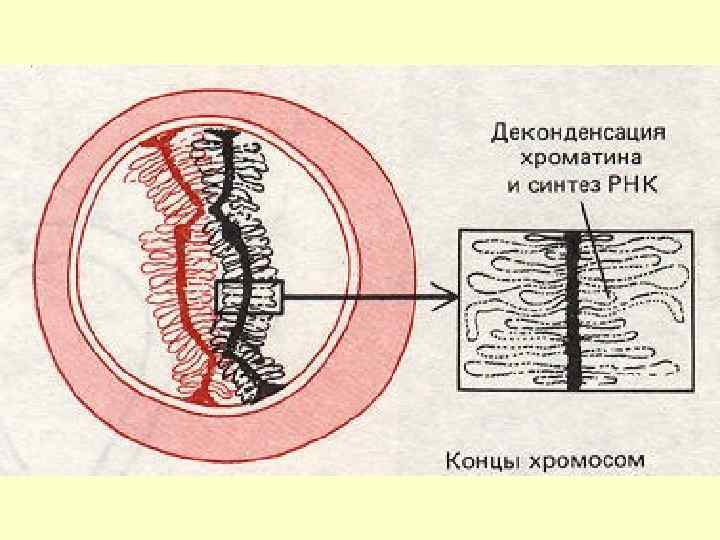
 В овоцитах диплотена може розтягтись на місяці, або навіть роки: на цій стадії хромосоми хоча й трошки вкорочуються та конденсуються, проте деякі їхні ділянки, навпаки, зазнають деконденсації (хромосоми набувають вигляд лампових щіток) й активно транскрибуються – відбувається утворення величезної кількості рибосом на ампліфікованих ядерцях і синтез м. РНК (що є необхідним для реалізації білкового синтезу на ранніх етапах розвитку зародка).
В овоцитах диплотена може розтягтись на місяці, або навіть роки: на цій стадії хромосоми хоча й трошки вкорочуються та конденсуються, проте деякі їхні ділянки, навпаки, зазнають деконденсації (хромосоми набувають вигляд лампових щіток) й активно транскрибуються – відбувається утворення величезної кількості рибосом на ампліфікованих ядерцях і синтез м. РНК (що є необхідним для реалізації білкового синтезу на ранніх етапах розвитку зародка).
 ДІАКІНЕЗ - синтез РНК не відбувається, хромосоми конденсуються, потовщуються й відділяються від ядерної мембрани. Тепер чітко видно, що кожний бівалент містить чотири окремі хроматиди, причому кожна пара сестринських хроматид сполучена центромерою, тоді як несестринські хроматиди, де відбувся кросинговер, зв’язані хіазмами. Хіазми поступово сповзають до термінальних зон бівалентів (кінці хромосом), тому біваленти набувають форми кільцеподібних структур. Закінчення діакінезу означає закінчення профази I.
ДІАКІНЕЗ - синтез РНК не відбувається, хромосоми конденсуються, потовщуються й відділяються від ядерної мембрани. Тепер чітко видно, що кожний бівалент містить чотири окремі хроматиди, причому кожна пара сестринських хроматид сполучена центромерою, тоді як несестринські хроматиди, де відбувся кросинговер, зв’язані хіазмами. Хіазми поступово сповзають до термінальних зон бівалентів (кінці хромосом), тому біваленти набувають форми кільцеподібних структур. Закінчення діакінезу означає закінчення профази I.

 В період діакінезу матеріал овоциту не є інертним: у яйцеклітині людини (як і в більшості яйцеклітин із солітарним і фолікулярним типом живлення) він виконує роль матриці для синтезу всіх типів РНК, утворення яких відбувається переважно про запас (тобто для використання вже заплідненою яйцеклітиною). До моменту свого використання такі запасені РНК, у першу чергу м. РНК, заблоковуються білковими молекулами (такі міцні рибонуклеопротеїнові комплекси називають інформосомами). Аналогічно, про запас, накопичуються трофічні включення, і у першу чергу, жовток.
В період діакінезу матеріал овоциту не є інертним: у яйцеклітині людини (як і в більшості яйцеклітин із солітарним і фолікулярним типом живлення) він виконує роль матриці для синтезу всіх типів РНК, утворення яких відбувається переважно про запас (тобто для використання вже заплідненою яйцеклітиною). До моменту свого використання такі запасені РНК, у першу чергу м. РНК, заблоковуються білковими молекулами (такі міцні рибонуклеопротеїнові комплекси називають інформосомами). Аналогічно, про запас, накопичуються трофічні включення, і у першу чергу, жовток.
 Особливості профази 1 мейозу: 1. Займає великий проміжок часу. 2. Складається із декількох фаз. 3. Спостерігається ріст овоцитів (накопичення поживних речовин) 4. На відміну від мітозу, хромосоми здатні до синтезу РНК, часткового синтезу ДНК, тобто приймають участь у певних подіях.
Особливості профази 1 мейозу: 1. Займає великий проміжок часу. 2. Складається із декількох фаз. 3. Спостерігається ріст овоцитів (накопичення поживних речовин) 4. На відміну від мітозу, хромосоми здатні до синтезу РНК, часткового синтезу ДНК, тобто приймають участь у певних подіях.
 Метафаза 1 Слід зазначити, що під час першого мейотичного поділу хромосоми прикріплюються до ахроматинових ниток інакше, ніж при звичайному мітозі. До однієї нитки підходить зразу пара гомологічних хромосом і займає місце, яке при звичайному мітозі властиве одній хромосомі. При цьому одна пара гомологічних хромосом прикріплюється до нитки, що тягнеться до одного полюса, а друга - до нитки, що тягнеться до протилежного.
Метафаза 1 Слід зазначити, що під час першого мейотичного поділу хромосоми прикріплюються до ахроматинових ниток інакше, ніж при звичайному мітозі. До однієї нитки підходить зразу пара гомологічних хромосом і займає місце, яке при звичайному мітозі властиве одній хромосомі. При цьому одна пара гомологічних хромосом прикріплюється до нитки, що тягнеться до одного полюса, а друга - до нитки, що тягнеться до протилежного.
 Анафаза 1 На цій стадії, гомологічні хромосоми, кожна з яких складається з двох хроматид, розходяться до протилежних полюсів. Отже, на кожному полюсі зосереджується вдвічі менша кількість подвійних хромосом, що відображається в назві цього поділу: його називають редукційним
Анафаза 1 На цій стадії, гомологічні хромосоми, кожна з яких складається з двох хроматид, розходяться до протилежних полюсів. Отже, на кожному полюсі зосереджується вдвічі менша кількість подвійних хромосом, що відображається в назві цього поділу: його називають редукційним
 До кінця першого ділення хромосомний набір редукується , перетворюючись із тетраплоїдного в диплоїдний, і як при мітозі, утворюється 2 клітини. Відмінність у тому, що при 1 поділі мейозу в кожну клітину попадають 2 сестринські хроматиди, об’єднані центромерою, а при мітозі – розділені хроматиди.
До кінця першого ділення хромосомний набір редукується , перетворюючись із тетраплоїдного в диплоїдний, і як при мітозі, утворюється 2 клітини. Відмінність у тому, що при 1 поділі мейозу в кожну клітину попадають 2 сестринські хроматиди, об’єднані центромерою, а при мітозі – розділені хроматиди.
 Таким чином, редукційний поділ приводить до утворення двох дочірніх клітин, кількість ДНК в яких становить 2 с, а хромосом – 2 n (кожна хромосома в парі гомологів складається з двох хроматид).
Таким чином, редукційний поділ приводить до утворення двох дочірніх клітин, кількість ДНК в яких становить 2 с, а хромосом – 2 n (кожна хромосома в парі гомологів складається з двох хроматид).
 Слідом за телофазою I настає короткий період між закінченням першого та початком другого мейотичного поділу клітини. Цей період дістав назву інтеркінезу. Він відрізняється від інтерфази відсутністю стадій G 1, S і G 2. Отже, між двома поділами редуплікація не відбувається й хромосоми залишаються компактними, зберігаючи свої морфологічні особливості.
Слідом за телофазою I настає короткий період між закінченням першого та початком другого мейотичного поділу клітини. Цей період дістав назву інтеркінезу. Він відрізняється від інтерфази відсутністю стадій G 1, S і G 2. Отже, між двома поділами редуплікація не відбувається й хромосоми залишаються компактними, зберігаючи свої морфологічні особливості.
 Другий мейотичний поділ Профаза II - значно коротша ніж профаза I. Хромосоми складаються зі з’єднаних центромерою двох хроматид, які починають відділятися одна від одної вздовж екваційної щілини (утримуючись лише центромерою). Швидко формується веретено поділу, зникають ядерця та ядерна оболонка. Слід підкреслити, що в результаті кросинговеру відбувається втрата ізогенності хроматидами, які входять до складу хромосом. Тому пара сестринських хроматид завжди є гетерогенною
Другий мейотичний поділ Профаза II - значно коротша ніж профаза I. Хромосоми складаються зі з’єднаних центромерою двох хроматид, які починають відділятися одна від одної вздовж екваційної щілини (утримуючись лише центромерою). Швидко формується веретено поділу, зникають ядерця та ядерна оболонка. Слід підкреслити, що в результаті кросинговеру відбувається втрата ізогенності хроматидами, які входять до складу хромосом. Тому пара сестринських хроматид завжди є гетерогенною
 Метафаза II. Ця стадія триває недовго. Кожна подвійна хромосома розміщується в екваторіальній площині клітини, а до їхньої центромери прикріпляються мікротрубочки веретена поділу. Анафаза II. На цій стадії відбувається остаточне відокремлення хроматид, і вони починають рухатись до протилежних полюсів. Тепер кожна дочірня хроматида набуває статусу хромосоми.
Метафаза II. Ця стадія триває недовго. Кожна подвійна хромосома розміщується в екваторіальній площині клітини, а до їхньої центромери прикріпляються мікротрубочки веретена поділу. Анафаза II. На цій стадії відбувається остаточне відокремлення хроматид, і вони починають рухатись до протилежних полюсів. Тепер кожна дочірня хроматида набуває статусу хромосоми.
 Телофаза II. Ця стадія є заключною. Хромосоми повністю деспіралізуються, формується ядро, ядерце, відбувається цитотомія, й, врешті-решт, утворюються дві клітини.
Телофаза II. Ця стадія є заключною. Хромосоми повністю деспіралізуються, формується ядро, ядерце, відбувається цитотомія, й, врешті-решт, утворюються дві клітини.
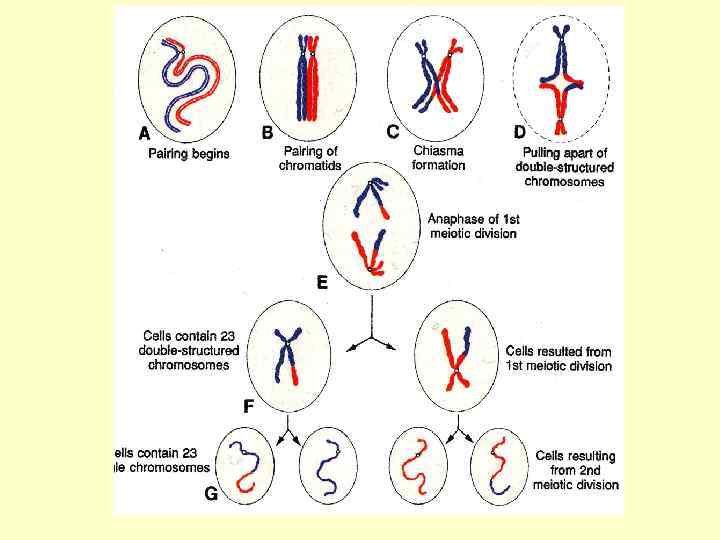
 Біологічне значення мейозу : кожна з чотирьох новоутворених клітин буде мати вдвічі менше хромосом, а кожна хромосома – удвічі меншу кількість ДНК. Отже, у цілому кількість хромосомних молекул ДНК зменшується в чотири рази. Крім того, при мейотичному поділі відбувається випадковий розподіл хромосом по дочірнім клітинам. Це й створює генетичну різноманітність у виникаючих гаплоїдних статевих клітинах.
Біологічне значення мейозу : кожна з чотирьох новоутворених клітин буде мати вдвічі менше хромосом, а кожна хромосома – удвічі меншу кількість ДНК. Отже, у цілому кількість хромосомних молекул ДНК зменшується в чотири рази. Крім того, при мейотичному поділі відбувається випадковий розподіл хромосом по дочірнім клітинам. Це й створює генетичну різноманітність у виникаючих гаплоїдних статевих клітинах.
 Овогенез
Овогенез
 - ріст – овоцит 1 порядку.") овогенез - розмноження - оогонії (к-ть поділів видоспецифічна) - ріст – овоцит 1 порядку. До настання статевої зрілості – період малого росту. Великий ріст – з набуттям статевої зрілості - дозрівання - мейоз
овогенез - розмноження - оогонії (к-ть поділів видоспецифічна) - ріст – овоцит 1 порядку. До настання статевої зрілості – період малого росту. Великий ріст – з набуттям статевої зрілості - дозрівання - мейоз
 Період розмноження У деяких тварин, наприклад у риб та амфібій, періодичність мітотичних поділів овогоній пов’язана з сезонним розмноженням і повторюється протягом усього життя. У ссавців овоґенез починається під час внутрішньоутробного розвитку й завершується до моменту народження.
Період розмноження У деяких тварин, наприклад у риб та амфібій, періодичність мітотичних поділів овогоній пов’язана з сезонним розмноженням і повторюється протягом усього життя. У ссавців овоґенез починається під час внутрішньоутробного розвитку й завершується до моменту народження.
 Кількість овоцитів в яєчнику людини у різні періоди онтогенезу: ПСК 100 Двомісячний плід 600 тис. П’ятимісячний плід 6, 8 млн. Дев’ятимісячний плід 1, 0 млн. 7 років 300 тис. Статева зрілість 40 – 80 тис. Використано впродовж життя 400 – 500 овоцитів
Кількість овоцитів в яєчнику людини у різні періоди онтогенезу: ПСК 100 Двомісячний плід 600 тис. П’ятимісячний плід 6, 8 млн. Дев’ятимісячний плід 1, 0 млн. 7 років 300 тис. Статева зрілість 40 – 80 тис. Використано впродовж життя 400 – 500 овоцитів
 З переходом до профази І овоцити першого порядку переходять до фази росту, яка складається з двох етапів: етапу малого й великого росту. Початок малого росту, або превітелогенез, співпадає з моментом вступу овоциту в профазу мейозу й триває до середини диплотени. На цьому етапові ріст клітини є незначним: незначно й пропорційно збільшуються як об'єм ядра, так і об'єм цитоплазми. У цитоплазмі накопичуються РНК, білки, рибосоми тощо.
З переходом до профази І овоцити першого порядку переходять до фази росту, яка складається з двох етапів: етапу малого й великого росту. Початок малого росту, або превітелогенез, співпадає з моментом вступу овоциту в профазу мейозу й триває до середини диплотени. На цьому етапові ріст клітини є незначним: незначно й пропорційно збільшуються як об'єм ядра, так і об'єм цитоплазми. У цитоплазмі накопичуються РНК, білки, рибосоми тощо.
 Приблизно з середини диплотени овоцити першого порядку вступають у період великого росту, при цьому різко збільшується об'єм цитоплазми (у сотні й тисячі разів): вона заповнюються жовтковими включеннями. Крім жовтка в овоциті в цей період відкладаються в резерв й інші компоненти - вуглеводи, ліпіди тощо, проте головною подією великого росту, безсумнівно, є вітелогенез.
Приблизно з середини диплотени овоцити першого порядку вступають у період великого росту, при цьому різко збільшується об'єм цитоплазми (у сотні й тисячі разів): вона заповнюються жовтковими включеннями. Крім жовтка в овоциті в цей період відкладаються в резерв й інші компоненти - вуглеводи, ліпіди тощо, проте головною подією великого росту, безсумнівно, є вітелогенез.
 Наступна фаза – фаза поділів дозрівання (під час якої здійснюється мейоз від метафази І до телофази ІІ) й вихід з фази діакінезу приурочена до досягнення самкою статевої зрілості й зумовлюється впливом статевих гормонів: гонадотропні гормони аденогіпофізу впливають на фолікулярний епітелій, який у відповідь продукує прогестерон або його аналоги.
Наступна фаза – фаза поділів дозрівання (під час якої здійснюється мейоз від метафази І до телофази ІІ) й вихід з фази діакінезу приурочена до досягнення самкою статевої зрілості й зумовлюється впливом статевих гормонів: гонадотропні гормони аденогіпофізу впливають на фолікулярний епітелій, який у відповідь продукує прогестерон або його аналоги.
 Овоцит зв’язаний з фолікулярними клітинами щілинними контактами, через які з останніх у овоцит надходить ц. АМФ. Високий рівень ц. АМФ блокує (шляхом ц. АМФ-залежного фосфорилювання) білковий фактор дозрівання, наявний в цитоплазмі овоциту. Під дією прогестерону (або його аналогів) щілинні контакти «розмикаються» , концентрація ц. АМФ у цитоплазмі овоциту падає, і, як наслідок, фактор дозрівання деблокується (дефосфорилюється). Цей фактор у активному стані викликає дезінтеграцію оболонки овоциту, розбирання ядерець та міграцію хромосом до майбутнього анімального полюсу, де і відбудуться поділи дозрівання.
Овоцит зв’язаний з фолікулярними клітинами щілинними контактами, через які з останніх у овоцит надходить ц. АМФ. Високий рівень ц. АМФ блокує (шляхом ц. АМФ-залежного фосфорилювання) білковий фактор дозрівання, наявний в цитоплазмі овоциту. Під дією прогестерону (або його аналогів) щілинні контакти «розмикаються» , концентрація ц. АМФ у цитоплазмі овоциту падає, і, як наслідок, фактор дозрівання деблокується (дефосфорилюється). Цей фактор у активному стані викликає дезінтеграцію оболонки овоциту, розбирання ядерець та міграцію хромосом до майбутнього анімального полюсу, де і відбудуться поділи дозрівання.
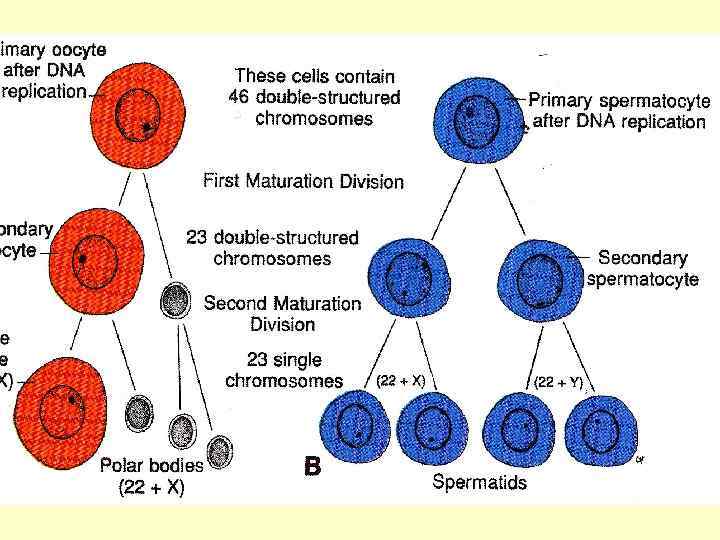
 Під час першого поділу дозрівання овоцит першого порядку ділиться на дві клітини – овоцит другого порядку та маленьку клітину – редукційне, або полярне тільце (первинне). У подальшому полярне тільце поділяється на дві такі ж маленькі клітини (вторинні полярні тільця), які ніякої участі у подальшому розвиткові не приймають. Овоцит же другого порядку здійснює другий поділ дозрівання шляхом виділення другого редукційного тільця таких же розмірів, як і перше. Після його виділення овоцит другого порядку перетворюється у зрілу яйцеклітину.
Під час першого поділу дозрівання овоцит першого порядку ділиться на дві клітини – овоцит другого порядку та маленьку клітину – редукційне, або полярне тільце (первинне). У подальшому полярне тільце поділяється на дві такі ж маленькі клітини (вторинні полярні тільця), які ніякої участі у подальшому розвиткові не приймають. Овоцит же другого порядку здійснює другий поділ дозрівання шляхом виділення другого редукційного тільця таких же розмірів, як і перше. Після його виділення овоцит другого порядку перетворюється у зрілу яйцеклітину.
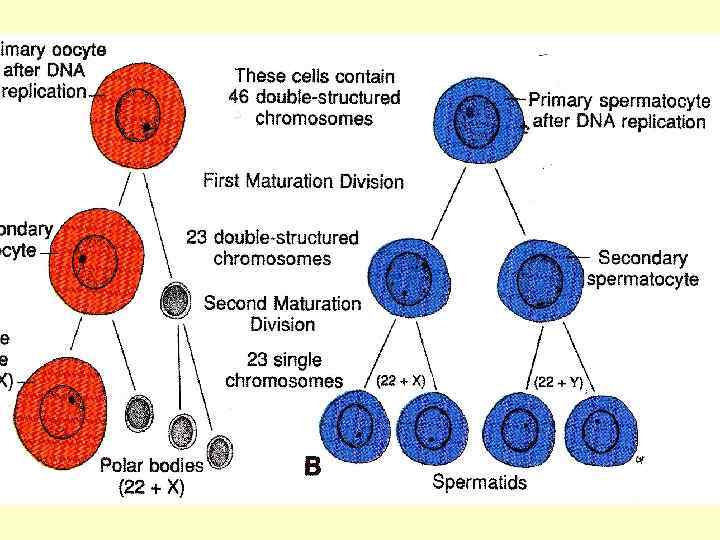
 Схема періодів овогенеза
Схема періодів овогенеза
 Превітелогенез Транскрипція р. РНК, т. РНК, білків припадає на стадії лептотени, зиготени, пахітени. Синтез р. РНК (5, 8 S, 18 S і 28 S р. РНК) проходитъ у амфібій шляхом ампліфікаціі рибосомних генів. Під час такого синтезу кількість р. РНК зростає більше, ніж у 1000 разів. Для 5 S р. РНК і т. РНК вистачає їхнього синтезу на всіх копіях генів, що є у геномі (5 S р. РНКгенів є декілька тисяч копій, т. РНК-генів – сотні копій).
Превітелогенез Транскрипція р. РНК, т. РНК, білків припадає на стадії лептотени, зиготени, пахітени. Синтез р. РНК (5, 8 S, 18 S і 28 S р. РНК) проходитъ у амфібій шляхом ампліфікаціі рибосомних генів. Під час такого синтезу кількість р. РНК зростає більше, ніж у 1000 разів. Для 5 S р. РНК і т. РНК вистачає їхнього синтезу на всіх копіях генів, що є у геномі (5 S р. РНКгенів є декілька тисяч копій, т. РНК-генів – сотні копій).
 У X. leavis за 3 -6 місяців утворюється 1011 рибосом, у ссавців – 106 рибосом. Швидкість їхнього утворення – 300 тис/сек. Якщо б синтез рибосом проходив звичайним шляхом, необхідно було б 400 років для синтезу такої кількості рибосом. Синтез м. РНК відбувається на стадії пахітени, диплотени й, іноді, діакінезу. Синтезується близько 4 х104 різновидностей м. РНК. Синтез м. РНК здійснюється на хромосомах типу лампових щіток у ссавців і амфібій і на політенних хромосомах у комах.
У X. leavis за 3 -6 місяців утворюється 1011 рибосом, у ссавців – 106 рибосом. Швидкість їхнього утворення – 300 тис/сек. Якщо б синтез рибосом проходив звичайним шляхом, необхідно було б 400 років для синтезу такої кількості рибосом. Синтез м. РНК відбувається на стадії пахітени, диплотени й, іноді, діакінезу. Синтезується близько 4 х104 різновидностей м. РНК. Синтез м. РНК здійснюється на хромосомах типу лампових щіток у ссавців і амфібій і на політенних хромосомах у комах.
 складається з однієї молекули ліповітеліну (ліпопротеїн") Ліпофосфопротеїн або вітелогенін (молекулярна маса 470 k. D) складається з однієї молекули ліповітеліну (ліпопротеїн 400 k. D) i двох молекул фосвітину (фосфопротеїд 40 к. D). У процесі синтезу жовтка формується полярність клітини: анімальний і вегетативний полюси – А-Ввісь або дорзо-вентральна (спинно-черевна) вісь. Одночасно з жовтком з апарату Гольджі утворюються кортикальні гранули, що розміщуються по периферії цитоплазми, синтезується велика кількість мітохондрій і інших органел.
Ліпофосфопротеїн або вітелогенін (молекулярна маса 470 k. D) складається з однієї молекули ліповітеліну (ліпопротеїн 400 k. D) i двох молекул фосвітину (фосфопротеїд 40 к. D). У процесі синтезу жовтка формується полярність клітини: анімальний і вегетативний полюси – А-Ввісь або дорзо-вентральна (спинно-черевна) вісь. Одночасно з жовтком з апарату Гольджі утворюються кортикальні гранули, що розміщуються по периферії цитоплазми, синтезується велика кількість мітохондрій і інших органел.
 ТИПИ ЖИВЛЕННЯ ЯЙЦЕКЛІТИН Фагоцитарний тип – у тварин, які не мають стабільних статевих залоз (губки, кишковопорожнинні). Жовточні гранули не утворюються Солітарний тип – у колоніальних гідроїдних поліпів, голкошкірих, ланцетника, деяких червів, безкрилих комах. Аліментарний тип: нутріментарний фолікулярний (черви членистоногі) (більшість тварин)
ТИПИ ЖИВЛЕННЯ ЯЙЦЕКЛІТИН Фагоцитарний тип – у тварин, які не мають стабільних статевих залоз (губки, кишковопорожнинні). Жовточні гранули не утворюються Солітарний тип – у колоніальних гідроїдних поліпів, голкошкірих, ланцетника, деяких червів, безкрилих комах. Аліментарний тип: нутріментарний фолікулярний (черви членистоногі) (більшість тварин)
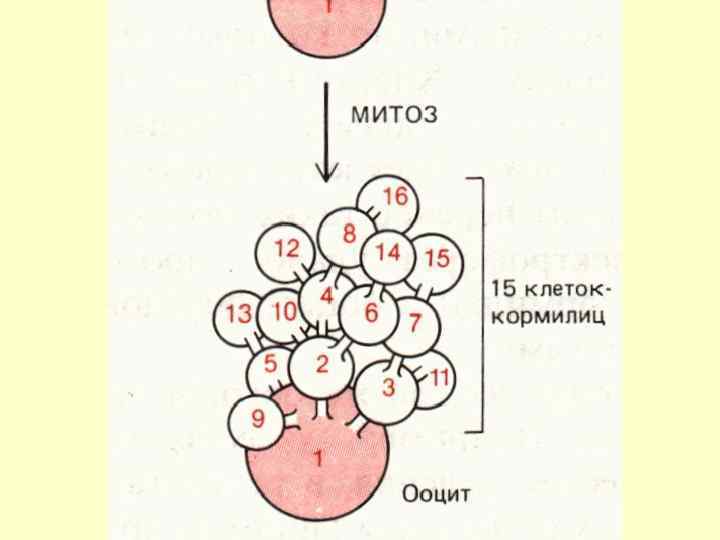
 БЛОКАДА МЕЙОЗУ: У відповідності зі стадією мейозу , на якій наступає блок мейозу тварини розподіляються наступним чином: 1. Мейоз зупиняється на стадії діакінезу профази 1. (Губки, деякі плоскі, круглі і кільчасті черви, молюски, а також для лисиці, собаки та коней). 2. На метафазі 1 мейоз блокується у губок, немертин, кільчастих червів, молюсків, майже у всіх комах. 3. На метафазі II блокується майже у всіх хордових (виключення – летючі миші, у них - на анафазі II).
БЛОКАДА МЕЙОЗУ: У відповідності зі стадією мейозу , на якій наступає блок мейозу тварини розподіляються наступним чином: 1. Мейоз зупиняється на стадії діакінезу профази 1. (Губки, деякі плоскі, круглі і кільчасті черви, молюски, а також для лисиці, собаки та коней). 2. На метафазі 1 мейоз блокується у губок, немертин, кільчастих червів, молюсків, майже у всіх комах. 3. На метафазі II блокується майже у всіх хордових (виключення – летючі миші, у них - на анафазі II).
 веретено") Овогенний мейоз має певні особливості порівняно з цим поділом під час сперматогенезу: 1) веретено поділу зміщене на периферію клітини; 2) цитокінез відбувається нерівномірно; 3) утворюються дві нерівнозначні клітини: овоцит ІІ і полярне тільце. Ссавцям також властиве явище вибірковості овоцитів, яке полягає в овуляції одного або декількох зрілих овоцитів під час статевої зрілості за дії гормонів.
Овогенний мейоз має певні особливості порівняно з цим поділом під час сперматогенезу: 1) веретено поділу зміщене на периферію клітини; 2) цитокінез відбувається нерівномірно; 3) утворюються дві нерівнозначні клітини: овоцит ІІ і полярне тільце. Ссавцям також властиве явище вибірковості овоцитів, яке полягає в овуляції одного або декількох зрілих овоцитів під час статевої зрілості за дії гормонів.
 Механізми росту Затримка завершення мейозу майже до кінця дозрівання яйцеклітини, тобто яйцеклітина має тетраплоїдний набір хромосом протягом більшої частини періоду свого росту. Яйцеклітина зберігає на цей період материнську і батьківську копії кожного гену, чим запобігає прояву рецесивних летальних мутацій. В деяких яйцеклітинах утворюються додаткові копії окремих генів.
Механізми росту Затримка завершення мейозу майже до кінця дозрівання яйцеклітини, тобто яйцеклітина має тетраплоїдний набір хромосом протягом більшої частини періоду свого росту. Яйцеклітина зберігає на цей період материнську і батьківську копії кожного гену, чим запобігає прояву рецесивних летальних мутацій. В деяких яйцеклітинах утворюються додаткові копії окремих генів.
 В оогенезі в ооцитах накопичуються гістони, що забезпечує майбутню реплікацію ДНК при дробленні. В соматичних клітинах гістони не накопичуються, вони синтезуються сурово координовано з реплікацією ДНК.
В оогенезі в ооцитах накопичуються гістони, що забезпечує майбутню реплікацію ДНК при дробленні. В соматичних клітинах гістони не накопичуються, вони синтезуються сурово координовано з реплікацією ДНК.
 Таким чином яйцеклітина містить весь апарат трансляції, структурні білки, резерв багатьох ферментів, необхідних для ранніх стадій розвитку: ферменти синтезу ДНК і РНК (ДНК- іРНК-полімерази), ферменти для синтезу дезоксинуклеозидтрифосфатів (фосфатази, редуктази). Є фактори, які виконують різноманітні регуляторні функції (необхідні для активації запасених ферментів, рибосом, іРНК, т. РНК, процесів синтезу ДНК і РНК, збирання нуклеосом і інше)
Таким чином яйцеклітина містить весь апарат трансляції, структурні білки, резерв багатьох ферментів, необхідних для ранніх стадій розвитку: ферменти синтезу ДНК і РНК (ДНК- іРНК-полімерази), ферменти для синтезу дезоксинуклеозидтрифосфатів (фосфатази, редуктази). Є фактори, які виконують різноманітні регуляторні функції (необхідні для активації запасених ферментів, рибосом, іРНК, т. РНК, процесів синтезу ДНК і РНК, збирання нуклеосом і інше)
 фолікул") Водночас з овогенезом у яєчниках відбувається розвиток фолікулів – фолікулогенез: примордіальний (первісний) фолікул → первинний (преантральний) фолікул → вторинний (антральний) фолікул → третинний (зрілий, Граафів пухирець) фолікул → атретичний фолікул → жовте тіло → білуваті тіла
Водночас з овогенезом у яєчниках відбувається розвиток фолікулів – фолікулогенез: примордіальний (первісний) фолікул → первинний (преантральний) фолікул → вторинний (антральний) фолікул → третинний (зрілий, Граафів пухирець) фолікул → атретичний фолікул → жовте тіло → білуваті тіла
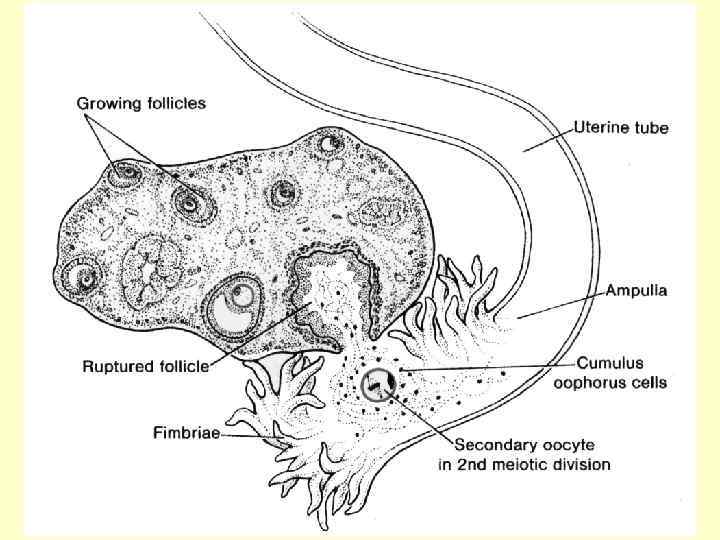
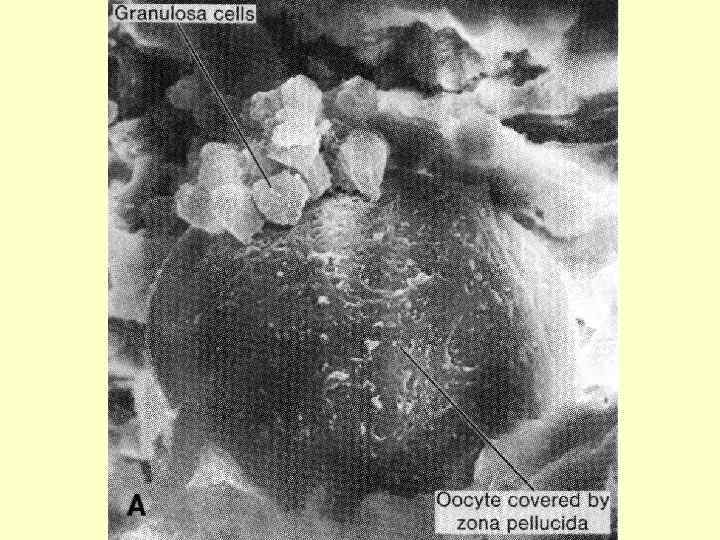
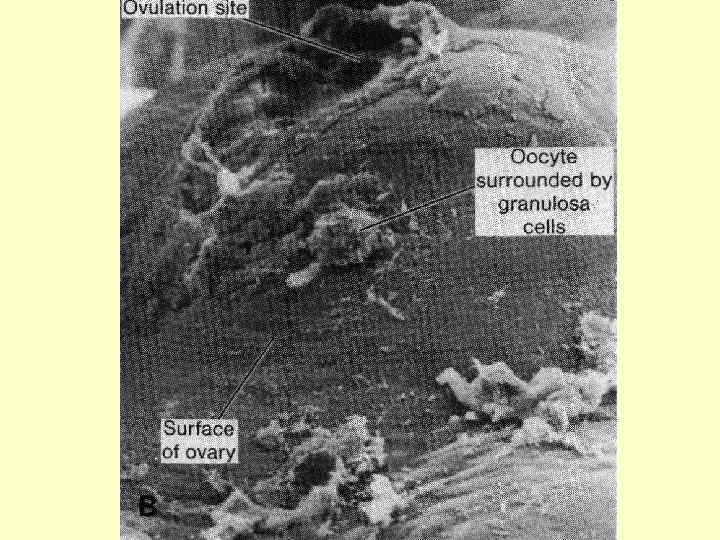
 У людини формування овоцитів другого порядку відбувається поетапно у фолікулах, які носять назву первинного, вторинного й третинного (Граафів пухирець): на стадії третинного фолікула який вже містить в собі овоцит другого порядку, відбувається овуляція. Зі сформованого овоциту другого порядку утворюється зріла гаплоїдна яйцеклітина (та друге редукційне тільце) під час другого поділу і вже у маточних трубах.
У людини формування овоцитів другого порядку відбувається поетапно у фолікулах, які носять назву первинного, вторинного й третинного (Граафів пухирець): на стадії третинного фолікула який вже містить в собі овоцит другого порядку, відбувається овуляція. Зі сформованого овоциту другого порядку утворюється зріла гаплоїдна яйцеклітина (та друге редукційне тільце) під час другого поділу і вже у маточних трубах.
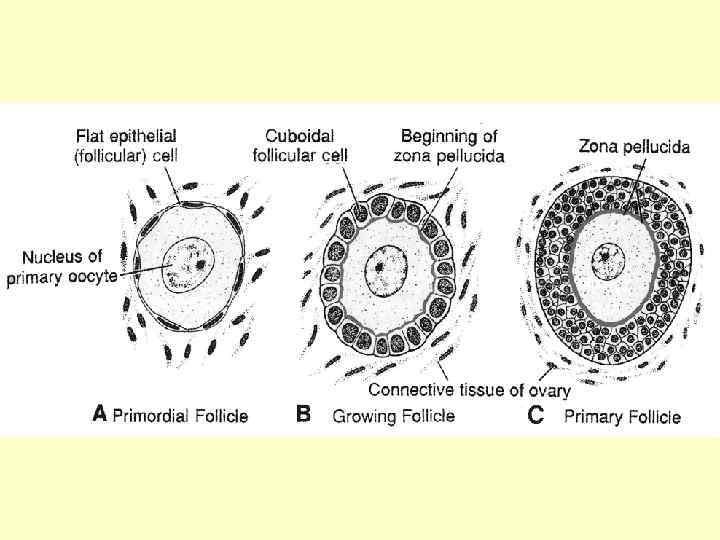
 Примордіальні фолікули залягають безпосередньо у корковій речовині в декілька рядів під білковою капсулою, яка зверху вкрита зачатковим кубічним епітелієм. Примордіальні фолікули називають первісними, оскільки вони з’являються в зачатках яєчників найпершими. Їхня кількість становить при народженні дівчинки близько 1 млн. , тоді як до статевого дозрівання залишається близько 100 тис. У порожнині цього фолікула знаходиться овоцит I порядку, що перебуває на стадії діакінезу профази першого мейотичного поділу.
Примордіальні фолікули залягають безпосередньо у корковій речовині в декілька рядів під білковою капсулою, яка зверху вкрита зачатковим кубічним епітелієм. Примордіальні фолікули називають первісними, оскільки вони з’являються в зачатках яєчників найпершими. Їхня кількість становить при народженні дівчинки близько 1 млн. , тоді як до статевого дозрівання залишається близько 100 тис. У порожнині цього фолікула знаходиться овоцит I порядку, що перебуває на стадії діакінезу профази першого мейотичного поділу.

, який відрізняється кількома (1 -2 -4)") Наступним етапом є утворення первинного фолікулу (преантрального фолікулу), який відрізняється кількома (1 -2 -4) рядами кубічних або призматичних фолікулярних клітин. Він ще не має порожнини, проте навколо нього вже з’являються поодинокі клітини сполучнотканинної оболонки – теки (thecae folliculi). Саме цей фолікул вступає у фазу росту й досягає розміру 150200 мкм.
Наступним етапом є утворення первинного фолікулу (преантрального фолікулу), який відрізняється кількома (1 -2 -4) рядами кубічних або призматичних фолікулярних клітин. Він ще не має порожнини, проте навколо нього вже з’являються поодинокі клітини сполучнотканинної оболонки – теки (thecae folliculi). Саме цей фолікул вступає у фазу росту й досягає розміру 150200 мкм.
 фолікулу, проте у") ФСГ стимулює ріст і примордіального, і первинного (наступного за ним) фолікулу, проте у первинному фолікулі фолікулярних клітин (а це єдині клітини, що мають рецептори до ФСГ) більше, ніж у примордіальному, а значить і "доза" захопленого ФСГ є вищою, і, як наслідок, шансів досягти піку розвитку більше. Цікаво, що ріст примордіального фолікулу до первинного триває 85 днів, а значить, фолікул, який дозріває в даному менструальному циклі, починає свій ріст як мінімум за два цикли до цього.
ФСГ стимулює ріст і примордіального, і первинного (наступного за ним) фолікулу, проте у первинному фолікулі фолікулярних клітин (а це єдині клітини, що мають рецептори до ФСГ) більше, ніж у примордіальному, а значить і "доза" захопленого ФСГ є вищою, і, як наслідок, шансів досягти піку розвитку більше. Цікаво, що ріст примордіального фолікулу до первинного триває 85 днів, а значить, фолікул, який дозріває в даному менструальному циклі, починає свій ріст як мінімум за два цикли до цього.
 З моменту утворення навколо овоциту першого порядку вторинної оболонки, що складається з двох шарів фолікулярних клітин, закінчується фаза малого росту (превітелогенез), і починається фаза великого росту (вітелогенез), яка супроводжується інтенсивним нагромадженням жовтка. Вітелогенез супроводжується змінами у "комунікаційних відносинах" між овоцитом I порядку й фолікулярним епітелієм, що призводить до появи первинної оболонки (вона складається з двох зон: блискуча зона (оболонка) - zona pellucida й променистий вінець (corona rаdiata).
З моменту утворення навколо овоциту першого порядку вторинної оболонки, що складається з двох шарів фолікулярних клітин, закінчується фаза малого росту (превітелогенез), і починається фаза великого росту (вітелогенез), яка супроводжується інтенсивним нагромадженням жовтка. Вітелогенез супроводжується змінами у "комунікаційних відносинах" між овоцитом I порядку й фолікулярним епітелієм, що призводить до появи первинної оболонки (вона складається з двох зон: блискуча зона (оболонка) - zona pellucida й променистий вінець (corona rаdiata).
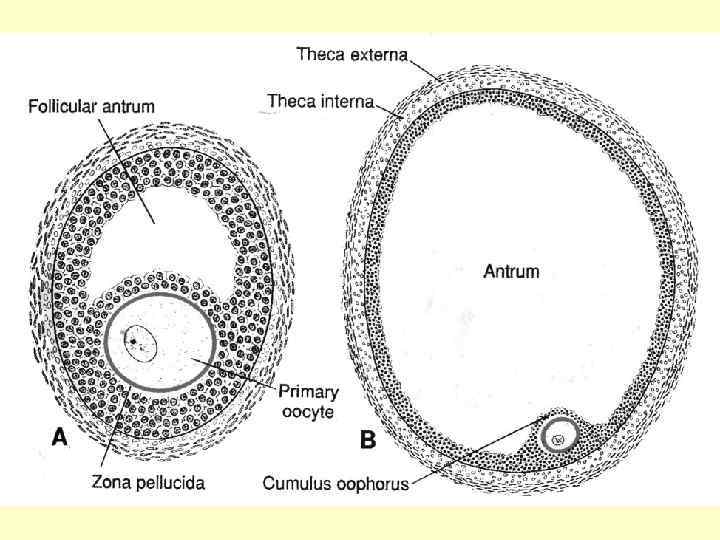
 і його розмір досягає 500 мкм") Коли фолікул стає багатошаровим (більше п’яти шарів клітин) і його розмір досягає 500 мкм у діаметрі - вторинний або антральний. Утворення порожнини є процесом поступовим: спочатку в товщі шарів фолікулярних клітин утворюється невелика щілина, потім невелика порожнина, що заповнюється фолікулярною рідиною, яка розтягує сам фолікул. Збільшення кількості фолікулярних клітин є наслідком їхнього розмноження під впливом ФСГ. Більш того, саме ФСГ стимулює секрецію фолікулярними клітинами глікозаміногліканів у порожнину фолікула.
Коли фолікул стає багатошаровим (більше п’яти шарів клітин) і його розмір досягає 500 мкм у діаметрі - вторинний або антральний. Утворення порожнини є процесом поступовим: спочатку в товщі шарів фолікулярних клітин утворюється невелика щілина, потім невелика порожнина, що заповнюється фолікулярною рідиною, яка розтягує сам фолікул. Збільшення кількості фолікулярних клітин є наслідком їхнього розмноження під впливом ФСГ. Більш того, саме ФСГ стимулює секрецію фолікулярними клітинами глікозаміногліканів у порожнину фолікула.
 У порожнині вторинного фолікула міститься овоцит I порядку, що перебуває на стадії першого мейотичного поділу. Навколо ж самого фолікулу формується сполучнотканинна оболонка (thecae folliculi), яку поділяють на зовнішню theca externa, що складається з волокнистої сполучної тканини, і внутрішню theca interna, яка сильно васкуляризована й утворена гормоносинтезуючими клітинами (вона, як і фолікулярні клітини, приймає участь в утворенні фолікулярної рідини). І ендокриноцити теки (під впливом ЛГ), і фолікулярні клітини (ФСГ) синтезують жіночий гормон естрадіол.
У порожнині вторинного фолікула міститься овоцит I порядку, що перебуває на стадії першого мейотичного поділу. Навколо ж самого фолікулу формується сполучнотканинна оболонка (thecae folliculi), яку поділяють на зовнішню theca externa, що складається з волокнистої сполучної тканини, і внутрішню theca interna, яка сильно васкуляризована й утворена гормоносинтезуючими клітинами (вона, як і фолікулярні клітини, приймає участь в утворенні фолікулярної рідини). І ендокриноцити теки (під впливом ЛГ), і фолікулярні клітини (ФСГ) синтезують жіночий гормон естрадіол.
, що він набуває велетенського") Фолікулярна рідина розтягує фолікул настільки (витончуючи при цьому його стінки), що він набуває велетенського розміру (у людини він досягає 20 мм). Кількість фолікулярної рідини у такому фолікулі в 100 разів більше, ніж в антральному. Такий фолікул називають пухирчастим, третинним (зрілим), або Граафовим пухирцем (преовуляторним фолікулом). У порожнину Граафова пухирця (рідина в якій містить естрадіол) обернено яйценосний горбик, утворений овоцитом першого - другого порядку, оточеним 1 -3 шарами фолікулярних клітин - променистим вінцем.
Фолікулярна рідина розтягує фолікул настільки (витончуючи при цьому його стінки), що він набуває велетенського розміру (у людини він досягає 20 мм). Кількість фолікулярної рідини у такому фолікулі в 100 разів більше, ніж в антральному. Такий фолікул називають пухирчастим, третинним (зрілим), або Граафовим пухирцем (преовуляторним фолікулом). У порожнину Граафова пухирця (рідина в якій містить естрадіол) обернено яйценосний горбик, утворений овоцитом першого - другого порядку, оточеним 1 -3 шарами фолікулярних клітин - променистим вінцем.

 Приблизно 1 зі 100 фолікулів, що розвиваються, здійснює повний розвиток, інші ж піддаються атрезії, причини якого до кінця ще не вивчені. При цьому утворюються атретичні фолікули (тіла), які у подальшому інволюють. Такі атретичні фолікули інтенсивно іннервовані. Їхня поява є не просто атрофією фолікулу, а його функціональною перебудовою. Атретичні тіла мають в центрі залишки зморщеної блискучої оболонки, оточеної сполучною тканиною, у складі якої наявні численні гіпертрофовані інтерстиціальні (гормоносинтезуючі) клітини. Ці тіла функціонують недовго, а після інволюції на їхньому місці лишається сполучна тканина й скупчення інтерстиціальних клітин.
Приблизно 1 зі 100 фолікулів, що розвиваються, здійснює повний розвиток, інші ж піддаються атрезії, причини якого до кінця ще не вивчені. При цьому утворюються атретичні фолікули (тіла), які у подальшому інволюють. Такі атретичні фолікули інтенсивно іннервовані. Їхня поява є не просто атрофією фолікулу, а його функціональною перебудовою. Атретичні тіла мають в центрі залишки зморщеної блискучої оболонки, оточеної сполучною тканиною, у складі якої наявні численні гіпертрофовані інтерстиціальні (гормоносинтезуючі) клітини. Ці тіла функціонують недовго, а після інволюції на їхньому місці лишається сполучна тканина й скупчення інтерстиціальних клітин.
 На стадії розквіту клітини жовтого тіла синтезують прогестерон, естроген, невелику кількість андрогену, окситоцину, інгібіну, а при настанні вагітності й релаксину. Підвищення рівня прогестерону призводить до зниження концентрації ЛГ.
На стадії розквіту клітини жовтого тіла синтезують прогестерон, естроген, невелику кількість андрогену, окситоцину, інгібіну, а при настанні вагітності й релаксину. Підвищення рівня прогестерону призводить до зниження концентрації ЛГ.
 Регуляція овогенезу знаходиться під контролем гормонів гіпоталамо-гіпофізарно-нейросекреторної системи. У першу чергу, це два гонадотропних гормони аденогіпофізу ЛГ і ФСГ, які у ссавців і людини виробляються циклічно: у першу половину місяця спостерігається пік секреції ФСГ, у другу – ЛГ. При сперматогенезі секреція ЛГ і ФСГ відбувається за тонічним принципом, тобто не залежить від днів місяця. Процеси фолікулогенезу стимулює ФСГ. Для того ж, щоб у антральному фолікулі почався синтез естрадіолу необхідний додатковий вплив ЛГ: ФСГ посилює синтез і секрецію естрогенів фолікулярними клітинами яєчника, а ЛГ здійснює аналогічний вплив на гормоносинтезуючі клітини теки.
Регуляція овогенезу знаходиться під контролем гормонів гіпоталамо-гіпофізарно-нейросекреторної системи. У першу чергу, це два гонадотропних гормони аденогіпофізу ЛГ і ФСГ, які у ссавців і людини виробляються циклічно: у першу половину місяця спостерігається пік секреції ФСГ, у другу – ЛГ. При сперматогенезі секреція ЛГ і ФСГ відбувається за тонічним принципом, тобто не залежить від днів місяця. Процеси фолікулогенезу стимулює ФСГ. Для того ж, щоб у антральному фолікулі почався синтез естрадіолу необхідний додатковий вплив ЛГ: ФСГ посилює синтез і секрецію естрогенів фолікулярними клітинами яєчника, а ЛГ здійснює аналогічний вплив на гормоносинтезуючі клітини теки.
 СПЕРМАТОГЕНЕЗ
СПЕРМАТОГЕНЕЗ
 Схема періодів сперматогенезу
Схема періодів сперматогенезу
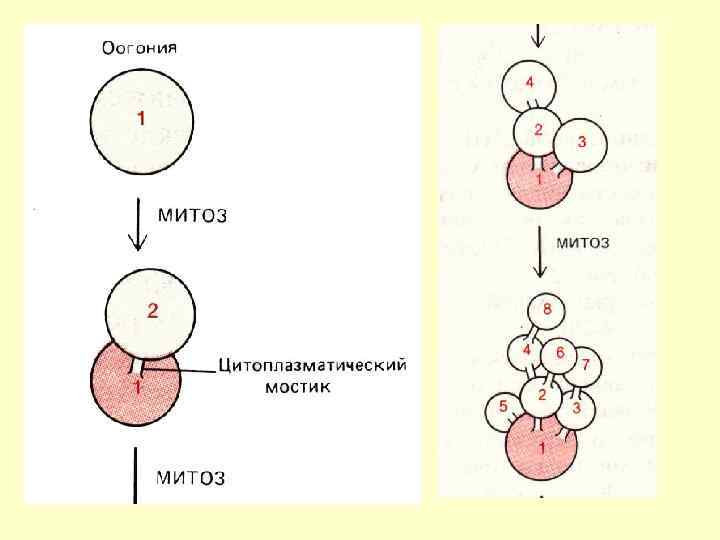
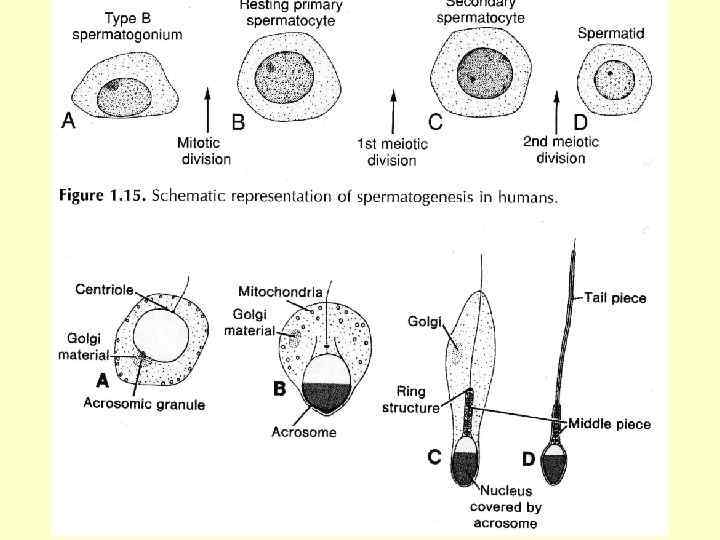
 Клітини типу А темні - резервні, власне стовбурові клітини, мають тривалий клітинний цикл і рідко вступають у мітоз. Мітотична активність їхня росте зі зменшенням загальної кількості сперматогоній. Клітини типу А світлі - напівстовбурові клітини, які знаходяться в постійно змінних клітинних циклах. У результаті мітотичного поділу такої клітини формуються або дві клітини типу Б, або одна Б і одна А (світла клітина);
Клітини типу А темні - резервні, власне стовбурові клітини, мають тривалий клітинний цикл і рідко вступають у мітоз. Мітотична активність їхня росте зі зменшенням загальної кількості сперматогоній. Клітини типу А світлі - напівстовбурові клітини, які знаходяться в постійно змінних клітинних циклах. У результаті мітотичного поділу такої клітини формуються або дві клітини типу Б, або одна Б і одна А (світла клітина);
 При мітотичному поділі клітин Б не відбувається повного цитокінезу, дочірні клітини залишаються зв'язаними між собою цитоплазматичними містками: формується синцитій. У подальшому, клітини, що диференціюються, залишаються зв'язаними одна з одною та у складі синцитіального клону переміщуються від базального шару звивистого сім'яного канальця до його просвіту. Мітотичний поділ сперматогоній Б останньої генерації призводить до утворення передлептотенних первинних сперматоцитів.
При мітотичному поділі клітин Б не відбувається повного цитокінезу, дочірні клітини залишаються зв'язаними між собою цитоплазматичними містками: формується синцитій. У подальшому, клітини, що диференціюються, залишаються зв'язаними одна з одною та у складі синцитіального клону переміщуються від базального шару звивистого сім'яного канальця до його просвіту. Мітотичний поділ сперматогоній Б останньої генерації призводить до утворення передлептотенних первинних сперматоцитів.

 Період росту В сперматоцитах в основному синтезуються м. РНК з генів, продукти яких необхідні для забезпечення руху спермія або для зв'язування його з яйцеклітиною. Це гени тубуліну і біндину. Біндин накопичується в міхурцях, які зливаються з акросомою, і забезпечує видоспецифічність запліднення. Тубулін бере участь в утворенні джгутика сперматозоїда та веретена поділу. Якщо інгібувати їхній синтез – мейоз блокується. Період короткотривалий.
Період росту В сперматоцитах в основному синтезуються м. РНК з генів, продукти яких необхідні для забезпечення руху спермія або для зв'язування його з яйцеклітиною. Це гени тубуліну і біндину. Біндин накопичується в міхурцях, які зливаються з акросомою, і забезпечує видоспецифічність запліднення. Тубулін бере участь в утворенні джгутика сперматозоїда та веретена поділу. Якщо інгібувати їхній синтез – мейоз блокується. Період короткотривалий.

 Період формування 1. Формування акросоми. З апарату Гольджі формується органела, оточена мембраною, в якій містяться вуглеводи і ферменти лізосомальної природи: - акрозин –(руйнує прозору оболонку овоциту); - пенетраза (дисоціює клітини променистого вінця); - гіалуронідаза (розщеплює гіалуронову кислоту); - кисла фосфатаза ( руйнує форсхолін при проходженні сперміїв через плазмолему овоциту), а також багато аутоантигенів.
Період формування 1. Формування акросоми. З апарату Гольджі формується органела, оточена мембраною, в якій містяться вуглеводи і ферменти лізосомальної природи: - акрозин –(руйнує прозору оболонку овоциту); - пенетраза (дисоціює клітини променистого вінця); - гіалуронідаза (розщеплює гіалуронову кислоту); - кисла фосфатаза ( руйнує форсхолін при проходженні сперміїв через плазмолему овоциту), а також багато аутоантигенів.
 2. Формування ядра. 3. Формування шийки. Кількість витків спіралі мітохондрії може бути різною. Напр. , у ссавців нараховують до 15 -ти витків, хоча у гризунів їх може бути до 300. 4. Формування хвоста.
2. Формування ядра. 3. Формування шийки. Кількість витків спіралі мітохондрії може бути різною. Напр. , у ссавців нараховують до 15 -ти витків, хоча у гризунів їх може бути до 300. 4. Формування хвоста.
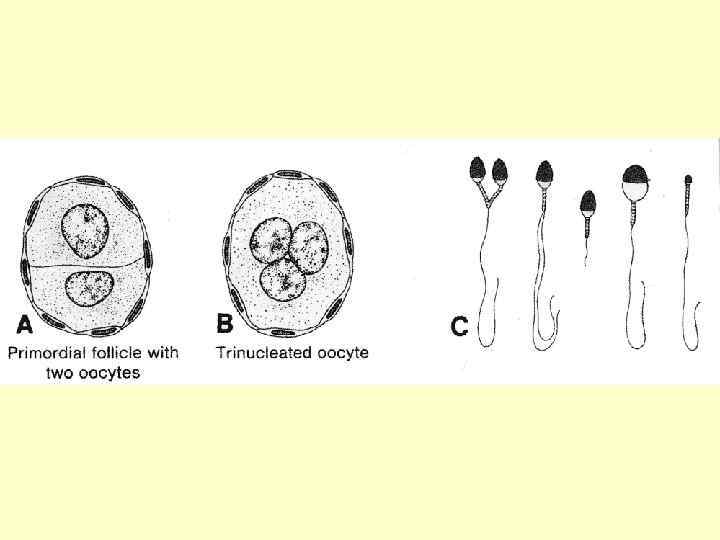
 1 -людина, 2 -тритон, 3 -річковий рак, 4 -морська свинка, 5 -свиня, 6 –бик, 7 -півень, 8 -гіллястовусий рак, 9 -десятиногий рак, 10 -кінська аскарида, 11 - гострик
1 -людина, 2 -тритон, 3 -річковий рак, 4 -морська свинка, 5 -свиня, 6 –бик, 7 -півень, 8 -гіллястовусий рак, 9 -десятиногий рак, 10 -кінська аскарида, 11 - гострик
 Як можна відрізнити сперматозоїди, що несуть Х-хромосому, від сперматозоїдів з Yхромосомою? За допомогою біохімічних досліджень. Наприклад, за визначенням молекулярної маси ДНК: у сперматозоїдів з Х-хромосомою на 3 -4% більше ДНК; за електрофоретичною рухливістю: сперматозоїди з Х-хромосомою швидше рухаються до анода; за допомогою центрифугування: сперматозоїди з Ххромосомою осаджуються у важкій фракції; за допомогою метода проточної цитометрії, тобто вимірювання хромосомного вмісту. Але в кожному випадку не має стовідсоткової гарантії і не завжди результати є відтворюваними
Як можна відрізнити сперматозоїди, що несуть Х-хромосому, від сперматозоїдів з Yхромосомою? За допомогою біохімічних досліджень. Наприклад, за визначенням молекулярної маси ДНК: у сперматозоїдів з Х-хромосомою на 3 -4% більше ДНК; за електрофоретичною рухливістю: сперматозоїди з Х-хромосомою швидше рухаються до анода; за допомогою центрифугування: сперматозоїди з Ххромосомою осаджуються у важкій фракції; за допомогою метода проточної цитометрії, тобто вимірювання хромосомного вмісту. Але в кожному випадку не має стовідсоткової гарантії і не завжди результати є відтворюваними
; –") Функції клітин Сертолі – трофічна (доставка кисню та поживних речовин для клітин сперматогенезу); – фагоцитарна (фагоцитують фрагменти сперматид та сперматозоїдів, значна частини яких гине шляхом апоптозу); – участь у формуванні гематотестикулярного бар’єру; – ендокринна; продукують гормони інгібін і β-естрадіол 17 (β 17), які спрямовані до гіпофізу і клітин Лейдіга, відповідно. – вироблення рідини сім’яних канальців.
Функції клітин Сертолі – трофічна (доставка кисню та поживних речовин для клітин сперматогенезу); – фагоцитарна (фагоцитують фрагменти сперматид та сперматозоїдів, значна частини яких гине шляхом апоптозу); – участь у формуванні гематотестикулярного бар’єру; – ендокринна; продукують гормони інгібін і β-естрадіол 17 (β 17), які спрямовані до гіпофізу і клітин Лейдіга, відповідно. – вироблення рідини сім’яних канальців.
 ГТБ підтримує температуру в сім’янику на 4 -5 Сº нижчу від температури тіла; зберігає внутрішнє середовище від проникнення шкідливих речовин. ГТБ є проникним для води, алкоголю, сечовини та деяких інших речовин, але водночас захищає геном від вітальних та акридинових барвників, ізолює аутоантигени від імунного апарату організму. ГТБ відповідає за гормональну регуляцію сперматогенезу
ГТБ підтримує температуру в сім’янику на 4 -5 Сº нижчу від температури тіла; зберігає внутрішнє середовище від проникнення шкідливих речовин. ГТБ є проникним для води, алкоголю, сечовини та деяких інших речовин, але водночас захищає геном від вітальних та акридинових барвників, ізолює аутоантигени від імунного апарату організму. ГТБ відповідає за гормональну регуляцію сперматогенезу
 При хронічному вживанні алкоголю у чоловіків розвивається найважча форма репродуктивного порушення – азооспермія (відсутність у спермі сперматозоїдів). Гістологічний аналіз тканини сім’яників у таких чоловіків виявив, що причина порушення полягає у блокуванні дозрівання клітин на стадії пахітени. Процеси сперматогенезу можуть поступово відновлюватися після припинення вживання алкоголю; нормальні показники сперми спостерігаються не раніше, ніж через три місяці.
При хронічному вживанні алкоголю у чоловіків розвивається найважча форма репродуктивного порушення – азооспермія (відсутність у спермі сперматозоїдів). Гістологічний аналіз тканини сім’яників у таких чоловіків виявив, що причина порушення полягає у блокуванні дозрівання клітин на стадії пахітени. Процеси сперматогенезу можуть поступово відновлюватися після припинення вживання алкоголю; нормальні показники сперми спостерігаються не раніше, ніж через три місяці.
 Аутоантигени – це гліко- або ліпопротеїди, що входять до складу чоловічих статевих клітин вищих тварин і людини та сприймаються клітинами імунної системи як чужорідні. Це зумовлено тим, що формування статевих клітин починається лише після досягнення статевої зрілості, коли імунна система вже сформована. Тому необхідна система захисту від імунізації, якщо їй не запобігти, то виникає аутоімунний орхіт (ураження сім'яника), який веде до інфільтрації тканини і порушення сперміогенезу.
Аутоантигени – це гліко- або ліпопротеїди, що входять до складу чоловічих статевих клітин вищих тварин і людини та сприймаються клітинами імунної системи як чужорідні. Це зумовлено тим, що формування статевих клітин починається лише після досягнення статевої зрілості, коли імунна система вже сформована. Тому необхідна система захисту від імунізації, якщо їй не запобігти, то виникає аутоімунний орхіт (ураження сім'яника), який веде до інфільтрації тканини і порушення сперміогенезу.
 Між звивистими сім’яними канальцями знаходиться стромальна сполучна тканина з розташованими в ній клітинами Лейдіга (ендокриноцитами). Клітини Лейдіга мають рецептори до ЛГ аденогіпофіза, під впливом якого синтезують гормон тестостерон, який необхідний як для процесу сперматогенезу, так і для розвитку вторинно-статевих ознак за чоловічим типом (важкий скелет, розвинені м’язи, низький голос, ріст волосся на обличчі й тілі за чоловічим типом, посилений еритропоез і потовиділення, статева поведінка тощо).
Між звивистими сім’яними канальцями знаходиться стромальна сполучна тканина з розташованими в ній клітинами Лейдіга (ендокриноцитами). Клітини Лейдіга мають рецептори до ЛГ аденогіпофіза, під впливом якого синтезують гормон тестостерон, який необхідний як для процесу сперматогенезу, так і для розвитку вторинно-статевих ознак за чоловічим типом (важкий скелет, розвинені м’язи, низький голос, ріст волосся на обличчі й тілі за чоловічим типом, посилений еритропоез і потовиділення, статева поведінка тощо).
 Клітини Сертолі - антимюлерівський гормон, який трансформує гоноцити у сперматогонії та до статевого дозрівання пригнічує мейоз статевих клітин. Клітини Лейдіга - інгібіни й тестостерон забезпечує диференціювання не лише гонади, але й придаткових залоз і зовнішніх статевих органів. У відсутності такої стимуляції закладання гонади здійснюється автономно за жіночим типом, під впливом гормонів плаценти й материнських гормонів.
Клітини Сертолі - антимюлерівський гормон, який трансформує гоноцити у сперматогонії та до статевого дозрівання пригнічує мейоз статевих клітин. Клітини Лейдіга - інгібіни й тестостерон забезпечує диференціювання не лише гонади, але й придаткових залоз і зовнішніх статевих органів. У відсутності такої стимуляції закладання гонади здійснюється автономно за жіночим типом, під впливом гормонів плаценти й материнських гормонів.
 ДЯКУЮ ЗА УВАГУ!
ДЯКУЮ ЗА УВАГУ!