

Лекция_СибГМУ.pptx
- Количество слайдов: 57
 Функциональная морфология мозжечка
Функциональная морфология мозжечка
 Мозжечок
Мозжечок
 Основные отделы мозжечка
Основные отделы мозжечка
 Ядра мозжечка 1. 2. 3. 4. 5. 6. Ядро шатра Шаровидное ядро Пробковидное ядро Зубчатое ядро Червь мозжечка Полушария мозжечка
Ядра мозжечка 1. 2. 3. 4. 5. 6. Ядро шатра Шаровидное ядро Пробковидное ядро Зубчатое ядро Червь мозжечка Полушария мозжечка
 Строение мозжечка
Строение мозжечка
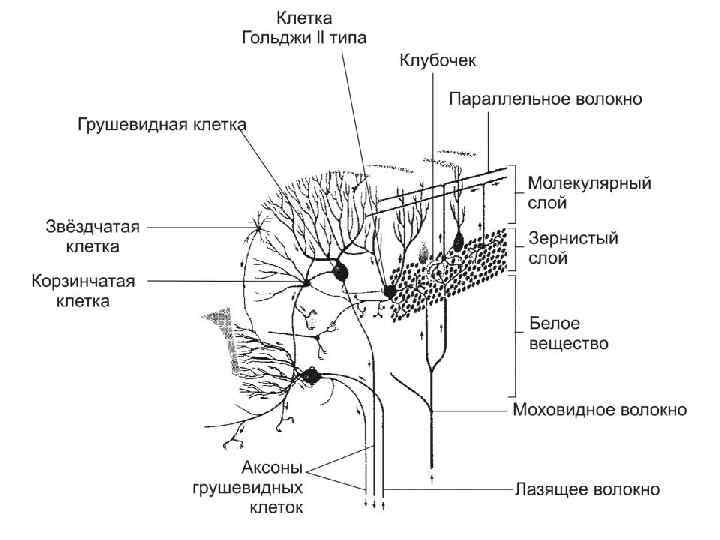
; 2 — ганглионарный (грушевидные клетки") Строение мозжечка 1 — молекулярный (звездчатые и корзинчатые клетки); 2 — ганглионарный (грушевидные клетки Пуркинье); 3 — зернистый (клетки-зерна, клетки Гольджи и веретеновидные клетки). 4 — белое вещество мозжечка; образовано миелиновыми нервными волокнами: — одни волокна (афферентные) идут к коре мозжечка от спинного и продолговатого мозга; — другие (эфферентные) — от коры мозжечка к его подкорковым ядрам.
Строение мозжечка 1 — молекулярный (звездчатые и корзинчатые клетки); 2 — ганглионарный (грушевидные клетки Пуркинье); 3 — зернистый (клетки-зерна, клетки Гольджи и веретеновидные клетки). 4 — белое вещество мозжечка; образовано миелиновыми нервными волокнами: — одни волокна (афферентные) идут к коре мозжечка от спинного и продолговатого мозга; — другие (эфферентные) — от коры мозжечка к его подкорковым ядрам.
 Мозжечок. Участок коры
Мозжечок. Участок коры
 Молекулярный слой Звездчатые клетки: Корзинчатые клетки: дендритами связаны с параллельными волокнами молекулярного слоя аксонами охватывают тела клеток Пуркинье, формируя корзинки вокруг них и тормозные синапсы Короткоаксонные: связаны с параллельными волокнами, аксоны формируют тормозные синапсы на дендритах клеток Пуркинье Лежат выше тел корзинчатых клеток Длинноаксонные: имеют аксон, участвующий в образовании корзинки вокруг клеток Пуркинье
Молекулярный слой Звездчатые клетки: Корзинчатые клетки: дендритами связаны с параллельными волокнами молекулярного слоя аксонами охватывают тела клеток Пуркинье, формируя корзинки вокруг них и тормозные синапсы Короткоаксонные: связаны с параллельными волокнами, аксоны формируют тормозные синапсы на дендритах клеток Пуркинье Лежат выше тел корзинчатых клеток Длинноаксонные: имеют аксон, участвующий в образовании корзинки вокруг клеток Пуркинье
 Ганглионарный слой Содержит клетки Пуркинье – грушевидные нейроны, лежащие в один ряд, оплетенные коллатералями корзинчатых клеток
Ганглионарный слой Содержит клетки Пуркинье – грушевидные нейроны, лежащие в один ряд, оплетенные коллатералями корзинчатых клеток
 Клетка Пуркинье Аксон – отходит от основания клетки, – миелинизируется, – идет через зернистый слой, – образует тормозные синапсы на телах соседних клеток Пуркинье, – тормозные синапсы на клетках Гольджи – заканчивается на ядрах мозжечка 2 -3 дендрита имеют множество шипиков, на которых находятся – возбуждающие синапсы с параллельными волокнами (аксонами клеток-зерен), - тормозные синапсы с лазящими волокнами
Клетка Пуркинье Аксон – отходит от основания клетки, – миелинизируется, – идет через зернистый слой, – образует тормозные синапсы на телах соседних клеток Пуркинье, – тормозные синапсы на клетках Гольджи – заканчивается на ядрах мозжечка 2 -3 дендрита имеют множество шипиков, на которых находятся – возбуждающие синапсы с параллельными волокнами (аксонами клеток-зерен), - тормозные синапсы с лазящими волокнами
 Зернистый слой Клетки-зерна: 1. многочисленные 2. дендриты – «птичьи лапки» 3. Аксоны – т-образно делятся, образуя параллельные волокна молекулярного слоя 4. Образуют возбуждающие синапсы на дендритах клеток Пуркинье, корзинчатых и звездчатых клеток Клубочки мозжечка – контактные зоны между моховидными волокнами, дендритами клеток-зерен и аксонами клеток Гольджи Большие клетки-зерна (клетки Гольджи) Аксоны – в клубочках связаны с дендритами клеток-зерен Дендриты – связаны с параллельными волокнами. Угнетают деятельность клетокзерен 3 вида: 1. Звездчатые с коротким нейритом 2. Звездчатые с длинным нейритом 3. Горизонтальные веретенообразные
Зернистый слой Клетки-зерна: 1. многочисленные 2. дендриты – «птичьи лапки» 3. Аксоны – т-образно делятся, образуя параллельные волокна молекулярного слоя 4. Образуют возбуждающие синапсы на дендритах клеток Пуркинье, корзинчатых и звездчатых клеток Клубочки мозжечка – контактные зоны между моховидными волокнами, дендритами клеток-зерен и аксонами клеток Гольджи Большие клетки-зерна (клетки Гольджи) Аксоны – в клубочках связаны с дендритами клеток-зерен Дендриты – связаны с параллельными волокнами. Угнетают деятельность клетокзерен 3 вида: 1. Звездчатые с коротким нейритом 2. Звездчатые с длинным нейритом 3. Горизонтальные веретенообразные
 волокна: идут в составе оливомозжечкового и мостомозжечкового путей") Волокна мозжечка • • Моховидные (мшистые) волокна: идут в составе оливомозжечкового и мостомозжечкового путей опосредованно (через клетки-зерна) оказывают на грушевидные клетки возбуждающее действие. заканчиваются в клубочках (glomerulus) зернистого слоя мозжечка, где вступают в контакт с дендритами клетокзерен. Каждое волокно дает ветви ко многим клубочкам мозжечка, и каждый клубочек получает ветви от многих моховидных волокон. Аксоны клеток-зерен по параллельным волокнам молекулярного слоя передают импульс дендритам грушевидных, корзинчатых, звездчатых нейронов, больших звездчатых нейронов зернистого слоя.
Волокна мозжечка • • Моховидные (мшистые) волокна: идут в составе оливомозжечкового и мостомозжечкового путей опосредованно (через клетки-зерна) оказывают на грушевидные клетки возбуждающее действие. заканчиваются в клубочках (glomerulus) зернистого слоя мозжечка, где вступают в контакт с дендритами клетокзерен. Каждое волокно дает ветви ко многим клубочкам мозжечка, и каждый клубочек получает ветви от многих моховидных волокон. Аксоны клеток-зерен по параллельным волокнам молекулярного слоя передают импульс дендритам грушевидных, корзинчатых, звездчатых нейронов, больших звездчатых нейронов зернистого слоя.
 Волокна мозжечка Лазящие волокна: • поступают в кору мозжечка, по-видимому, по спинно-мозжечковому и вестибуломозжечковому путям • пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности синапсами • лазящие волокна передают возбуждение непосредственно грушевидным нейронам
Волокна мозжечка Лазящие волокна: • поступают в кору мозжечка, по-видимому, по спинно-мозжечковому и вестибуломозжечковому путям • пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности синапсами • лазящие волокна передают возбуждение непосредственно грушевидным нейронам
 Клубочки мозжечка • скопление терминальных ветвлений отростков различных нейронов мозжечка и моховидных волокон • клубочек окружён капсулой из глиальных клеток • вокруг клубочка расположены скопления клеток-зёрен
Клубочки мозжечка • скопление терминальных ветвлений отростков различных нейронов мозжечка и моховидных волокон • клубочек окружён капсулой из глиальных клеток • вокруг клубочка расположены скопления клеток-зёрен
 Клубочки содержат • розетки - окончания моховидных волокон, • птичьи лапки - терминали дендритов клеток -зёрен, • аксоны клеток Гольджи (формируют аксональные синапсы) • часть разветвлений дендритов клеток Гольджи.
Клубочки содержат • розетки - окончания моховидных волокон, • птичьи лапки - терминали дендритов клеток -зёрен, • аксоны клеток Гольджи (формируют аксональные синапсы) • часть разветвлений дендритов клеток Гольджи.
 Клубочек 1. Аксон большого звездчатого нейрона 2. Дендриты клеток-зерен 3. Моховидные волокна 4. Синаптические пузырьки
Клубочек 1. Аксон большого звездчатого нейрона 2. Дендриты клеток-зерен 3. Моховидные волокна 4. Синаптические пузырьки
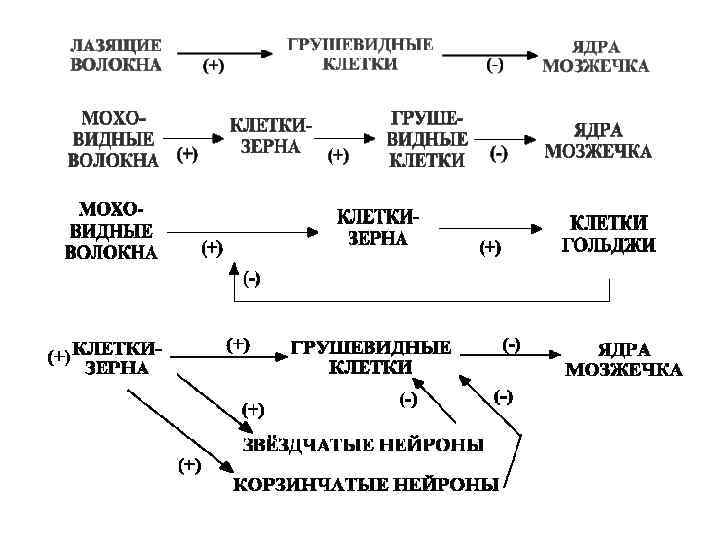
 • Моховидные и лазящие волокна в коре мозжечка образуют возбуждающие синапсы. • Волокна клеток-зёрен заканчиваются возбуждающими синапсами. • Остальные типы связей в коре мозжечка формируют тормозящие синапсы.
• Моховидные и лазящие волокна в коре мозжечка образуют возбуждающие синапсы. • Волокна клеток-зёрен заканчиваются возбуждающими синапсами. • Остальные типы связей в коре мозжечка формируют тормозящие синапсы.
 Схема строения коры мозжечка
Схема строения коры мозжечка
 Основные медиаторы • Возбуждающие – глутамат, аспартат • Тормозные – γ-аминомаслянная кислота • нейропептиды
Основные медиаторы • Возбуждающие – глутамат, аспартат • Тормозные – γ-аминомаслянная кислота • нейропептиды
, оболочки") Глиальные элементы коры мозжечка • Астроциты – формируют периваскулярные пограничные мембраны (компоненты ГЭБ), оболочки вокруг клубочков мозжечка • Клетки (волокна) Бергмана – их отростки охватывают тело клеток Пуркинье, окружают и поддерживают их дендриты • Олигодендроглия – миелинизация волокон • Микроглия - фагоцитирующие клетки, относящиеся к системе мононуклеарных макрофагов
Глиальные элементы коры мозжечка • Астроциты – формируют периваскулярные пограничные мембраны (компоненты ГЭБ), оболочки вокруг клубочков мозжечка • Клетки (волокна) Бергмана – их отростки охватывают тело клеток Пуркинье, окружают и поддерживают их дендриты • Олигодендроглия – миелинизация волокон • Микроглия - фагоцитирующие клетки, относящиеся к системе мононуклеарных макрофагов
 • Специальные нейропсихологические исследования у больных с поражениями мозжечка различного генеза выявили когнитивные нарушения — снижение памяти, интеллекта, нарушение вербального и невербального мышления, оптико-пространственного восприятия, а также эмоционально-личностные изменения
• Специальные нейропсихологические исследования у больных с поражениями мозжечка различного генеза выявили когнитивные нарушения — снижение памяти, интеллекта, нарушение вербального и невербального мышления, оптико-пространственного восприятия, а также эмоционально-личностные изменения
 Функциональная морфология коры головного мозга
Функциональная морфология коры головного мозга
 • Кора полушарий головного мозга – высший нервный центр экранного типа, обеспечивающий регуляцию разнообразных функций организма и сложные формы поведения
• Кора полушарий головного мозга – высший нервный центр экранного типа, обеспечивающий регуляцию разнообразных функций организма и сложные формы поведения
 Ранние стадии развития нервной системы Двухслойный зародыш на подстадии 6 а (досомитный период, 12— 13 дней после оплодотворения, 0, 2 мм).
Ранние стадии развития нервной системы Двухслойный зародыш на подстадии 6 а (досомитный период, 12— 13 дней после оплодотворения, 0, 2 мм).
 Ранние стадии развития нервной системы а — графическая реконструкция первичной полоски с каудальной стороны (от прикрепляющего стебелька); б— схема сагиттального сечения зародыша при формировании нотохорда; в— гистологический срез через первичный бугорок, амнион и желточный мешок; г — формирование изгибов эпибласта и гипобласта на протяжении стадии первичной полоски.
Ранние стадии развития нервной системы а — графическая реконструкция первичной полоски с каудальной стороны (от прикрепляющего стебелька); б— схема сагиттального сечения зародыша при формировании нотохорда; в— гистологический срез через первичный бугорок, амнион и желточный мешок; г — формирование изгибов эпибласта и гипобласта на протяжении стадии первичной полоски.
 Нейруляционный период Нейруляция на 8 -й и 9 -й стадиях (начало сомитного периода развития, 18— 21 -й день после оплодотворения, 1, 5— 2 мм). а— поверхность нервной пластинки в период начала нейруляции и формирования 1 -го сомита; б — нервная пластинка на стадии 3 сомитов (подстадия 9 с) с временными медиальными и пресомитными валиками; в— срез через нервную пластинку до начала подъёма нервных валиков (ростральнее 1 -го сомита); г — срез через нервный желобок позади 1 -го сомита
Нейруляционный период Нейруляция на 8 -й и 9 -й стадиях (начало сомитного периода развития, 18— 21 -й день после оплодотворения, 1, 5— 2 мм). а— поверхность нервной пластинки в период начала нейруляции и формирования 1 -го сомита; б — нервная пластинка на стадии 3 сомитов (подстадия 9 с) с временными медиальными и пресомитными валиками; в— срез через нервную пластинку до начала подъёма нервных валиков (ростральнее 1 -го сомита); г — срез через нервный желобок позади 1 -го сомита
 Нейруляционный период Нейруляция на 9 -й стадии (сомитный период, 19— 21 -й день после оплодотворения, 1, 5— 2 мм). а — срез через 2 -й сомит на трёхсомитной стадии (подстадия 9 с); б — каудальный участок зародыша на уровне формирования кишечномозговой пластинки; в— нервная пластинка в головном регионе зародыша.
Нейруляционный период Нейруляция на 9 -й стадии (сомитный период, 19— 21 -й день после оплодотворения, 1, 5— 2 мм). а — срез через 2 -й сомит на трёхсомитной стадии (подстадия 9 с); б — каудальный участок зародыша на уровне формирования кишечномозговой пластинки; в— нервная пластинка в головном регионе зародыша.
 Схемы поперечных сечений зародыша на стадиях нейруляции а — досомитный этап нейруляции; б — начало сомитогенеза, появление нервного желобка и сомитов; в — замыкание нервной трубки, начало миграции клеток нервного гребня и исчезновение сомитов.
Схемы поперечных сечений зародыша на стадиях нейруляции а — досомитный этап нейруляции; б — начало сомитогенеза, появление нервного желобка и сомитов; в — замыкание нервной трубки, начало миграции клеток нервного гребня и исчезновение сомитов.
 и глиобласт в промежуточном слое коры мозга. Плод") Развитие коры головного мозга Нейробласт (вверху) и глиобласт в промежуточном слое коры мозга. Плод 9 недель развития, контрольная группа. Ув. 24 000.
Развитие коры головного мозга Нейробласт (вверху) и глиобласт в промежуточном слое коры мозга. Плод 9 недель развития, контрольная группа. Ув. 24 000.
 Нейроны коры Пирамидные: Непирамидные: 50 -90% Гигантские Шипиковые звездчатые Крупные Аксо-аксонные Клетки - «канделябры» Клетки с двойным букетом дендритов Горизонтальные клетки Кахаля Клетки Мартинотти и др. Средние Малые Корзинчатые
Нейроны коры Пирамидные: Непирамидные: 50 -90% Гигантские Шипиковые звездчатые Крупные Аксо-аксонные Клетки - «канделябры» Клетки с двойным букетом дендритов Горизонтальные клетки Кахаля Клетки Мартинотти и др. Средние Малые Корзинчатые
 Большой пирамидный нейрон
Большой пирамидный нейрон
 Образование эфферентных путей") Функции клеток Пирамидные: Непирамидные: Интеграция внутри коры (средние и малые клетки) Образование эфферентных путей (гигантские и крупные клетки) Восприятие поступающих афферентных сигналов Распространение импульсов в пределах коры на пирамидные клетки
Функции клеток Пирамидные: Непирамидные: Интеграция внутри коры (средние и малые клетки) Образование эфферентных путей (гигантские и крупные клетки) Восприятие поступающих афферентных сигналов Распространение импульсов в пределах коры на пирамидные клетки
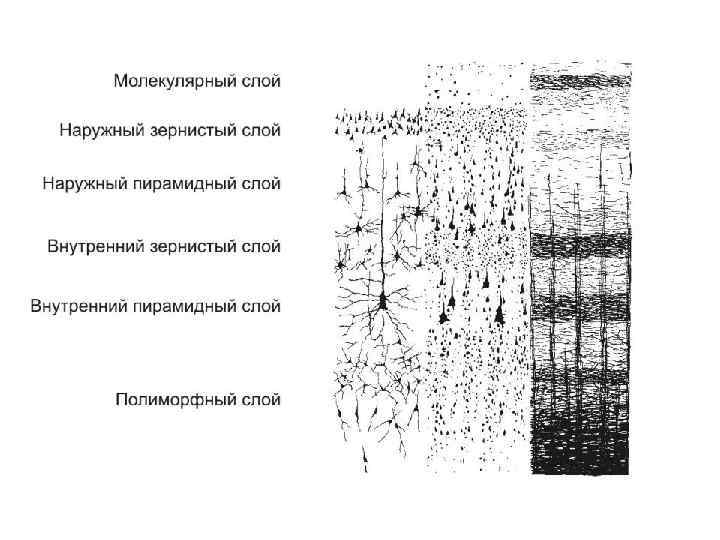
 Цитоархитектоника коры головного мозга
Цитоархитектоника коры головного мозга
 Цитоархитектоника коры головного мозга
Цитоархитектоника коры головного мозга
 Миелоархитектоника Нервные волокна коры Афферентные: Формируют радиальные лучи Оканчиваются на клетках 4 слоя коры Ассоциативные и комиссуральные: Соединяют разные участки коры Формируют полоски Бехтерева, Байярже Эфферентные: Соединяют кору с подкорковыми структурами
Миелоархитектоника Нервные волокна коры Афферентные: Формируют радиальные лучи Оканчиваются на клетках 4 слоя коры Ассоциативные и комиссуральные: Соединяют разные участки коры Формируют полоски Бехтерева, Байярже Эфферентные: Соединяют кору с подкорковыми структурами
 Миелоархитектоника 1. 2. 3. 4. 5. Тангенциальные волокна Полоска Бехтерева Радиальные лучи Наружная полоска Байярже Внутренняя полоска Байярже
Миелоархитектоника 1. 2. 3. 4. 5. Тангенциальные волокна Полоска Бехтерева Радиальные лучи Наружная полоска Байярже Внутренняя полоска Байярже
 5 3 1 6 4 3 1 1 – пирамидные нейроны 2 – шипиковые звездчатые нейроны 3 – корзинчатые нейроны 4 – аксональные нейроны 5 – нейроны с аксональной кисточкой 6 – нейроны с двойным букетом дендритов 7 – афферентные волокна 8 – эфферентные волокна 2 1 7 8 Нейроны: пирамидные синие, звёздчатые (возбуждающие) - жёлтые, тормозные - чёрные. Волокна: афферентные - красные, эфферентные - синие.
5 3 1 6 4 3 1 1 – пирамидные нейроны 2 – шипиковые звездчатые нейроны 3 – корзинчатые нейроны 4 – аксональные нейроны 5 – нейроны с аксональной кисточкой 6 – нейроны с двойным букетом дендритов 7 – афферентные волокна 8 – эфферентные волокна 2 1 7 8 Нейроны: пирамидные синие, звёздчатые (возбуждающие) - жёлтые, тормозные - чёрные. Волокна: афферентные - красные, эфферентные - синие.
 Типы строения коры Агранулярный: моторные центры Развиты III, IV, VI слои коры Слабо развиты II и IV слои Гранулярный: сенсорные центры коры Слабо развиты пирамидные слои Выражены зернистые слои II и IV
Типы строения коры Агранулярный: моторные центры Развиты III, IV, VI слои коры Слабо развиты II и IV слои Гранулярный: сенсорные центры коры Слабо развиты пирамидные слои Выражены зернистые слои II и IV
 Нейромедиаторные системы коры • В коре выделено более десятка различных нейромедиаторов и рецепторов к ним • Норадреналин • Серотонин • Ацетилхолин • Глутамат • Холецистокинин • Гамма-аминомаслянная кислота и др.
Нейромедиаторные системы коры • В коре выделено более десятка различных нейромедиаторов и рецепторов к ним • Норадреналин • Серотонин • Ацетилхолин • Глутамат • Холецистокинин • Гамма-аминомаслянная кислота и др.
") Система вторичных мессенджеров (ц. АМФ)
Система вторичных мессенджеров (ц. АМФ)
 Синтез дофамина, норадреналина и адреналина
Синтез дофамина, норадреналина и адреналина
 Локализация норадреналина в мозге
Локализация норадреналина в мозге
 • • • locus coeruleus ствол мозга мозжечок hypothalamus вся кора головного мозга • предложена «норадреналиновая гипотеза» для описания расстройств настроения.
• • • locus coeruleus ствол мозга мозжечок hypothalamus вся кора головного мозга • предложена «норадреналиновая гипотеза» для описания расстройств настроения.
 Локализация дофамина в мозге
Локализация дофамина в мозге
 • Есть три подсистемы мозга, где дофамин используется как первичный медиатор. Они все локализуются в вентральной тегментальной области. • Одна группа, вокруг substantia nigra, проецируется на хвостатое ядро и putamen и называется нигростриатум. • Второй главный трактат, названный mesocortical или mesolimbic (или mesocorticolimbic), проецируется в префронтальную кору и temporolimbic областям типа миндалины и hippocampus. • Третий компонент системы дофамина находится в hypothalamus и проецируется в гипофиз. • познание и эмоции • шизофрения объяснялась в соответствии с «дофаминовой гипотезой» . Нарушения в мышлении и эмоциях возникают из-за сверхактивности в системе дофамина.
• Есть три подсистемы мозга, где дофамин используется как первичный медиатор. Они все локализуются в вентральной тегментальной области. • Одна группа, вокруг substantia nigra, проецируется на хвостатое ядро и putamen и называется нигростриатум. • Второй главный трактат, названный mesocortical или mesolimbic (или mesocorticolimbic), проецируется в префронтальную кору и temporolimbic областям типа миндалины и hippocampus. • Третий компонент системы дофамина находится в hypothalamus и проецируется в гипофиз. • познание и эмоции • шизофрения объяснялась в соответствии с «дофаминовой гипотезой» . Нарушения в мышлении и эмоциях возникают из-за сверхактивности в системе дофамина.
 Локализация серотонина в мозге
Локализация серотонина в мозге
 • Серотонергические нейроны имеют распределение, поразительно подобное нейронам норадреналина. Места локализации: • вся кора головного мозга, • базальные ганглии, • темпоролимбические области, • hypothalamus, • мозжечок, • ствол мозга. • Предложена «гипотеза серотониновой депрессии» в значительной степени потому, что лечение некоторыми антидепрессантами (типа Prozac) облегчают серотонергическую передачу, блокируя обратный захват. Новые атипичные нейролептики, типа risperidone, также влияют на систему серотонина, что указывает на участие серотонина и в механизмах развития шизофрении.
• Серотонергические нейроны имеют распределение, поразительно подобное нейронам норадреналина. Места локализации: • вся кора головного мозга, • базальные ганглии, • темпоролимбические области, • hypothalamus, • мозжечок, • ствол мозга. • Предложена «гипотеза серотониновой депрессии» в значительной степени потому, что лечение некоторыми антидепрессантами (типа Prozac) облегчают серотонергическую передачу, блокируя обратный захват. Новые атипичные нейролептики, типа risperidone, также влияют на систему серотонина, что указывает на участие серотонина и в механизмах развития шизофрении.
 Локализация ацетилхолина в мозге
Локализация ацетилхолина в мозге
 • • • Тела клеток главной группы нейронов ацетилхолина расположены в базальном ядре Meynert, который находится в средней части globus pallidus. Нейроны от базального ядра Meynert проецируются по всей коре. Вторая группа нейронов ацетилхолина находится в диагональной полоске Broca и связана с hippocampus и gyrus cinguli. Третья группа холинергических нейронов - местные нейроны в пределах базальных ганглиев. • Система ацетилхолина играет главную роль в зашифровывании памяти, хотя точные механизмы пока еще не поняты. Пациенты с болезнью Альцгеймера имеют нарушения распределения холинергичных нейронов в коре и hippocampus. • Дофамин и ацетилхолин имеют высокие концентрации в пределах базальных ганглиев. • Предполагают участие холинергических механизмов в модуляции моторной деятельности и, возможно, в возникновении психозов. Холинергические антагонисты могут также вредить познавательным функциям, ухудшать память пациентов, которым они назначены. Многие обычно используемые лекарства, типа некоторых противозастойных средств, имеют антихолинергические эффекты.
• • • Тела клеток главной группы нейронов ацетилхолина расположены в базальном ядре Meynert, который находится в средней части globus pallidus. Нейроны от базального ядра Meynert проецируются по всей коре. Вторая группа нейронов ацетилхолина находится в диагональной полоске Broca и связана с hippocampus и gyrus cinguli. Третья группа холинергических нейронов - местные нейроны в пределах базальных ганглиев. • Система ацетилхолина играет главную роль в зашифровывании памяти, хотя точные механизмы пока еще не поняты. Пациенты с болезнью Альцгеймера имеют нарушения распределения холинергичных нейронов в коре и hippocampus. • Дофамин и ацетилхолин имеют высокие концентрации в пределах базальных ганглиев. • Предполагают участие холинергических механизмов в модуляции моторной деятельности и, возможно, в возникновении психозов. Холинергические антагонисты могут также вредить познавательным функциям, ухудшать память пациентов, которым они назначены. Многие обычно используемые лекарства, типа некоторых противозастойных средств, имеют антихолинергические эффекты.
") ЛОКАЛИЗАЦИЯ ГАММА-АМИНОМАСЛЯНОЙ КИСЛОТЫ (GABA)
ЛОКАЛИЗАЦИЯ ГАММА-АМИНОМАСЛЯНОЙ КИСЛОТЫ (GABA)
 Локализация GABAергических нейронов • • • хвостатое ядро putamen globus pallidus субстанция нигра мозжечок • GABA- система является главной тормозной системой мозга. • Потеря GABA нейронов, соединяющие n. caudatus и globus pallidus высвобождает последнюю структуру из под тормозящего влияния, таким образом разрешая globus pallidus производить хореиформные движения, которые характеризуют болезнь Гентингтона.
Локализация GABAергических нейронов • • • хвостатое ядро putamen globus pallidus субстанция нигра мозжечок • GABA- система является главной тормозной системой мозга. • Потеря GABA нейронов, соединяющие n. caudatus и globus pallidus высвобождает последнюю структуру из под тормозящего влияния, таким образом разрешая globus pallidus производить хореиформные движения, которые характеризуют болезнь Гентингтона.
 Локализация глутамата в мозге
Локализация глутамата в мозге
, возбуждающий медиатор, продуцируется большими пирамидными клетками всюду в") Роль глутамата • Глутамат (глутаминовая кислота), возбуждающий медиатор, продуцируется большими пирамидными клетками всюду в коре головного мозга и клетками hippocampus. • Глутамат, в дополнение к тому, что является медиатором, может также быть нейротоксином, если содержится в количествах, которые вызывают чрезмерное возбуждение нейрона. • Глутамат, возможно, вовлечен в механизм развития психозов и нейродегенеративных болезней типа болезни Гентингтона. • Избыток глутамата приводит к оцепенению, дезорганизации мышления, нарушениям речи и галлюцинациям.
Роль глутамата • Глутамат (глутаминовая кислота), возбуждающий медиатор, продуцируется большими пирамидными клетками всюду в коре головного мозга и клетками hippocampus. • Глутамат, в дополнение к тому, что является медиатором, может также быть нейротоксином, если содержится в количествах, которые вызывают чрезмерное возбуждение нейрона. • Глутамат, возможно, вовлечен в механизм развития психозов и нейродегенеративных болезней типа болезни Гентингтона. • Избыток глутамата приводит к оцепенению, дезорганизации мышления, нарушениям речи и галлюцинациям.
 Взаимозависимость нейротрансмиттеров и связанных с ними состояний
Взаимозависимость нейротрансмиттеров и связанных с ними состояний