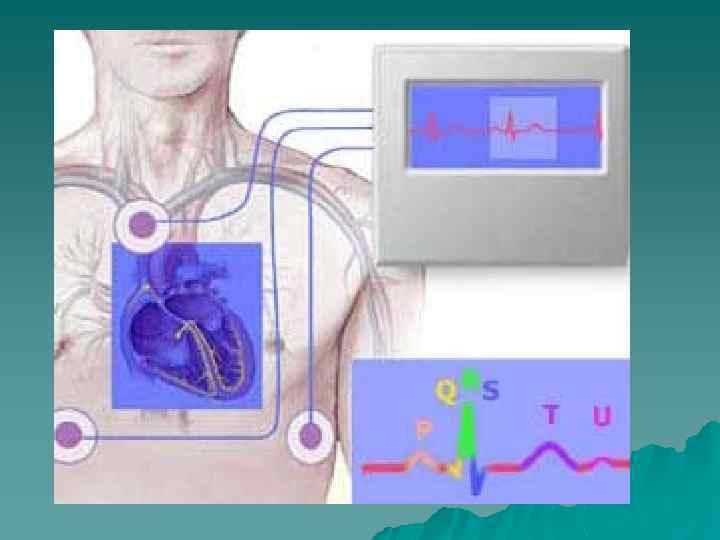
Функции сердца.ppt
- Количество слайдов: 21
 Функции сердца
Функции сердца
 Основные функции сердца u u u Автоматизм(способность генерировать электрический импульс автономно, без постороннего воздействия, и , соответственно, способность сердца ритмично сокращаться в физиологических условиях). Проводимость(способность проводить генерированный импульс по системе проводящих путей, представляющих собой атипичные кардиомиоциты. Проведение импульса происходит путем образования ТМПД в клетках проводящей системы; передача потенциалов происходит за счет межклеточных нексусов). Возбудимость(способность сердца к самовозбуждению, благодаря работе проводящей системы, путем образования трансмембранного потенциала действия. ТМПД, а также в ответ на воздействие химических и физических факторов). Сократимость(способность к выполнению насосной функции сердца), при этом сила сокращения пропорциональна начальной длине мышечного волокна в диастолу. Рефрактерность (состояние временной невозбудимости тканей сердца, в норме обеспечивает востановление их энергетического потенциала).
Основные функции сердца u u u Автоматизм(способность генерировать электрический импульс автономно, без постороннего воздействия, и , соответственно, способность сердца ритмично сокращаться в физиологических условиях). Проводимость(способность проводить генерированный импульс по системе проводящих путей, представляющих собой атипичные кардиомиоциты. Проведение импульса происходит путем образования ТМПД в клетках проводящей системы; передача потенциалов происходит за счет межклеточных нексусов). Возбудимость(способность сердца к самовозбуждению, благодаря работе проводящей системы, путем образования трансмембранного потенциала действия. ТМПД, а также в ответ на воздействие химических и физических факторов). Сократимость(способность к выполнению насосной функции сердца), при этом сила сокращения пропорциональна начальной длине мышечного волокна в диастолу. Рефрактерность (состояние временной невозбудимости тканей сердца, в норме обеспечивает востановление их энергетического потенциала).
 u u u В общем смысле, основной функцией сердца является ритмическое нагнетание крови из вен в артерии, то есть создание градиента давления, вследствие которого происходит её постоянное движение. Это означает, что основной функцией сердца является обеспечение кровообращения сообщением крови кинетической энергии. Сердце поэтому часто ассоциируют с насосом. Его отличают исключительно высокие производительность, скорость и гладкость переходных процессов, запас прочности и постоянное обновление тканей. Для обеспечения нормального существования организма в различных условиях сердце может работать в достаточно широком диапазоне частот, и способность эта определяется вышеупомянутыми свойствами: автоматизм, сократимость и т. д. ). Нагнетание крови обеспечивается посредством попеременного сокращения (систола) и расслабления (диастола) миокарда. Волокна сердечной мышцы сокращаются вследствие электрических импульсов (процессов возбуждения), образующихся в мембране (оболочке) клеток. Эти импульсы появляются ритмически в самом сердце. Свойство сердечной мышцы самостоятельно генерировать периодические импульсы возбуждения называется автоматией. Мышечное сокращение в сердце - хорошо организованный периодический процесс. Функция периодической (хронотропной) организации этого процесса обеспечивается работой проводящей системы.
u u u В общем смысле, основной функцией сердца является ритмическое нагнетание крови из вен в артерии, то есть создание градиента давления, вследствие которого происходит её постоянное движение. Это означает, что основной функцией сердца является обеспечение кровообращения сообщением крови кинетической энергии. Сердце поэтому часто ассоциируют с насосом. Его отличают исключительно высокие производительность, скорость и гладкость переходных процессов, запас прочности и постоянное обновление тканей. Для обеспечения нормального существования организма в различных условиях сердце может работать в достаточно широком диапазоне частот, и способность эта определяется вышеупомянутыми свойствами: автоматизм, сократимость и т. д. ). Нагнетание крови обеспечивается посредством попеременного сокращения (систола) и расслабления (диастола) миокарда. Волокна сердечной мышцы сокращаются вследствие электрических импульсов (процессов возбуждения), образующихся в мембране (оболочке) клеток. Эти импульсы появляются ритмически в самом сердце. Свойство сердечной мышцы самостоятельно генерировать периодические импульсы возбуждения называется автоматией. Мышечное сокращение в сердце - хорошо организованный периодический процесс. Функция периодической (хронотропной) организации этого процесса обеспечивается работой проводящей системы.
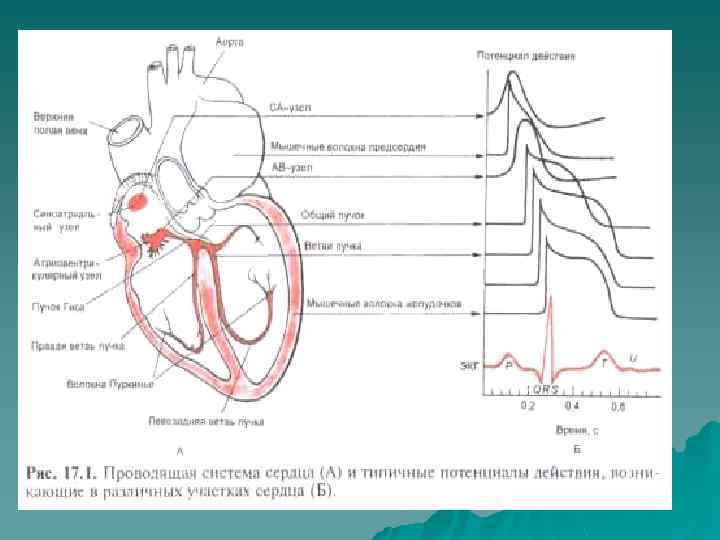
 Трансмембранный потенциал действия u u u ТМПД как проявление функции возбудимости : АРП-абсолютно рефрактерный период( фазы 0, 1, 2 -кардиомиоциты невозбудимы); ОРП-относительно рефрактрный период( фаза 3 -возбудимость частично восстанавливается). Фаза 0 - быстрая реполяризация, длительнось 0. 01 с Фаза 1 -фаза начальной быстрой реполяризации(ТМПД изменяется от +2 до 0 м. В) Фаза 2 -фаза плато, длительность 0. 2 с , ТМПД const. Фаза 3 -фаза конечной быстрой реполяризации, смена заряда мембраны кмц Фаза 4 -фаза диастолы, ТМПД 90 м. В, восстановление концентрации ионов наружной и внутренней поверхности мембраны кмц; кмц в фазе возбуждения.
Трансмембранный потенциал действия u u u ТМПД как проявление функции возбудимости : АРП-абсолютно рефрактерный период( фазы 0, 1, 2 -кардиомиоциты невозбудимы); ОРП-относительно рефрактрный период( фаза 3 -возбудимость частично восстанавливается). Фаза 0 - быстрая реполяризация, длительнось 0. 01 с Фаза 1 -фаза начальной быстрой реполяризации(ТМПД изменяется от +2 до 0 м. В) Фаза 2 -фаза плато, длительность 0. 2 с , ТМПД const. Фаза 3 -фаза конечной быстрой реполяризации, смена заряда мембраны кмц Фаза 4 -фаза диастолы, ТМПД 90 м. В, восстановление концентрации ионов наружной и внутренней поверхности мембраны кмц; кмц в фазе возбуждения.
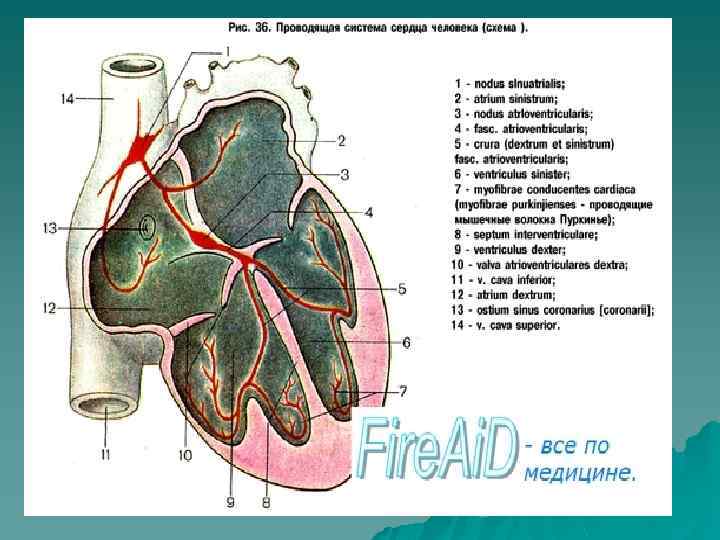
") Распространение возбуждения по миокарду (схема)
Распространение возбуждения по миокарду (схема)
.") Распространение возбуждения по проводящей системе сердца(длительность проведения импульса в норме).
Распространение возбуждения по проводящей системе сердца(длительность проведения импульса в норме).
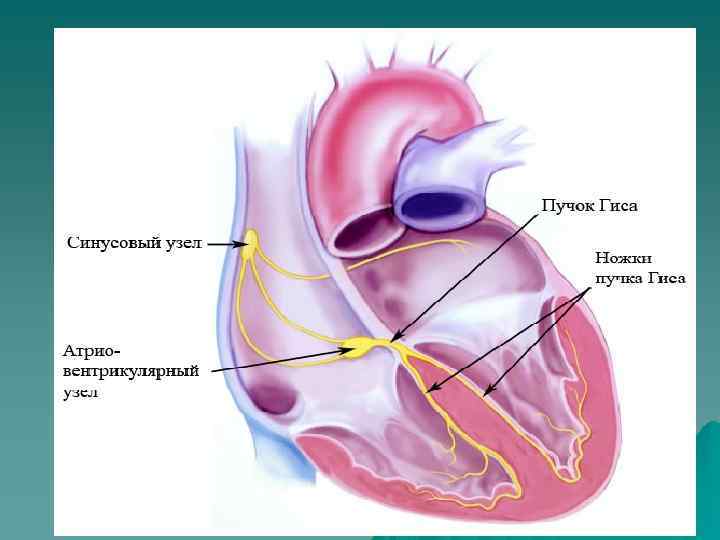
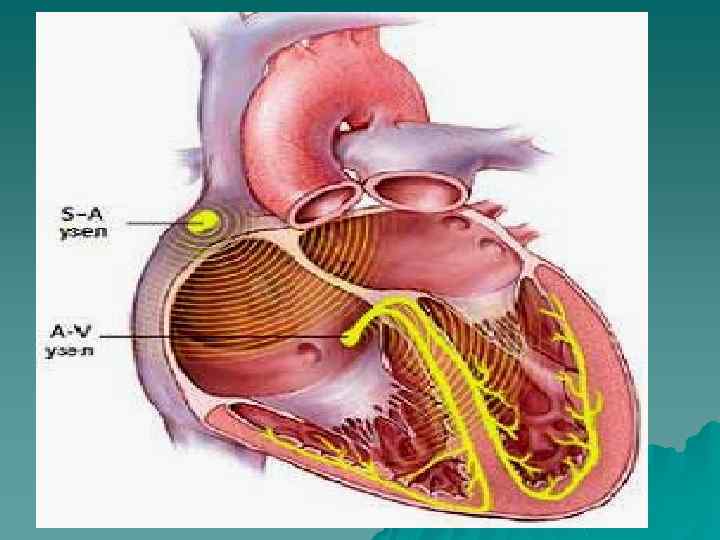
 Иерархия проводящей системы u В проводящей системе существуют центры автоматизма различного порядка: СА-узел генерирует импульсы с ЧСС 60 -90 в минуту; далее в АВ-узле снижается скорость проведения на 3 -5 см/сек, а ЧСС из АВ-узла 40 -50 в минуту; в желудочках по волокнам п. Гиса скорость проведения ещё более снижается, распостранение возбуждения происходит в направлении от субэндокарда к субэпикарду; ЧСС , генерируемое волокнами Пуркинье в условиях блокады-16 -20 в минуту.
Иерархия проводящей системы u В проводящей системе существуют центры автоматизма различного порядка: СА-узел генерирует импульсы с ЧСС 60 -90 в минуту; далее в АВ-узле снижается скорость проведения на 3 -5 см/сек, а ЧСС из АВ-узла 40 -50 в минуту; в желудочках по волокнам п. Гиса скорость проведения ещё более снижается, распостранение возбуждения происходит в направлении от субэндокарда к субэпикарду; ЧСС , генерируемое волокнами Пуркинье в условиях блокады-16 -20 в минуту.
 Сократимость миокарда u В целом, сократительная способность миокарда подчиняется закону СТАРЛИНГА: чем больше растяжение желудочка и кровенаполнение во время диастолы(ударный объем), тем больше сила сокращения во время систолы(чем больше КДО и длина кардиомиоцитов, тем выше инотропная функция). Однако, данный закон применим лишь для здорового сердца.
Сократимость миокарда u В целом, сократительная способность миокарда подчиняется закону СТАРЛИНГА: чем больше растяжение желудочка и кровенаполнение во время диастолы(ударный объем), тем больше сила сокращения во время систолы(чем больше КДО и длина кардиомиоцитов, тем выше инотропная функция). Однако, данный закон применим лишь для здорового сердца.
 Клеточные механизмы сократимости миокарда
Клеточные механизмы сократимости миокарда
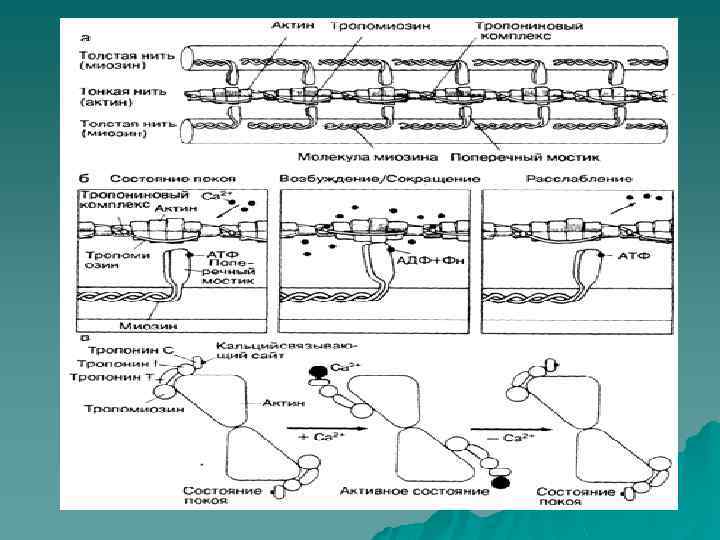
, диаметр которых в норме составлят") u Миокард состоит из отдельных поперечнополосатых мышечных клеток (волокон), диаметр которых в норме составлят 10— 15 мкм, а длина — 30— 60 мкм (рис. 181 -1, а). Каждое волокно включает в себя множество пересекающихся и соединенных между собой нитей (миофибрилл), которые идут на всем протя жении волокна и в свою очередь состоят из периодически повторяющихся струк тур —саркомеров. В цитоплазме между миофибриллами располагаются одно центрально расположенное ядро, многочисленные митохондрии и внутриклеточ ные системы мембран. u Каждый саркомер, являющийся структурной и функциональной едини цей сокращений, ограничен с двух сторон темными линиями, так называемыми Zлиниями (см. рис. 181 -1). Расстояние между Z-линиями зависит от степени сокращения или растяжения мышц и варьирует от 1, 6 до 2, 2 мкм. Внутри саркомера можно видеть чередующиеся светлые и темные полосы, придающие волок нам миокарда характерный исчерченный вид. В центресаркомера расположена широкая темная полоса постоянной ширины (1, 5 мкм) — А-полоса, с двух сторон ее окружают две более светлые I-полосы, ширина которых может меняться. Саркомер сердечной мышцы, так же как и скелетной, построен из миофиламентов двух типов. Более толстые филаменты, состоящие главным образом из белка миозина, идут в продольном направлении и ограничиваются А -полосами. Их диаметр около 100 А, длина— 1, 5— 1, 6 мкм; к периферии они сужаются. Тонкие филаменты состоят прежде всего из актина. Они протянуты между Z-линиями и проходят через I-и А-полосы Диаметр их составляет приблизительно 50 А, длина 1, 0 мкм. Таким образом, толстые и тонкие филаменты накладываются друг на друга только в пределах полос А, полосы I содержат исключительно тонкие фи ламенты(см. рис. 181 -1). При электронной микроскопии можно обнаружить, что между толстыми и тонкими филаментами, проходящими в полосе А, тянутся поперечные мостики.
u Миокард состоит из отдельных поперечнополосатых мышечных клеток (волокон), диаметр которых в норме составлят 10— 15 мкм, а длина — 30— 60 мкм (рис. 181 -1, а). Каждое волокно включает в себя множество пересекающихся и соединенных между собой нитей (миофибрилл), которые идут на всем протя жении волокна и в свою очередь состоят из периодически повторяющихся струк тур —саркомеров. В цитоплазме между миофибриллами располагаются одно центрально расположенное ядро, многочисленные митохондрии и внутриклеточ ные системы мембран. u Каждый саркомер, являющийся структурной и функциональной едини цей сокращений, ограничен с двух сторон темными линиями, так называемыми Zлиниями (см. рис. 181 -1). Расстояние между Z-линиями зависит от степени сокращения или растяжения мышц и варьирует от 1, 6 до 2, 2 мкм. Внутри саркомера можно видеть чередующиеся светлые и темные полосы, придающие волок нам миокарда характерный исчерченный вид. В центресаркомера расположена широкая темная полоса постоянной ширины (1, 5 мкм) — А-полоса, с двух сторон ее окружают две более светлые I-полосы, ширина которых может меняться. Саркомер сердечной мышцы, так же как и скелетной, построен из миофиламентов двух типов. Более толстые филаменты, состоящие главным образом из белка миозина, идут в продольном направлении и ограничиваются А -полосами. Их диаметр около 100 А, длина— 1, 5— 1, 6 мкм; к периферии они сужаются. Тонкие филаменты состоят прежде всего из актина. Они протянуты между Z-линиями и проходят через I-и А-полосы Диаметр их составляет приблизительно 50 А, длина 1, 0 мкм. Таким образом, толстые и тонкие филаменты накладываются друг на друга только в пределах полос А, полосы I содержат исключительно тонкие фи ламенты(см. рис. 181 -1). При электронной микроскопии можно обнаружить, что между толстыми и тонкими филаментами, проходящими в полосе А, тянутся поперечные мостики.
 u. Процесс сокращения. «Скользящая» модель мышечного сокраще ния основана на фундаментальном положении о том, что длина как толстых, так и тонких филаментов остается постоянной и в покое, и во время сокращения. При активации саркомера нити актина и миозина начинают взаимодействовать между собой на уровне соединяющих мостиков, в результате чего нити актина продвигаются глубже внутрь полосы А. В течение всего сокращения ширина полосы А остается постоянной, в то время как полоса I становится уже, а ли нии Z двигаются по направлению друг к другу в одном направлении. При этом глобу лярная часть молекулы отклоняется в сторону, так что получает возможность взаимодействовать с актином, генерируя энергию и вызывая сокращение. М. масса актина 47000. Тонкие филаменты состоят из двойных спи ралей, формируя две цепи молекул актина, переплетенных между собой и тесно связанных с регуляторными белками — тропомиозином и тропонином (тропонины С, I и Т). В отличие от миозина актин не обладает существенной энзиматической активностью, однако он способен обратимо связываться с мио зином в присутствии АТФ и ионов магния, которые активируют. АТФазу миозина. В расслабленной мышце это взаимодействие подавляется тропомиозином. Во время активации ионы кальция присоединяются к тропонину С, что приводит к конформационным изменениям, в результате которых связывающие мостики актина перемещаются кнаружи и становятся доступными для взаимодействия. Физические изменения в связывающих мостиках вызывают скольжение актина вдоль нитей миозина, что неизбежно приводит к укорочению мышцы или разви тию напряжения. Расщепление АТФ сопровождается диссоциацией соединяю щих мостиков между миозином и актином. Образование и разрыв связей между нитями актина и миозина происходят циклично в соответствии с колебаниями концентрации ионов Са. Связи нарушаются, когда концентрация ионов Са сни жается ниже критического уровня, а комплекс тропонин — тропомиозин предот вращает взаимодействия между соединительными мостиками миозина и нитями актина. Ионизированный кальций служит основным медиатором, регулирующим инотропное состояние сердца. Большинство препаратов, усиливающих инотропную функцию миокарда, включая сердечные гликозиды и катехоламины, опосредуют свое действие через повышение доставки ионов кальция к миофиламентам. u. Саркоплазматическая сеть представляет собой сложную цепь соединенных между собой мембранных внутриклеточных каналов, обволакивающую миофибриллы. Она состоит из множества продольно расположенных переплетенных между собой мембранных канальцев, тесно прилегающих к поверхности каждого саркомера. Саркоплазматическая сеть не имеет непосредственного продолжения за грани цами клетки. С саркоплазматической сетью тесно связана система поперечных канальцев, или Т-система, образо ваннаяканалоподобными выпячиваниями сарколеммы, которые проникают внутрь миокардиального волокна вдоль Z-линий, т. е. концевых частей саркомеров.
u. Процесс сокращения. «Скользящая» модель мышечного сокраще ния основана на фундаментальном положении о том, что длина как толстых, так и тонких филаментов остается постоянной и в покое, и во время сокращения. При активации саркомера нити актина и миозина начинают взаимодействовать между собой на уровне соединяющих мостиков, в результате чего нити актина продвигаются глубже внутрь полосы А. В течение всего сокращения ширина полосы А остается постоянной, в то время как полоса I становится уже, а ли нии Z двигаются по направлению друг к другу в одном направлении. При этом глобу лярная часть молекулы отклоняется в сторону, так что получает возможность взаимодействовать с актином, генерируя энергию и вызывая сокращение. М. масса актина 47000. Тонкие филаменты состоят из двойных спи ралей, формируя две цепи молекул актина, переплетенных между собой и тесно связанных с регуляторными белками — тропомиозином и тропонином (тропонины С, I и Т). В отличие от миозина актин не обладает существенной энзиматической активностью, однако он способен обратимо связываться с мио зином в присутствии АТФ и ионов магния, которые активируют. АТФазу миозина. В расслабленной мышце это взаимодействие подавляется тропомиозином. Во время активации ионы кальция присоединяются к тропонину С, что приводит к конформационным изменениям, в результате которых связывающие мостики актина перемещаются кнаружи и становятся доступными для взаимодействия. Физические изменения в связывающих мостиках вызывают скольжение актина вдоль нитей миозина, что неизбежно приводит к укорочению мышцы или разви тию напряжения. Расщепление АТФ сопровождается диссоциацией соединяю щих мостиков между миозином и актином. Образование и разрыв связей между нитями актина и миозина происходят циклично в соответствии с колебаниями концентрации ионов Са. Связи нарушаются, когда концентрация ионов Са сни жается ниже критического уровня, а комплекс тропонин — тропомиозин предот вращает взаимодействия между соединительными мостиками миозина и нитями актина. Ионизированный кальций служит основным медиатором, регулирующим инотропное состояние сердца. Большинство препаратов, усиливающих инотропную функцию миокарда, включая сердечные гликозиды и катехоламины, опосредуют свое действие через повышение доставки ионов кальция к миофиламентам. u. Саркоплазматическая сеть представляет собой сложную цепь соединенных между собой мембранных внутриклеточных каналов, обволакивающую миофибриллы. Она состоит из множества продольно расположенных переплетенных между собой мембранных канальцев, тесно прилегающих к поверхности каждого саркомера. Саркоплазматическая сеть не имеет непосредственного продолжения за грани цами клетки. С саркоплазматической сетью тесно связана система поперечных канальцев, или Т-система, образо ваннаяканалоподобными выпячиваниями сарколеммы, которые проникают внутрь миокардиального волокна вдоль Z-линий, т. е. концевых частей саркомеров.
 u u u Активация клеток миокарда. В покое клетка миокарда поляри зована. т. е. внутренняя поверхность мембраны имеет отрицательный заряд по отношению к наружной поверхности. При этом трансмембранный потенциал со ставляет от — 80 до -100 м. В Главную роль в создании этого потен циала покоя играет сарколемма, которая в состоянии покоя практически непро ницаема для ионов. Na и имеет натрий-калийзависимый насос, изгоняющий ионы Na из клетки. Для работы этого насоса требуется АТФ. Таким образом, внутри клетки накапливается относительно большое количество ионов К и значительно меньше ионов Na, в то время как внеклеточная среда богата ионами Nа и бедна ионами К. В свою очередь в состоянии покоя количе ство ионов. Са вне клетки значительно превышает содержание свободных ионов Са внутри нее. Схематически показано взаимодействие сократительных белков, а также роль кальция как активирующего посредника. (СЛАЙД 14): а — показано относительное расположение сократительных (миозина и актина) и регуляторных белков (тропонинового комплекса и тропомиозина) в миофиламенте; б — сокращение происходит, когда головки молекул миозина, образующие поперечные мостики толстых нитей, связываются с актином. Затем меняется ориентация поперечных мостиков, что приводит к смещению тонких нитей но направлению к центру саркомера. Для активации необходимо связывание ионов Са с тропониновым комплексом, в результате чего снимается тормозящее влияние связи миозина и актина. Одной из серий химических реакций, лежащих в основе мышечного сокращения, является гидролиз АТФ, вызывающий смещение поперечных мостиков. Релаксация наступает после того, как ионы Са 2+ отщепляются от тропонина; в — молекулярная перестройка на уровне тонких нитей затрагивает регуляторные белки ( тропомиозин и тропонины С, I и Т) и заключается в их аллостерических изменениях. Связываясь с тропонином С, кальций ослабляет связь между тропонином I и актином. Развивающаяся диссоциация тропо нина. Т и актиновоп основы тонких нитей приводит к смещению тропомиозина таким образом, что его активные участки становятся доступными для взаимодействия с миозином. По данным: А. М. Katz, V. Е. Smith. Hosp. Proc. , 1984, 19 (1), 69. |Во время плато потенциала действия (фаза 2) отмечается медленный ток ионов внутрь клетки. Он обусловлен движением ионов Са, хотя абсолютные количества. этого иона, относительно малы и сами по себе не могут вызвать полноценную активацию контрактильного аппарата. Деполяризующий ток заря дов распространяется не только по поверхности клетки, но и проникает глубоко в нее, что обеспечивается разветвленной Т-системой. Следствием транссарколеммального движения ионов Са является высвобождение значительно больших его количеств из саркоплазматической сети. Этот процесс получил название «реге неративного высвобождения» ионов. Са. Ионы Са диффундируют по направлению к саркомеру и, как было описано выше, связываются с тропонином, блокируя этот ингибитор сокращения, и акти вируютмиофиламенты, вызывая сокращение. Затем ионы Са вновь накаплива ются в саркоплазматической сети, что, естественно, влечет снижение концентра ции этого иона в миофибриллах до уровня, при котором взаимодействие актина и миозина между собой, лежащее в основе сокращения, становится невозмож ным. Таким образом происходит расслабление мышцы. Очевидно, что в основе периодически сменяющихся сокращений и расслаблении сердечной мышцы лежит способность клеточной мембраны, поперечных канальцев и саркоплазматической сети распространять в пространстве потенциал действия, высвобождать и вновь накапливать ионы Са.
u u u Активация клеток миокарда. В покое клетка миокарда поляри зована. т. е. внутренняя поверхность мембраны имеет отрицательный заряд по отношению к наружной поверхности. При этом трансмембранный потенциал со ставляет от — 80 до -100 м. В Главную роль в создании этого потен циала покоя играет сарколемма, которая в состоянии покоя практически непро ницаема для ионов. Na и имеет натрий-калийзависимый насос, изгоняющий ионы Na из клетки. Для работы этого насоса требуется АТФ. Таким образом, внутри клетки накапливается относительно большое количество ионов К и значительно меньше ионов Na, в то время как внеклеточная среда богата ионами Nа и бедна ионами К. В свою очередь в состоянии покоя количе ство ионов. Са вне клетки значительно превышает содержание свободных ионов Са внутри нее. Схематически показано взаимодействие сократительных белков, а также роль кальция как активирующего посредника. (СЛАЙД 14): а — показано относительное расположение сократительных (миозина и актина) и регуляторных белков (тропонинового комплекса и тропомиозина) в миофиламенте; б — сокращение происходит, когда головки молекул миозина, образующие поперечные мостики толстых нитей, связываются с актином. Затем меняется ориентация поперечных мостиков, что приводит к смещению тонких нитей но направлению к центру саркомера. Для активации необходимо связывание ионов Са с тропониновым комплексом, в результате чего снимается тормозящее влияние связи миозина и актина. Одной из серий химических реакций, лежащих в основе мышечного сокращения, является гидролиз АТФ, вызывающий смещение поперечных мостиков. Релаксация наступает после того, как ионы Са 2+ отщепляются от тропонина; в — молекулярная перестройка на уровне тонких нитей затрагивает регуляторные белки ( тропомиозин и тропонины С, I и Т) и заключается в их аллостерических изменениях. Связываясь с тропонином С, кальций ослабляет связь между тропонином I и актином. Развивающаяся диссоциация тропо нина. Т и актиновоп основы тонких нитей приводит к смещению тропомиозина таким образом, что его активные участки становятся доступными для взаимодействия с миозином. По данным: А. М. Katz, V. Е. Smith. Hosp. Proc. , 1984, 19 (1), 69. |Во время плато потенциала действия (фаза 2) отмечается медленный ток ионов внутрь клетки. Он обусловлен движением ионов Са, хотя абсолютные количества. этого иона, относительно малы и сами по себе не могут вызвать полноценную активацию контрактильного аппарата. Деполяризующий ток заря дов распространяется не только по поверхности клетки, но и проникает глубоко в нее, что обеспечивается разветвленной Т-системой. Следствием транссарколеммального движения ионов Са является высвобождение значительно больших его количеств из саркоплазматической сети. Этот процесс получил название «реге неративного высвобождения» ионов. Са. Ионы Са диффундируют по направлению к саркомеру и, как было описано выше, связываются с тропонином, блокируя этот ингибитор сокращения, и акти вируютмиофиламенты, вызывая сокращение. Затем ионы Са вновь накаплива ются в саркоплазматической сети, что, естественно, влечет снижение концентра ции этого иона в миофибриллах до уровня, при котором взаимодействие актина и миозина между собой, лежащее в основе сокращения, становится невозмож ным. Таким образом происходит расслабление мышцы. Очевидно, что в основе периодически сменяющихся сокращений и расслаблении сердечной мышцы лежит способность клеточной мембраны, поперечных канальцев и саркоплазматической сети распространять в пространстве потенциал действия, высвобождать и вновь накапливать ионы Са.
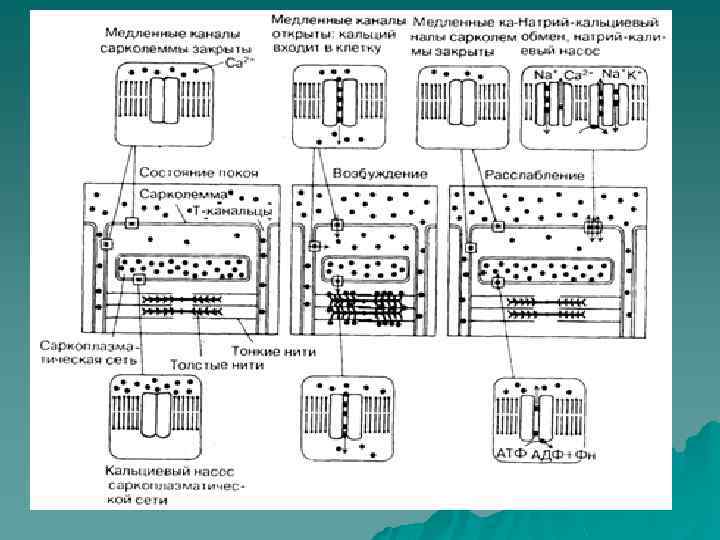
 u u Наиболее мощное сокращение саркомера наблюдают при длине 2, 2 мкм. Именно при такой длине саркомера расположение обоих видов миофиламентов по отно шению друг к другу наиболее благоприятно для их взаимодействия. Имеются данные о том, что длина саркомера определяет также степень актив ности контрактильной системы, т. е. степень ее чувствительности к ионам Са. Максимальная активность установлена при длине саркомера 2, 2 мкм. Если длина саркомера увеличивается до 3, 65 мкм, то создаваемое напряжение падает до нуля, а тонкие нити полностью выходят за пределы А-полосы. С другой стороны, если длина саркомера менее 2, 0 мкм, то происходит скручивание тонких нитей и их двойной перегиб. Одновременно снижается чувствительность контрактильных локусов к ионам Са, а следовательно, и сила сокращения. Зависимость развиваемой силы сокращения от исходной длины мышечных волокон является решающим фактором, определяющим функцию сердечной мышцы. Она лежит в основе правила Франка — Старлинга (закона сердца Старлинга), которое утверждает, что в определенных границах увеличение ис ходного объема желудочка, являющегося производным от длины мышцы, при водит к усилению сокращения желудочка. Было установлено, что в сердечной мышце длина саркомера прямо пропорциональна длине мышцы. Эта зависимость соответствует восходящему колену кривой «длина — активное напряжение мыш цы» . По мере уменьшения длины мышцы до того момента, когда создаваемое напряжение приближается к нулю, а длина саркомера — к 1, 5 мкм, I-полосы сначала сужаются, а затем и вовсе исчезают, в то время как ширина А-полос остается постоянной. В этот момент Z-линии упираются в края А-полос. Таким образом, кривая зависимости силы активного напряжения мышцы от длины саркомера отражает ультраструктурный механизм Старлинга для мышцы сердца. Схема движения ионов кальция(СЛАЙД 15). Кальциевые токи, активирующие мышечное сокращение, направлены вниз, вызывающие расслабление мышц — вверх. Как видно, в покое кальциевые каналы мембран сарко леммы клеток сердечной мышцы закрыты, а внутриклеточный кальций находится в саркоплазматической сети. При возбуждении и деполяризации мембраны натриевые каналы , чувствительные к изменению электрического напряжения, и кальциевые каналы сарколеммы открываются, обусловливая быстрое поступление в клетку внекле точного натрия и кальция. В настоящее время считается, что вхождение ионов Са в клетку извне вызывает его высвобождение из саркоплазматической сети, что и индуцирует сокращение. Условием расслабления сердечной мышцы явля ется повторный захват кальция. АТФ-зависимым кальциевым насосом, расположенным в саркоплазматической сети. Важно то, что сокращение активируется главным образом пассивным током ионов Са из саркоплазматической сети. Напротив, во время диастолы кальций должен активно выкачиваться из цитозоля, обеспечивая возможность релакса ции. Во время диастолы также затрачивается энергия на восстановление градиентов концентраций натрия и кальция по обе стороны сарколеммы, что необходимо для обеспечения деполяризующих ионных токов, участвующих в генерации потенциала действия. Транспорт ионов Na осуществляется сарколеммальным натриевым насосом (натрийкалиевая АТФаза), использующим энергию АТФ для выкачивания натрия из клетки в обмен на калий. Образующийся в результате градиент концентрации натрия и является основным фактором, обеспечивающим активный транспорт ионов Са из клет ки во время релаксации посредством натрий-кальциевого обмена.
u u Наиболее мощное сокращение саркомера наблюдают при длине 2, 2 мкм. Именно при такой длине саркомера расположение обоих видов миофиламентов по отно шению друг к другу наиболее благоприятно для их взаимодействия. Имеются данные о том, что длина саркомера определяет также степень актив ности контрактильной системы, т. е. степень ее чувствительности к ионам Са. Максимальная активность установлена при длине саркомера 2, 2 мкм. Если длина саркомера увеличивается до 3, 65 мкм, то создаваемое напряжение падает до нуля, а тонкие нити полностью выходят за пределы А-полосы. С другой стороны, если длина саркомера менее 2, 0 мкм, то происходит скручивание тонких нитей и их двойной перегиб. Одновременно снижается чувствительность контрактильных локусов к ионам Са, а следовательно, и сила сокращения. Зависимость развиваемой силы сокращения от исходной длины мышечных волокон является решающим фактором, определяющим функцию сердечной мышцы. Она лежит в основе правила Франка — Старлинга (закона сердца Старлинга), которое утверждает, что в определенных границах увеличение ис ходного объема желудочка, являющегося производным от длины мышцы, при водит к усилению сокращения желудочка. Было установлено, что в сердечной мышце длина саркомера прямо пропорциональна длине мышцы. Эта зависимость соответствует восходящему колену кривой «длина — активное напряжение мыш цы» . По мере уменьшения длины мышцы до того момента, когда создаваемое напряжение приближается к нулю, а длина саркомера — к 1, 5 мкм, I-полосы сначала сужаются, а затем и вовсе исчезают, в то время как ширина А-полос остается постоянной. В этот момент Z-линии упираются в края А-полос. Таким образом, кривая зависимости силы активного напряжения мышцы от длины саркомера отражает ультраструктурный механизм Старлинга для мышцы сердца. Схема движения ионов кальция(СЛАЙД 15). Кальциевые токи, активирующие мышечное сокращение, направлены вниз, вызывающие расслабление мышц — вверх. Как видно, в покое кальциевые каналы мембран сарко леммы клеток сердечной мышцы закрыты, а внутриклеточный кальций находится в саркоплазматической сети. При возбуждении и деполяризации мембраны натриевые каналы , чувствительные к изменению электрического напряжения, и кальциевые каналы сарколеммы открываются, обусловливая быстрое поступление в клетку внекле точного натрия и кальция. В настоящее время считается, что вхождение ионов Са в клетку извне вызывает его высвобождение из саркоплазматической сети, что и индуцирует сокращение. Условием расслабления сердечной мышцы явля ется повторный захват кальция. АТФ-зависимым кальциевым насосом, расположенным в саркоплазматической сети. Важно то, что сокращение активируется главным образом пассивным током ионов Са из саркоплазматической сети. Напротив, во время диастолы кальций должен активно выкачиваться из цитозоля, обеспечивая возможность релакса ции. Во время диастолы также затрачивается энергия на восстановление градиентов концентраций натрия и кальция по обе стороны сарколеммы, что необходимо для обеспечения деполяризующих ионных токов, участвующих в генерации потенциала действия. Транспорт ионов Na осуществляется сарколеммальным натриевым насосом (натрийкалиевая АТФаза), использующим энергию АТФ для выкачивания натрия из клетки в обмен на калий. Образующийся в результате градиент концентрации натрия и является основным фактором, обеспечивающим активный транспорт ионов Са из клет ки во время релаксации посредством натрий-кальциевого обмена.