

 Фотосинтез
Фотосинтез
 • Процесс фотосинтеза — это основной путь, по которому практически вся энергия входит в нашу биосферу. • В течение года в результате фотосинтеза на Земле: • усваивается около 200 млрд. тонн углекислого газа, • образуется около 100 млрд. тонн органического вещества и • выделяется около 145 млрд. тонн молекулярного кислорода. • Без притока энергии от Солнца, в основном преобразуемой в хлоропластах эукариотических клеток, жизнь на нашей планете, постепенно подчиняясь неумолимому второму закону термодинамики, вероятно, прекратилась бы навсегда.
• Процесс фотосинтеза — это основной путь, по которому практически вся энергия входит в нашу биосферу. • В течение года в результате фотосинтеза на Земле: • усваивается около 200 млрд. тонн углекислого газа, • образуется около 100 млрд. тонн органического вещества и • выделяется около 145 млрд. тонн молекулярного кислорода. • Без притока энергии от Солнца, в основном преобразуемой в хлоропластах эукариотических клеток, жизнь на нашей планете, постепенно подчиняясь неумолимому второму закону термодинамики, вероятно, прекратилась бы навсегда.
 История изучения фотосинтеза • Немногим более 300 лет назад в одном из первых тщательно продуманных биологических экспериментов голландский врач Ян Баптист ван Гельмонт (1577— 1644) представил первое доказательство того, что не одна почва кормит растение. • Ван Гельмонт выращивал маленькое дерево ивы в глиняном горшке, добавляя в него только воду. Через пять лет масса ивы увеличилась на 74, 4 кг, в то время как масса почвы уменьшилась только на 57 г. • На основании этих данных ван Гельмонт сделал вывод о том, что все вещества растения образуются из воды, а не из почвы и воздуха!
История изучения фотосинтеза • Немногим более 300 лет назад в одном из первых тщательно продуманных биологических экспериментов голландский врач Ян Баптист ван Гельмонт (1577— 1644) представил первое доказательство того, что не одна почва кормит растение. • Ван Гельмонт выращивал маленькое дерево ивы в глиняном горшке, добавляя в него только воду. Через пять лет масса ивы увеличилась на 74, 4 кг, в то время как масса почвы уменьшилась только на 57 г. • На основании этих данных ван Гельмонт сделал вывод о том, что все вещества растения образуются из воды, а не из почвы и воздуха!
 Опыты Ван-Гельмонта ВОДА дождевая Масса почвы – 91 кг Масса ивового побега – 2, 25 кг. Масса почвы – 91 кг 57 г. = 90, 43 кг. Масса ивового побега – 75 кг.
Опыты Ван-Гельмонта ВОДА дождевая Масса почвы – 91 кг Масса ивового побега – 2, 25 кг. Масса почвы – 91 кг 57 г. = 90, 43 кг. Масса ивового побега – 75 кг.
 сообщил,") История изучения фотосинтеза В конце XVIII в. английский ученый Джозеф Пристли (1733— 1804) сообщил, что он «случайно обнаружил метод исправления воздуха, который был испорчен горением свечей» . 17 августа 1771 г. Пристли «поместил живую веточку мяты в закрытый сосуд, в котором сгорела восковая свеча» , а 27 го числа того же месяца обнаружил, что «другая свеча снова могла гореть в этом же сосуде» . «Исправляющим началом, которым для этих целей пользуется природа, — полагал Пристли, — было растение» . Он расширил свои наблюдения и скоро показал, что воздух, «исправляемый» растением, не был «совсем неподходящим для мыши» .
История изучения фотосинтеза В конце XVIII в. английский ученый Джозеф Пристли (1733— 1804) сообщил, что он «случайно обнаружил метод исправления воздуха, который был испорчен горением свечей» . 17 августа 1771 г. Пристли «поместил живую веточку мяты в закрытый сосуд, в котором сгорела восковая свеча» , а 27 го числа того же месяца обнаружил, что «другая свеча снова могла гореть в этом же сосуде» . «Исправляющим началом, которым для этих целей пользуется природа, — полагал Пристли, — было растение» . Он расширил свои наблюдения и скоро показал, что воздух, «исправляемый» растением, не был «совсем неподходящим для мыши» .
 подтвердил работу Пристли и") История изучения фотосинтеза Позднее голландский врач Ян Ингснхауз (1730— 1799) подтвердил работу Пристли и показал, что воздух «исправляется» только на солнечном свету и только зелеными частями растения. В 1796 г. Ингснхауз предположил, что углекислота разлагается при фотосинтезе с образованием углерода и кислорода, а кислород выделяется в виде газа. Впоследствии было обнаружено, что соотношение атомов углерода, водорода и кислорода в сахарах и крахмале таково, что один атом углерода приходится на одну молекулу воды (СН 20), на что и указывает слово «углеводы» . Таким образом, в суммарном уравнении фотосинтеза С 02+Н 20+Энергия света = (СН 20)+02 Считалось общепринятым, что углеводы образуются из углерода и воды, а кислород выделяется из углекислоты. Эта вполне разумная гипотеза была широко признана, но, как позднее выяснилось, она была совершенно неверной.
История изучения фотосинтеза Позднее голландский врач Ян Ингснхауз (1730— 1799) подтвердил работу Пристли и показал, что воздух «исправляется» только на солнечном свету и только зелеными частями растения. В 1796 г. Ингснхауз предположил, что углекислота разлагается при фотосинтезе с образованием углерода и кислорода, а кислород выделяется в виде газа. Впоследствии было обнаружено, что соотношение атомов углерода, водорода и кислорода в сахарах и крахмале таково, что один атом углерода приходится на одну молекулу воды (СН 20), на что и указывает слово «углеводы» . Таким образом, в суммарном уравнении фотосинтеза С 02+Н 20+Энергия света = (СН 20)+02 Считалось общепринятым, что углеводы образуются из углерода и воды, а кислород выделяется из углекислоты. Эта вполне разумная гипотеза была широко признана, но, как позднее выяснилось, она была совершенно неверной.
 Исследователем, который опроверг эту общепринятую теорию, был Корнелис ван Ниль из Станфордского университета, который, будучи еще студентом дипломником, исследовал метаболизм различных фотосинтезирующих бактерий. Одна группа таких бактерий, а именно пурпурные серные бактерии, восстанавливает углерод до углеводов, но не выделяет кислород. Пурпурным серным бактериям для фотосинтеза необходим сероводород. В результате фотосинтеза внутри бактериальных клеток накапливаются частицы серы. Ван Ниль обнаружил, что для этих бактерий уравнение фотосинтеза может быть записано следующим образом: C 02+2 H 2 S → свет (CH 20)+H 20+2 S. Этот факт не привлекал внимания исследователей до тех пор, пока ван Ниль не сделал смелого обобщения и не предложил следующее суммарное уравнение фотосинтеза: С 02+2 Н 2 А→ свет (CH 20)+H 20+2 А. В этом уравнении Н 2 А представляет собой либо воду, либо другое окисляемое вещество, например сероводород или свободный водород. У водорослей и зеленых растений Н 2 А — это вода. Таким образом, ван Ниль предположил, что вода, а не углекислота разлагается при фотосинтезе.
Исследователем, который опроверг эту общепринятую теорию, был Корнелис ван Ниль из Станфордского университета, который, будучи еще студентом дипломником, исследовал метаболизм различных фотосинтезирующих бактерий. Одна группа таких бактерий, а именно пурпурные серные бактерии, восстанавливает углерод до углеводов, но не выделяет кислород. Пурпурным серным бактериям для фотосинтеза необходим сероводород. В результате фотосинтеза внутри бактериальных клеток накапливаются частицы серы. Ван Ниль обнаружил, что для этих бактерий уравнение фотосинтеза может быть записано следующим образом: C 02+2 H 2 S → свет (CH 20)+H 20+2 S. Этот факт не привлекал внимания исследователей до тех пор, пока ван Ниль не сделал смелого обобщения и не предложил следующее суммарное уравнение фотосинтеза: С 02+2 Н 2 А→ свет (CH 20)+H 20+2 А. В этом уравнении Н 2 А представляет собой либо воду, либо другое окисляемое вещество, например сероводород или свободный водород. У водорослей и зеленых растений Н 2 А — это вода. Таким образом, ван Ниль предположил, что вода, а не углекислота разлагается при фотосинтезе.
 История изучения фотосинтеза Около двухсот лет назад было обнаружено, что для процесса, называемого сейчас фотосинтезом, необходим свет. В настоящее время известно, что фотосинтез проходит в две стадии, но только одна из них — на свету. Доказательства двухстадийности процесса впервые были получены в 1905 г. английским физиологом растений Ф. Ф. Блэкменом, который исследовал влияние освещенности и температуры на скорость фотосинтеза. Блэкмсн сделал следующие выводы: Имеется одна группа светозависимых реакций, которые не зависят от температуры. Скорость этих реакций в диапазоне низких освещенностей могла возрастать с увеличением освещенности, но не с увеличением температуры! Имеется вторая группа реакций, зависимых от температуры, а не от света. Оказалось, что обе группы реакций необходимы для осуществления фотосинтеза.
История изучения фотосинтеза Около двухсот лет назад было обнаружено, что для процесса, называемого сейчас фотосинтезом, необходим свет. В настоящее время известно, что фотосинтез проходит в две стадии, но только одна из них — на свету. Доказательства двухстадийности процесса впервые были получены в 1905 г. английским физиологом растений Ф. Ф. Блэкменом, который исследовал влияние освещенности и температуры на скорость фотосинтеза. Блэкмсн сделал следующие выводы: Имеется одна группа светозависимых реакций, которые не зависят от температуры. Скорость этих реакций в диапазоне низких освещенностей могла возрастать с увеличением освещенности, но не с увеличением температуры! Имеется вторая группа реакций, зависимых от температуры, а не от света. Оказалось, что обе группы реакций необходимы для осуществления фотосинтеза.
 Световые и темновые реакции Световые реакции: Темновые реакции: Зависят от света Не зависят от температуры Зависят от температуры Быстрые < 10 ( 5) сек Медленные ~ 10 ( 2) сек Протекают на мембранах Протекают в строме Хл
Световые и темновые реакции Световые реакции: Темновые реакции: Зависят от света Не зависят от температуры Зависят от температуры Быстрые < 10 ( 5) сек Медленные ~ 10 ( 2) сек Протекают на мембранах Протекают в строме Хл
 СВЕТОВЫЕ РЕАКЦИИ Первый этап преобразования световой энергии в химическую — это поглощение света. Пигмент представляет собой соединение, которое поглощает видимый свет. Некоторые пигменты поглощают свет любой длины волны и поэтому кажутся черными. Другие — свет только определенной длины волны, а остальной пропускают или отражают. Хлорофилл — это пигмент, который обусловливает зеленый цвет листьев; он поглощает в основном фиолетовые и синие лучи, а также красный свет, при этом отражая зеленый (по этому он и кажется зеленым). Основные классы фотосинтетических пигментов: хлорофиллы; каротиноиды и фикобилины
СВЕТОВЫЕ РЕАКЦИИ Первый этап преобразования световой энергии в химическую — это поглощение света. Пигмент представляет собой соединение, которое поглощает видимый свет. Некоторые пигменты поглощают свет любой длины волны и поэтому кажутся черными. Другие — свет только определенной длины волны, а остальной пропускают или отражают. Хлорофилл — это пигмент, который обусловливает зеленый цвет листьев; он поглощает в основном фиолетовые и синие лучи, а также красный свет, при этом отражая зеленый (по этому он и кажется зеленым). Основные классы фотосинтетических пигментов: хлорофиллы; каротиноиды и фикобилины
.") Хлорофиллы с Хлорофиллы тетрапирролы, образующие циклическую структуру хлорофилла (магний порфирины).
Хлорофиллы с Хлорофиллы тетрапирролы, образующие циклическую структуру хлорофилла (магний порфирины).
 Конъюгированная система сопряженных связей у всех хлорофиллов представляет собой основную хромофорную группу, ответственную за избирательное поглощение световой энергии. Циклическая система сопряженных двойных связей, образующая делокализованную орбиту, имеет ряд дополнительных сопряжений: винильную группу в I кольце, полуизолированная связь во II кольце, С=О группа в V кольце. В результате возникает сложно разветвленная конъюгированная система, которая обеспечивает высокую поглощающую способность Mg порфиринов.
Конъюгированная система сопряженных связей у всех хлорофиллов представляет собой основную хромофорную группу, ответственную за избирательное поглощение световой энергии. Циклическая система сопряженных двойных связей, образующая делокализованную орбиту, имеет ряд дополнительных сопряжений: винильную группу в I кольце, полуизолированная связь во II кольце, С=О группа в V кольце. В результате возникает сложно разветвленная конъюгированная система, которая обеспечивает высокую поглощающую способность Mg порфиринов.
 Встречаемость хлорофиллов у представителей различных отделов Хлорофилл а характерен для всех фотосинтезирующих эукариот и цианобактерий. Полагают, что это основной пигмент, участвующий в процессах фотосинтеза у данных организмов. Сосудистые растения, мхи, зеленые и эвгленовыс водоросли содержат и хлорофилл Ь. Это вспомогательный пигмент, который расширяет спектр поглощения света в процессе фотосинтеза. Когда молекула хлорофилла b поглощает свет, то возбужденный электрон передает свою энергию хлорофиллу а, который затем в процессе фотосинтеза преобразует ее в энергию химических связей. Поскольку хлорофилл b поглощает свет других длин волн, нежели хлорофилл а, то тем самым расширяется диапазон длин волн, которые могут ис пользоваться для фотосинтеза. Содержание хлорофилла b в листьях зеленых растений составляет в целом 1/4 общего количества хлорофилла. У некоторых групп водорослей, в основном бурых и диатомовых, вместо хлорофилла b функционирует хлорофилл с. Фотосинтсзирующие бактерии (не цианобактерии) не могут использовать электроны воды и поэтому не выделяют кислород. Они содержат либо бактериохлорофилл (пурпурные бактерии), либо хлоробиум хлорофилл (зеленые серные бактерии).
Встречаемость хлорофиллов у представителей различных отделов Хлорофилл а характерен для всех фотосинтезирующих эукариот и цианобактерий. Полагают, что это основной пигмент, участвующий в процессах фотосинтеза у данных организмов. Сосудистые растения, мхи, зеленые и эвгленовыс водоросли содержат и хлорофилл Ь. Это вспомогательный пигмент, который расширяет спектр поглощения света в процессе фотосинтеза. Когда молекула хлорофилла b поглощает свет, то возбужденный электрон передает свою энергию хлорофиллу а, который затем в процессе фотосинтеза преобразует ее в энергию химических связей. Поскольку хлорофилл b поглощает свет других длин волн, нежели хлорофилл а, то тем самым расширяется диапазон длин волн, которые могут ис пользоваться для фотосинтеза. Содержание хлорофилла b в листьях зеленых растений составляет в целом 1/4 общего количества хлорофилла. У некоторых групп водорослей, в основном бурых и диатомовых, вместо хлорофилла b функционирует хлорофилл с. Фотосинтсзирующие бактерии (не цианобактерии) не могут использовать электроны воды и поэтому не выделяют кислород. Они содержат либо бактериохлорофилл (пурпурные бактерии), либо хлоробиум хлорофилл (зеленые серные бактерии).
 Каротиноиды — это красные, оранжевые или желтые жирорастворимые пигменты, обнаруженные в хлоропластах и у цианобактсрий. Подобно хлорофиллам, каротиноиды хлоропластов погружены в тилакоидные мембраны. В хлоропластах обычно присутствуют две группы каротиноидов — каротины и ксантофиллы (последние в отличие от каротинов содержат кислород). Бета каротин, обнаруженный в растениях, служит основным источником витамина А, необхо димого человеку и животным. В зеленых листьях каротиноиды маскируются большим количеством хлорофилла.
Каротиноиды — это красные, оранжевые или желтые жирорастворимые пигменты, обнаруженные в хлоропластах и у цианобактсрий. Подобно хлорофиллам, каротиноиды хлоропластов погружены в тилакоидные мембраны. В хлоропластах обычно присутствуют две группы каротиноидов — каротины и ксантофиллы (последние в отличие от каротинов содержат кислород). Бета каротин, обнаруженный в растениях, служит основным источником витамина А, необхо димого человеку и животным. В зеленых листьях каротиноиды маскируются большим количеством хлорофилла.
 Фикобилины открытые тетрапиррольные структуры Фикобилины характерны для цианобактсрий и хлоропластов красных водорослей. В отличие от каротиноидов фикобилины растворяются в воде.
Фикобилины открытые тетрапиррольные структуры Фикобилины характерны для цианобактсрий и хлоропластов красных водорослей. В отличие от каротиноидов фикобилины растворяются в воде.
 Спектры поглощения Хлорофиллы: в красной области спектра 640 700 нм в синей 400 450 нм Каротиноиды: 400 550 нм главный максимум: 480 нм
Спектры поглощения Хлорофиллы: в красной области спектра 640 700 нм в синей 400 450 нм Каротиноиды: 400 550 нм главный максимум: 480 нм
 Все производные магний порфиринов имеют характерные спектры поглощения. В спектрах хлорофиллов а и в главными являются две интенсивные полосы поглощения – в красной области спектра (640 700 нм), что обусловлено Sо S 1* электронным переходом, и в синей области, так называемая «полоса Соре» (400 450 нм), соответствующая Sо S 2* электронному переходу. У хлорофилла в оба максимума расположены между двумя максимумами поглощения хлорофилла а, что определяет различия в их окраске – сине зеленый оттенок у хлорофилла а и желто зеленый у хлорофилла в. Кроме того, одна и та же химическая структура в живом листе в комплексе с белками и липидами образует целую серию так называемых «нативных» форм. Так, для хлорофилла а известно около 10 таких спектрально различных форм, из них 4 формы считаются универсальными – 661, 670, 678, 683 нм (в эфире). Для каротиноидов известно несколько различных состояний одной и той же структуры, например, максимум поглощения β каротина мономерного – 450 нм, в комплексе с белком – 455 460 нм, кристаллического – 540 нм.
Все производные магний порфиринов имеют характерные спектры поглощения. В спектрах хлорофиллов а и в главными являются две интенсивные полосы поглощения – в красной области спектра (640 700 нм), что обусловлено Sо S 1* электронным переходом, и в синей области, так называемая «полоса Соре» (400 450 нм), соответствующая Sо S 2* электронному переходу. У хлорофилла в оба максимума расположены между двумя максимумами поглощения хлорофилла а, что определяет различия в их окраске – сине зеленый оттенок у хлорофилла а и желто зеленый у хлорофилла в. Кроме того, одна и та же химическая структура в живом листе в комплексе с белками и липидами образует целую серию так называемых «нативных» форм. Так, для хлорофилла а известно около 10 таких спектрально различных форм, из них 4 формы считаются универсальными – 661, 670, 678, 683 нм (в эфире). Для каротиноидов известно несколько различных состояний одной и той же структуры, например, максимум поглощения β каротина мономерного – 450 нм, в комплексе с белком – 455 460 нм, кристаллического – 540 нм.
 Спектры поглощения
Спектры поглощения
 Спектр поглощении хлорофилла характеризует долю энергии поглощенного света в зависимости от длины волны. Доказательство того, что хлорофилл — это основной пигмент, участвующий в фотосинтезе, заключается в сходстве его спектра поглощения со спектром действия фотосинтеза. Спектр действия определяет относительную эффективность различных длин волн света для таких светозависимых процессов, как фотосинтез, цветение, фототропизм (изгибание растения по направлению к свету). Сходство спектра поглощения пигмента и спектра действия процесса считается доказательством того, что именно данный пигмент обеспечивает осуществление этого процесса.
Спектр поглощении хлорофилла характеризует долю энергии поглощенного света в зависимости от длины волны. Доказательство того, что хлорофилл — это основной пигмент, участвующий в фотосинтезе, заключается в сходстве его спектра поглощения со спектром действия фотосинтеза. Спектр действия определяет относительную эффективность различных длин волн света для таких светозависимых процессов, как фотосинтез, цветение, фототропизм (изгибание растения по направлению к свету). Сходство спектра поглощения пигмента и спектра действия процесса считается доказательством того, что именно данный пигмент обеспечивает осуществление этого процесса.
 Когда пигменты поглощают свет и электроны поднимаются на более высокий энергетический уровень, дальнейшие события могут развиваться следующим образом: • энергия электрона рассеивается в виде тепла; • немедленно выделяется в виде световой энергии большей длины волны (это явление называют флуоресценцией, однако когда происходит испускание света с некоторой задержкой, то данное явление называют фосфоресценцией) или • энергия запасается в форме химических связей, что и происходит в процессе фотосинтеза. Если выделить молекулы хлорофилла и осветить их, они флуоресцируют. Иными словами, молекулы пигмента поглощают световую энергию, поэтому электроны немедленно поднимаются на более высокий энергетический уровень, а затем спускаются на более низкий, при этом выделяя большую часть поглощенной энергии в виде света. Свет, поглощенный изолированными молекулами хлорофилла, не может преобразоваться в какую либо другую форму энергии, полезную для живых систем. Хлорофилл способен преобразовать энергию света в химическую только в комплексе с определенными белками, находящимися в тилакоидах.
Когда пигменты поглощают свет и электроны поднимаются на более высокий энергетический уровень, дальнейшие события могут развиваться следующим образом: • энергия электрона рассеивается в виде тепла; • немедленно выделяется в виде световой энергии большей длины волны (это явление называют флуоресценцией, однако когда происходит испускание света с некоторой задержкой, то данное явление называют фосфоресценцией) или • энергия запасается в форме химических связей, что и происходит в процессе фотосинтеза. Если выделить молекулы хлорофилла и осветить их, они флуоресцируют. Иными словами, молекулы пигмента поглощают световую энергию, поэтому электроны немедленно поднимаются на более высокий энергетический уровень, а затем спускаются на более низкий, при этом выделяя большую часть поглощенной энергии в виде света. Свет, поглощенный изолированными молекулами хлорофилла, не может преобразоваться в какую либо другую форму энергии, полезную для живых систем. Хлорофилл способен преобразовать энергию света в химическую только в комплексе с определенными белками, находящимися в тилакоидах.
 Хлорофилл способен к окислительно-восстановительным превращениям «реакция Красновского» h Донор е Акцептор е Хлорофилл 1 – флуоресценция S*2 2 – миграция энергии 1 S*1 2 3 – химическая энергия 3 4 - фосфоресценция T 430 S 0 680 4 3
Хлорофилл способен к окислительно-восстановительным превращениям «реакция Красновского» h Донор е Акцептор е Хлорофилл 1 – флуоресценция S*2 2 – миграция энергии 1 S*1 2 3 – химическая энергия 3 4 - фосфоресценция T 430 S 0 680 4 3
 Известно два основных типа возбужденных состояний – синглетные и триплетные. Они отличаются по энергии и состоянию спина электрона. В синглетном возбужденном состоянии спины электронов на основном и возбужденном уровнях остаются антипараллельными, при переходе в триплетное состояние происходит поворот спина возбужденного электрона с образованием бирадикальной системы. При поглощении фотона молекула хлорофилла переходит из основного (S 0) в одно из возбужденных синглетных состояний – S*1 или S*2, что сопровождается переходом электрона на возбужденный уровень с более высокой энергией. Возбужденное состояние S*2 очень не стабильно, электрон быстро (в течение 10 13 сек) теряет часть энергии в виде тепла и опускается на нижний колебательный уровень S*1, где он может находиться в течение 10 9 сек. В состоянии S*1 возможно обращение спина электрона и переход в триплетное состояние Т 1, энергия которого ниже S*1.
Известно два основных типа возбужденных состояний – синглетные и триплетные. Они отличаются по энергии и состоянию спина электрона. В синглетном возбужденном состоянии спины электронов на основном и возбужденном уровнях остаются антипараллельными, при переходе в триплетное состояние происходит поворот спина возбужденного электрона с образованием бирадикальной системы. При поглощении фотона молекула хлорофилла переходит из основного (S 0) в одно из возбужденных синглетных состояний – S*1 или S*2, что сопровождается переходом электрона на возбужденный уровень с более высокой энергией. Возбужденное состояние S*2 очень не стабильно, электрон быстро (в течение 10 13 сек) теряет часть энергии в виде тепла и опускается на нижний колебательный уровень S*1, где он может находиться в течение 10 9 сек. В состоянии S*1 возможно обращение спина электрона и переход в триплетное состояние Т 1, энергия которого ниже S*1.
 Фотосистемы В хлоропластах хлорофилл и другие пигменты, погруженные в тилакоиды, собраны в функциональные единицы, называемые фотосистемами. Каждая фотосистема содержит около 250— 400 молекул пигментов.
Фотосистемы В хлоропластах хлорофилл и другие пигменты, погруженные в тилакоиды, собраны в функциональные единицы, называемые фотосистемами. Каждая фотосистема содержит около 250— 400 молекул пигментов.
, но только одна молекула хлорофилла") Все пигменты фотосистемы могут поглощать фотоны (частицы световой энергии), но только одна молекула хлорофилла данной фотосистемы может использовать поглощенную энергию в фотохимических реакциях. Эта молекула хлорофилла называется реакционным центром фотосистемы, а другие молекулы пигментов называются антенными, поскольку они, подобно антеннам, собирают свет. Внутренняя поверхность тилакоида. Частицы, погруженные в мембраны представляют собой фотосистемы
Все пигменты фотосистемы могут поглощать фотоны (частицы световой энергии), но только одна молекула хлорофилла данной фотосистемы может использовать поглощенную энергию в фотохимических реакциях. Эта молекула хлорофилла называется реакционным центром фотосистемы, а другие молекулы пигментов называются антенными, поскольку они, подобно антеннам, собирают свет. Внутренняя поверхность тилакоида. Частицы, погруженные в мембраны представляют собой фотосистемы
 Расположение белковых комплексов в мембране тилакоида
Расположение белковых комплексов в мембране тилакоида
 Световая энергия, поглощенная молекулой пигмента, переносится на другую молекулу, пока не достигнет реакционного центра, где локализована специальная молекула хлорофилла а. Когда эта молекула поглощает световую энергию, электроны поднимаются на более высокий энергетический уровень и переносятся на молекулу акцептора, вызывая поток электронов. Таким образом, молекула хлорофилла окисляется и становится положительно заряженной. Фотореакционный центр: 1 – свет; 2 – светособирающие пигменты; 3 – донор электрона; 4 акцептор электрона
Световая энергия, поглощенная молекулой пигмента, переносится на другую молекулу, пока не достигнет реакционного центра, где локализована специальная молекула хлорофилла а. Когда эта молекула поглощает световую энергию, электроны поднимаются на более высокий энергетический уровень и переносятся на молекулу акцептора, вызывая поток электронов. Таким образом, молекула хлорофилла окисляется и становится положительно заряженной. Фотореакционный центр: 1 – свет; 2 – светособирающие пигменты; 3 – донор электрона; 4 акцептор электрона
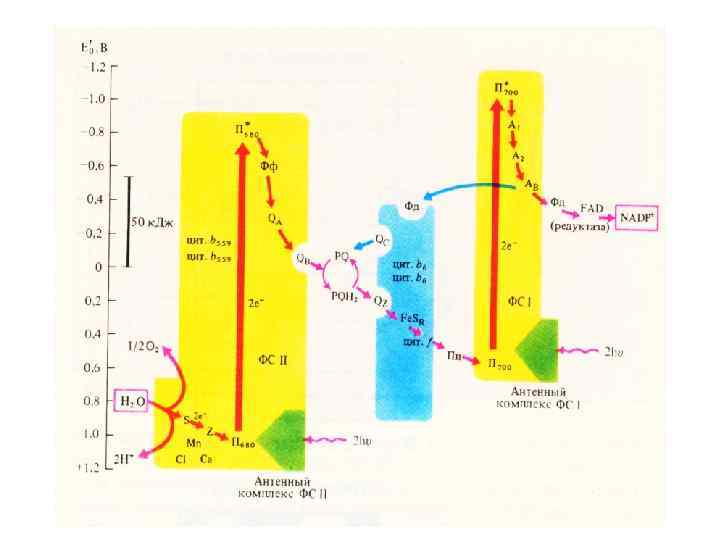
 Имеются доказательства существования двух типов фотосистем. В фотосистеме I реакционный центр образован специфической молекулой хлорофилла а и обозначается как Р 700(Р означает пигмент, от англ. pigment; 700 — оптимум поглощения, нм). Реакционный центр фотосистемы II тоже образован специфической молекулой хлорофилла а и обо значается Р 680 (оптимум поглощения — при 680 нм). В целом обе фотосистемы работают синхронно и непрерывно. ССК и фотосинтетическая единица. В листе все светособирающие хлорофиллы нековалентно связаны с белками.
Имеются доказательства существования двух типов фотосистем. В фотосистеме I реакционный центр образован специфической молекулой хлорофилла а и обозначается как Р 700(Р означает пигмент, от англ. pigment; 700 — оптимум поглощения, нм). Реакционный центр фотосистемы II тоже образован специфической молекулой хлорофилла а и обо значается Р 680 (оптимум поглощения — при 680 нм). В целом обе фотосистемы работают синхронно и непрерывно. ССК и фотосинтетическая единица. В листе все светособирающие хлорофиллы нековалентно связаны с белками.
 В фотосистеме I при поглощении света хлорофиллом энергия электронов возрастает до уровня, на котором они могут восстанавливать неидентифицированный акцептор электронов X. От него электроны через ферредоксин (Fd) передаются НАДФ и восстанавливают его. Этот восстановленный НАДФ сам может затем поставлять электроны, необходимые для восстановления атмосферного СО 2 до глюкозы. Таким путем световая энергия превращается в химическую. Источником протонов для синтеза НАДФH является вода.
В фотосистеме I при поглощении света хлорофиллом энергия электронов возрастает до уровня, на котором они могут восстанавливать неидентифицированный акцептор электронов X. От него электроны через ферредоксин (Fd) передаются НАДФ и восстанавливают его. Этот восстановленный НАДФ сам может затем поставлять электроны, необходимые для восстановления атмосферного СО 2 до глюкозы. Таким путем световая энергия превращается в химическую. Источником протонов для синтеза НАДФH является вода.
 В фотосистеме II электроны хлорофилла при поглощении света переходят на энергетический уровень, на котором они могут присоединяться к акцептору электронов Q C 550. В фотосистеме II богатые энергией электроны возбужденного хлорофилла передаются системе переноса электронов и замещаются бедными энергией электронами, поступающими от воды.
В фотосистеме II электроны хлорофилла при поглощении света переходят на энергетический уровень, на котором они могут присоединяться к акцептору электронов Q C 550. В фотосистеме II богатые энергией электроны возбужденного хлорофилла передаются системе переноса электронов и замещаются бедными энергией электронами, поступающими от воды.
 Нециклический поток электронов
Нециклический поток электронов
 Эти электроны передаются затем в серии окислительно восстановительных реакций, включающих пластохинон, цитохром и пластоцианин, хлорофиллу фотосистемы I.
Эти электроны передаются затем в серии окислительно восстановительных реакций, включающих пластохинон, цитохром и пластоцианин, хлорофиллу фотосистемы I.
 Компоненты этой цепи световых реакций напоминают компоненты электронотранспортной цепи дыхания; в се состав входят цитохромы, белки, содержащие железо и серу, хиноны, а также хлорофилл и белок пластоцианин, содержащий медь. Электронотранспортная цепь между фотосистемами организована так, что АТР может образовываться из ADP и Р(, причем этот процесс аналогичен окислительному фосфорилированию, происходящему в митохондриях. В хлоропластах этот процесс называется фотофосфоршшрованием
Компоненты этой цепи световых реакций напоминают компоненты электронотранспортной цепи дыхания; в се состав входят цитохромы, белки, содержащие железо и серу, хиноны, а также хлорофилл и белок пластоцианин, содержащий медь. Электронотранспортная цепь между фотосистемами организована так, что АТР может образовываться из ADP и Р(, причем этот процесс аналогичен окислительному фосфорилированию, происходящему в митохондриях. В хлоропластах этот процесс называется фотофосфоршшрованием
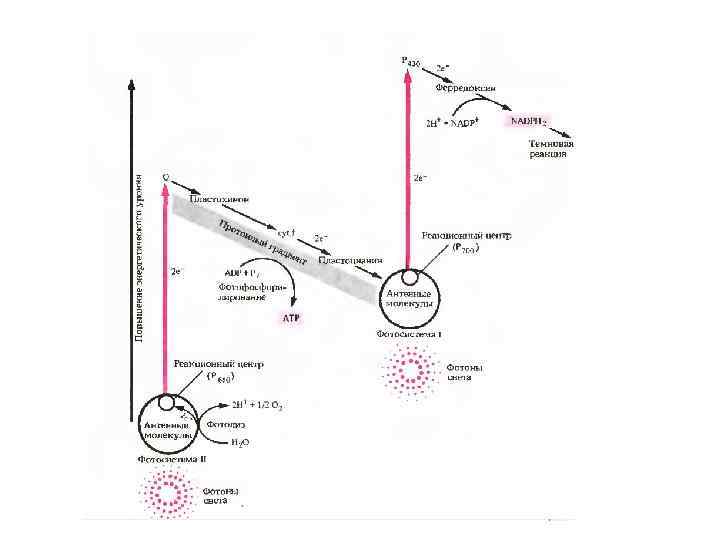
 показывает путь потока электронов") Нециклический поток электронов и фотофосфорилирование. Эта зигзагообразная схема (Z схема) показывает путь потока электронов от воды (внизу слева) к NADP (вверху справа) и энергетические взаимоотношения. Чтобы поднять энергию электронов от воды до уровня, необходимого для восстановления NADP до NADPH 2, электроны должны высвобождаться дважды (коричневые линии) под действием фотонов света, поглощаемого фотосистемами I и II. После этого высокоэнергетические потоки электронов спускаются по путям, указанным черными стрелками. Процесс фотофосфорилирования ADP с образованием А ТР сопряжен с потоком электронов в электронотранспортной цепи, связывающей фотосистему II с фотосистемой I. Ферредоксин является донором электронов для восстановления NADP до NADPH Кроме того, ферредоксин выполняет и другие функции в хлоропласте: он отдает свои электроны ферментам, участвующим в биосинтезе аминокислот и жирных кислот
Нециклический поток электронов и фотофосфорилирование. Эта зигзагообразная схема (Z схема) показывает путь потока электронов от воды (внизу слева) к NADP (вверху справа) и энергетические взаимоотношения. Чтобы поднять энергию электронов от воды до уровня, необходимого для восстановления NADP до NADPH 2, электроны должны высвобождаться дважды (коричневые линии) под действием фотонов света, поглощаемого фотосистемами I и II. После этого высокоэнергетические потоки электронов спускаются по путям, указанным черными стрелками. Процесс фотофосфорилирования ADP с образованием А ТР сопряжен с потоком электронов в электронотранспортной цепи, связывающей фотосистему II с фотосистемой I. Ферредоксин является донором электронов для восстановления NADP до NADPH Кроме того, ферредоксин выполняет и другие функции в хлоропласте: он отдает свои электроны ферментам, участвующим в биосинтезе аминокислот и жирных кислот
 Циклический поток электронов B&MBP, 2000
Циклический поток электронов B&MBP, 2000
 внутрь (тылакоидное пространство) с помощью") Протоны «накачиваются» через тилакоидную мембрану из стромы (внешняя среда) внутрь (тылакоидное пространство) с помощью переносчиков электронов, определенным образом расположенных в мембране. Концентрация протонов в тилакоидном пространстве увеличивается частично за счет расщепления воды, частично в результате окисления пластохинона (PQ) на внутренней стороне мембраны. Когда протоны идут обратно по градиенту из тилакоидного пространства в строму, ADP фосфорилируется до АТР с помощью АТР синтетазы. На каждые три протона, которые проходят через АТР синтетазу, синтезируется одна молекула АТР.
Протоны «накачиваются» через тилакоидную мембрану из стромы (внешняя среда) внутрь (тылакоидное пространство) с помощью переносчиков электронов, определенным образом расположенных в мембране. Концентрация протонов в тилакоидном пространстве увеличивается частично за счет расщепления воды, частично в результате окисления пластохинона (PQ) на внутренней стороне мембраны. Когда протоны идут обратно по градиенту из тилакоидного пространства в строму, ADP фосфорилируется до АТР с помощью АТР синтетазы. На каждые три протона, которые проходят через АТР синтетазу, синтезируется одна молекула АТР.
 АТФ азный комплекс Механизм работы АТФ синтазы Ротационноый механизм. O – open «открыто» , T tight «закрыто» и L – loose «слабосвязанно»
АТФ азный комплекс Механизм работы АТФ синтазы Ротационноый механизм. O – open «открыто» , T tight «закрыто» и L – loose «слабосвязанно»
 Циклическое фотофосфорилирование Фотосистема I может работать независимо от фотосистемы II. В этом процессе, называемом циклическим потоком электронов, электроны передаются от Р 700 на Р 430 при освещении фотосистемы I. Вместо того чтобы идти к NADP, электроны идут по «запасному пути» , связанному с I и II фотосистемами, и затем возвращаются в реакционный центр фотосистемы I.
Циклическое фотофосфорилирование Фотосистема I может работать независимо от фотосистемы II. В этом процессе, называемом циклическим потоком электронов, электроны передаются от Р 700 на Р 430 при освещении фотосистемы I. Вместо того чтобы идти к NADP, электроны идут по «запасному пути» , связанному с I и II фотосистемами, и затем возвращаются в реакционный центр фотосистемы I.
 Предполагают, что это самый примитивный механизм, и, очевидно, он имеет место у некоторых фотосинтезирующих бактерий. Эукариотические клетки тоже способны синтезировать АТР при циклическом переносе электронов. Однако при этом не происходит разложения Н 20, выделения 02 и образования NADPH 2. Полагают, что циклический транспорт электронов и фотофосфорилирование происходят в том случае, когда клетка с избытком снабжается восстановителем в форме NADPH 2, но при этом требуется дополнительный АТР для других метаболических реакций.
Предполагают, что это самый примитивный механизм, и, очевидно, он имеет место у некоторых фотосинтезирующих бактерий. Эукариотические клетки тоже способны синтезировать АТР при циклическом переносе электронов. Однако при этом не происходит разложения Н 20, выделения 02 и образования NADPH 2. Полагают, что циклический транспорт электронов и фотофосфорилирование происходят в том случае, когда клетка с избытком снабжается восстановителем в форме NADPH 2, но при этом требуется дополнительный АТР для других метаболических реакций.
 ТЕМНОВЫЕ РЕАКЦИИ ФОТОСИНТЕЗА
ТЕМНОВЫЕ РЕАКЦИИ ФОТОСИНТЕЗА
 На второй стадии фотосинтеза химическая энергия, запасенная в световых реакциях, используется для восстановления углерода. Углерод, доступный для фотосинтезирующих клеток, поставляется в виде двуокиси углерода. Обнаружено, что водоросли и цианобактерии усваивают двуокись углерода, растворенную в воде. У большинства растений С 02 поступает к фотосинтезирующим клеткам через специальные отверстия, называемые устьицами, которые находятся в листьях и зеленых стеблях.
На второй стадии фотосинтеза химическая энергия, запасенная в световых реакциях, используется для восстановления углерода. Углерод, доступный для фотосинтезирующих клеток, поставляется в виде двуокиси углерода. Обнаружено, что водоросли и цианобактерии усваивают двуокись углерода, растворенную в воде. У большинства растений С 02 поступает к фотосинтезирующим клеткам через специальные отверстия, называемые устьицами, которые находятся в листьях и зеленых стеблях.
 Цикл Кальвина: С 3 путь Восстановление углерода происходит в строме хлоропласта в цикле реакций, которые известны как цикл Кальвина (названного в честь Мелвина Кальвина, который получил Нобелевскую премию за его открытие). Цикл Кальвина аналогичен циклу Кребса, поскольку в конце цикла происходит регенерация исходного соединения. Исходное соединение цикла Кальвина — пятиуглеродный сахар с двумя фосфатными группами — рибулозо 1, 5 бисфосфат (РБФ). Процесс начинается, когда двуокись углерода входит в цикл и фиксируется на РБФ. Образуемое при этом соединение затем расщепляется на две молекулы 3 фосфоглицерата. Каждая молекула 3 фосфоглицерата состоит из трех атомов углерода, отсюда другое название цикла Кальвина — С 3 путь. Рибулозобисфосфаткарбоксилаза — фермент, катализирующий эти ключевые реакций, — самый распространенный в хлоропласте; его содержание вставляет 15% от общего количества белка хлоропласта. Данный фермент локализован на поверхности тилакоидных мембран.
Цикл Кальвина: С 3 путь Восстановление углерода происходит в строме хлоропласта в цикле реакций, которые известны как цикл Кальвина (названного в честь Мелвина Кальвина, который получил Нобелевскую премию за его открытие). Цикл Кальвина аналогичен циклу Кребса, поскольку в конце цикла происходит регенерация исходного соединения. Исходное соединение цикла Кальвина — пятиуглеродный сахар с двумя фосфатными группами — рибулозо 1, 5 бисфосфат (РБФ). Процесс начинается, когда двуокись углерода входит в цикл и фиксируется на РБФ. Образуемое при этом соединение затем расщепляется на две молекулы 3 фосфоглицерата. Каждая молекула 3 фосфоглицерата состоит из трех атомов углерода, отсюда другое название цикла Кальвина — С 3 путь. Рибулозобисфосфаткарбоксилаза — фермент, катализирующий эти ключевые реакций, — самый распространенный в хлоропласте; его содержание вставляет 15% от общего количества белка хлоропласта. Данный фермент локализован на поверхности тилакоидных мембран.

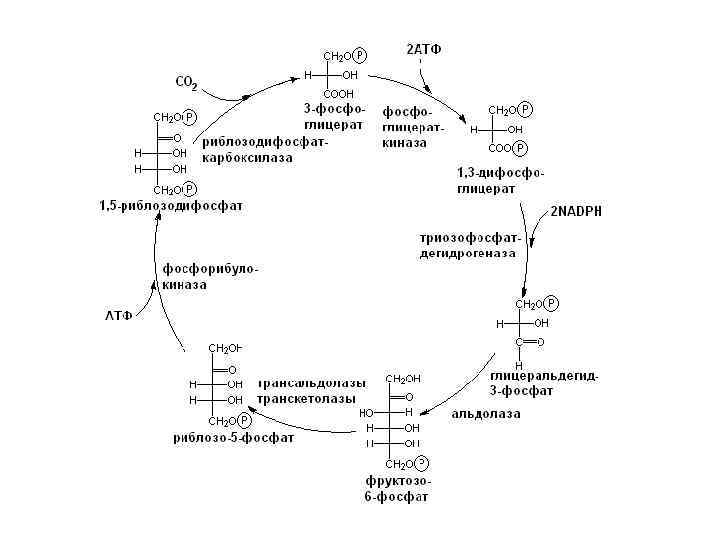
 Этапы цикла Кальвина 1. Карбоксилирование. Молекулы рибулозо 5 фосфата фосфорилируются с участием АТР и фосфорибулозокиназы, в результате чего образуются молекулы рибулозо 1, 5 дифосфата, к которым в свою очередь присоединяется СО 2 с помощью рибулозодифосфаткарбоксилазы. Полученный продукт расщепляется на две триозы: 2 молекулы 3 фосфо глидериновой кислоты (3 ФГК). 2. Фаза восстановления. 3 ФГК восстанавливается до 3 фосфоглицеринового альдегида (3 ФГА) в два этапа. Сначала происходит фосфорилирование 3 ФГК при участии АТР и фосфоглицераткиназы до 1, 3 дифосфоглицериновой кислоты, а затем восстановление 1, 3 ФГК с помощью NADPH и дегидрогеназы фосфоглицеринового альдегида.
Этапы цикла Кальвина 1. Карбоксилирование. Молекулы рибулозо 5 фосфата фосфорилируются с участием АТР и фосфорибулозокиназы, в результате чего образуются молекулы рибулозо 1, 5 дифосфата, к которым в свою очередь присоединяется СО 2 с помощью рибулозодифосфаткарбоксилазы. Полученный продукт расщепляется на две триозы: 2 молекулы 3 фосфо глидериновой кислоты (3 ФГК). 2. Фаза восстановления. 3 ФГК восстанавливается до 3 фосфоглицеринового альдегида (3 ФГА) в два этапа. Сначала происходит фосфорилирование 3 ФГК при участии АТР и фосфоглицераткиназы до 1, 3 дифосфоглицериновой кислоты, а затем восстановление 1, 3 ФГК с помощью NADPH и дегидрогеназы фосфоглицеринового альдегида.
 З. Фаза регенерации первичного акцептора диоксида углерода и синтеза конечного продукта фотосинтеза. В результате описанных выше реакций при фиксации трех молекул СО 2 и образовании шести молекул восстановленных 3 фосфотриоз пять из них используются затем для регенерации рибулозо 5 фосфата, а один — для синтеза глюкозы. 3 ФГА под действием триозофосфатизомеразы изомеризуется в фосфодиоксиацетон. При участии альдолазы 3 ФГА и фосфодиоксиацетон конденсируются с образованием фруктозо 1, 6 дифосфата, у которого отщепляется один фосфат с помощью фруктозо 1, 6 дифосфатазы. В дальнейших реакциях, связанных с регенера цией первичного акцептора СО 2, последовательно принимают участие транскетолаза и альдолаза. Транскетолаза катализирует перенос содержащего два углерода гликолевого альдегида от кетозы на альдозу. Таким образом, для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 NADPH и 18 АТР, которые поставляются в результате фотохимических реакций фотосинтеза.
З. Фаза регенерации первичного акцептора диоксида углерода и синтеза конечного продукта фотосинтеза. В результате описанных выше реакций при фиксации трех молекул СО 2 и образовании шести молекул восстановленных 3 фосфотриоз пять из них используются затем для регенерации рибулозо 5 фосфата, а один — для синтеза глюкозы. 3 ФГА под действием триозофосфатизомеразы изомеризуется в фосфодиоксиацетон. При участии альдолазы 3 ФГА и фосфодиоксиацетон конденсируются с образованием фруктозо 1, 6 дифосфата, у которого отщепляется один фосфат с помощью фруктозо 1, 6 дифосфатазы. В дальнейших реакциях, связанных с регенера цией первичного акцептора СО 2, последовательно принимают участие транскетолаза и альдолаза. Транскетолаза катализирует перенос содержащего два углерода гликолевого альдегида от кетозы на альдозу. Таким образом, для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 NADPH и 18 АТР, которые поставляются в результате фотохимических реакций фотосинтеза.
 Шесть оборотов цикла с поглощением шести атомов углерода необходимо совершить, чтобы образовался шестиуглеродный сахар — глюкоза. Суммарное уравнение синтеза глюкозы можно записать следующим образом: Суммарная реакция цикла Кальвина: 18 АТФ + 12 НАДФH + 6 CO 2 → C 6 H 12 O 6 + 18 H 3 PO 4 + 18 АДФ + 6 H 2 O + 12 НАДФ Таким образом, суммарная реакция фотосинтеза имеет следующий вид: 6 СО 2 + 12 Н 2 О + hν → С 6 Н 12 О 6 + 6 О 2 ↑ + 6 Н 2 О
Шесть оборотов цикла с поглощением шести атомов углерода необходимо совершить, чтобы образовался шестиуглеродный сахар — глюкоза. Суммарное уравнение синтеза глюкозы можно записать следующим образом: Суммарная реакция цикла Кальвина: 18 АТФ + 12 НАДФH + 6 CO 2 → C 6 H 12 O 6 + 18 H 3 PO 4 + 18 АДФ + 6 H 2 O + 12 НАДФ Таким образом, суммарная реакция фотосинтеза имеет следующий вид: 6 СО 2 + 12 Н 2 О + hν → С 6 Н 12 О 6 + 6 О 2 ↑ + 6 Н 2 О
 Карбоксилирование ФГК РБФ С 3 С 5 РБФК ФГК С 3
Карбоксилирование ФГК РБФ С 3 С 5 РБФК ФГК С 3
 Восстановление СООН НСОН АТФ Н 2 СО Р СОО Р НСОН Н 2 СО Р О НАДФН С-Н НСОН 3 -ФГА Н 2 СО Р 3 -ФГК 1, 3 -ФГК Н 2 С - ОН Транскетолазные и трансальдолазные реакции Регенерация субстрата С=О Н 2 СО Р ДГАФ
Восстановление СООН НСОН АТФ Н 2 СО Р СОО Р НСОН Н 2 СО Р О НАДФН С-Н НСОН 3 -ФГА Н 2 СО Р 3 -ФГК 1, 3 -ФГК Н 2 С - ОН Транскетолазные и трансальдолазные реакции Регенерация субстрата С=О Н 2 СО Р ДГАФ
 Дальний транспорт продуктов ФС 25% общего V листа Общая длина проводящих пучков – 100% Крупные пучки – 5% Флоэма: симпластный транспорт Ксилема: апопластный транспорт Мелкие жилки – 95%
Дальний транспорт продуктов ФС 25% общего V листа Общая длина проводящих пучков – 100% Крупные пучки – 5% Флоэма: симпластный транспорт Ксилема: апопластный транспорт Мелкие жилки – 95%
 SEL: Ситовидные") Загрузка и разгрузка флоэмы Ситовидные трубки, клетки спутники, перенхимные клетки (передаточные, запасные) SEL: Ситовидные трубки флоэмы 10 к. Да Клетки спутники 10 к. Да
Загрузка и разгрузка флоэмы Ситовидные трубки, клетки спутники, перенхимные клетки (передаточные, запасные) SEL: Ситовидные трубки флоэмы 10 к. Да Клетки спутники 10 к. Да