Biology-7.1.ppt
- Количество слайдов: 22



ФОТОСИНТЕЗ

Фотосинтез представляет собой ряд фотохимических и биохимических процессов, в результате которых живые существа синтезируют органические вещества за счет энергии солнечного света.
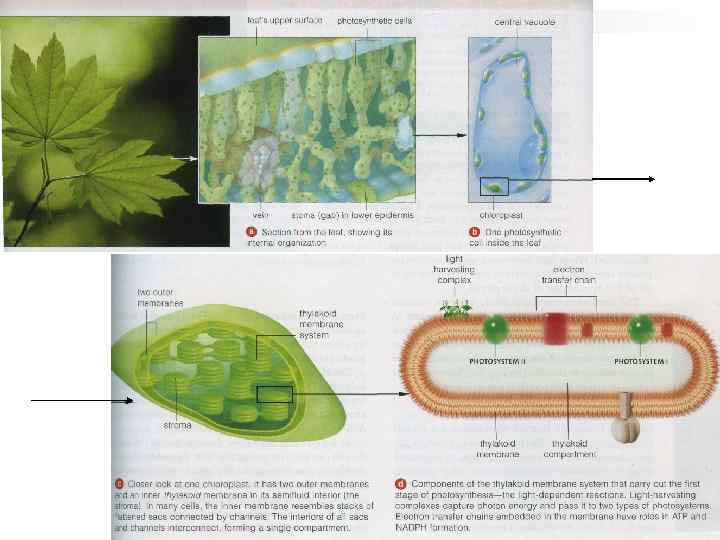

В зеленых водорослях и высших растениях фотосинтез происходит в хлоропластах. Это органеллы, которые, подобно митохондриям, окружены двумя мембранами и содержат собственную ДНК. Во внутреннем пространстве (строме) находятся тилакоиды, уплощенные мембранные мешки, которые будучи сложены стопками образуют граны. Внутреннее содержимое тилакоида называют люменом. Световые реакции катализируются ферментами тилакоидной мембраны, в то время как темновые реакции происходят в строме.

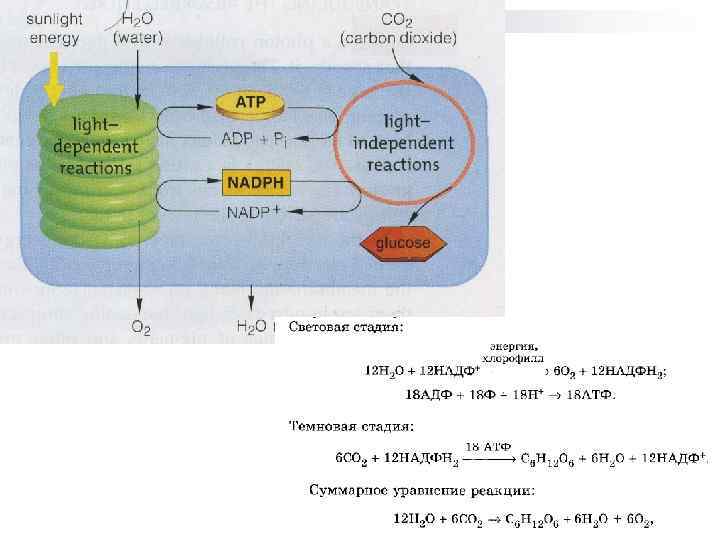
Как и в дыхательной цепи в световых реакциях электроны переносятся по электронтранспортной цепи. Однако в дыхательной цепи электроны переносятся с НАДН на О 2 с образованием воды и выделением энергии, а при фотосинтезе электроны переносятся с воды на НАДФ+ при затрате энергии. Таким образом, фотосинтетический перенос электронов в энергетическом отношении подобен «подъему в гору» . Возбуждение электронов за счет энергии поглощенного света происходит в двух реакционных центрах (фотосистемах). Это белковые комплексы, содержащие молекулы хлорофилла и разных пигментов. Другим компонентом транспортной цепи является комплекс цитохрома b/f. Функции мобильных переносчиков электронов выполняют подобный убихинону пластохинон и два растворимых белка — медьсодержащий пластоцианин и ферредоксин.

Так как фотосистема II и комплекс цитохрома b/f передают протоны от восстановленного пластохинона в люмен, фотосинтетический электронный транспорт формирует электрохимический градиент, который используется АТФсинтазой для образования АТФ.

Возбуждение Хлорофилл — зеленый, содержащий ионы Mg 2+ тетрапиррольный пигмент, несущий дополнительно остаток фитола.

В пиррольном кольце молекул хлорофиллов к остатку пропионовой кислоты обычно присоединены остатки высокомолекулярных изопреноидных спиртов которые придают хлорофиллам способность встраиваться в липидные слои мембран хлоропластов. хлорофилл a: R 1 = СН — СН 2, R 2 = СН 3, R 3 = С 2 Н 5, R 4 = CH 2 C(0)Y Хлорофилл b: R 1 = СН – СН 2, R 2 = СНО, R 3 = C 2 H 5, R 4 = CH 2 C(O)Y Хлорофилл d: R 1 = СНО, R 2 = СН 3, R 3 = С 2 Н 5, R 4 = CH 2 C(O)Y

Из высших растений, водорослей и фотосинтезирующих бактерий выделено и структурно охарактеризовано свыше 50 различных хлорофиллов. Высшие растения и зелёные водоросли содержат хлорофилл а и b, бурые и диатомовые водоросли — а и с, красные водоросли — а и d. В фотосинтезирующих бактериях присутствуют близкие аналоги — бактериохлорофиллы (БХ), отличающиеся от растительных хлорофиллов типом макроцикла и замещающими группами. Они имеют несколько модификаций: так, из пурпурных бактерий выделены БХ а и b, из зеленых бактерий - БХ а, с, d и е, из серных бактерий - БХ с, d и е; обнаружены также фотосинтезирующие бактерии, содержащие БХ g.
 Хлорофилл а")
Пигмент Природный источник x 10 -3 или отношение интенсивностей к основному пику) Хлорофилл а Все аэробные организмы 662(90), 516(15), 578(8), 534(4), 430(118), 410(76) Хлорофилл b Зеленые растения, водоросли 644(56), 595(12), 549(6), 455(159), 430(57) Хлорофилл с1 Бурые водоросли 628(1, 0)*, 578(0, 6), 444(9, 9) Хлорофилл с2 Бурые водоросли 629(1, 0)*, 582(1 , 2), 448(14, 1) Хлорофилл с3 Бурые водоросли 626(1, 0)*, 585(3, 8), 451(32, 1) Хлорофилл d Хлорелла — Бактериохло рофилл а Пурпурные бактерии 773(91), 697(9), 577(21), 530(3), 391(48), 358(73) Бактериохло рофилл b Пурпурные бактерии 794(100), 676(18), 578(25), 408(78), 368(81) Бактериохло рофилл g Азотфиксирующие бактерии 763(1, 0)*, 575(0, 4), 470(0, 5), 418(1, 9), 408(2, 0) Бактериохло рофилл с Зеленые и бурые бактерии 668(64), 624(14), 574(11), 431(100), 384(65), 356(56) Бактериохло рофилл d Зеленые и бурые бактерии 654(61), 608(17), 424(100), 408(87), 330(45) Бактериохло рофилл е Зеленые и бурые бактерии 647(34), 592(19), 458(100), 337(48) НЕКОТОРЫЕ ХАРАКТЕРИСТИКИ ХЛОРОФИЛЛОВ И БАКТЕРИОХЛОРОФИЛЛОВ

При освещении светособирающие пигменты выполняют функцию фотоантенн, которые улавливают кванты света и передают энергию возбуждения молекуле хлорофилла а, расположенной в реакционном центре.

Фотосинтетический перенос электронов у растений начинается с фотосистемы II, которая содержит реакционный центр P 680 (хлорофилл а ), феофитин (хлорофилл, не содержащий ионов Mg 2+), два связанных хинона (QA и QB) и пластохинон (QP). Только около 1% молекул хлорофилла в ФС II непосредственно участвуют в фотохимическом переносе электронов. Основная часть связана с другими пигментами в так называемом комплексе светособирающей антенны (окрашен в зеленый цвет).

Поступающая от светособирающей антенны анергия света переводит электрон реакционного центра молекулы хлорофилла в возбужденное «синглетное состояние» . Возбужденный электрон немедленно переносится на соседний феофитин. Вследствие этого в реакционном центре остается «электронная дыра» , т. е. положительно заряженный радикал Р 680. Эта дыра заполняется электроном, который отнимается от молекулы водорасщепляющим ферментом. Возбужденный электрон переносится с феофитина через QA на акцептор QB, переводя его в состояние радикала (семихиноновый радикал). QB полностью восстанавливается вторым возбужденным электроном и, наконец, обменивается на окисленный пластохинон (QP).

У современных растений функционируют 2 фотосистемы: ФС-1 поглощает свет с длиной волны 700 -730 нм и впервые возникла у фототрофных бактерий. ФС-2 поглощает свет с д. в. 680 -700 нм и появилась позже у цианобактерий. Донором электронов для фотосистемы 2 является вода, а для фотосистемы 1 – ЭТЦ фотосистемы 2.
,")
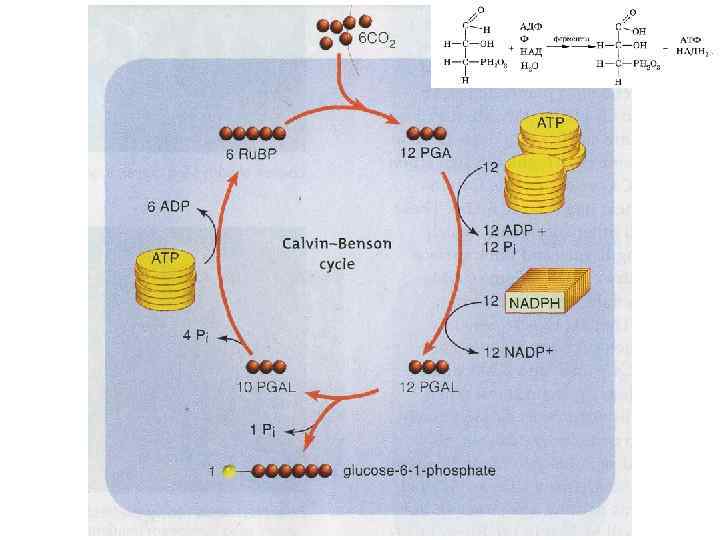
Циклический перенос электронов Нециклический перенос электронов При работе 2 -х фотосистем (при нормальном освещении), осуществляется нециклический перенос электронов. При отсутствии коротковолновых лучей протекает циклический перенос электронов. В этом случае не происходит восстановление НАДФ+ но, благодаря транслокации протонов, синтезируется АТФ. Источником электронов и протонов для ФС-1 служат восстановленые органические соединения. У бактерий это могут быть Н 2 S, Н 2 и др. Поэтому в процессе бактериального фотосинтеза выделение О 2 не происходит. a/L. d.

Плазматическая мембрана бактерий семейства Halobacterium содержит белок, подобный родопсину глаза и потому названный бактериородопсином. Этот белок способен непосредственно использовать энергию солнечного света для создания электрохимического градиента. В основе процесса, как и при зрительном процессе, лежит индуцируемая светом цис-трансизомеризация ретиналя. Белковая часть молекулы бактериородопсина в основном состоит из 7 α-спиралей, пронизывающих мембрану и образующих полый цилиндр. Внутри цилиндра расположена молекула ретиналя. В темноте ретиналь находится полностью в транс-форме. При освещении ретиналь перегруппировывается в 13 -цисформу и отдает протон, который «откачивается» наружу двумя аспартатными остатками.

, использующий энергию света.")
Протонные градиенты формируются различными способами. Необычным протонным насосом является бактериородопсин (1), использующий энергию света. При фотосинтезе восстановленный пластохинон (QH 2) переносит протоны вместе с электронами через мембрану (Q-цикл) (2). Образование протонного градиента в дыхательной цепи также сопряжено с окислительно-восстановительным процессом. В комплексе III, повидимому, как и при фотосинтезе, за перенос протона ответствен Q-цикл. В цитохром с-оксидазе (комплекс IV, 3) H+-транспорт сопряжен с электронным потоком от цитохрома с на О 2. В каждом из этих случаев протонный градиент используется в синтезе АТФ-синтазой (4).

Из высших растений, водорослей и фотосинтезирующих бактерий выделено и структурно охарактеризовано свыше 50 различных хлорофиллов. Высшие растения и зелёные водоросли содержат хлорофилл а и b, бурые и диатомовые водоросли — а и с, красные водоросли — а и d. В фотосинтезирующих бактериях присутствуют близкие аналоги — бактериохлорофиллы (БХ) В большинстве фотосинтезирующих бактерий обнаружены отличающиеся от хлорофилла а типом макроцикла и замещающими группами в цикле. Они имеют несколько модификаций: так, из пурпурных бактерий выделены БХ а и b, из зеленых бактерий - БХ а, с, d и е, из серных бактерий - БХ с, d и е; обнаружены также фотосинтезирующие бактерии, содержащие БХ g. Из бурых и диатомовых водорослей выделены хлорофиллы а и с, из красных морских водорослей - хлорофиллы а и d. Формально хлорофиллы представляют собой производные порфина, молекулы к-рых содержат циклопентаноновое кольцо, конденсированное с порфириновым макроциклом, центральный атом Mg и разл. заместители; одно или два пиррольных цикла в молекулах частично гидрированы, см. , напр. , ф-лу I
Biology-7.1.ppt