Общ микробиол Лекция 3-5 ИН Строение клетки.ppt
- Количество слайдов: 125



Формы и размеры бактериальных клеток

Прокариоты • C середине XX века стало понятно, что бактерии обладают уникальной клеточной организацией • Все организмы в настоящее время делят по строению клеток на прокариоты и эукариоты • Клетки прокариот не имеют оформленного ядра, окруженного ядерной мембраной • Генетический материал, представлен, как правило, кольцевой молекулой ДНК, которая находится у прокариот прямо в цитоплазме • Молекула ДНК находится в зоне, которую называют нуклеоидом • В клетках прокариот отсутствуют мембранные органоиды и эндоплазматическая сеть • Цитоплазматическая мембрана является единственной мембранной структурой у прокариот

Прокариоты и эукариоты • У эукариот имеется настоящее ядро, т. е. у них генетический материал окружен ядерной мембраной • В ядре у эукариот имеется более одной хромосомы • У клетках эукариот имеется эндоплазматический ретикулум и мембранные органоиды • Прокариоты отличаются от эукариот еще по целому ряду других признаков

Прокариотные и эукариотные микроорганизмы • Среди микроорганизмов есть как прокариотные, так и эукариотные организмы • Прокариотными являются бактерии (эубактерии = истинные бактерии) и архебактерии • К эукариотным микроорганизмам относятся грибы, микроскопические водоросли и простейшие • Понятие микроорганизмы не имеет таксономического значения, поскольку к микроорганизмам относятся представители разных крупных таксонов • Особое место среди микроорганизмов занимают вирусы, которые не имеют клеточного строения и являются неклеточной (доклеточной) формой жизни

Формы бактериальных клеток • Бактерии по форме клеток можно разделить на несколько групп, а именно: сферические, цилиндрические, спиральные, необычной формы и нитчатые • Сферические бактерии или кокки имеют округлую форму • Цилиндрические формы называют палочками • Спиральные формы подразделяют на вибрионы, спириллы и спирохеты • Подавляющее большинство бактерий – одноклеточные организмы • К многоклеточным бактериям относятся нитчатые цианобактерии и актиномицеты
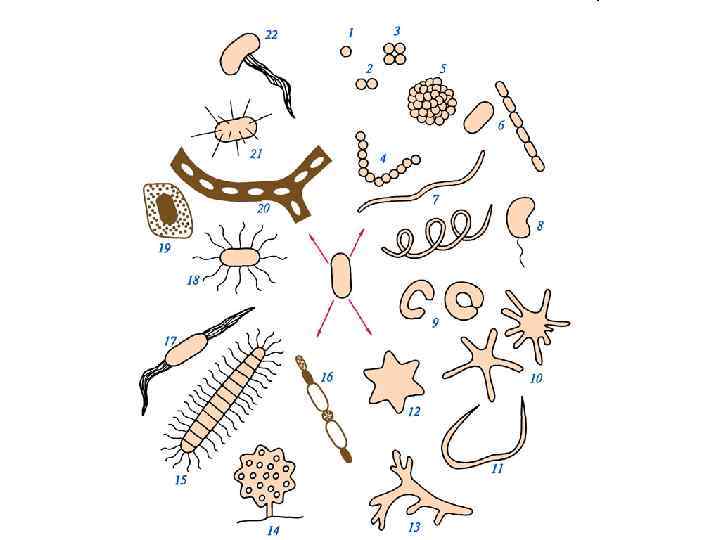

Кокки. Расположение клеток друг относительно друга • • • В зависимости от того, как клетки располагаются относительно друга после деления, кокки подразделяют на несколько групп Диплококки - кокки, которые делятся в одной плоскости, и после деления клетки располагаются попарно (например, возбудители гонореи и менингита - Neisseria gonorrhoeae, Neisseria meningitidis). Стрептококки - это кокки, которые делятся в одной плоскости, и после деления между клетками сохраняется связь, и они располагаются в виде цепочек (Streptococcus lactis, Str. faecalis, Str. pyogenes) Сарцины - это кокки, которые делятся в трех взаимно перпендикулярных плоскостях, и после деления клетки располагаются в виде пакетов из 8, 16, 32, 64 клеток, например, S. flava или они образуют колонии сферической формы Стафилококки – это кокки, которые делятся в нескольких плоскостях, после деления клетки располагаются в виде виноградной грозди, например, Staphylococcus aureus.

Кокки. Расположение клеток друг относительно друга

Диплококки

Стрептококки

Стафилококки

Палочковидные бактерии • Цилиндрическая или палочковидная форма характерна для очень многих видов бактерий • Палочковидные бактерии подразделяются на образующие эндоспоры (например, бактерии родов Bacillus и Clostridium) и не образующие эндоспоры, например, Escherichia, Pseudomonas и Xanthomonas • Палочковидные формы бактерий различаются по длине, по поперечному диаметру, по форме концов клеток • Различают палочки длинные (более 3 мкм, например Bacillus megaterium), короткие – длиной около 1 мкм (бактерии кишечной группы – например, кишечная палочка) и очень короткие (менее 1 мкм), длина которых слегка превышает диаметр клетки (их называют еще коккобактериями)

Палочковидные бактерии • По поперечному диаметру они палочковидные бактерии делятся на тонкие, например, Mycobacterium tuberculosis – туберкулезная палочка и толстые, например, Bacillus subtilis – сенная палочка • После деления клетки могут не расходиться, и тогда образуются цепочки клеток, которые называют стрептобациллы • У коринебактерий дифтерии клетки располагаются под углом друг к другу и образуют фигуры, напоминающие римские пятерки или китайский буквы

Escherichia coli

Mycobacterium tuberculosis окраска по Цилю — Нельсену

Bacillus anthracis

Палочки Расположение клеток друг относительно друга

Расположение клеток друг относительно друга у коринебактерий дифтерии ПОД УГЛОМ

Спиральные формы бактерий • Спиральные формы бактерий различаются количеством и характером завитков, длиной и толщиной клеток • Их можно подразделить на негнущиеся формы (вибрионы, спириллы) и изгибающиеся формы (спирохеты) • Вибрионы имеют вид изогнутой палочки или запятой, например Vibrio cholerae - возбудитель холеры • Спириллы – это спирально изогнутые клетки, имеющие большой поперечный диаметр и малое число высоких завитков • Спирохеты - это изгибающиеся, тонкие, спирально изогнутые клетки

Спиральные формы бактерий

Vibrio cholerae

Спирохеты Borrelia burgdorferi Treponema pallidum

Спирохета в продольном разрезе

Необычные формы бактериальных клеток • Бактериальные клетки имеют вид замкнутого или разомкнутого кольца; • Червеобразная форма; • Простекобактерии имеют клеточные выросты – простеки; • Форма шестиугольной звезды; • Ветвящиеся формы (актиномицеты). • Широко распространено среди бактерий явление плеоморфизма – в зависимости от условий среды обитания они могут иметь вид палочек, кокков или слабо ветвиться.

Размеры бактериальных клеток • Размеры бактериальных клеток в зависимости от вида могут варьировать в довольно широких пределах • Подавляющее большинство бактерий палочковидной формы имеют толщину не более 1 мкм и длину не более 5 мкм • Диаметр шаровидных бактерий обычно от 0, 5 мкм до 2 мкм • В среднем линейные размеры бактерий лежат в пределах 0, 5 -3 мкм • Клетки нитчатой серобактерии Beggiatoa alba имеют диаметр до 50 мкм, некоторые виды бактерий родов Chromatium и Achromatium имеют длину до 100 мкм • Длина клеток некоторых спирохет может достигать 500 мкм • Крупнейшей из известных бактерий является Thiomargarita namibiensis , достигающая размера в 750 мкм (0, 75 мм) • Самые мелкие из известных прокариотных клеток - бактерии, принадлежащие к группе микоплазм с диаметром клеток 0, 2 и даже 0, 1 мкм.

Строение прокариотной клетки

Строение прокариотной клетки • Бактериальная клетка состоит из протопласта, ограниченного цитоплазматической мембраной и поверхностных структур • Поверхностными структурами называют все структуры, расположенные снаружи от цитоплазматической мембраны • Цитоплазматическая мембрана вместе с цитоплазмой составляют протопласт

Поверхностные структуры • Структуры, расположенные снаружи от цитоплазматической мембраны называют поверхностными структурами – клеточная стенка – капсула – слизистый чехол – жгутики – ворсинки

Архитектура строения поверхности клетки • Архитектура строения поверхности бактериальной клетки включает: – слои биополимеров, покрывающие клетку (клеточную стенку, капсулу или слизистый чехол – выросты или придаточные структуры (жгутики, фимбрии и пили)

Структура прокариотной клетки

Клеточная стенка • Клеточная стенка – важный и обязательный структурный элемент подавляющего большинства прокариотных клеток • Исключение составляют микоплазмы, Lформы, некоторые планктомицеты и некоторые архебактерии, у которых клеточная стенка отсутствует • Клеточная стенка располагается между цитоплазматической мембраной и капсулой или слизистым чехлом • На долю клеточной стенки приходится от 5 до 50% сухих веществ клетки

Функции клеточной стенки • Клеточная стенка служит механическим барьером между протопластом и внешней средой - защитная функция • Клеточная стенка придает клеткам микроорганизмов определенную, присущую им форму – формообразовательная функция • Клеточная стенка защищает клетку от осмотического лизиса – уравновешивает гидростатическое давление • Клеточные стенки многих бактерий имеют антигены, специфические рецепторы, которые обеспечивают сигнальную функцию • Клеточная стенка выполняет транспортную функцию (пассивный транспорт веществ и ионов) и препятствует проникновению многих токсических веществ; • Клеточная стенка служит мишенью для антибиотиков и литических ферментов

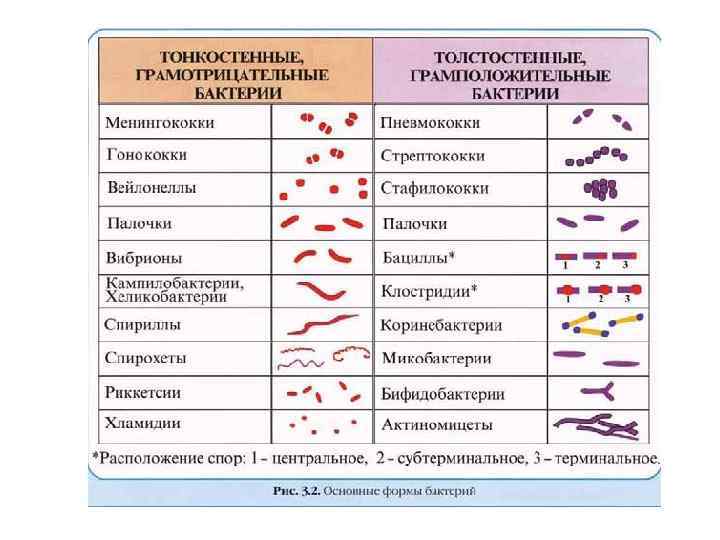
Окраска по Граму • Различия в окраске у грамположительных и грамотрицательных бактерий связаны с различиями в строении их клеточной стенки • Метод окраски, на основании которого бактерии разделяют на грамположительные и грамотрицательные предложил датский ученый Христиан Грам

Окраска по Граму • Суть метода окраски по Граму – При обработке фиксированных нагреванием клеток бактерий раствором кристаллического фиолетового, а затем йодом, образуется окрашенный комплекс – При последующей обработке мазка спиртом в зависимости от строения клеточной стенки достигается разная степень обесцвечивания мазка – У грамположительных видов бактерий окрашенный комплекс удерживается клеткой и, таким образом, бактерии остаются окрашенными в фиолетовый цвет – У грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются – На последнем этапе мазок окрашивают контрастным красителем, например фуксином, в результате чего обесцвеченные грамотрицательные бактерии приобретают розовую или красную окраску, тогда как грамположительные бактерии сохраняют фиолетовую окраску первого красителя
 Staphylococcus aureus")
Окраска по Граму (гр+) Staphylococcus aureus
 Escherichia coli")
Окраска по Граму (гр-) Escherichia coli
 и Staphylococcus aureus (гр+)")
Окраска по Граму Escherichia coli (гр-) и Staphylococcus aureus (гр+)

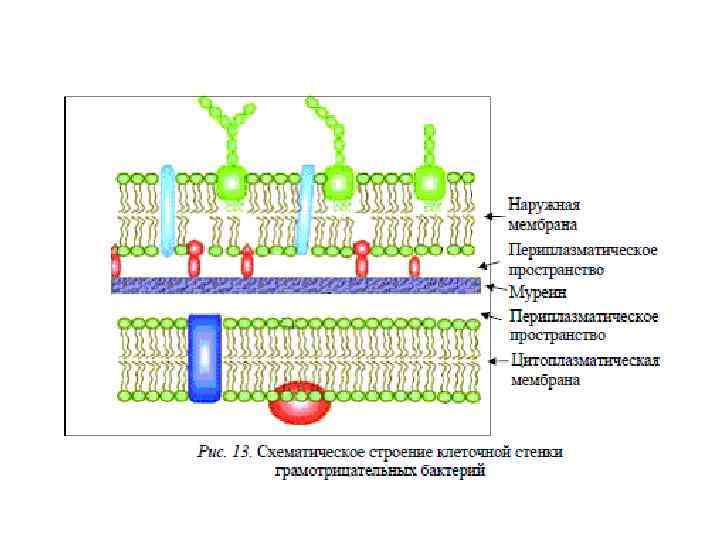
Ультраструктура клеточной стенки • То, что разные виды бактерий дают разную окраску при окрашивании по Граму связано с различиями в ультраструктуре их клеточных стенок • Под электронным микроскопом видно, что у грамположительных бактерий клеточная стенка выглядит как гомогенный (однообразный) толстый слой (толщиной 20 -80 nm), лежащий снаружи от цитоплазматической мембраны • Клеточная стенка грамотрицательных бактерий под электронным микроскопом выглядит тоньше (толщина около 10 нм) и состоит из нескольких слоев • Клеточная стенка грамотрицательных бактерий имеет так называемую внешнюю мембрану, которая похожа на типичную цитоплазматическую мембрану

Сравнение строения грам+ и грам- клеточных стенок

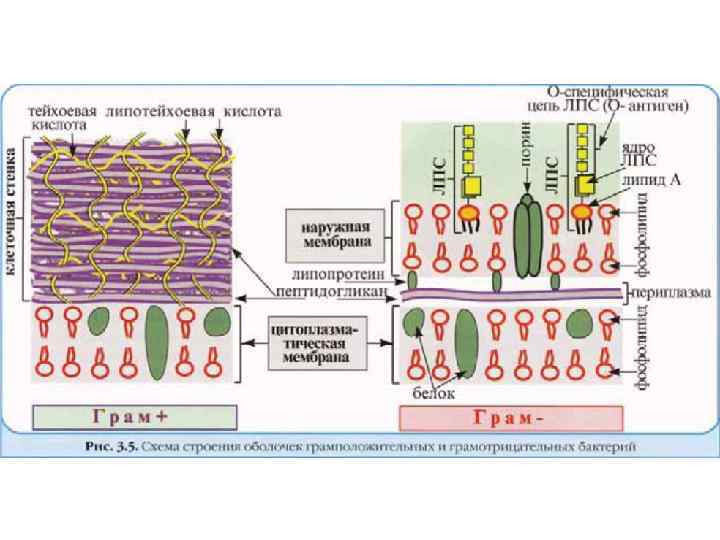
Химический состав клеточной стенки • Ультраструктурные различия связаны с различиями химического состава и молекулярной структуры клеточных стенок у грамположительных и грамотрицательных бактерий • Клеточные стенки грамположительных бактерий содержат как основной компонент пептидогликан (обычно он составляет более 30% от массы клеточной стенки). • Клеточные стенки грамположительных бактерий включают также белки, полисахариды, тейхоевые, липотейхоевые или тейхуроновые кислоты

Химический состав клеточной стенки • Клеточные стенки грамотрицательных бактерий в основном состоят из липополисахаридов, фосфолипидов, белка, липопротеинов и включают относительно немного пептидогликана (обычно менее чем 10% от общей массы клеточной стенки) • У грамотрицательных организмов липиды являются обязательными компонентами клеточных стенок • Грамположительные бактерии, в отличие от грамотрицательных, содержат гораздо больше пептидогликана, и, кроме того, практически не содержат каких-либо липидов
 • Гетерополимер пептидогликан является основным компонентом клеточной стенки большинства бактерий • Пептидогликан")
Пептидогликан (муреин) • Гетерополимер пептидогликан является основным компонентом клеточной стенки большинства бактерий • Пептидогликан содержится в клеточных стенках как у грамположительных, так и у грамотрицательных бактерий • Грамположительные бактерии, в отличие от грамотрицательных, содержат гораздо больше пептидогликана • У архебактерий пептидогликан отсутствует • Структура и химический состав клеточной стенки у архебактерий уникальны • По строению клеточной стенки архебактерии отличаются от грамположительных и грамотрицательных эубактерий

Пептидогликан • Пептидогликан присутствует только в клеточных стенках эубактерий • Пептидогликана нет в каких-либо других структурах бактериальной клетки • Пептидогликан является уникальным биополимером прокариот • Пептидогликан присутствует в клеточных стенках цианобактерий • Пептидогликан не обнаружен в клеточных стенках эукариот

Биополимеры клеточных стенок эукариот • У растений основным компонентом клеточной стенки является целлюлоза • У грибов основным компонентом клеточной стенки является хитин, глюканы, маннаны или другие полисахариды • В связи с тем, что пептидогликан является важным компонентом клеточной стенки эубактерий и в то же время отсутствует у эукариот, его биосинтез является очень удобной мишенью для подавления роста болезнетворных бактерий • Такие антибиотики, как пенициллин и антибиотики цефаллоспориновой группы, избирательно подавляют рост бактерий за счет блокирования синтеза пептидогликана
, остов")
Строение пептидогликана • Пептидогликан состоит из пептидного и углеводного компонентов • Углеводный (полисахаридный), остов молекулы пептидогликана построен из чередующихся остатков Nацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных между собой посредством – 1, 4 -гликозидных связей • К N-ацетилмурамовой кислоте прикреплена короткая пептидная цепочка

Строение пептидогликана

Строение пептидогликана • Неразветвленные гетерополимерные цепи составляют основу пептидогликана • Обычная для многих бактерий толщина клеточной стенки в 30 -40 нм соответствует приблизительно 40 молекулам пептидогликана • Неразветвленные гетерополимерные цепи пептидогликана связываются между собой пептидными мостиками

Пептидные мостики между гетерополимерными цепочками; Г – N-ацетилглюкозамин: М – N-ацетилмурамовая кислота

Строение пептидных цепочек пептидогликана • Пептидные цепочки прикрепленные к Nацетилмурамовой кислоте через лактильную группу (т. е. остаток молочной кислоты) состоят из небольшого числа аминокислот, обычно 4 -х (тетрапептиды) или 5 -и (пентапептиды) • К типичным аминокислотам пептидогликана относятся L-аланин, D-глутаминовая кислота, мезодиаминопимелиновая кислота или L-лизин и Dаланин • Особенности пептидных цепочек пептидогликана: – наличие аминокислот в D-форме – высокое содержание аминокислот с двумя аминогруппами (мезо-диаминопимелиновая кислота или L-лизин)

Пептидный промежуточный мостик • Иногда связь между тетрапептидами или пентапептидами разных гликановых цепей осуществляется с помощью других аминокислот, образующих пептидный промежуточный мостик • В клеточной стенке у грамположительной бактерии Staphylococcus aureus такой промежуточный пептидный мостик между двумя пептидными цепочками образован из аминокислоты глицина • У большинства грамотрицательных бактерий, в том числе и у Escherichia coli такие промежуточные пептидные мостики отсутствуют

Пептидные мостики А – Escherichia coli; Б – Staphylococcus aureus

Муреиновый мешок • Тетрапептидными или пентапептидными цепочками и пептидными мостиками гетерополимерные цепи связаны между собой в мешкообразную гигантскую молекулу – муреиновый мешок • Муреиновый мешок является опорным каркасом клеточной стенки • На муреиновом мешке откладываются и его инкрустируют различные другие вещества (полисахариды, белки, липиды) • Частота «сшивок» пептидными мостиками в муреиновом мешке различна у разных бактерий, поскольку не все пептидные цепочки участвуют в формировании межцепочечных связей • Некоторые пептидные цепочки образуют ковалентные связи с другими молекулами, входящими в состав клеточной стенки • Часть тетрапептидных цепочек находится в свободном состоянии

Синтез пептидогликана – мишень для антибиотиков • Синтез пептидогликана особенно чувствителен к воздействию антибиотиков и других антимикробных агентов • Ингибирование любой стадии синтеза пептидогликана ослабляет клеточную стенку и может привезти к осмотическому лизису клетки • Многие антибиотики могут воздействовать на синтез пептидогликана • Пенициллин ингибирует реакции транспептидации, а бацитрацин блокирует дефосфорилирование пирофосфата бактопренола

Клеточная стенка грамположительных бактерий • У грамположительных бактерий клеточная стенка имеет однородную структуру • Она состоит из толстого многослойного (до 40 слоев) пептидогликанового комплекса, составляющего до 70% массы клеточной стенки грамположительных бактерий • Пептидогликан, окружающий протопласт грамположительных эубактерий - одна гигантская молекула, «сшитая» с помощью гликозидных и пептидных связей • Кроме пептидогликана в состав клеточных стенок грамположительных бактерий входят тейхоевые кислоты

Клеточная стенка грамположительных бактерий

Тейхоевые кислоты • Тейхоевые кислоты могут пронизывать весь пептидогликановый слой, достигая внешней поверхности клеточной стенки • В этом случае, тейхоевые кислоты могут являться основными антигенами грамположительных бактерий • Остающиеся свободные гидроксилы фосфорной кислоты придают тейхоевой кислоте свойства полианиона • Как полианионы тейхоевые кислоты определяют отрицательный поверхностный заряд клеток грамположительных бактерий

Тейхоевые кислоты • Сахарные компоненты тейхоевых кислот входят в состав рецепторов для некоторых бактериофагов и определяют возможность адсорбции фагов на клеточной поверхности • Тейхоевые кислоты не обнаружены у грамотрицательных бактерий

Клеточная стенка грамотрицательных бактерий • Клеточная стенка грамотрицательных бактерий устроена гораздо более сложно, хотя она тоньше клеточной стенки грамположительных бактерий • Клеточная стенка грамотрицательных бактерий включает тонкий пептидогликановый слой, периплазматическое пространство и так называемую внешнюю мембрану

Электронно-микроскопический снимок клеточной стенки грамотрицательных бактерий


Пептидогликановый слой грамотрицательных бактерий • Тонкий пептидогликановый слой толщиной всего 1 -3 nm неплотно прилегает к цитоплазматической мембране • Масса пептидогликанового слоя грамотрицательных бактерий составляет всего лишь 5 -10% от общей массы клеточной стенки, т. е. значительно меньше, чем у грамположительных бактерий • У E. coli толщина пептидогликанового слоя составляет около 1 nm, и он содержит один или два слоя пептидогликана с весьма редкими поперечными связями между двумя гетерополимерными цепями • Химическая структура пептидогликана грамотрицательных эубактерий в основном сходна со структурой типичного пептидогликана грамположительных эубактерий

Периплазматическое пространство • Пространство между цитоплазматической и наружной мембранами у грамотрицательных бактерий называется периплазматическим пространством • Вещество, которое заполняет периплазматическое пространство, называется периплазмой • Величина периплазматического пространства у грамотрицательных бактерий колеблется в пределах от 1 до 70 nm • Ферменты, которые заполняют периплазматическое пространство называют периплазматическими ферментами

Наружная мембрана грамотрицательных бактерий • Наружная мембрана грамотрицательных бактерий имеет сложное строение • Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеинов и липополисахаридов • Внешний и внутренний слои наружной мембраны ассиметричны • Внутренний слой наружной мембраны более гомогенный и напоминает слои обычной ЦПМ • Внешний слой наружной мембраны более гетерогенный и отличается от слоев мембран обычной ЦПМ • Наружная мембрана отличается от обычных биологических мембран и защищает клетку от химических воздействий

Липополисахариды • Наиболее необычными и специфическими компонентами наружной мембраны являются липополисахариды • Липополисахариды находятся во внешнем слое наружной мембраны и занимают около 30 -40% ее поверхности • Липополисахариды наружной мембраны клеточных стенок грамотрицательных бактерий имеют сложное молекулярное строение • Липополисахариды включают в свой состав липиды и углеводы (полисахариды) • Липополисахариды состоят из 3 -х частей: липида А, корового полисахарида и О-боковой цепи (Оантигена)

Строение липополисахаридов

Бактериальная капсула • Капсула и слизистые слои находятся снаружи от клеточной стенки • В том случае, когда слизистый слой хорошо организован и практически не смывается водой с поверхности бактериальной клетки, говорят, что бактериальная клетка имеет капсулу • Бактериальная капсула — слизистая структура, находящаяся снаружи от клеточной стенки бактерий, прочно связанная с клеточной стенкой и имеющая четко очерченные внешние границы • Капсула различима в мазках-отпечатках из патологического материала • В чистых культурах бактерий капсула образуется реже • Капсула выявляется при специальных методах окраски, создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы

Капсула стрептококков, выявляемая методом негативного контрастирования

Функции бактериальной капсулы • Бактериальные капсулы имеют множество функций • Капсулы имеют защитную функцию, поскольку они предохраняют клетки бактерий от неблагоприятных условий среды обитания • Капсулы содержат много воды и могут предохранять клетки от высыхания • Капсула создаёт дополнительный осмотический барьер • Капсула может являться источником запасных питательных веществ • Капсулы могут изолировать клетки от вирусов бактерий, а также от различных токсических для бактерий веществ • У патогенных бактерий капсула является важным фактором вирулентности

Функции бактериальной капсулы • Капсулы могут защищать клетки патогенных бактерий от фагоцитоза • Классический пример - это бактерии Streptococcus pneumoniae, образующие полисахаридную капсулу и вызывающие пневмонию • Мутантные штаммы S. pneumoniae, утратившие способность образовывать капсулу, легко разрушаются альвеолярными макрофагами и теряют способность вызывать это заболевание • Бактерии Bacillus anthracis способны выживать внутри фагоцитов, потому что их поли-D-глутаматная капсула защищает их от разрушения лизосомальными ферментами фагоцитов • Капсула и слизистые слои, покрывающие поверхность клетки, играют адгезивную функцию у многих патогенных и непатогенных бактерий

Химический состав капсул • Капсулы и слизистые слои обычно состоят из полисахаридов, однако иногда они могут состоять и из других веществ • Возбудитель сибирской язвы, Bacillus anthracis имеет капсулу, состоящую из поли-D -глутаминовой кислоты (полипептидную капсулу) • Bacillus megaterium образует касулу состоящую из полипептидов и полисахаридов

Архитектура строения поверхности клетки • Архитектура строения поверхности бактериальной клетки включает: – слои биополимеров, покрывающие клетку (клеточную стенку, капсулу или слизистый чехол – выросты или придаточные структуры (жгутики, фимбрии и пили)

Бактериальные жгутики • Бактериальные жгутики - довольно тонкие, но достаточно прочные и длинные структуры • Их длина достигает 15 -20 мкм, а толщина всего 20 нм • Жгутики бактерий значительно тоньше, чем жгутики эукариот • Бактериальные жгутики - настолько тонкие, что их невозможно увидеть в световой микроскоп, но их можно окрасить специальными красителями, так что они становятся видимыми • Смысл окраски заключается в том, что при окрашивании жгутик утолщается и становится видимым • Детальную структуру жгутика можно рассмотреть только при помощи электронного микроскопа

Характер расположения жгутиков • Виды бактерий могут различаться друг от друга по характеру расположения жгутиков на поверхности клетки • Монотрихи имеют только один жгутик • Амфитрихи имеют два одиночных жгутика на полюсах клетки • Лофотрихи имеют пучок жгутиков на одном или на обоих полюсах клетки • Перитрихи покрыты жгутиками по всей поверхности
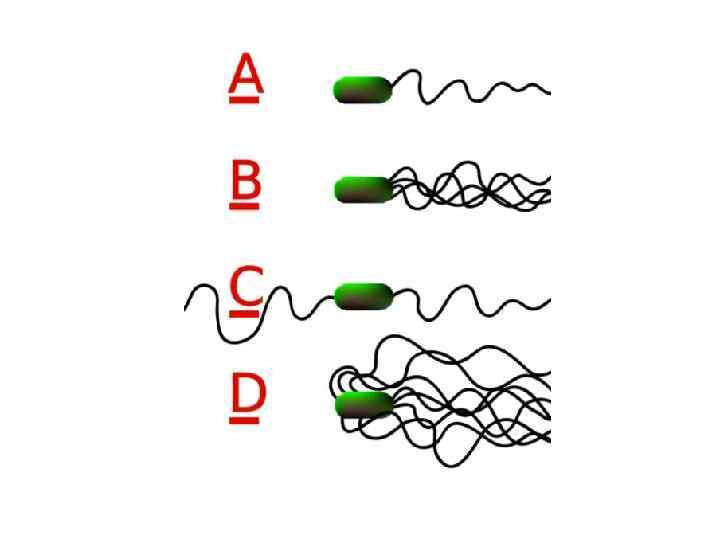

Vibrio cholerae

Патогенные факультативно анаэробные грамотрицательные палочки Shigella Salmonella

Фимбрии: их строение и адгезивные свойства • Многие грамотрицательные бактерии имеют короткие, волосоподобные выросты, которые называются фимбрии, ворсинки или пили • Фимбрии тоньше и жестче, чем жгутики • Фимбрии никак не связаны с функцией движения • Одна бактериальная клетка может нести на себе до 1000 фимбрий • Фимбрии можно увидеть только с помощью электронного микроскопа, поскольку они очень тонкие – 3 -10 нм в диаметре • Длина фимбрий составляет до 10 мкм • Фимбрии представляют собой тонкие трубочки, стенки которых построены из спирально расположенных субъединиц белка пиллина • Фимбрии помогают бактериям прикрепляться к твердым поверхностям, таким, например, как камни в водоемах или ткани в организме хозяина

Неспецифическая и специфическая адгезия • Все микроорганизмы обладают выраженной способностью прикрепляться к органическим и неорганическим поверхностям • Явление адгезии заключается в том, что клетки микроорганизмов прикрепляются или прилипают к поверхности эпителия • Различают две группы механизмов адгезии – неспецифические и специфические • Неспецифическая адгезия опосредована физикохимическими взаимодействиями бактерий с поверхностями. К ним относятся: – электростатические взаимодействия; – гидрофобные взаимодействия; – ван-дер Ваальсовы взаимодействия; – броуновское движение Неспецифическая адгезия, как правило, обратима

Специфическая адгезия • Специфическая адгезия происходит в результате молекулярных взаимодействий между адгезином микробной клетки и рецептором клетки хозяина • Под адгезинами понимают поверхностные структуры микробных клеток и входящие в их состав макромолекулы, обычно белки, посредством которых осуществляется прикрепление к специфическим поверхностям • Под рецептором подразумевают структуру, комплементарную адгезину и находящуюся на поверхности эукариотической клетки • Функцию рецепторов в процессе адгезии выполняют углеводные или пептидные (белковые) фрагменты, локализованные на мембране эукариотических клеток • У грамотрицательных микроорганизмов молекулы адгезинов входят в состав фимбрий, вследствие чего они названы фимбриальными адгезинами • Фимбриальные адгезины располагаются на кончиках фимбрий

Взаимодействие фимбриальных адгезинов бактерий и рецепторов
 • Некоторые бактерии имеют половые ворсинки или секспили (F-пили) • Секс пили")
Секс-пили (F-пили) • Некоторые бактерии имеют половые ворсинки или секспили (F-пили) • Секс пили имеют больший размер, чем обычные пили (они длиннее и толще - около 10 нм в диаметре), а количество их значительно меньше • У E coli их обычно от 1 до 10 на клетку • Секс-пили генетически детерминированы секс факторами или конъюгативными плазмидами (F-фактор у E. coli) и играют важную роль в конъюгации бактерий • Секс-пили служат «мостиками» , по которым ДНК переносится из одной клетки в другую в процессе конъюгации • Некоторые бактериофаги избирательно прикрепляются к рецепторам, находящимся на секс-пили

Конъюгация у бактерий при помощи секс-пили

Строение протопласта • Протопласт включает в себя цитоплазму, окруженную цитоплазматической мембраной • Цитоплазма – это сложная коллоидная система, которую называют также коллоидный матрикс • До 80% массы цитоплазмы вегетативных клеток составляет вода • Основными компонентами цитоплазмы являются белки, в том числе разнообразные ферменты, нуклеиновые кислоты • В цитоплазме расположены бактериальная хромосома, которая находится в зоне, которую называют нуклеоид, плазмиды, рибосомы, разнообразные включения. • Органеллы, характерные для эукариот, у бактерий отсутствуют и их функции выполняют цитоплазматическая мембрана и ее производные

Цитоплазматическая мембрана прокариот • Цитоплазматическая мембрана ограничивает протопласт, располагаясь непосредственно под клеточной стенкой • У большинства прокариотных клеток цитоплазматическая мембрана – это единственная мембрана • У прокариот отсутствуют автономные мембранные структуры, т. е. мембранные органоиды, такие как митохондрии, хлоропласты, аппарат Гольджи и др. • Однако цитоплазматическая мембрана прокариот может образовывать выпячивания внутрь прокариотической клетки, выполняющие важные функции, при этом они сохраняют связь с цитоплазматической мембраной. • Цитоплазматическая мембрана является универсальной структурой, характерной как для клеток эукариот, так и для прокариот • По своему строению мембраны бактериальных, животных и растительных клеток очень схожи •

Цитоплазматическая мембрана прокариот • • В настоящее время общепринята жидко-мозаичная модель строения мембраны, предложенная Сингером и Никольсеном Цитоплазматическая мембрана состоит из двойного липидного (фосфолипидного) слоя, в который встроены белки Белки «плавают» в этом слое, будучи погружены в него частично или же пронизывают его насквозь Липиды двойного липидного слоя представлены главным образом, фосфолипидами Гидрофобные жирнокислотные концы молекул фосфолипидов направлены внутрь, а гидрофильные «головки» наружу Таким образом, внешние поверхности фосфолипидного бислоя являются гидрофильными, а внутренняя часть мембраны является гидрофобной Благодаря гидрофобным взаимодействиям между остатками жирных кислот, входящих в состав липидов, и электростатическому взаимодействию между гидрофильными «головками» мембрана стабилизируется

Цитоплазматическая мембрана

Цитоплазматическая мембрана прокариот • Мембраны такого типа получили название элементарных • Хотя цитоплазматические мембраны прокариот имеют общий план строения, характерный для эукариотических организмов, тем не менее, они обладают некоторыми существенными отличиями • На молекулярном уровне строение цитоплазматических мембран разных бактерий может иметь определенные особенности, что может быть использовано при их идентификации • В частности большое значение имеет жирнокислотный состав мембран • ЦПМ некоторых прокариот включают специализированные белки

Функции цитоплазматической мембраны • Защитная функция. Цитоплазматическая мембрана заключает в себе цитоплазму, что особенно важно для тех видов или форм микроорганизмов, которые не имеют клеточной стенки • Барьерная функция. Цитоплазматическая мембрана обеспечивает поддержание неравенства состава и концентрации растворенных веществ в цитоплазме и во внешней среде, т. е. служит избирательным барьером. Она обеспечивает проникновение внутрь или наружу клетки одних компонентов и в то же время предотвращает проникновение других • Транспортная функция. В мембране располагаются транспортные системы необходимые для поглощения питательных веществ, т. е. транспорт веществ в клетку и удаления продуктов метаболизма, а также секреции внеклеточных белков

Функции цитоплазматической мембраны • Энергетическая функция. На цитоплазматической мембране или ее производных осуществляется перенос электронов и мембранное фосфорилирование, обеспечивающее прокариоты энергией. Часто эти процессы происходят на особых структурах, представляющих собой выпячивания цитоплазматической мембраны внутрь клетки • Биосинтетическая функция. На цитоплазматической мембране осуществляется биосинтез фосфолипидов, компонентов клеточной стенки и капсулы • Рецепторная функция. Цитоплазматическая мембрана содержат специальные рецепторные молекулы, которые позволяют бактериям отвечать на те или иные сигналы, поступающие из внешней среды

Производные цитоплазматической мембраны • К нефотосинтетическим производным цитоплазматической мембраны относятся мезосомы, а также ламеллы хемолитоавтотрофов и хемолитогетеротрофов • Мезосомы являются мембранными структурами - производными цитоплазматической мембраны, на которых локализованы компоненты дыхательной цепи • Мезосомы бывают ламеллярные (пластинчатые), тубулярные (трубчатые), или везикулярные (пузырчатые образования)

Мезосомы • Главная функция мезосом – энергетическая • На мембранах мезосом создается электрохимический градиент ионов водорода (ΔµН+) и синтезируется АТФ • Кроме того, предполагают, что мезосомы служат местом прикрепления бактериальной хромосомы, а также участвуют в синтезе материала клеточной стенки при формировании поперечной перегородки, а, следовательно, играют важную роль в делении бактериальных клеток

Ламеллы хемолитоавтотрофов и хемолитогетеротрофов • Ламеллы хемолитоавтотрофов и хемолитогетеротрофов также выполняют энергетическую функцию • Они являются местом локализации ферментов, принимающих участие в окислении неорганических и органических субстратов • У нитрифицирующих и метанолокисляющих бактерий имеется мощно развитая внутриклеточная сеть мембран, что, повидимому, связано с использованием этими бактериями газообразных субстратов • У разных видов нитрифицирующих бактерий эти структуры имеют форму ламелл, расположенных на периферии клетки параллельно клеточной стенке, по всей клетке или на одном из ее полюсов • У метанолокисляющих бактерий встречаются дисковидные стопки мембран, распределенные по всей цитоплазме, или ламеллярные структуры, расположенные параллельно клеточной стенке • Ламеллярные структуры обнаружены у метанобразующих архебактерий

Мембранные структуры метилотрофных бактерий

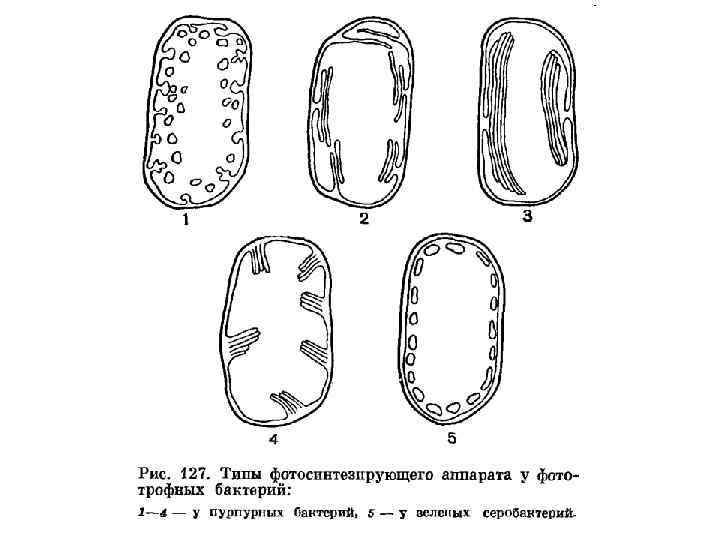
Тилакоиды • К фотосинтетическим производным цитоплазматической мембраны относятся мембранные структуры – тилакоиды, обнаруженные у фототрофных бактерий • На тилакоидах локализованы пигменты фотосинтеза (бактериохлорофиллы и каротиноиды), системы транспорта электронов и фотофосфорилирования • Тилакоиды имеют форму пузырьков или трубочек, образующих внутриклеточную мембранную сеть, сохраняющую связь с ЦПМ • У некоторых видов пурпурных бактерий они представлены стопками ламеллярных мембран • Эти структуры могут исчезать в темноте и вновь формироваться на свету

Цитоплазма • Цитоплазма – это сложная коллоидная система • До 80% массы цитоплазмы вегетативных клеток составляет вода (у спор значительно меньше) • Основными компонентами цитоплазмы являются белки, в том числе разнообразные ферменты • Состав цитоплазмы и ее свойства непрерывно изменяются в процессе жизнедеятельности клетки

Генетический аппарат прокариотической клетки • Генетический аппарат прокариотической клетки включает нуклеоид, содержащий бактериальную хромосому, и плазмиды • В цитоплазме всех бактерий содержится дезоксирибонуклеиновая кислота (ДНК), в которой закодирована наследственная информация • Зону, в которой локализована ДНК, называют нуклеоидом • Ядерной мембраны и ядрышка у бактерий не обнаружено • В нуклеоиде содержится, как правило, единственная гигантская молекула ДНК (бактериальная хромосома), имеющая форму замкнутой в кольцо двойной спирали

Плазмиды • Клетки многих бактерий могут содержать небольшие внехромосомные кольцевые молекулы ДНК, называемые плазмидами • Плазмиды в 20 -1000 раз меньше бактериальной хромосомы и способны к автономной репликации • На плазмидах закодирована дополнительная генетическая информация, которая расширяет адаптационные свойства многих бактерий
 и белков (около 40%) и")
Рибосомы • Рибосомы состоят из РНК (около 60% массы) и белков (около 40%) и имеют форму круглых или слегка удлиненных телец диаметром 15 -20 нм • Показатель седиментации (скорости осаждения) при центрифугировании бактериальных рибосом составляет 70 ед. Сведберга (условное обозначение 70 S) • Рибосомы бактерий несколько мельче и легче, чем рибосомы эукариот, в цитоплазме которых содержатся 80 S-рибосомы • Рибосомы в митохондриях и хлоропластах эукариотных клеток имеют, как и у прокариот константу седиментации 70 S • В цитоплазме бактериальной клетки присутствует большое количество (от 5000 до 90000) рибосом • Функцией рибосом является биосинтез белков

Включения • В цитоплазме прокариот обнаруживаются различные включения • Одни из включений следует рассматривать как активно функционирующие структуры, другие как продукты клеточного метаболизма, откладывающиеся внутри клетки • Некоторые из этих продуктов являются запасными питательными веществами, отложение которых клеткой происходит в условиях избытка питательных веществ в окружающей среде, а потребление наблюдается, когда организм попадает в условия голодания

Хлоросомы • К числу цитоплазматических включений, выполняющих определенную функцию в фотосинтезе, относятся хлоросомы, которые обнаруживаются у зеленых бактерий и фикобилисомы у цианобактерий • В этих структурах локализованы пигменты, поглощающие кванты света и передающие их в реакционные центры, т. е. выполняющие роль антенн • Хлоросомы имеют форму продолговатых пузырьков, окруженных однослойной мембраной построенной только из белка
 • Каждая аэросома состоит")
Аэросомы • В клетках некоторых бактерий содержатся аэросомы (газовые вакуоли) • Каждая аэросома состоит из многочисленных продолговатых газовых пузырьков • При сжатии пузырьков клетки опускаются, при увеличении объема и числа всплывают • Газовые вакуоли позволяют бактериям находится во взвешенном состоянии в водной среде

Карбоксисомы • В клетках некоторых фотоавтотрофных и хемолитоавтотрофных бактерий обнаружены карбоксисомы - включения в форме многогранников (диаметром около 500 нм) • Карбоксисомы состоят в основном из фермента рибулозодифосфаткарбоксилазы, катализирующей включение СО 2 в процессе его автотрофной фиксации

Магнитосомы • Магнитосомы присутствуют в клетках бактерий, обладающих магнитотаксисом • У разных видов бактерий форма, число и характер распределения магнитосом в клетке могут различаться • Благодаря магнитосомам бактерии способны ориентироваться в магнитном поле и перемещаться в направлении линий магнитного поля

Частицы магнетита в клетках Aquaspirillum

Гранулы запасных веществ • В клетках бактерий могут откладываться гранулы запасных веществ, наиболее распространенным из которых является поли-бета-оксимасляная кислота (ПОМ) • Гранулы ПОМ окружены белковой мембраной • Накопление этих гранул в клетках часто усиливается при выращивании бактерий на среде с ацетатом, так как синтез ПОМ начинается с ацетил-Ко. А • ПОМ образуют многие аэробные бактерии, а также цианобактерии и анаэробные фототрофные бактерии • Липидные (жиропоподобные) вещества накапливают в форме липидных капель в клетках микобактерии, нокардии и актиномицеты (до 40%)

Гранулы запасных веществ • Полисахаридные гранулы в клетках некоторых прокариот - гликоген и гранулеза - состоят из остатков Д-глюкозы • В клетках многих бактерий часто обнаруживаются гранулы волютина (метахроматина), в состав которых входят преимущественно неорганические полифосфаты, являющиеся источником фосфатных групп и энергии • В цитоплазме пурпурных и некоторых бесцветных серных бактерий обнаружены включения молекулярной серы

Полифосфатные гранулы в клетках цианобактерий Anacystis

Включения серы в клетках Chromatium

Бактериальные эндоспоры и их функции • Некоторые грамположительные бактерии способны образовывать эндоспоры • Споры - это специальные покоящиеся резистентные структуры • Споры образуются при неблагоприятных условиях, например, при исчерпании питательных веществ или накоплении вредных продуктов метаболизма в окружающей среде • Образование спор отмечается в частности у грамположительных палочковидных бактерий родов Bacillus и Clostridium • Образование эндоспор может происходить только у представителей прокариот

Бактериальные эндоспоры и их функции • Эндоспоры можно наблюдать как с помощью светового, так и с помощью электронного микроскопа • В связи с тем, что эндоспоры непроницаемы для большинства красителей, их можно наблюдать как бесцветные области в окрашенных клетках бактерий • Споры сильно преломляют свет • В каждой клетке, как правило, образуется только одна эндоспора • Спорообразование у бактерий является не способом размножения, ибо число особей при этом не увеличивается, а способом перенесения неблагоприятных условий • В окружающей среде эндоспоры помогают выживать бактериям при недостатке влаги и питательных веществ

Бактериальные эндоспоры и их функции • Эндоспоры развиваются внутри цитоплазмы вегетативных клеток, вследствие чего их и называют эндоспорами в отличие от экзоспор • Бактериальные эндоспоры – это особый тип покоящихся клеток, которые обладают специфическими структурами и характеризуются очень высокой устойчивостью к неблагоприятным факторам внешней среды, таким как нагревание, ультрафиолетовая радиация, химические дезинфектанты и высушивание • Поэтому в микробиологической практике используется стерилизация под давлением в автоклавах именно для того, чтобы инактивировать бактериальные эндоспоры

Bacillus anthracis

Споры и их расположение в клетке • Спорообразование, форма и расположение спор в клетке являются видовым признаком, что может быть использовано при идентификации и определении видовой принадлежности спорообразующих бактерий • Форма спор может быть овальной, и шаровидной • Спора может располагаться в клетке терминально – на конце клетки, субтерминально – т. е. ближе к концу клетки (но не на самом конце) или занимать центральное положение

Расположение спор в клетке

Спорообразование • Процесс спорообразования состоит из нескольких этапов или стадий • Формированию эндоспоры предшествует репликация ДНК • Спорообразование начинается с формирования у одного из полюсов клетки спорогенной зоны, этот процесс имеет место внутри бактериальной клетки • Cпорогенная зона представляет собой уплотненный участок цитоплазмы, куда переходит одна или несколько реплицированных хромосом • Затем происходит врастание (инвагинация) цитоплазматической мембраны от периферии к центру, что ведет к образованию споровой перегородки и изоляции спорогенной зоны от остальной части цитоплазмы •

Спорообразование • Затем наступает этап формирования проспоры • Происходит окружение протопласта споры протопластом материнской клетки • Наблюдается как бы обтекание или обрастание отсеченного участка спорогенной зоны цитоплазматической мембраной вегетативной материнской клетки • В результате образуется проспора – структура, расположенная внутри материнской клетки и полностью отделенная от нее двумя элементарными мембранами: наружной и внутренней по отношению к проспоре

Спорообразование у бактерий

Спорообразование • После образования проспоры происходит образование кортекса и оболочек споры • Кортекс или кортикальный слой – это особый слой пептидогликана, который формируется между наружным и внутренним мембранными слоями проспоры • В дальнейшем внешняя сторона мембраны покрывается плотной оболочкой, в состав которой входят белки, липиды и другие соединения, не встречающиеся у вегетативных клеток • В частности в оболочках споры накапливается дипиколиновая кислота и кальций • У некоторых бактерий поверх покровов споры формируется еще один дополнительный слой, который называется экзоспориум • Таким образом, спора оказывается покрытой многослойной оболочкой

Строение зрелой споры

Спорообразование • После того как спора сформировалась, может происходить разрушение материнской клетки и спора выходит в окружающую среду • В благоприятных условиях спора может прорастать, при этом из одной споры образуется одна бактерия • Кроме эндоспор прокариоты могут образовывать другие покоящиеся формы – экзоспоры, цисты и акинеты • Образование экзоспор, в частности, характерно для актиномицетов • Образование цист характерно для азотобактера и некоторых других бактерий • Некоторые цианобактерии образую акинеты

Сопоставление некоторых черт прокариотной и эукариотнои клеточной организации • Прокариотная клетка • Эукариотная клетка • Нуклеоид (ДНК не отделена от цитоплазмы мембраной) • Ядро (ДНК отделена от цитоплазмы ядерной оболочкой) • Больше одной хромосомы • Деление ядра путем митоза • Органеллы имеются • Рибосомы 80 S типа • Клеточная стенка не содержит пептидогликана • Каждый жгутик содержит набор микротрубочек, собранных в группы: (2 х 9) + 2 • • • Одна кольцевая хромосома Митоз отсутствует Органеллы отсутствуют Рибосомы 70 S типа Клеточная стенка содержит пептидогликан • Нить жгутика построена из белковых субъединиц, образующих спираль
Общ микробиол Лекция 3-5 ИН Строение клетки.ppt