Физиология сердца.ppt
- Количество слайдов: 74



Физиология сердца

Общий план строения сердца Основные структуры сердца n камеры; n магистральные сосуды (входные и выходные); n клапаны. Камеры сердца: n правое предсердие; n правый желудочек; n левое предсердие; n левый желудочек.

Общий план строения сердца Камеры: ЛЖ – левый желудочек; ЛП – левое предсердие; ПЖ – правый желудочек; ПП – правое предсердие. Сосуды: А – аорта; ЛС – легочный ствол; Л В – легочные вены; ПВ – полые вены. Клапаны: АК – аортальный клапан; ЛК – клапан легочного ствола; МК – митральный клапан; ТК – трехстворчатый клапан.

Общий план строения сердца Магистральные сосуды: n в правое предсердие впадают верхняя и нижняя полые вены; n из правого желудочка выходит легочный ствол; n в левое предсердие впадают четыре легочные вены; n из левого желудочка выходит аорта

Клапаны сердца Между предсердиями и желудочками – атриовентрикулярные клапаны: n между правым предсердием и правым желудочком – трехстворчатый, или трикуспидальный клапан; n между левым предсердием и левым желудочком – двустворчатый, или митральный клапан; Между желудочками и магистральными артериями – полулунные клапаны: n между правым желудочком и легочным стволом – клапан легочного ствола; n между левым желудочком и аортой – аортальный клапан.

Сердце связано с сосудистой системой следующим образом: n левый желудочек через аорту выбрасывает кровь в большой круг; n из большого круга через полые вены кровь возвращается в правое предсердие; n из правого предсердия кровь поступает в правый желудочек и далее выбрасывается через легочный ствол в малый круг; n из малого круга через четыре легочные вены (по две от каждого легкого) кровь оттекает в левое предсердие и оттуда — в левый желудочек

Общие представления о работе сердца. Насосная функция сердца Цель работы сердца — перекачивать кровь из вен в артерии. Способ работы сердца — принцип пульсирующего насоса: чередование выброса крови в артерии и заполнения кровью из вен.
 и расслабления (диастолы)")
Работа сердца осуществляется за счет трех главных особенностей: чередования сокращения (систолы) и расслабления (диастолы) каждой из камер. При систоле происходит выброс крови из камеры, при диастоле — ее заполнение; n последовательности сокращения предсердий и желудочков. Сначала сокращаются предсердия, вбрасывая кровь в желудочки, затем желудочки, выбрасывая кровь в артерии; n деятельности клапанов. Клапаны сердца расположены на входе и выходе желудочков, открываясь в направлении от венозного конца к артериальному (то есть пропускают кровь из предсердий в желудочки и из желудочков — в артерии). Тем самым клапаны препятствуют обратному току крови. n

Работа сердца осуществляется за счет трех главных особенностей: Таким образом, работа сердца состоит из трех основных фаз: n систола предсердий, во время которой кровь из предсердий вбрасывается в желудочки. n систола желудочков, во время которой кровь из желудочков выбрасывается в артерии (аорту и легочный ствол). n общая пауза, во время которой расслабленное сердце заполняется кровью из вен.
; n")
Физиологические свойства сердца следующие: n возбудимость (способность генерировать ПД в ответ на раздражитель); n автоматизм (способность генерировать ПД самопроизвольно, без раздражителя); n проводимость (способность проводить ПД); n сократимость (способность сокращаться).

Физиологические свойства сердца Изолированное сердце, лишенное какойлибо внешней регуляции, способно сокращаться не только ритмично, но и в обычной последовательности (систола предсердий — систола желудочков — общая пауза).

Физиологические свойства сердца Это может быть обусловлено исключительно особенностями физиологи ческих свойств самого сердца, которые обеспречивают: n ритмичные сокращения сердца; n последовательность сокращения камер сердца; n чередование сокращения и расслабления.

Физиологические свойства сердца Сердце состоит из двух типов мышечных волокон: n рабочих кардиомиоцитов, обеспечивающих сокращение сердца; n атипичных кардиомиоцитов, обеспечивающих ритмичность и последовательность сокращения камер сердца.

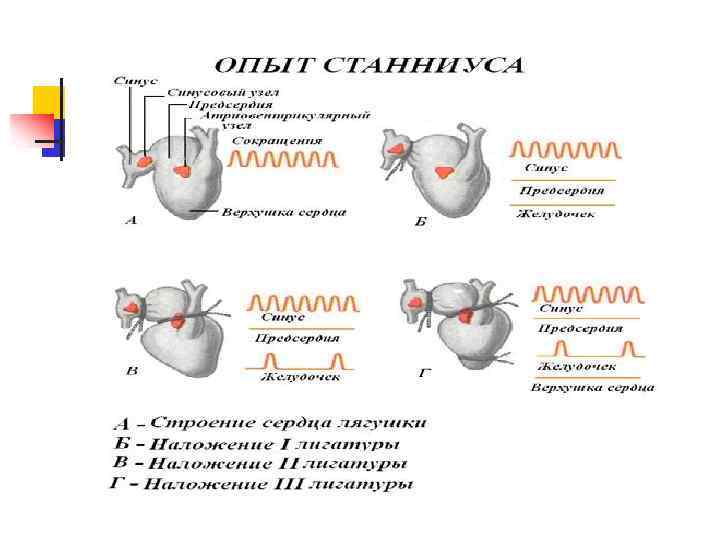
Атипичные кардиомиоциты образуют проводящую систему сердца

Проводящая система состоит из: синусового узла, расположенного в правом верхнем углу правого предсердия в области устьев полых вен; n атриовентрикулярного узла, расположенного в правом предсердии на границе между правым предсердием и правым желудочком; n
. Эту систему образуют: n пучок")
Проводящая система состоит из: внутрижелудочковой проводящей системы (системы Гиса—Пуркинье). Эту систему образуют: n пучок Гиса, отходящий от атриовентрикулярного узла; n правая и левая ножки пучка Гиса, последняя делится на левую переднюю и левую заднюю ветви; n волокна Пуркинье — конечные ветвления внутрижелудочковой проводящей системы, отходящие от ножек пучка Гиса вглубь миокарда n
. Автоматизм сердца обусловлен")
Автоматизм сердца — это его способность сокращаться самопроизвольно (без внешних раздражителей). Автоматизм сердца обусловлен n способностью определенных его самопроизвольно генерировать ПД. клеток Способностью к автоматизму обладают все атипичные кардиомиоциты (клетки проводящей системы сердца), но не обладают рабочие кардиомиоциты. В связи с этим клетки и структуры проводящей системы сердца называют водителями ритма, или пейсмекерами. n

Автоматизм обусловлен электрофизиологическими особенностями атипичных кардиомиоцитов, главная из которых заключается в том, что у этих клеток нет стабильного потенциала покоя: по окончании ПД эти клетки медленно самопроизвольно деполяризуются, в результате мембранный потенциал постепенно приближается к Екр, и по достижении этого уровня возникает следующий ПД.

Спонтанная диастолическая деполяризация Мембранный потенциал пейсмекерной клетки. МДП — максимальный диастолический потенциал; СДД — спонтанная диастолическая деполяризация.

Автоматизм В проводящей системе сердца существует градиент автоматизма: способность к автоматизму у разных структур проводящей системы сердца выражена по разному: частота самопроизвольных разрядов убывает в направлении от основания к верхушке сердца, составляя: n у синусового узла – 80 100 в минуту; n у атриовентрикулярного узла – 40 60 в минуту; n у волокон Пуркинье – 15 40 в минуту.

Автоматизм Физиологический смысл градиента автоматизма заключается в том, что с одной стороны, все клетки проводящей системы сердца должны обладать автоматизмом. Если бы способностью к автоматизму обладал только синусовый узел, то его выход из строя сопровождался остановкой сердца и смертью

Автоматизм С другой стороны, проявлять автоматизм должен только синусовый узел (возбуждение, а следовательно, сокращение сердца должно начинаться от предсердий), в противном случае вместо строго последовательного сокращения сердца наблюдались бы нерегулярные некоординированные сокращения, начинающиеся то от одного, то от другого отдела.

Автоматизм других клеток должен быть в норме подавлен, а проявляться он должен только при повреждении синусового узла.
 приходит к атриовентрикулярному")
Подавление автоматизма латентного пейсмекера Волна возбуждения от синусового узла (пунктирные стрелки) приходит к атриовентрикулярному узлу и вызывает его разряд раньше, чем его мембранный потенциал достиг бы Екр в результате его собственной спонтанной диастолической деполяризации (пунктирная кривая).

Автоматизм Градиент автоматизма проявляется в том, что: n обладая самой высокой частотой разрядов, синусовый узел первым генерирует ПД; n этот ПД проводится к другим структурам проводящей системы сердца (например, к атриовентрикулярному узлу) и вызывает их разряд прежде, чем их мембранный потенциал самопроизвольно достигнет Екр. Таким образом, автоматизм атриовентрикулярного узла и клеток внутрижелудочковой проводящей системы в норме не успевает проявиться — они возбуждаются не самопроизвольно, а под действием импульсов из синусового узла.

Автоматизм В норме автоматизм проявляет только синусовый узел, и поэтому он называется истинным или ведущим водителем ритма (пейсмекером). Остальные структуры проводящей системы сердца проявляют автоматизм только при нарушении поступления импульсов от синусового узла и поэтому называются скрытыми, или латентными, водителями ритма (пейсмекерами).

Автоматизм В случае выхода из строя синусового узла роль водителя ритма принимает атриовентрикулярный узел, а при его повреждении – волокна Пуркинье. Частота сердечных сокращений при этом последовательно уменьшается.

Автоматизм Еще одна особенность латентных водителей ритма заключается в наличии преавтоматической паузы. Это явление заключается в том, что при внезапной остановке ведущего водителя ритма латентные водители ритма включаются не сразу, а после более или менее длительной паузы. Механизмы преавтоматической паузы не до конца изучены. Клиническое значение: из за этого явления внезапный отказ синусового узла чреват временной остановкой кровообращения с трагическими или более менее благополучными последствиями.

Проводимость Возбуждение должно охватывать сердце в строго определенной последовательности: начаться в предсердиях для нормальной последовательности сокращений: предсердия — желудочки; n начаться от области устьев полых вен для того, чтобы устья этих крупных сосудов были пережаты и не было обратного тока крови при сокращении предсердий; n охватив предсердия, перейти на желудочки с некоторой задержкой, чтобы предсердия успели сократиться перед систолой желудочков; n распространиться по желудочкам с большой скоростью, чтобы их сокращение было как можно более синхронным и, следовательно, мощным. n

Проводимость Эти требования реализуются благодаря особенностям проводимости сердца как на тканевом, так и на органном уровне.
")
Проводимость На тканевом уровне проводимость сердца отличается наличием между кардиомиоцитами (и рабочими, и атипичными) щелевых контактов, или нексусов. Через эти контакты возбуждение свободно переходит с одного кардиомиоцита на другой. Следовательно, импульс, возникший в одной клетке сердца, способен охватить все сердце.

Сердечная мышца - синцитий Темные зоны – вставочные диски, в этой зоне мембраны клеток сливаются друг с другом и формируют щелевые контакты, через которые свободно диффундируют ионы, что обеспечивает беспрепятственное распространение ПД. Т. е. миокард можно рассматривать как синцитий: возбуждение одной клетки ведет к распространению ПД на все остальные.

Сердечная мышца - синцитий Сердце состоит функциональных из двух синцитиев: предсердного синцития (мышечная стенка предсердий), желудочкового синцития (мышечная стенка желудочков). Предсердия отделены от желудочков фиброзной перегородкой, в которой имеются атриовентрикулярные отверстия, снабженные клапанами. Возбуждение не может пройти от предсердного синцития к желудочковому непосредственно через фиброзную ткань.

Сердечная мышца - синцитий Возбуждение передается только с помощью атриовентрикулярного пучка диаметром в несколько миллиметров, состоящего из волокон проводящей системы сердца. Наличие в сердце двух функциональных синцитиальных систем позволяет предсердиям сокращаться раньше, чем начнется сокращение желудочков.

Проводимость На органном уровне проводимость сердца отличается наличием проводящей системы, отвечающей за последовательность распро странения возбуждения по сердцу. Эта последовательность обусловлена: n локализацией структур (узлов и пучков) проводящей системы n распределением скоростей проведения в этих структурах.

Проводимость Скорости проведения в структурах проводящей системы, а также в рабочем миокарде следующие: n во внутрижелудочковой проводящей системе – до 5 м/с; n в рабочем миокарде – до 1 м/с; n в атриовентрикулярном узле – несколько сантиметров в секунду.

Возбуждение распространяется по сердцу следующим образом: Выходит из синусового узла, вызывая прежде всего сокращение миокарда в области устьев полых вен и их пережатие. n Распространяется со средней скоростью (до 1 м/с) по рабочему миокарду предсердий. n
")
Возбуждение распространяется по сердцу следующим образом: Медленно (со скоростью порядка нескольких сантиметров в секунду) проводится по атриовентрикулярному узлу, обеспечивая атриовентрикулярную задержку (поскольку стенки предсердий отделены от стенок желудочков фиброзными кольцами, возбуждение может перейти с предсердий на желудочки только через атриовентрикулярный узел и пучок Гиса). n
 распространяется по")
Возбуждение распространяется по сердцу следующим образом: С высокой скоростью (до 5 м/с) распространяется по внутрижелудочковой проводящей системе, обеспечивая синхронный выход возбуждения сразу на многие участки рабочего миокарда желудочков. n На небольшое расстояние (от одного волокна Пуркинье до другого, от окончания волокна Пуркинье до эпикардиальной поверхности желудочка) со средней скоростью (до 1 м/с) проводится по рабочему миокарду желудочков. n

Возбудимость Кроме определенной последовательности сокращения к работе сердца предъявляется еще одно важное требование: сокращение обязательно должно чередоваться с расслаблением. Это требование осуществляется благодаря особенностям возбуждения (потенциала действия) кардиомиоцитов.

Возбудимость ПД рабочего кардиомиоцита отличается от ПД волокна скелетной мышцы наличием фазы плато: после пика ПД сначала начинается реполяризация, но затем она прекращается, и мембранный потенциал на 100— 300 мс почти застывает на постоянном уровне около 0 м. В (точнее, реполяризация продолжается, но резко замедляется). Это и есть фаза плато. После нее следует быстрая ПД рабочего кардиомиоцита реполяризация до уровня потенциала покоя.

ПД рабочего кардиомиоцита Благодаря фазе плато сердце обладает длительным рефрак -терным периодом: он примерно соответствует по времени этой фазе. Вследствие этого в сердце невозможен тетанус. Уточнение: ПД рабочего кардиомиоцита подразделяется на фазы деполяризации, ранней быстрой реполяризации, медленной реполяризации (плато) и поздней быстрой ре поляризации.

Вспоминаем Тетанус в скелетной мышце возникает, если очередное раздражение наносится и вызывает ПД (следовательно и сокращение) в тот момент, когда мышца еще не расслабилась, но рефрактерный период предыдущего ПД уже закончился. В сердечной мышце рефрактерный период по времени соизмерим с длительностью сокращения; следовательно, очередной раздражитель может вызвать возбуждение (и сокращение) только тогда, когда сердечная мышца успела расслабиться. Именно поэтому сокращение сердца всегда чередуется с расслаблением.
")
Рефрактерный период скелетной и сердечной мышцы В скелетной мышце рефрактерный период (изображен серыми прямоугольниками) значительно короче, чем сокращение, поэтому частые стимулы (стрелки) могут вызвать очередной ПД и очередное сокращение, когда предыдущее сокращение еще не закончилось; возникает тетанус. В сердечной мышце ПД и рефрактерный период гораздо длительнее и соответствуют времени сокращения, поэтому стимулы, приходящиеся на период рефрактерности, не могут вызвать очередной ПД и очередное сокращение; тетанус отсутствует.

Сократимость Особенности сократимости сердца вытекают из особенностей его возбудимости и проводимости. Сокращение сердца подчиняется закону «все или ничего» — то есть сила сокращения не зависит от силы раздражения: на подпороговые раздражители сердце не реагирует ( «ничего» ), а на пороговые и сверхпороговые реагирует сокращениями одинаковой силы ( «все» ).

Сократимость Сокращения сердца одинаковой сила обусловлены особенностями проводимости сердечной мышцы: наличием щелевых контактов. В скелетной мышце сила сокращений зависит от силы раздражителя, так как мышечные волокна возбуждаются отдельно друг от друга, и чем сильнее раздражитель, тем больше вовлекается в возбуждение (и сокращение) мышечных волокон (например, сильный ток распространяется на большее число волокон, чем слабый). В сердце возбуждение, распространяясь от клетки к клетке через щелевые контакты, всегда охватывает все сердце, и в сокращение независимо от силы раздражителя вовлекаются все волокна.

Сократимость сердца регулируется иначе, чем в скелетной мышце. В скелетной мышце сила сокращения зависит от количества одновременно сокращающихся волокон: чем больше волокон участвует в сокращении, тем оно сильнее, при этом сила сокращения каждого отдельного волокна всегда одинакова. В сердце количество сокращающихся волокон всегда одинаково (в каждом сокращении участвуют все волокна), но зато сила сокращения отдельных волокон может регулироваться.

Ионно-молекулярные механизмы возбудимости и сократимости Главное отличие мембранных потенциалов рабочих кардиомиоцитов от мембранных потенциалов клеток скелетных мышц — наличие фазы плато

Ионно-молекулярные механизмы возбудимости и сократимости Фаза плато обусловлена наличием у рабочих кардиомиоцитов медленных кальциевых каналов. Через эти каналы во время фазы плато в кардиомиоцит входит кальций. В остальном ионные токи рабочих кардиомиоцитов и скелетных мышц в основном сходны: в рабочих кардиомиоцитах, как и в скелетных мышцах, имеются безворотные калиевые каналы (обусловливающие постоянно высокую проницаемость для калия в покое), быстрые натриевые каналы и потенциал чувствительные калиевые каналы
 и инактивационными")
Медленный кальциевый канал Этот канал, как и быстрый натриевый, обладает активационными (наружными) и инактивационными (внутренними) воротами. В состоянии покоя инактивационные ворота открыты, но активационные — закрыты; в результате канал закрыт. В ответ на деполяризацию сначала открываются активационные ворота, канал переходит в открытое состояние (активация); затем закрываются инактивационные ворота, канал переходит в закрытое состояние (инактивация). А — активационные ворота; И — инактивационные ворота; МП — мембранный потенциал; g. Ca — проводимость медленных кальциевых каналов.

Медленные кальциевые каналы во многом похожи на быстрые натриевые каналы: n они также обладают двумя воротами — наружными активационными и внутренними инактивационными; n активационные ворота при потенциале покоя закрыты, а при деполяризации открываются; n инактивационные ворота при потенциале покоя открыты, а при деполяризации закрываются; n активационные ворота срабатывают быстрее инактивационных, и поэтому в ответ на деполяризацию канал на какое то время открывается, а затем закрывается.

Отличия медленных кальциевых каналов от быстрых натриевых видны из самого названия этих каналов: n медленные кальциевые каналы преимущественно пропускают Са 2+; n ворота медленных кальциевых каналов (и активационные, и инактивационные) срабатывают значительно медленнее, чем соответствующие ворота быстрых натриевых каналов.

Поскольку внеклеточная концентрация Са 2+ всегда выше внутриклеточной, концентрационный градиент для Ca 2+ направлен внутрь. Заряд внутри клетки в покое отрицателен, электрический градиент для Са 2+ также направлен внутрь. Следовательно, при открывании медленных кальциевых каналов Са 2+ входит в кардиомиоцит

Основные ионные токи в рабочем кардиомиоците В ответ на раздражитель открываются быстрые натриевые каналы, входит Na+. Это фаза деполяризации. n В ответ на вызванную входом Na+ деполяризацию с некоторой задержкой открываются медленные кальциевые каналы, начинается вход Са 2+. Одновременно повышается выход из клетки К+, так как при деполяризации электрический градиент для К+ становится направленным наружу (а концентрационный градиент для К+ направлен наружу всегда). В течение некоторого времени вход Са 2+ уравновешивается выходом К+, и в результате мембранный потенциал удерживается на постоянном уровне. Это фаза плато. n
 постепенно закрываются")
Основные ионные токи в рабочем кардиомиоците На фоне продолжающейся деполяризации (фазы плато) постепенно закрываются медленные кальциевые каналы (инактивационные ворота), и вход Са 2+ прекращается. Остается только выход К+, который дополнительно усиливается из-за активации потенциалчувствительных калиевых каналов. Это фаза реполяризации (более точное название — фаза поздней быстрой реполяризации).

Основные ионные токи в рабочем кардиомиоците Основные фазы ПД рабочих кардиомиоцитов обусловлены следующими токами: n деполяризация — вход Na+; n плато — вход Са 2+ на фоне выхода К+; n реполяризация — выход К+

Основные ионные токи в рабочем кардиомиоците Входящие токи изображены стрелками, направленными внутрь ПД, выходящие – наружу. IСа – кальциевый ток; IК – калиевый ток; INa – натриевый ток.
 и рабочего (жирная кривая) кардиомиоцитов Mембранные потенциалы клеток синусового")
Мембранные потенциалы пейсмекерного (тонкая кривая) и рабочего (жирная кривая) кардиомиоцитов Mембранные потенциалы клеток синусового узла отличаются: n меньшей поляризованностью (у рабочих кардиомиоцитов потенциал покоя около -90 м. В, у клеток синусового узла мембранный потенциал почти никогда не бывает отрицательнее -75 м. В); n меньшей амплитудой и крутизной ПД; наличием спонтанной диастолической деполяризации — это главная особенность пеqсмекерных клеток. n
 и рабочего (жирная кривая) кардиомиоцитов Все эти особенности обусловлены")
Мембранные потенциалы пейсмекерного (тонкая кривая) и рабочего (жирная кривая) кардиомиоцитов Все эти особенности обусловлены одним главным свойством: у пейсмекерных клеток имеется постоянная высокая проницаемость для Na+ (обусловленная не быстрыми натриевыми каналами, а постоянно открытыми каналами утечки).
 и рабочего (жирная кривая) кардиомиоцитов Из этого следует: из")
Мембранные потенциалы пейсмекерного (тонкая кривая) и рабочего (жирная кривая) кардиомиоцитов Из этого следует: из за постоянного высокого входа Na+ клетки синусового узла относительно деполяри зованы;
 и рабочего (жирная кривая) кардиомиоцитов на фоне постоянной деполяризации")
Мембранные потенциалы пейсмекерного (тонкая кривая) и рабочего (жирная кривая) кардиомиоцитов на фоне постоянной деполяризации быстрые натриевые каналы в этих клетках инактивированы; следовательно, ПД создается медленным кальциевым током. Отсюда — меньшая амплитуда и крутизна ПД
 и рабочего (жирная кривая) кардиомиоцитов после окончания ПД продолжается")
Мембранные потенциалы пейсмекерного (тонкая кривая) и рабочего (жирная кривая) кардиомиоцитов после окончания ПД продолжается вход Na+ в клетку. Это приводит к по степеннойдеполяризации, то есть спонтанной диастолической деполяри зации;
 и рабочего (жирная кривая) кардиомиоцитов спонтанная деполяризация приводит к")
Мембранные потенциалы пейсмекерного (тонкая кривая) и рабочего (жирная кривая) кардиомиоцитов спонтанная деполяризация приводит к открыванию медленных кальциевых каналов; вход по ним Са 2+, в свою очередь, еще больше усиливает деполяризацию, которая по достижении Екр перерастает в ПД. Следовательно, кальциевый ток в клетках синусового узла не только создает ПД, но и участвует в спонтанной диастолической деполяризации. Таким образом, спонтанная диастолическая деполяризация обусловлена постоянным входом Na+, а в последней трети — еще и входом Са 2+.

Таким образом, основные фазы ПД синусового узла обусловлены следующими токами: спонтанная диастолическая деполяризация – вход Na+, а затем еще и вход Са 2+ деполяризация – вход Са 2+ ;

реполяризация — выход К Натрий, входящий в клетку по постоянно открытым каналам, удаляется с помощью Na+К+ АТФазы

Основные ионные токи в клетке синусового узла Входящие токи изображены стрелками, направленными внутрь ПД, выходящие — наружу.

Сократимость Основная последовательность сокращения скелетной и сердечной мышц одинакова. 1. На клеточной мембране возникает ПД. 2. ПД вызывает вход кальция в цитоплазму. 3. Кальций запускает взаимодействие актина с миозином, то есть сокращение. 4. Затем кальций удаляется из цитоплазмы, взаимодействие актина с миозином прекращается, возникает расслабление.

Сократимость Главное отличие касается источников кальция в скелетной и сердечной мышцах: в скелетной мышце кальций поступает в цитоплазму из саркоплазматического ретикулума, а удаляется путем обратного закачивания снова в ретикулум; n в сердечной мышце кальций не только поступает в цитоплазму из саркоплазматического ретикулума, но также входит через поверхностную мембрану (сарколемму); соответственно, он удаляется не только обратно в ретикулум, но и через сарколемму наружу. n

Сократимость Вход Са 2+ через сарколемму осуществляется по медленным кальциевым каналам во время ПД.

Сократимость Выход Са 2+ через сарколемму осуществляется благодаря двум механизмам: 1 - Са 2+-АТФазе (кальциевому насосу) сарколеммы. Перенос Са 2+ идет против электрохимического градиента, а источником энергии служит АТФ (первичный активный транспорт);

Сократимость 2 натрий кальциевому обменнику сарколеммы, выводящему из клетки Са 2+ в обмен на вход в клетку Na+. Перенос Са 2+ идет против электрохимического градиента, а источником энергии служит электрохимический градиент для Na+ (вторичный активный транспорт).

Сократимость Таким образом, в рабочих кардиомиоцитах имеются следующие основные виды транспорта кальция: n Вход в цитоплазму из наружной среды через медленные кальциевые каналы сарколеммы. n Активное выведение через сарколемму Са 2+-АТФазой сарколеммы. n Активное выведение через сарколемму натрийкальциевым обменником. n Выход в цитоплазму через кальциевые каналы саркоплазматического ретикулума. n Активное закачивание в саркоплазматический ретикулум Са 2+-АТФазой саркоплазматического ретикулума.

Транспорт кальция в рабочем кардиомиоците 1 – кальциевый канал сарколеммы; 2 – Са 2+-АТФаза сарколеммы; 3 – натрий-кальциевый обменник; 4 – кальциевый канал саркоплазматического ретикулума; 5 – Са 2+-АТФаза саркоплазматического ретикулума.
Физиология сердца.ppt