

Частная физиология.ppt
- Количество слайдов: 120
 Физиология различных отделов ЦНС
Физиология различных отделов ЦНС
 Физиология спинного мозга • • • Спинной мозг – наиболее древнее образование ЦНС. Характерная особенность строения – сегментарность. Нейроны спинного мозга образуют его серое вещество в виде передних и задних (столбов) рогов. Задние рога содержат интернейроны, которые реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения и передают импульсы в вышележащие центры, в симметричные структуры противоположной стороны, к передним рогам спинного мозга. Передние рога содержат нейроны (мотонейроны), дающие аксоны к мышцам, они являются эфферентными. Все нисходящие пути ЦНС, осуществляющие двигательные реакции, заканчиваются в передних рогах. В боковых рогах шейных и двух поясничных сегментов располагаются нейроны симпатического отдела вегетативной нервной системы, во втором— четвертом крестцовых сегментах – парасимпатического. Белое вещество спинного мозга образовано нервными волокнами и выполняет проводниковую роль.
Физиология спинного мозга • • • Спинной мозг – наиболее древнее образование ЦНС. Характерная особенность строения – сегментарность. Нейроны спинного мозга образуют его серое вещество в виде передних и задних (столбов) рогов. Задние рога содержат интернейроны, которые реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения и передают импульсы в вышележащие центры, в симметричные структуры противоположной стороны, к передним рогам спинного мозга. Передние рога содержат нейроны (мотонейроны), дающие аксоны к мышцам, они являются эфферентными. Все нисходящие пути ЦНС, осуществляющие двигательные реакции, заканчиваются в передних рогах. В боковых рогах шейных и двух поясничных сегментов располагаются нейроны симпатического отдела вегетативной нервной системы, во втором— четвертом крестцовых сегментах – парасимпатического. Белое вещество спинного мозга образовано нервными волокнами и выполняет проводниковую роль.
 волокна, а задние") Закон Белла - Мажанди • Передний корешки содержат эфферентные двигательные (выходящие) волокна, а задние корешки содержат афферентные чувствительные (входящие) волокна
Закон Белла - Мажанди • Передний корешки содержат эфферентные двигательные (выходящие) волокна, а задние корешки содержат афферентные чувствительные (входящие) волокна
 • • • Спинной мозг человека содержит около 13 млн. нейронов, из них только 3% — мотонейроны, а 97% — вставочные. Функционально нейроны спинного мозга можно разделить на 4 основные группы: 1) мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки; 2) интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти афферентные нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения и передают импульсы в вышележащие центры, в симметричные структуры противоположной стороны, к передним рогам спинного мозга; 3) симпатические, парасимпатические нейроны расположены в боковых рогах. В боковых рогах шейных и двух поясничных сегментов располагаются нейроны симпатического отдела вегетативной нервной системы, во II—IV сегментах крестцовых – парасимпатического. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков и направляются к клеткам ганглиев симпатической цепочки и к ганглиям внутренних органов; 4) ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами. Так, в основании заднего рога находится крупное скопление нервных клеток, образующих промежуточное ядро спинного мозга. Его нейроны имеют короткие аксоны, которые в основном идут в передний рог и образуют там синаптические контакты с мотонейронами. Аксоны некоторых из этих нейронов распространяются на 2 -3 сегмента, но никогда не выходят за пределы спинного мозга.
• • • Спинной мозг человека содержит около 13 млн. нейронов, из них только 3% — мотонейроны, а 97% — вставочные. Функционально нейроны спинного мозга можно разделить на 4 основные группы: 1) мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки; 2) интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти афферентные нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения и передают импульсы в вышележащие центры, в симметричные структуры противоположной стороны, к передним рогам спинного мозга; 3) симпатические, парасимпатические нейроны расположены в боковых рогах. В боковых рогах шейных и двух поясничных сегментов располагаются нейроны симпатического отдела вегетативной нервной системы, во II—IV сегментах крестцовых – парасимпатического. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков и направляются к клеткам ганглиев симпатической цепочки и к ганглиям внутренних органов; 4) ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами. Так, в основании заднего рога находится крупное скопление нервных клеток, образующих промежуточное ядро спинного мозга. Его нейроны имеют короткие аксоны, которые в основном идут в передний рог и образуют там синаптические контакты с мотонейронами. Аксоны некоторых из этих нейронов распространяются на 2 -3 сегмента, но никогда не выходят за пределы спинного мозга.
 Мотонейроны • Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на мотонейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем» . • Выделяют три типа мотонейронов: альфа, бета и гамма. • Альфа-мотонейроны представлены крупными мультиполярными клетками с диаметром тела 25 -75 мкм; их аксоны иннервируют двигательные мышцы, которые способны развивать значительную силу. • Бета-мотонейроны - это мелкие нейроны, которые иннервируют тонические мышцы. • Гамма-мотонейроны еще мельче - диаметр их тела 15 -25 мкм. Они локализованы в двигательных ядрах вентральных рогов среди альфа- и бета-мотонейронов. Гамма-мотонейроны осуществляют двигательную иннервацию мышечных рецепторов (мышечных веретен). Аксоны мотонейронов составляют основную массу передних корешков спинного мозга.
Мотонейроны • Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на мотонейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем» . • Выделяют три типа мотонейронов: альфа, бета и гамма. • Альфа-мотонейроны представлены крупными мультиполярными клетками с диаметром тела 25 -75 мкм; их аксоны иннервируют двигательные мышцы, которые способны развивать значительную силу. • Бета-мотонейроны - это мелкие нейроны, которые иннервируют тонические мышцы. • Гамма-мотонейроны еще мельче - диаметр их тела 15 -25 мкм. Они локализованы в двигательных ядрах вентральных рогов среди альфа- и бета-мотонейронов. Гамма-мотонейроны осуществляют двигательную иннервацию мышечных рецепторов (мышечных веретен). Аксоны мотонейронов составляют основную массу передних корешков спинного мозга.
 Спинной мозг выполняет рефлекторную и проводниковую функции. • • Рефлекторная функция позволяет реализовать все двигательные рефлексы тела, рефлексы внутренних органов, терморегуляции и т. д. Рефлекс – реакция организма на раздражение рецепторов, которая осуществляется с участием ЦНС. Структурной основой рефлекса является рефлекторная дуга. Рефлекторная дуга – последовательно соединенная цепочка нервных клеток, которая обеспечивает осуществление реакции, ответа на раздражение. Рефлекторные дуги могут быть двух видов: 1) простые – моносинаптические рефлекторные дуги, состоящие из 2 нейронов (рецепторного (афферентного) и эффекторного), между ними имеется 1 синапс; 2) сложные – полисинаптические рефлекторные дуги. В их состав входят 3 или больше нейронов – чувствителный, один или несколько вставочных и эффекторный (двигательный). Проводниковая функция обеспечивает связь нейронов спинного мозга друг с другом или с вышележащими отделами ЦНС.
Спинной мозг выполняет рефлекторную и проводниковую функции. • • Рефлекторная функция позволяет реализовать все двигательные рефлексы тела, рефлексы внутренних органов, терморегуляции и т. д. Рефлекс – реакция организма на раздражение рецепторов, которая осуществляется с участием ЦНС. Структурной основой рефлекса является рефлекторная дуга. Рефлекторная дуга – последовательно соединенная цепочка нервных клеток, которая обеспечивает осуществление реакции, ответа на раздражение. Рефлекторные дуги могут быть двух видов: 1) простые – моносинаптические рефлекторные дуги, состоящие из 2 нейронов (рецепторного (афферентного) и эффекторного), между ними имеется 1 синапс; 2) сложные – полисинаптические рефлекторные дуги. В их состав входят 3 или больше нейронов – чувствителный, один или несколько вставочных и эффекторный (двигательный). Проводниковая функция обеспечивает связь нейронов спинного мозга друг с другом или с вышележащими отделами ЦНС.
 В рефлекторной дуге различают пять звеньев: • рецептор; • чувствительное волокно, проводящее возбуждение к центрам; • нервный центр, где происходит переключение возбуждения с чувствительных клеток на двигательные; • двигательное волокно, несущее нервные импульсы на периферию; • действующий орган - мышца или железа.
В рефлекторной дуге различают пять звеньев: • рецептор; • чувствительное волокно, проводящее возбуждение к центрам; • нервный центр, где происходит переключение возбуждения с чувствительных клеток на двигательные; • двигательное волокно, несущее нервные импульсы на периферию; • действующий орган - мышца или железа.
 экстероцептивные - возникают при раздражении факторами внешней") Рефлексы делятся на: • • • 1) экстероцептивные - возникают при раздражении факторами внешней среды сенсорных раздражителей; 2) интероцептивные - возникают при раздражении механо-, хемо-, терморецепторов: висцеро-висцеральные – рефлексы с одного внутреннего органа на другой, висцеро-мышечные – рефлексы с внутренних органов на скелетную мускулатуру; 3) проприоцептивные (собственные) рефлексы с самой мышцы и связанных с ней образований. Они имеют моносинаптическую рефлекторную дугу. Проприоцептивные рефлексы регулируют двигательную активность за счет сухожильных и позотонических рефлексов. Сухожильные рефлексы (коленный, ахиллов, с трехглавой мышцы плеча и т. д. ) возникают при растяжении мышц и вызывают расслабление или сокращение мышцы, возникают при каждом мышечном движении. Исследование проприоцептивных рефлексов производится для определения возбудимости и степени поражения ЦНС; Позотонические рефлексы (возникают при возбуждении вестибулярных рецепторов при изменении скорости движения и положения головы по отношению к туловищу, что приводит к перераспределению тонуса мышц (повышению тонуса разгибателей и уменьшению сгибателей) и обеспечивает равновесие тела).
Рефлексы делятся на: • • • 1) экстероцептивные - возникают при раздражении факторами внешней среды сенсорных раздражителей; 2) интероцептивные - возникают при раздражении механо-, хемо-, терморецепторов: висцеро-висцеральные – рефлексы с одного внутреннего органа на другой, висцеро-мышечные – рефлексы с внутренних органов на скелетную мускулатуру; 3) проприоцептивные (собственные) рефлексы с самой мышцы и связанных с ней образований. Они имеют моносинаптическую рефлекторную дугу. Проприоцептивные рефлексы регулируют двигательную активность за счет сухожильных и позотонических рефлексов. Сухожильные рефлексы (коленный, ахиллов, с трехглавой мышцы плеча и т. д. ) возникают при растяжении мышц и вызывают расслабление или сокращение мышцы, возникают при каждом мышечном движении. Исследование проприоцептивных рефлексов производится для определения возбудимости и степени поражения ЦНС; Позотонические рефлексы (возникают при возбуждении вестибулярных рецепторов при изменении скорости движения и положения головы по отношению к туловищу, что приводит к перераспределению тонуса мышц (повышению тонуса разгибателей и уменьшению сгибателей) и обеспечивает равновесие тела).
 Введем некоторые важные определения. • Минимальная сила раздражения, которая вызывает рефлекс, называется порогом (или пороговым стимулом) данного рефлекса. • Всякий рефлекс имеет рецептивное поле, т. е. совокупность рецепторов, раздражение которых вызывает рефлекс с наименьшим порогом. • Время, прошедшее от момента нанесения раздражения до ответа на него, называется временем рефлекса. • Каждый рефлекс имеет свою локализацию (место расположения) в центральной нервной системе, т. е. тот ее участок, который необходим для его осуществления. Например, центр расширения зрачка - в верхнем грудном сегменте спинного мозга. При разрушении соответствующего отдела рефлекс отсутствует.
Введем некоторые важные определения. • Минимальная сила раздражения, которая вызывает рефлекс, называется порогом (или пороговым стимулом) данного рефлекса. • Всякий рефлекс имеет рецептивное поле, т. е. совокупность рецепторов, раздражение которых вызывает рефлекс с наименьшим порогом. • Время, прошедшее от момента нанесения раздражения до ответа на него, называется временем рефлекса. • Каждый рефлекс имеет свою локализацию (место расположения) в центральной нервной системе, т. е. тот ее участок, который необходим для его осуществления. Например, центр расширения зрачка - в верхнем грудном сегменте спинного мозга. При разрушении соответствующего отдела рефлекс отсутствует.
 Рефлексы спинного мозга • Наиболее простыми рефлексами спинного мозга являются собственные рефлексы мышц –рефлексы растяжения и сухожильные рефлексы. Для осуществления данных рефлексов достаточно всего лишь одного сегмента спинного мозга. • Рефлекс растяжения проявляется в том, что мышца, у которой сохранена связь с мозгом, развивает значительно большее противодействие растяжению, чем та же мышца после перерезки нерва. Рефлекс растяжения в определенных пределах пропорционален силе растяжения. Однако чрезмерное растяжение мышцы может вызвать противоположный эффект; вместо противодействия мышца внезапно расслабляется. Это расслабление получило образное название эффекта “складного ножа”. Cухожильные рефлексы (коленный рефлекс, возникающий при ударе по сухожилию четырехглавой мышцы бедра; ахиллов рефлекс, вызываемый легким ударом по ахиллову сухожилию; и локтевой рефлекс вызываемый ударом молоточка по сухожилию четырехглавой мышцы). Сухожильные рефлексы очень хорошо взаимодействуют с другими рефлексами и подчиняются определенному контролю со стороны вышерасположенных центров. И хотя нельзя произвольно затормозить сухожильный рефлекс, однако наличие постоянного тормозящего контроля со стороны вышерасположенных центров можно легко продемонстрировать. Если отвлечь внимание испытуемого от определения у него коленного рефлекса (например, предложив ему с силой растягивать сцепленные руки), то этот рефлекс сразу же существенно усиливается. Основным отличием рефлексов растяжения от сухожильных рефлексов являются временные характеристики их течения. Если сухожильный рефлекс представляет собой кратковременную (фазную) реакцию, то рефлексы растяжения носят тонический характер. • •
Рефлексы спинного мозга • Наиболее простыми рефлексами спинного мозга являются собственные рефлексы мышц –рефлексы растяжения и сухожильные рефлексы. Для осуществления данных рефлексов достаточно всего лишь одного сегмента спинного мозга. • Рефлекс растяжения проявляется в том, что мышца, у которой сохранена связь с мозгом, развивает значительно большее противодействие растяжению, чем та же мышца после перерезки нерва. Рефлекс растяжения в определенных пределах пропорционален силе растяжения. Однако чрезмерное растяжение мышцы может вызвать противоположный эффект; вместо противодействия мышца внезапно расслабляется. Это расслабление получило образное название эффекта “складного ножа”. Cухожильные рефлексы (коленный рефлекс, возникающий при ударе по сухожилию четырехглавой мышцы бедра; ахиллов рефлекс, вызываемый легким ударом по ахиллову сухожилию; и локтевой рефлекс вызываемый ударом молоточка по сухожилию четырехглавой мышцы). Сухожильные рефлексы очень хорошо взаимодействуют с другими рефлексами и подчиняются определенному контролю со стороны вышерасположенных центров. И хотя нельзя произвольно затормозить сухожильный рефлекс, однако наличие постоянного тормозящего контроля со стороны вышерасположенных центров можно легко продемонстрировать. Если отвлечь внимание испытуемого от определения у него коленного рефлекса (например, предложив ему с силой растягивать сцепленные руки), то этот рефлекс сразу же существенно усиливается. Основным отличием рефлексов растяжения от сухожильных рефлексов являются временные характеристики их течения. Если сухожильный рефлекс представляет собой кратковременную (фазную) реакцию, то рефлексы растяжения носят тонический характер. • •
 Рефлекс растяжения
Рефлекс растяжения
 Рефлекс растяжения проявляется в том, что мышца, у которой сохранена связь с мозгом, развивает значительно большее противодействие растяжению, чем та же мышца после перерезки нерва. Очевидно, что мышца, у которой связь с мозгом нарушена, противодействует растяжению только за счет эластических свойств мышечного волокна. У мышцы же, у которой эта связь сохранена, к эластическому сопротивлению добавляется активное сопротивление. Следовательно, растяжение мышцы вызывает рефлекторное тоническое сокращение мышечных волокон, которое и противодействует растяжению. Рефлекс растяжения в определенных пределах пропорционален силе растяжения. Однако чрезмерное растяжение мышцы может вызвать противоположный эффект; вместо противодействия мышца внезапно расслабляется. Это расслабление получило образное название эффекта “складного ножа”.
Рефлекс растяжения проявляется в том, что мышца, у которой сохранена связь с мозгом, развивает значительно большее противодействие растяжению, чем та же мышца после перерезки нерва. Очевидно, что мышца, у которой связь с мозгом нарушена, противодействует растяжению только за счет эластических свойств мышечного волокна. У мышцы же, у которой эта связь сохранена, к эластическому сопротивлению добавляется активное сопротивление. Следовательно, растяжение мышцы вызывает рефлекторное тоническое сокращение мышечных волокон, которое и противодействует растяжению. Рефлекс растяжения в определенных пределах пропорционален силе растяжения. Однако чрезмерное растяжение мышцы может вызвать противоположный эффект; вместо противодействия мышца внезапно расслабляется. Это расслабление получило образное название эффекта “складного ножа”.
 Рефлексы спинного мозга • • Рефлексы конечностей: Коленный рефлекс, возникает при ударе по сухожилию четырехглавой мышцы бедра, при этом наблюдается внезапное сокращение четырехглавой мышцы бедра, в результате голень разгибается; рефлекторная дуга замыкается на уровне L 3–L 4. Ахиллов рефлекс –выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию; рефлекторная дуга замыкается на уровне крестцовых сегментов – S 1 -S 2. Подошвенный рефлекс – кожный, выражается в сгибании стопы и пальцев при штриховом раздражении подошвы; рефлекторная дуга замыкается на уровне S 1 -S 2.
Рефлексы спинного мозга • • Рефлексы конечностей: Коленный рефлекс, возникает при ударе по сухожилию четырехглавой мышцы бедра, при этом наблюдается внезапное сокращение четырехглавой мышцы бедра, в результате голень разгибается; рефлекторная дуга замыкается на уровне L 3–L 4. Ахиллов рефлекс –выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию; рефлекторная дуга замыкается на уровне крестцовых сегментов – S 1 -S 2. Подошвенный рефлекс – кожный, выражается в сгибании стопы и пальцев при штриховом раздражении подошвы; рефлекторная дуга замыкается на уровне S 1 -S 2.
 • Реализация сухожильного рефлекса была бы невозможна, если бы одновременно с сокращением мышцразгибателей не расслаблялись мышцы-сгибатели. • Поэтому во время разгибательного коленного рефлекса мотонейроны мышцсгибателей тормозятся с помощью вставочных тормозных клеток Реншоу (реципрокное торможение).
• Реализация сухожильного рефлекса была бы невозможна, если бы одновременно с сокращением мышцразгибателей не расслаблялись мышцы-сгибатели. • Поэтому во время разгибательного коленного рефлекса мотонейроны мышцсгибателей тормозятся с помощью вставочных тормозных клеток Реншоу (реципрокное торможение).
 Рефлексы спинного мозга • • Более сложно организованы рефлекторные ответы, выражаются в координированном сгибании и разгибании мышц конечностей. Примером являются сгибательные рефлексы, направленные на избежание различных повреждающих воздействий. Рецептивное поле сгибательного рефлекса достаточно сложно и включает различные рецепторные образования и различные по скорости проведения афферентные пути. Сгибательный рефлекс возникает при раздражении болевых рецепторов кожи, мышц и внутренних органов. Все разнообразные афферентные волокна, импульсация по которым приводит к развитию сгибательного рефлекса, объединяют под названием афферентов сгибательного рефлекса. Сгибательные рефлексы отличаются от собственных рефлексов мышц не только большим числом синаптических переключений на пути к мотонейронам, но и вовлечением ряда мышц, координированное сокращение которых обусловливает движение целой конечности. Одновременно с возбуждением мотонейронов, иннервирующих мышцысгибатели, происходит реципрокное торможение мотонейронов мышцразгибателей. При достаточно интенсивном раздражении рецепторов нижней конечности происходят иррадиация возбуждения и вовлечение в реакцию мышц верхней конечности и туловища. При активации мотонейронов противоположной стороны тела наблюдается не сгибание, а разгибание мышц противоположной конечности - перекрестный разгибательный рефлекс.
Рефлексы спинного мозга • • Более сложно организованы рефлекторные ответы, выражаются в координированном сгибании и разгибании мышц конечностей. Примером являются сгибательные рефлексы, направленные на избежание различных повреждающих воздействий. Рецептивное поле сгибательного рефлекса достаточно сложно и включает различные рецепторные образования и различные по скорости проведения афферентные пути. Сгибательный рефлекс возникает при раздражении болевых рецепторов кожи, мышц и внутренних органов. Все разнообразные афферентные волокна, импульсация по которым приводит к развитию сгибательного рефлекса, объединяют под названием афферентов сгибательного рефлекса. Сгибательные рефлексы отличаются от собственных рефлексов мышц не только большим числом синаптических переключений на пути к мотонейронам, но и вовлечением ряда мышц, координированное сокращение которых обусловливает движение целой конечности. Одновременно с возбуждением мотонейронов, иннервирующих мышцысгибатели, происходит реципрокное торможение мотонейронов мышцразгибателей. При достаточно интенсивном раздражении рецепторов нижней конечности происходят иррадиация возбуждения и вовлечение в реакцию мышц верхней конечности и туловища. При активации мотонейронов противоположной стороны тела наблюдается не сгибание, а разгибание мышц противоположной конечности - перекрестный разгибательный рефлекс.
 Схема сгибательного рефлекса
Схема сгибательного рефлекса
 Рефлексы позы Еще более сложный характер имеют рефлексы позы – перераспределение мышечного тонуса, возникающее при изменении положения тела или отдельных его частей. Они представляют собой большую группу рефлексов. Сгибательный тонический рефлекс позы можно наблюдать у лягушки и у млекопитающих, для которых характерно подогнутое положение конечностей (кролик). Для большинства млекопитающих и человека главное значение для поддержания положения тела играет не сгибательный, а разгибательный рефлекторный тонус.
Рефлексы позы Еще более сложный характер имеют рефлексы позы – перераспределение мышечного тонуса, возникающее при изменении положения тела или отдельных его частей. Они представляют собой большую группу рефлексов. Сгибательный тонический рефлекс позы можно наблюдать у лягушки и у млекопитающих, для которых характерно подогнутое положение конечностей (кролик). Для большинства млекопитающих и человека главное значение для поддержания положения тела играет не сгибательный, а разгибательный рефлекторный тонус.
 Шейные позные рефлексы На уровне спинного мозга особенно важную роль в рефлекторной регуляции разгибательного тонуса играют шейные позные рефлексы. Они активируются мышечными веретенами шейных мышц. Рефлекторная дуга полисинаптическая, замыкается на уровне I-III шейных сегментов. Импульсы от этих сегментов передаются мышцам туловища и конечностей, вызывая перераспределение их тонуса. Имеется две группы этих рефлексов – возникающие при наклоне и при повороте головы. Позные шейные рефлексы у кошки с удаленным вестибулярным аппаратом: a- до изменения положения головы; б- при пассивных поднимании (↑) и опускании (↓) головы. При поднятии головы передние конечности выпрямляются, а задние сгибаются. При наклоне - эффект обратный.
Шейные позные рефлексы На уровне спинного мозга особенно важную роль в рефлекторной регуляции разгибательного тонуса играют шейные позные рефлексы. Они активируются мышечными веретенами шейных мышц. Рефлекторная дуга полисинаптическая, замыкается на уровне I-III шейных сегментов. Импульсы от этих сегментов передаются мышцам туловища и конечностей, вызывая перераспределение их тонуса. Имеется две группы этих рефлексов – возникающие при наклоне и при повороте головы. Позные шейные рефлексы у кошки с удаленным вестибулярным аппаратом: a- до изменения положения головы; б- при пассивных поднимании (↑) и опускании (↓) головы. При поднятии головы передние конечности выпрямляются, а задние сгибаются. При наклоне - эффект обратный.
 и влево") Шейные позные рефлексы Изменение тонуса мышц конечностей при наклоне головы вправо (а) и влево (б): при повороте головы в сторону на одноименной стороне увеличивается тонус разгибателей и снижается тонус сгибателей, а на противоположной - наоборот.
Шейные позные рефлексы Изменение тонуса мышц конечностей при наклоне головы вправо (а) и влево (б): при повороте головы в сторону на одноименной стороне увеличивается тонус разгибателей и снижается тонус сгибателей, а на противоположной - наоборот.
 Ритмические рефлексы На уровне спинного мозга замыкаются также ритмические рефлексы – многократное повторное сгибание и разгибание конечностей. Примерами могут служить чесательный и шагательный рефлексы. Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
Ритмические рефлексы На уровне спинного мозга замыкаются также ритмические рефлексы – многократное повторное сгибание и разгибание конечностей. Примерами могут служить чесательный и шагательный рефлексы. Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
 Спинальные вегетативные рефлексы • • • На уровне спинного мозга осуществляются многие типы сегментарных вегетативных рефлексов. К ним относят: (1) изменения сосудистого тонуса в результате локального нагревания кожи; (2) потоотделение в результате локального нагревания поверхности тела; (3) кишечно-кишечные рефлексы, контролирующие некоторые моторные функции кишечника; (4) брюшинно-кишечные рефлексы, тормозящие моторную активность желудочно-кишечного тракта в ответ на раздражение брюшины; (5) эвакуационные рефлексы для опорожнения полного мочевого пузыря или переполненной толстой кишки. Эти рефлексы осуществляются при участии нейронов вегетативной нервной системы, расположенных в боковых рогах серого вещества. Вегетативные рефлексы спинного мозга осуществляются в ответ на раздражение кожи или внутренних органов и заканчиваются сокращением гладкой мускулатуры.
Спинальные вегетативные рефлексы • • • На уровне спинного мозга осуществляются многие типы сегментарных вегетативных рефлексов. К ним относят: (1) изменения сосудистого тонуса в результате локального нагревания кожи; (2) потоотделение в результате локального нагревания поверхности тела; (3) кишечно-кишечные рефлексы, контролирующие некоторые моторные функции кишечника; (4) брюшинно-кишечные рефлексы, тормозящие моторную активность желудочно-кишечного тракта в ответ на раздражение брюшины; (5) эвакуационные рефлексы для опорожнения полного мочевого пузыря или переполненной толстой кишки. Эти рефлексы осуществляются при участии нейронов вегетативной нервной системы, расположенных в боковых рогах серого вещества. Вегетативные рефлексы спинного мозга осуществляются в ответ на раздражение кожи или внутренних органов и заканчиваются сокращением гладкой мускулатуры.
 Нарушения функций спинного мозга • • В зависимости от характера повреждения, нарушения функций спинного мозга могут быть общими или избирательными. Прекращение поступления импульсов с чувствительных нервов в те или иные сегменты спинного мозга называется деафферентацией. • Деафферентацию можно вызвать у животного путем перерезки задних корешков в одном или нескольких сегментах спинного мозга. Синдром деафферентации сразу после операции выражается в явлениях паралича. Он возникает вследствие выключения механизма обратной связи оперированной конечности со спинным мозгом. В дальнейшем движения оперированной конечности восстанавливаются за счет регулирующих влияний со стороны здоровых конечностей и лежащих выше отделов центральной нервной системы. Эти влияния передаются через вставочные нейроны на мотонейроны деафферентированной конечности. • Восстановление движений деафферентированной конечности происходит, однако, не полностью. Сокращения ее мышц становятся слишком резкими, тонус мускулатуры ослаблен. Движения животного похожи на таковые у человека при спинной сухотке, когда поражаются задние столбы и соответственно проведение афферентных импульсов по спинному мозгу. Животное, лишенное ощущений с деафферентированной конечности, относится к ней как к постороннему телу. • Деафферентация существенно влияет на кровообращение и трофику ткани. Развиваются отек, кровоизлияния, дистрофические процессы, нередко появляются язвы.
Нарушения функций спинного мозга • • В зависимости от характера повреждения, нарушения функций спинного мозга могут быть общими или избирательными. Прекращение поступления импульсов с чувствительных нервов в те или иные сегменты спинного мозга называется деафферентацией. • Деафферентацию можно вызвать у животного путем перерезки задних корешков в одном или нескольких сегментах спинного мозга. Синдром деафферентации сразу после операции выражается в явлениях паралича. Он возникает вследствие выключения механизма обратной связи оперированной конечности со спинным мозгом. В дальнейшем движения оперированной конечности восстанавливаются за счет регулирующих влияний со стороны здоровых конечностей и лежащих выше отделов центральной нервной системы. Эти влияния передаются через вставочные нейроны на мотонейроны деафферентированной конечности. • Восстановление движений деафферентированной конечности происходит, однако, не полностью. Сокращения ее мышц становятся слишком резкими, тонус мускулатуры ослаблен. Движения животного похожи на таковые у человека при спинной сухотке, когда поражаются задние столбы и соответственно проведение афферентных импульсов по спинному мозгу. Животное, лишенное ощущений с деафферентированной конечности, относится к ней как к постороннему телу. • Деафферентация существенно влияет на кровообращение и трофику ткани. Развиваются отек, кровоизлияния, дистрофические процессы, нередко появляются язвы.
 Нарушения функций спинного мозга • В случаях изолированного поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх. Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность. • Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц. Они теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом. • В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так корковая иннервация этих центров является двусторонней.
Нарушения функций спинного мозга • В случаях изолированного поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх. Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность. • Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц. Они теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом. • В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так корковая иннервация этих центров является двусторонней.
 При травмах у человека в ряде случаев происходит полное или половинное пересечение спинного мозга. • При половинном латеральном повреждении спинного мозга развивается синдром Броун-Секара. Он проявляется в том, что на стороне поражения спинного мозга (ниже места поражения) развивается паралич двигательной системы вследствие повреждения пирамидных путей. На стороне поражения (ниже места поражения) нарушается проприоцептивная чувствительность. Это обусловлено тем, что восходящие пути глубокой чувствительности идут по своей стороне спинного мозга до продолговатого мозга, где происходит их перекрест. • На противоположной стороне туловища нарушается болевая чувствительность, так как проводящие пути болевой чувствительности кожи идут от спинального ганглия в задний рог спинного мозга, где переключаются на новый нейрон, аксон которого переходит на противоположную сторону. В итоге если повреждена левая половина спинного мозга, то исчезает болевая чувствительность правой половины туловища ниже повреждения. На противоположной поражению стороне движения сохраняются.
При травмах у человека в ряде случаев происходит полное или половинное пересечение спинного мозга. • При половинном латеральном повреждении спинного мозга развивается синдром Броун-Секара. Он проявляется в том, что на стороне поражения спинного мозга (ниже места поражения) развивается паралич двигательной системы вследствие повреждения пирамидных путей. На стороне поражения (ниже места поражения) нарушается проприоцептивная чувствительность. Это обусловлено тем, что восходящие пути глубокой чувствительности идут по своей стороне спинного мозга до продолговатого мозга, где происходит их перекрест. • На противоположной стороне туловища нарушается болевая чувствительность, так как проводящие пути болевой чувствительности кожи идут от спинального ганглия в задний рог спинного мозга, где переключаются на новый нейрон, аксон которого переходит на противоположную сторону. В итоге если повреждена левая половина спинного мозга, то исчезает болевая чувствительность правой половины туловища ниже повреждения. На противоположной поражению стороне движения сохраняются.
 Спинальный шок • • • После полного пересечения спинного мозга возникает спинальный шок. Это явление заключается в том, что все центры ниже перерезки перестают организовывать присущие им рефлексы. Нарушение рефлекторной деятельности после пересечения спинного мозга у разных животных длится разное время. У лягушек оно исчисляется десятками секунд, у кролика рефлексы восстанавливаются через 10— 15 мин, у собак отдельные рефлексы, например мышечного сокращения, восстанавливаются через несколько часов, другие — через несколько дней (рефлексы регуляции артериального давления), через недели восстанавливаются рефлексы мочеиспускания. У обезьян первые признаки восстановления рефлексов после перерезки спинного мозга появляются через несколько суток; у человека первые спинальные рефлексы восстанавливаются через несколько недель, а то и месяцев. Следовательно, чем сложнее организация ЦНС, тем сильнее контроль вышележащих отделов мозга над нижележащими. То, что причиной шока является нарушение супраспинальных влияний, доказывается повторной перерезкой спинного мозга ниже места первой перерезки. В этом случае спинальный шок не возникает, рефлекторная деятельность спинного мозга сохраняется. По истечении длительного периода времени после шока спинальные рефлексы резко усиливаются, что объясняется устранением тормозного влияния ретикулярной формации ствола мозга на рефлексы спинного мозга.
Спинальный шок • • • После полного пересечения спинного мозга возникает спинальный шок. Это явление заключается в том, что все центры ниже перерезки перестают организовывать присущие им рефлексы. Нарушение рефлекторной деятельности после пересечения спинного мозга у разных животных длится разное время. У лягушек оно исчисляется десятками секунд, у кролика рефлексы восстанавливаются через 10— 15 мин, у собак отдельные рефлексы, например мышечного сокращения, восстанавливаются через несколько часов, другие — через несколько дней (рефлексы регуляции артериального давления), через недели восстанавливаются рефлексы мочеиспускания. У обезьян первые признаки восстановления рефлексов после перерезки спинного мозга появляются через несколько суток; у человека первые спинальные рефлексы восстанавливаются через несколько недель, а то и месяцев. Следовательно, чем сложнее организация ЦНС, тем сильнее контроль вышележащих отделов мозга над нижележащими. То, что причиной шока является нарушение супраспинальных влияний, доказывается повторной перерезкой спинного мозга ниже места первой перерезки. В этом случае спинальный шок не возникает, рефлекторная деятельность спинного мозга сохраняется. По истечении длительного периода времени после шока спинальные рефлексы резко усиливаются, что объясняется устранением тормозного влияния ретикулярной формации ствола мозга на рефлексы спинного мозга.
 Физиология головного мозга Отделы головного мозга 1 - конечный мозг; 2 - промежуточный мозг; 3 - средний мозг; 4 - мост; 5 - мозжечок (задний мозг); 6 - продолговатый мозг.
Физиология головного мозга Отделы головного мозга 1 - конечный мозг; 2 - промежуточный мозг; 3 - средний мозг; 4 - мост; 5 - мозжечок (задний мозг); 6 - продолговатый мозг.
 Физиология продолговатого мозга и моста Структурные образования продолговатого мозга. 1. V–XII пары черепных нервов. 2. Тонкое и клиновидное ядра. 3. Ядро нижней оливы. 4. Ядра ретикулярной формации. Через продолговатый и задний мозг проходят нисходящие пути (кортикоспинальный и экстрапирамидный, ретикуло– и вестибулоспинальный, отвечающие за перераспределение мышечного тонуса и поддержание позы тела).
Физиология продолговатого мозга и моста Структурные образования продолговатого мозга. 1. V–XII пары черепных нервов. 2. Тонкое и клиновидное ядра. 3. Ядро нижней оливы. 4. Ядра ретикулярной формации. Через продолговатый и задний мозг проходят нисходящие пути (кортикоспинальный и экстрапирамидный, ретикуло– и вестибулоспинальный, отвечающие за перераспределение мышечного тонуса и поддержание позы тела).
 Функции продолговатого мозга и моста • • Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего мозга. Следовательно, жизнь бульбарных животных осуществляется за счет деятельности спинного и продолговатого мозга. У таких животных отсутствуют произвольные движения, отмечается потеря всех видов чувствительности, нарушается регуляция температуры тела (теплокровное животное превращается в холоднокровное). У бульбарных животных сохраняются рефлекторные реакции организма и осуществляется регуляция функций внутренних органов. У человека бульбарные нарушения проявляются нарушением глотания и фонации. В тяжелых случаях больные совершенно не могут глотать и говорить (признаки поражения ядер IX, X, XII пар черепных нервов). При поражении ядер V, VII и IX пар черепных нервов появляются, расстройства жевания, мимики, движения головой. Разрушение центров ромбовидного мозга приводит к мгновенной смерти из-за остановки сердца и остановки дыхания, паралича сосудов и падения АД (сосудистый коллапс). Основные функции продолговатого мозга и моста - сенсорная, рефлекторная и проводниковая.
Функции продолговатого мозга и моста • • Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего мозга. Следовательно, жизнь бульбарных животных осуществляется за счет деятельности спинного и продолговатого мозга. У таких животных отсутствуют произвольные движения, отмечается потеря всех видов чувствительности, нарушается регуляция температуры тела (теплокровное животное превращается в холоднокровное). У бульбарных животных сохраняются рефлекторные реакции организма и осуществляется регуляция функций внутренних органов. У человека бульбарные нарушения проявляются нарушением глотания и фонации. В тяжелых случаях больные совершенно не могут глотать и говорить (признаки поражения ядер IX, X, XII пар черепных нервов). При поражении ядер V, VII и IX пар черепных нервов появляются, расстройства жевания, мимики, движения головой. Разрушение центров ромбовидного мозга приводит к мгновенной смерти из-за остановки сердца и остановки дыхания, паралича сосудов и падения АД (сосудистый коллапс). Основные функции продолговатого мозга и моста - сенсорная, рефлекторная и проводниковая.
 Сенсорные функции Продолговатый мозг регулирует ряд сенсорных функций: • рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; • первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; • рецепцию слуховых раздражений — в ядре улиткового нерва; • рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. • В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). • На уровне продолговатого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
Сенсорные функции Продолговатый мозг регулирует ряд сенсорных функций: • рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; • первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; • рецепцию слуховых раздражений — в ядре улиткового нерва; • рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. • В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). • На уровне продолговатого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
.") Рефлекторные функции • • Через продолговатый мозг осуществляются следующие рефлексы: Жизненно важные рефлексы (дыхание). Сердечно-сосудистые рефлексы, регулирующие деятельность сердца и кровеносных сосудов (регуляция сосудистого тонуса). Защитные рефлексы: кашель, чиханье, мигание, слезоотделение, рвота. Пищевые рефлексы: сосание, глотание, секреция пищеварительных желез. Рефлексы поддержания позы. Вестибуло-вегетативные рефлексы. • • • Рефлекторные центры продолговатого мозга: 1. Дыхательный 2. Сосудодвигательный (вазомоторный) центр 3. Слюноотделительный 4. Сосательный 5. Жевательный 6. Глотательный 7. Рвоты 8. Кашля 9. Чихания 10. Моргания 11. Слезоотделения 12. Регуляции тонуса скелетных мышц. • •
Рефлекторные функции • • Через продолговатый мозг осуществляются следующие рефлексы: Жизненно важные рефлексы (дыхание). Сердечно-сосудистые рефлексы, регулирующие деятельность сердца и кровеносных сосудов (регуляция сосудистого тонуса). Защитные рефлексы: кашель, чиханье, мигание, слезоотделение, рвота. Пищевые рефлексы: сосание, глотание, секреция пищеварительных желез. Рефлексы поддержания позы. Вестибуло-вегетативные рефлексы. • • • Рефлекторные центры продолговатого мозга: 1. Дыхательный 2. Сосудодвигательный (вазомоторный) центр 3. Слюноотделительный 4. Сосательный 5. Жевательный 6. Глотательный 7. Рвоты 8. Кашля 9. Чихания 10. Моргания 11. Слезоотделения 12. Регуляции тонуса скелетных мышц. • •
 Дыхательный центр • • • Дыхательным центром называют совокупность нервных клеток, расположенных в разных отделах центральной нервной системы, обеспечивающих координированную ритмическую деятельность дыхательных мышц и приспособление дыхания к изменяющимся условиям внешней и внутренней среды организма. Нейроны ДЦ, расположенные в ретикулярной формации продолговатого мозга, составляют дыхательный центр в узком смысле слова. Нейронам ДЦ свойственна ритмическая автоматия. Нарушение функции этих клеток приводит к прекращению дыхания вследствие паралича дыхательных мышц. Дыхательные нейроны, активность которых вызывает инспирацию (вдох) или экспирацию (выдох), называются соответственно инспираторными и экспираторными нейронами. Инспираторные и экспираторные нейроны формируют соответствующие центры и иннервируют дыхательные мышцы. Автоматическое возбуждение дыхательного центра обусловлено протекающими в нем самом процессами обмена веществ и его высокой чувствительностью к углекислоте. Автоматия центра регулируется нервными импульсами, приходящими от рецепторов легких, сосудистых рефлексогенных зон, дыхательных и скелетных мышц, а также импульсами из вышележащих отделов центральной нервной системы и, наконец, гуморальными влияниями. В верхней части моста находится так называемый пневмотаксический центр, который контролирует деятельность расположенных ниже дыхательных центров вдоха и выдоха и обеспечивает нормальные адаптивное дыхание.
Дыхательный центр • • • Дыхательным центром называют совокупность нервных клеток, расположенных в разных отделах центральной нервной системы, обеспечивающих координированную ритмическую деятельность дыхательных мышц и приспособление дыхания к изменяющимся условиям внешней и внутренней среды организма. Нейроны ДЦ, расположенные в ретикулярной формации продолговатого мозга, составляют дыхательный центр в узком смысле слова. Нейронам ДЦ свойственна ритмическая автоматия. Нарушение функции этих клеток приводит к прекращению дыхания вследствие паралича дыхательных мышц. Дыхательные нейроны, активность которых вызывает инспирацию (вдох) или экспирацию (выдох), называются соответственно инспираторными и экспираторными нейронами. Инспираторные и экспираторные нейроны формируют соответствующие центры и иннервируют дыхательные мышцы. Автоматическое возбуждение дыхательного центра обусловлено протекающими в нем самом процессами обмена веществ и его высокой чувствительностью к углекислоте. Автоматия центра регулируется нервными импульсами, приходящими от рецепторов легких, сосудистых рефлексогенных зон, дыхательных и скелетных мышц, а также импульсами из вышележащих отделов центральной нервной системы и, наконец, гуморальными влияниями. В верхней части моста находится так называемый пневмотаксический центр, который контролирует деятельность расположенных ниже дыхательных центров вдоха и выдоха и обеспечивает нормальные адаптивное дыхание.
 центр • • • Сосудодвигательный центр — это совокупность структур ЦНС, расположенных") Сосудодвигательный (вазомоторный) центр • • • Сосудодвигательный центр — это совокупность структур ЦНС, расположенных на разных уровнях, основные из которых включают в себя спинной мозг, продолговатый мозг, гипоталамус, кору больших полушарий головного мозга. Главный компонент сосудодвигательного центра расположен в продолговатом мозге и состоит из прессорной и депрессорной зоны. Эти зоны не имеют четких границ и перекрывают друга. Однако только структуры прессорной области по нисходящим внутрицентральным путям связаны с центрами спинного мозга, которые передают эфферентные сосудодвигательные влияния к гладким мышцам сосудов. Сосудодвигательный центр продолговатого мозга находится в состоянии постоянного тонического возбуждения. Отсюда к гладким мышцам сосудов поступает постоянный поток тонической сосудосуживающей импульсации, которая «наслаивается» на миогенный базальный тонус. Депрессорная зона собственных выходов к центрам спинного мозга не имеет и оказывает свое влияние через изменение тонуса прессорной области. Сосудистодвигательный центр продолговатого мозга — это центр автоматической саморегуляции артериального давления. Он осуществляет «быстрые» рефлекторные ответы при гипоксии, гиперкапнии, при разных формах усиления мышечной деятельности. В составе сосудодвигательного центра продолговатого мозга есть кардиоингибирующие нейроны.
Сосудодвигательный (вазомоторный) центр • • • Сосудодвигательный центр — это совокупность структур ЦНС, расположенных на разных уровнях, основные из которых включают в себя спинной мозг, продолговатый мозг, гипоталамус, кору больших полушарий головного мозга. Главный компонент сосудодвигательного центра расположен в продолговатом мозге и состоит из прессорной и депрессорной зоны. Эти зоны не имеют четких границ и перекрывают друга. Однако только структуры прессорной области по нисходящим внутрицентральным путям связаны с центрами спинного мозга, которые передают эфферентные сосудодвигательные влияния к гладким мышцам сосудов. Сосудодвигательный центр продолговатого мозга находится в состоянии постоянного тонического возбуждения. Отсюда к гладким мышцам сосудов поступает постоянный поток тонической сосудосуживающей импульсации, которая «наслаивается» на миогенный базальный тонус. Депрессорная зона собственных выходов к центрам спинного мозга не имеет и оказывает свое влияние через изменение тонуса прессорной области. Сосудистодвигательный центр продолговатого мозга — это центр автоматической саморегуляции артериального давления. Он осуществляет «быстрые» рефлекторные ответы при гипоксии, гиперкапнии, при разных формах усиления мышечной деятельности. В составе сосудодвигательного центра продолговатого мозга есть кардиоингибирующие нейроны.
 Слюноотделительный центр • • • Отделение слюны является сложным рефлекторным актом, вызванным раздражением рецепторов ротовой полости пищей или другими веществами, а также раздражением зрительных и обонятельных рецепторов внешним видом и запахом пищи, видом обстановки, в которой происходит прием пищи. Во время приема пищи раздражаются тактильные, температурные и вкусовые рецепторы слизистой оболочки полости рта. Афферентные импульсы от них по чувствительным волокнам тройничного, языкоглоточного, лицевого и блуждающего нервов достигают слюноотделительного центра продолговатого мозга. При возбуждении верхнего слюноотделительного ядра центра эфферентные импульсы по преганглионарным парасимпатическим волокнам барабанной струны достигают подъязычного и нижнечелюстного ганглиев, где они переключаются на постганглионарные, парасимпатические волокна подъязычного нерва, иннервирующего подъязычные и подчелюстные слюнные железы. • При возбуждении нижнего слюноотделительного ядра центра эфферентные импульсы по преганглионарным парасимпатическим волокнам языкоглоточного нерва достигают ушного ганглия, где они переключаются на постганглионарные волокна ушновисочного нерва, иннервирующего околоушные слюнные железы. Центры слюноотделения рефлекторно могут не только возбуждаться, но и тормозиться. Например, при болевом раздражении, во время отрицательных эмоций и при умственном напряжении слюноотделение резко ослабляется или прекращается ( «пересыхает во рту» ).
Слюноотделительный центр • • • Отделение слюны является сложным рефлекторным актом, вызванным раздражением рецепторов ротовой полости пищей или другими веществами, а также раздражением зрительных и обонятельных рецепторов внешним видом и запахом пищи, видом обстановки, в которой происходит прием пищи. Во время приема пищи раздражаются тактильные, температурные и вкусовые рецепторы слизистой оболочки полости рта. Афферентные импульсы от них по чувствительным волокнам тройничного, языкоглоточного, лицевого и блуждающего нервов достигают слюноотделительного центра продолговатого мозга. При возбуждении верхнего слюноотделительного ядра центра эфферентные импульсы по преганглионарным парасимпатическим волокнам барабанной струны достигают подъязычного и нижнечелюстного ганглиев, где они переключаются на постганглионарные, парасимпатические волокна подъязычного нерва, иннервирующего подъязычные и подчелюстные слюнные железы. • При возбуждении нижнего слюноотделительного ядра центра эфферентные импульсы по преганглионарным парасимпатическим волокнам языкоглоточного нерва достигают ушного ганглия, где они переключаются на постганглионарные волокна ушновисочного нерва, иннервирующего околоушные слюнные железы. Центры слюноотделения рефлекторно могут не только возбуждаться, но и тормозиться. Например, при болевом раздражении, во время отрицательных эмоций и при умственном напряжении слюноотделение резко ослабляется или прекращается ( «пересыхает во рту» ).
 Центр сосательного рефлекса • Сосания центр - нервный центр, расположенный в продолговатом мозге. Сосательные движения возникают прикосновении к губам новорожденного. Рефлекс осуществляется при раздражении чувствительных окончаний тройничного нерва, возбуждение с которого переключается в продолговатом мозге на моторные ядра лицевого и подъязычного нервов.
Центр сосательного рефлекса • Сосания центр - нервный центр, расположенный в продолговатом мозге. Сосательные движения возникают прикосновении к губам новорожденного. Рефлекс осуществляется при раздражении чувствительных окончаний тройничного нерва, возбуждение с которого переключается в продолговатом мозге на моторные ядра лицевого и подъязычного нервов.
 Жевательный центр • • Жевательный рефлекс — это механический процесс размельчения пищи в ротовой полости. Жевание представляет двигательный акт, который может осуществляться рефлекторно в ответ на раздражение рецепторов ротовой полости и состоит в перемещении нижней челюсти по отношению к верхней. Центр этого рефлекса находится в продолговатом мозге, и потому жевание может быть вызвано у бульбарных животных (животные с сохраненным продолговатым мозгом). Более тонкая регуляция акта жевания достигается лишь при целости таламуса и моторных зон коры. Жевательные мышцы сокращаются рефлекторным путем. Когда пища попадает в рот, раздражаются находящиеся в слизистой оболочке ротовой полости рецепторы: осязательные, температурные, вкусовые, а иногда и болевые. При поступлении в центральную нервную систему афферентных импульсов от этих рецепторов, а также рецепторов жевательных мышц по эфферентным нервам регулируются движения, а кроме того, сила сокращения жевательных мышц, соответствующая консистенции пищи. Афферентные импульсы проводятся по ветвям тройничных нервов, лицевому и языкоглоточному нервам, эфферентные — по двигательным нервам жевательных мышц, по лицевому и по подъязычному нервам
Жевательный центр • • Жевательный рефлекс — это механический процесс размельчения пищи в ротовой полости. Жевание представляет двигательный акт, который может осуществляться рефлекторно в ответ на раздражение рецепторов ротовой полости и состоит в перемещении нижней челюсти по отношению к верхней. Центр этого рефлекса находится в продолговатом мозге, и потому жевание может быть вызвано у бульбарных животных (животные с сохраненным продолговатым мозгом). Более тонкая регуляция акта жевания достигается лишь при целости таламуса и моторных зон коры. Жевательные мышцы сокращаются рефлекторным путем. Когда пища попадает в рот, раздражаются находящиеся в слизистой оболочке ротовой полости рецепторы: осязательные, температурные, вкусовые, а иногда и болевые. При поступлении в центральную нервную систему афферентных импульсов от этих рецепторов, а также рецепторов жевательных мышц по эфферентным нервам регулируются движения, а кроме того, сила сокращения жевательных мышц, соответствующая консистенции пищи. Афферентные импульсы проводятся по ветвям тройничных нервов, лицевому и языкоглоточному нервам, эфферентные — по двигательным нервам жевательных мышц, по лицевому и по подъязычному нервам
 Центр глотания • Глотание является сложно координированным рефлекторным актом. В его реализации участвуют мышцы полости рта, глотки и начала пищевода. Акт глотания состоит из двух фаз: 1) формирование пищевого комка и подведение его к полости глотки; 2) проглатывание, при котором мышцы глотки сокращаются и одновременно поднимается небная занавеска, а надгортанник опускается. Первая часть этого механизма регулируется произвольно, а вторая - непроизвольно - безусловнорефлекторно. В акте глотания участвуют афферентные системы тройничного, языкоглоточного и блуждающего нервов. Центр глотания представляет функциональное объединение многих ядер, обеспечивающих осуществление этого рефлекторного акта. Ротовая стадия Глоточная стадия Пищеводная стадия
Центр глотания • Глотание является сложно координированным рефлекторным актом. В его реализации участвуют мышцы полости рта, глотки и начала пищевода. Акт глотания состоит из двух фаз: 1) формирование пищевого комка и подведение его к полости глотки; 2) проглатывание, при котором мышцы глотки сокращаются и одновременно поднимается небная занавеска, а надгортанник опускается. Первая часть этого механизма регулируется произвольно, а вторая - непроизвольно - безусловнорефлекторно. В акте глотания участвуют афферентные системы тройничного, языкоглоточного и блуждающего нервов. Центр глотания представляет функциональное объединение многих ядер, обеспечивающих осуществление этого рефлекторного акта. Ротовая стадия Глоточная стадия Пищеводная стадия
 Рвотный центр • Рвотный центр - нервный центр, расположенный в продолговатом мозге. • Рвота - рефлекторный акт, возникающий при раздражении рецепторов глотки и желудка, а также при раздражении вестибулорецепторов и некоторых других. Импульсы, поступающие от рецепторов по афферентным волокнам в продолговатый мозг, проходят ко многим эффекторным нейронам, находящимся как в продолговатом, так и в спинном мозге. • Во время акта рвоты происходит открытие входа в желудок, сокращение мускулатуры кишечника и стенок желудка, сокращение мышц брюшного пресса и диафрагмы, мышц глотки, гортани, языка и рта, секреция слюны и слез. • Во время акта рвоты изменяется состояние многих центров ЦНС в связи с участием в нем ретикулярной формации ствола мозга. Последняя своими множественными связями обеспечивает функциональное объединение и согласование деятельности нейронов, расположенных в разных участках продолговатого и спинного мозга и изменяет состояние вышележащих центров.
Рвотный центр • Рвотный центр - нервный центр, расположенный в продолговатом мозге. • Рвота - рефлекторный акт, возникающий при раздражении рецепторов глотки и желудка, а также при раздражении вестибулорецепторов и некоторых других. Импульсы, поступающие от рецепторов по афферентным волокнам в продолговатый мозг, проходят ко многим эффекторным нейронам, находящимся как в продолговатом, так и в спинном мозге. • Во время акта рвоты происходит открытие входа в желудок, сокращение мускулатуры кишечника и стенок желудка, сокращение мышц брюшного пресса и диафрагмы, мышц глотки, гортани, языка и рта, секреция слюны и слез. • Во время акта рвоты изменяется состояние многих центров ЦНС в связи с участием в нем ретикулярной формации ствола мозга. Последняя своими множественными связями обеспечивает функциональное объединение и согласование деятельности нейронов, расположенных в разных участках продолговатого и спинного мозга и изменяет состояние вышележащих центров.
 Центр чихания • Чихание представляет сложный рефлекторный акт, который возникает при раздражении рецепторов тройничного нерва в носовой полости. В начале чихания мягкое небо поднимается и закрывает внутренне носовое отверстие; затем за счет сокращения мышц выдоха создается повышенное давление в грудной полости, после этого носовое отверстие резко открывается и весь воздух с силой выходит через нос, удаляя вещество, раздражающее слизистую оболочку носа. В акте чихания принимают участие эфферентные волокна языкоглоточного, блуждающего, подъязычного и некоторых спинальных нервов.
Центр чихания • Чихание представляет сложный рефлекторный акт, который возникает при раздражении рецепторов тройничного нерва в носовой полости. В начале чихания мягкое небо поднимается и закрывает внутренне носовое отверстие; затем за счет сокращения мышц выдоха создается повышенное давление в грудной полости, после этого носовое отверстие резко открывается и весь воздух с силой выходит через нос, удаляя вещество, раздражающее слизистую оболочку носа. В акте чихания принимают участие эфферентные волокна языкоглоточного, блуждающего, подъязычного и некоторых спинальных нервов.
 Центр кашля • Кашель, как и чихание, является защитным дыхательным рефлексом, возникающим при раздражении слизистой оболочки гортани, трахеи и бронхов. При кашле, в отличие от чихания, замыкается не носовое отверстие, а голосовая щель, которая после создания необходимого давления в легких резко открывается и сильная струя воздуха удаляет раздражающий фактор. В акте кашля участвуют те же эфферентные волокна, что и в акте чихания, а афферентные сигналы передаются по волокнам блуждающего нерва.
Центр кашля • Кашель, как и чихание, является защитным дыхательным рефлексом, возникающим при раздражении слизистой оболочки гортани, трахеи и бронхов. При кашле, в отличие от чихания, замыкается не носовое отверстие, а голосовая щель, которая после создания необходимого давления в легких резко открывается и сильная струя воздуха удаляет раздражающий фактор. В акте кашля участвуют те же эфферентные волокна, что и в акте чихания, а афферентные сигналы передаются по волокнам блуждающего нерва.
 Центр мигания • Мигание – защитный рефлекс, происходит при раздражении роговой и конъюнктивальной оболочек глаза, иннервируемых афферентными волокнами тройничного нерва. Поступающие от них импульсы в продолговатом мозге переключаются на двигательное ядро лицевого нерва, волокна которого иннервируют круговую мышцу глаза; в результате происходит закрывание век.
Центр мигания • Мигание – защитный рефлекс, происходит при раздражении роговой и конъюнктивальной оболочек глаза, иннервируемых афферентными волокнами тройничного нерва. Поступающие от них импульсы в продолговатом мозге переключаются на двигательное ядро лицевого нерва, волокна которого иннервируют круговую мышцу глаза; в результате происходит закрывание век.
 Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса мышц. • Установочные рефлексы, обеспечивают тонус мускулатуры, необходимый для поддержания позы и осуществления рабочих актов. • Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо. • Импульсы от вестибулярных ядер усиливают тонус мышц разгибателей, что необходимо для организации позы. Неспецифические отделы продолговатого мозга (ядра РФ) наоборот снижают тонус мышц, в том числе и разгибателей. • Лабиринтные рефлексы, способствуют правильному распределению мышечного тонуса между отдельными группами мышц при изменении позы тела. • • Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы соответствующей моменту прямолинейного или вращательного движения. •
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса мышц. • Установочные рефлексы, обеспечивают тонус мускулатуры, необходимый для поддержания позы и осуществления рабочих актов. • Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо. • Импульсы от вестибулярных ядер усиливают тонус мышц разгибателей, что необходимо для организации позы. Неспецифические отделы продолговатого мозга (ядра РФ) наоборот снижают тонус мышц, в том числе и разгибателей. • Лабиринтные рефлексы, способствуют правильному распределению мышечного тонуса между отдельными группами мышц при изменении позы тела. • • Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы соответствующей моменту прямолинейного или вращательного движения. •
,") Лабиринтные рефлексы (распределение тонуса между отдельными группами мышц и установки определенной позы),
Лабиринтные рефлексы (распределение тонуса между отдельными группами мышц и установки определенной позы),
 Тонические рефлексы ствола мозга Статические наблюдаются в состоянии покоя Познотонические обеспечивают перераспреде ление мышечного тонуса при изменении позы, т. е. относительного положения частей тела Выпрямительные направлены на возвращение к адекватной для данного вида позе из неестественного положения Статокинетические возникают при движении тела с линейным или угловым ускорением Линейные, лифтные рефлексы наблюдаются при движении с линейным ускорением. Вращательные рефлексы (нистагм головы и глаз) наблюдается при движении с угловым ускорением
Тонические рефлексы ствола мозга Статические наблюдаются в состоянии покоя Познотонические обеспечивают перераспреде ление мышечного тонуса при изменении позы, т. е. относительного положения частей тела Выпрямительные направлены на возвращение к адекватной для данного вида позе из неестественного положения Статокинетические возникают при движении тела с линейным или угловым ускорением Линейные, лифтные рефлексы наблюдаются при движении с линейным ускорением. Вращательные рефлексы (нистагм головы и глаз) наблюдается при движении с угловым ускорением
 Проводниковые функции. • Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Через восходящие пути и черепные нервы продолговатый мозг получает импульсы от рецепторов мышц лица, шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, носовой и ротовой полости, от рецепторов слуха, вестибулярного аппарата, рецепторов гортани, трахеи, легких, интерорецепторов пищеварительного аппарата и сердечно-сосудистой системы. Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
Проводниковые функции. • Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Через восходящие пути и черепные нервы продолговатый мозг получает импульсы от рецепторов мышц лица, шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, носовой и ротовой полости, от рецепторов слуха, вестибулярного аппарата, рецепторов гортани, трахеи, легких, интерорецепторов пищеварительного аппарата и сердечно-сосудистой системы. Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
 Симптомы повреждения. • • Повреждение левой или правой половины продолговато мозга выше перекреста восходящих путей проприоцептивной чувствительности вызывает на стороне повреждения нарушения чувствительности и работы мышц лица и головы. В то же время на противоположной стороне относительно стороны повреждения наблюдаются нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это объясняется тем, что восходящие и нисходящие проводящие пути из спинного мозга и в спинной мозг перекрещиваются, а ядра черепных нервов иннервируют свою половину головы, т. е. черепные нервы не перекрещиваются. Повреждение моста мозга приводит к утрате разных видов чувствительности, волевой моторики, терморегуляции, но дыхание, величина артериального давления, рефлекторная активность при этом сохраняются.
Симптомы повреждения. • • Повреждение левой или правой половины продолговато мозга выше перекреста восходящих путей проприоцептивной чувствительности вызывает на стороне повреждения нарушения чувствительности и работы мышц лица и головы. В то же время на противоположной стороне относительно стороны повреждения наблюдаются нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это объясняется тем, что восходящие и нисходящие проводящие пути из спинного мозга и в спинной мозг перекрещиваются, а ядра черепных нервов иннервируют свою половину головы, т. е. черепные нервы не перекрещиваются. Повреждение моста мозга приводит к утрате разных видов чувствительности, волевой моторики, терморегуляции, но дыхание, величина артериального давления, рефлекторная активность при этом сохраняются.
 защитные рефлексы (слезотечение,") Заключение • Рефлекторная функция продолговатого и заднего мозга обеспечивает: • 1) защитные рефлексы (слезотечение, мигание, кашель, рвоту, чиханье); • 2) рефлексы голосообразования (ядра X, XII, VII черепномозговых нервов), дыхательный центр регулируют поток воздуха; • 3) рефлексы поддержания позы (лабиринтные рефлексы). Статические рефлексы поддерживают тонус мышц для сохранения позы тела, статокинетические перераспределяют тонус мышц для принятия позы, соответствующей моменту прямолинейного или вращательного движения; • 4) центры, расположенные в продолговатом и заднем мозге, регулируют деятельность многих систем: • Пищеварительной системы – регуляция секреции желудочных, кишечных желез, поджелудочной железы, секреторных клеток печени, слюнных желез, обеспечивает рефлексы сосания, жевания, глотания. • Дыхательный центр – запуск и регуляция вдоха и выдоха. • Сосудо-двигательный центр осуществляет регуляцию сосудистого тонуса.
Заключение • Рефлекторная функция продолговатого и заднего мозга обеспечивает: • 1) защитные рефлексы (слезотечение, мигание, кашель, рвоту, чиханье); • 2) рефлексы голосообразования (ядра X, XII, VII черепномозговых нервов), дыхательный центр регулируют поток воздуха; • 3) рефлексы поддержания позы (лабиринтные рефлексы). Статические рефлексы поддерживают тонус мышц для сохранения позы тела, статокинетические перераспределяют тонус мышц для принятия позы, соответствующей моменту прямолинейного или вращательного движения; • 4) центры, расположенные в продолговатом и заднем мозге, регулируют деятельность многих систем: • Пищеварительной системы – регуляция секреции желудочных, кишечных желез, поджелудочной железы, секреторных клеток печени, слюнных желез, обеспечивает рефлексы сосания, жевания, глотания. • Дыхательный центр – запуск и регуляция вдоха и выдоха. • Сосудо-двигательный центр осуществляет регуляцию сосудистого тонуса.
 Средний мозг представлен четверохолмием и ножками мозга. Эволюция среднего мозга связана с развитием зрения. • В строении среднего мозга полностью утрачиваются сегментарные признаки. Его клеточные элементы образуют ядра, относящиеся непосредственно к среднему мозгу, а также ядра ретикулярной формации, контролирующие состояние бодрствования. • Структурные единицы среднего мозга: 1) бугры четверохолмия; 2) красное ядро; 3) черная субстанция; 4) ядра III–IV пары черепно-мозговых нервов; 5) ядра ретикулярной формации. • • • Функции среднего мозга –сенсорная, рефлекторная и проводниковая.
Средний мозг представлен четверохолмием и ножками мозга. Эволюция среднего мозга связана с развитием зрения. • В строении среднего мозга полностью утрачиваются сегментарные признаки. Его клеточные элементы образуют ядра, относящиеся непосредственно к среднему мозгу, а также ядра ретикулярной формации, контролирующие состояние бодрствования. • Структурные единицы среднего мозга: 1) бугры четверохолмия; 2) красное ядро; 3) черная субстанция; 4) ядра III–IV пары черепно-мозговых нервов; 5) ядра ретикулярной формации. • • • Функции среднего мозга –сенсорная, рефлекторная и проводниковая.
 • Сенсорные функции реализуются за счет поступления в него зрительной и слуховой информации. • Рефлекторная функция реализуется за счет ядра блокового нерва, ядер глазодвигательного нерва, красного ядра и черного вещества. • Проводниковая функция: через средний мозг проходят все восходящие пути к таламусу, большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. • Мезенцефальное животное – это животное, у которого произведена перерезка ствола мозга выше бугров четверохолмия. Оно может осуществлять все спинальные, бульбарные рефлексы и также рефлексы за счет ядер среднего мозга.
• Сенсорные функции реализуются за счет поступления в него зрительной и слуховой информации. • Рефлекторная функция реализуется за счет ядра блокового нерва, ядер глазодвигательного нерва, красного ядра и черного вещества. • Проводниковая функция: через средний мозг проходят все восходящие пути к таламусу, большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. • Мезенцефальное животное – это животное, у которого произведена перерезка ствола мозга выше бугров четверохолмия. Оно может осуществлять все спинальные, бульбарные рефлексы и также рефлексы за счет ядер среднего мозга.
 Функции четверохолмия • • • Бугры четверохолмия выполняют афферентную функцию, остальные образования – эфферентную. Верхние холмики четверохолмия получают импульсы от сетчатки глаз. Являясь первичными зрительными центрами, верхние холмики обеспечивают сторожевые реакции с поворотом головы и глазных яблок к источнику внезапных зрительных раздражений. Верхние холмы с ядрами III–IV черепно-мозговых нервов обеспечивают реакцию конвергенции (схождение глазных яблок к срединной линии), движение глазных яблок. Нижние холмы четверохолмия получают импульсы от ядер слуховых нервов, лежащих в продолговатом мозге. Являясь первичными слуховыми центрами, нижние холмики обеспечивают сторожевые реакции с поворотом головы, движением ушных раковин к источнику внезапных слуховых раздражений, натяжением барабанной перепонки и слуховых косточек. Бугры четверохолмия тесным образом взаимодействуют с ядрами III–IV пар черепно-мозговых нервов, красным ядром, со зрительным трактом. За счет этого взаимодействия происходит обеспечение передними буграми ориентировочной рефлекторной реакции на свет, а задними – на звук. Обеспечивают жизненно важные рефлексы: старт-рефлекс – двигательная реакция на резкий необычный раздражитель (повышение тонуса сгибателей), ориентир-рефлекс – двигательная реакция на новый раздражитель (поворот тела, головы).
Функции четверохолмия • • • Бугры четверохолмия выполняют афферентную функцию, остальные образования – эфферентную. Верхние холмики четверохолмия получают импульсы от сетчатки глаз. Являясь первичными зрительными центрами, верхние холмики обеспечивают сторожевые реакции с поворотом головы и глазных яблок к источнику внезапных зрительных раздражений. Верхние холмы с ядрами III–IV черепно-мозговых нервов обеспечивают реакцию конвергенции (схождение глазных яблок к срединной линии), движение глазных яблок. Нижние холмы четверохолмия получают импульсы от ядер слуховых нервов, лежащих в продолговатом мозге. Являясь первичными слуховыми центрами, нижние холмики обеспечивают сторожевые реакции с поворотом головы, движением ушных раковин к источнику внезапных слуховых раздражений, натяжением барабанной перепонки и слуховых косточек. Бугры четверохолмия тесным образом взаимодействуют с ядрами III–IV пар черепно-мозговых нервов, красным ядром, со зрительным трактом. За счет этого взаимодействия происходит обеспечение передними буграми ориентировочной рефлекторной реакции на свет, а задними – на звук. Обеспечивают жизненно важные рефлексы: старт-рефлекс – двигательная реакция на резкий необычный раздражитель (повышение тонуса сгибателей), ориентир-рефлекс – двигательная реакция на новый раздражитель (поворот тела, головы).
 Сторожевые рефлексы верхних холмов четверохолмия • Ориентировочные зрительные и слуховые рефлексы - по Павлову рефлекс «Что такое? » . Ориентир-рефлекс возникает на новый раздражитель. У животных они проявляются в повороте головы и тела по направлению к свету и звуку, настораживании ушей. Старт-рефлекс – двигательная реакция на резкий необычный раздражитель (повышение тонуса сгибателей), готовность к борьбе или бегству.
Сторожевые рефлексы верхних холмов четверохолмия • Ориентировочные зрительные и слуховые рефлексы - по Павлову рефлекс «Что такое? » . Ориентир-рефлекс возникает на новый раздражитель. У животных они проявляются в повороте головы и тела по направлению к свету и звуку, настораживании ушей. Старт-рефлекс – двигательная реакция на резкий необычный раздражитель (повышение тонуса сгибателей), готовность к борьбе или бегству.
 • Передние бугры с ядрами III–IV черепно-мозговых нервов обеспечивают реакцию конвергенции (схождение глазных яблок к срединной линии), движение глазных яблок. • На уровне среднего мозга замыкаются также рефлексы, участвующие в обеспечении зрения (зрачковый рефлекс, аккомодация). Эти рефлексы улучшают ориентировку организма в пространстве.
• Передние бугры с ядрами III–IV черепно-мозговых нервов обеспечивают реакцию конвергенции (схождение глазных яблок к срединной линии), движение глазных яблок. • На уровне среднего мозга замыкаются также рефлексы, участвующие в обеспечении зрения (зрачковый рефлекс, аккомодация). Эти рефлексы улучшают ориентировку организма в пространстве.
 Красное ядро • Красное ядро принимает участие в регуляции перераспределения мышечного тонуса, в восстановлении позы тела (повышает тонус сгибателей, понижают тонус разгибателей), поддержании равновесия, подготавливает скелетные мышцы к произвольным и непроизвольным движениям.
Красное ядро • Красное ядро принимает участие в регуляции перераспределения мышечного тонуса, в восстановлении позы тела (повышает тонус сгибателей, понижают тонус разгибателей), поддержании равновесия, подготавливает скелетные мышцы к произвольным и непроизвольным движениям.
 Децеребрационная ригидность • Роль среднего мозга в регуляции мышечного тонуса лучше всего наблюдать на кошке, у которой сделан поперечный разрез между продолговатым и средним мозгом. У такой кошки резко повышается тонус, мышц, особенно разгибателей. Голова запрокидывается назад, резко выпрямляются лапы. Мышцы настолько сильно сокращены, что попытка согнуть конечность заканчивается неудачей - она сейчас же распрямляется. Животное, поставленное на вытянутые, как палки, лапы, может стоять. Такое состояние называется децеребрационной ригидностью. Если разрез сделать выше среднего мозга, то децеребрационная ригидность не возникает. Следовательно, нервные аппараты регуляции мышечного тонуса и функции стояния и ходьбы находятся в среднем мозге.
Децеребрационная ригидность • Роль среднего мозга в регуляции мышечного тонуса лучше всего наблюдать на кошке, у которой сделан поперечный разрез между продолговатым и средним мозгом. У такой кошки резко повышается тонус, мышц, особенно разгибателей. Голова запрокидывается назад, резко выпрямляются лапы. Мышцы настолько сильно сокращены, что попытка согнуть конечность заканчивается неудачей - она сейчас же распрямляется. Животное, поставленное на вытянутые, как палки, лапы, может стоять. Такое состояние называется децеребрационной ригидностью. Если разрез сделать выше среднего мозга, то децеребрационная ригидность не возникает. Следовательно, нервные аппараты регуляции мышечного тонуса и функции стояния и ходьбы находятся в среднем мозге.
 • Явления децеребрационной ригидности возникают как результат нарушения тормозящего влияния красного ядра на тонические рефлексы разгибания, осуществляемые латеральным вестибулярным ядром, лежащим в продолговатом мозге. • Красное ядро принимает участие в регуляции перераспределения мышечного тонуса, в восстановлении позы тела (повышает тонус сгибателей, понижает тонус разгибателей), поддержании равновесия, подготавливает скелетные мышцы к произвольным и непроизвольным движениям. • Красное ядро является переключательным центром связей мозжечка с периферией • Красные ядра не имеют непосредственной связи с рецепторами и эффекторами, но они связаны со всеми отделами центральной нервной системы. К ним подходят нервные волокна от мозжечка, базальных ядер, коры полушарий большого мозга. От красных ядер начинается нисходящий руброспинальный тракт, по которому передаются импульсы к двигательным нейронам спинного мозга.
• Явления децеребрационной ригидности возникают как результат нарушения тормозящего влияния красного ядра на тонические рефлексы разгибания, осуществляемые латеральным вестибулярным ядром, лежащим в продолговатом мозге. • Красное ядро принимает участие в регуляции перераспределения мышечного тонуса, в восстановлении позы тела (повышает тонус сгибателей, понижает тонус разгибателей), поддержании равновесия, подготавливает скелетные мышцы к произвольным и непроизвольным движениям. • Красное ядро является переключательным центром связей мозжечка с периферией • Красные ядра не имеют непосредственной связи с рецепторами и эффекторами, но они связаны со всеми отделами центральной нервной системы. К ним подходят нервные волокна от мозжечка, базальных ядер, коры полушарий большого мозга. От красных ядер начинается нисходящий руброспинальный тракт, по которому передаются импульсы к двигательным нейронам спинного мозга.
 Рефлексы среднего мозга • Ядра среднего мозга обеспечивают поддержание равновесия в условиях покоя и ускорения, с участием нейронов продолговатого мозга и моста. Это проявляется в наличии статических и статокинетических рефлексов • Статические: 1. Позно-тонические рефлексы 2. Выпрямительные рефлексы Статические рефлексы связаны главным образом с возбуждением рецепторов преддверия лабиринта и обеспечивают поддержание позы и равновесия тела при самых разнообразных его статических положениях в пространстве. Позно-тонические рефлексы начинаются от проприорецепторов мышц шеи или вестибулярного аппарата и направлены на перераспределение мышечного тонуса для поддержания позы тела, когда смещаются голова и шея. Например, у человека наклон головы вперед повышает тонус всех мышц сгибателей, а назад – разгибателей. У животных при наклоне головы вниз повышается тонус сгибателей передних конечностей и разгибателей – задних. При подъеме головы наблюдается противоположное: передние конечности разгибаются, а задние - сгибаются.
Рефлексы среднего мозга • Ядра среднего мозга обеспечивают поддержание равновесия в условиях покоя и ускорения, с участием нейронов продолговатого мозга и моста. Это проявляется в наличии статических и статокинетических рефлексов • Статические: 1. Позно-тонические рефлексы 2. Выпрямительные рефлексы Статические рефлексы связаны главным образом с возбуждением рецепторов преддверия лабиринта и обеспечивают поддержание позы и равновесия тела при самых разнообразных его статических положениях в пространстве. Позно-тонические рефлексы начинаются от проприорецепторов мышц шеи или вестибулярного аппарата и направлены на перераспределение мышечного тонуса для поддержания позы тела, когда смещаются голова и шея. Например, у человека наклон головы вперед повышает тонус всех мышц сгибателей, а назад – разгибателей. У животных при наклоне головы вниз повышается тонус сгибателей передних конечностей и разгибателей – задних. При подъеме головы наблюдается противоположное: передние конечности разгибаются, а задние - сгибаются.
 Выпрямительные рефлексы • Выпрямительные рефлексы определяют перераспределение тонуса мышц, приводящее к восстановлению естественной для данного вида животного позы из неестественной. • Рефлекс выпрямления головы (если, например, животное лежит на боку) возникает с рецепторов вестибулярного аппарата, кожи и сетчатки. При этом возбуждаются проприорецепторы мышц шеи и начинается выпрямление туловища. При выключении рецепторов кожи или вестибулярного аппарата выпрямительные рефлексы не нарушаются. При выключении обеих названных рефлексогенных зон выпрямительные рефлексы не возникают. Для осуществления выпрямительных рефлексов, т. е. восстановления нарушенной позы, необходимо обязательное участие красных ядер, их разрушение в эксперименте предотвращает возникновение выпрямительных рефлексов.
Выпрямительные рефлексы • Выпрямительные рефлексы определяют перераспределение тонуса мышц, приводящее к восстановлению естественной для данного вида животного позы из неестественной. • Рефлекс выпрямления головы (если, например, животное лежит на боку) возникает с рецепторов вестибулярного аппарата, кожи и сетчатки. При этом возбуждаются проприорецепторы мышц шеи и начинается выпрямление туловища. При выключении рецепторов кожи или вестибулярного аппарата выпрямительные рефлексы не нарушаются. При выключении обеих названных рефлексогенных зон выпрямительные рефлексы не возникают. Для осуществления выпрямительных рефлексов, т. е. восстановления нарушенной позы, необходимо обязательное участие красных ядер, их разрушение в эксперименте предотвращает возникновение выпрямительных рефлексов.
 • Выпрямительный рефлекс состоит из двух фаз. Первая заключается в том, что животное, как и человек, поднимает голову, которая запускается с рецепторов вестибулярного аппарата. Затем наступает вторая фаза, когда животное встает на лапы, а человек на ноги. Эта фаза выпрямительного рефлекса осуществляется с проприорецепторов мышц шеи, которые раздражаются вследствие изменения положения головы.
• Выпрямительный рефлекс состоит из двух фаз. Первая заключается в том, что животное, как и человек, поднимает голову, которая запускается с рецепторов вестибулярного аппарата. Затем наступает вторая фаза, когда животное встает на лапы, а человек на ноги. Эта фаза выпрямительного рефлекса осуществляется с проприорецепторов мышц шеи, которые раздражаются вследствие изменения положения головы.
 координация актов жевания и") • Черная субстанция обеспечивает три основных функции: • 1) координация актов жевания и глотания, • 2) координация фазных движений пальцев рук, • 3) регуляция мышечного тонуса.
• Черная субстанция обеспечивает три основных функции: • 1) координация актов жевания и глотания, • 2) координация фазных движений пальцев рук, • 3) регуляция мышечного тонуса.
 • Черное вещество регулирует: • акты жевания, глотания, • кровяное давление (патология черного вещества мозга ведет к повышению кровяного давления), • дыхание, • участвует в регуляции пластического тонуса мышц (особенно при выполнении мелких движений пальцами рук) • Поддержание позы. • Гуморальная регуляция тонуса мышц обеспечивается черным веществом путем выделения его пигментными клетками вещества дофамина. • Дефицит дофамина проявляется как болезнь Паркинсона (дрожательный паралич).
• Черное вещество регулирует: • акты жевания, глотания, • кровяное давление (патология черного вещества мозга ведет к повышению кровяного давления), • дыхание, • участвует в регуляции пластического тонуса мышц (особенно при выполнении мелких движений пальцами рук) • Поддержание позы. • Гуморальная регуляция тонуса мышц обеспечивается черным веществом путем выделения его пигментными клетками вещества дофамина. • Дефицит дофамина проявляется как болезнь Паркинсона (дрожательный паралич).
 Заключение • Средний мозг регулирует тонус мышц, участвует в его распределении, что является необходимым условием для координированных движений. • Средний мозг обеспечивает регуляцию ряда вегетативных функций организма (жевание, глотание, давление крови, дыхание). • Средний мозг за счет сторожевых зрительных и слуховых рефлексов, усиления тонуса мышц-сгибателей подготавливает организм к ответу на внезапное раздражение. На уровне среднего мозга реализуются статические и статокинетические рефлексы. • Тонические рефлексы восстанавливают нарушенное равновесие, нарушенную при изменении положения позу. Они возникают при изменении положения тела и головы в пространства за счет возбуждения проприорецепторов, рецепторов вестибулярного аппарата и тактильных рецепторов кожи.
Заключение • Средний мозг регулирует тонус мышц, участвует в его распределении, что является необходимым условием для координированных движений. • Средний мозг обеспечивает регуляцию ряда вегетативных функций организма (жевание, глотание, давление крови, дыхание). • Средний мозг за счет сторожевых зрительных и слуховых рефлексов, усиления тонуса мышц-сгибателей подготавливает организм к ответу на внезапное раздражение. На уровне среднего мозга реализуются статические и статокинетические рефлексы. • Тонические рефлексы восстанавливают нарушенное равновесие, нарушенную при изменении положения позу. Они возникают при изменении положения тела и головы в пространства за счет возбуждения проприорецепторов, рецепторов вестибулярного аппарата и тактильных рецепторов кожи.
 ФИЗИОЛОГИЯ МОЗЖЕЧКА • Мозжечок — одна из интегративных структур головного мозга, принимающая участие в координации и регуляции произвольных и непроизвольных движений, в регуляции вегетативных и поведенческих функций.
ФИЗИОЛОГИЯ МОЗЖЕЧКА • Мозжечок — одна из интегративных структур головного мозга, принимающая участие в координации и регуляции произвольных и непроизвольных движений, в регуляции вегетативных и поведенческих функций.
 «Как скульптор избирательно удаляет резцом все лишнее из первоначально бесформенного камня, так и мозжечок, подавляя торможением лишние возбуждения, добивается четкой формы двигательной реакции» • Экклз, 1969
«Как скульптор избирательно удаляет резцом все лишнее из первоначально бесформенного камня, так и мозжечок, подавляя торможением лишние возбуждения, добивается четкой формы двигательной реакции» • Экклз, 1969
 • Мозжечок связан с другими отделами мозга с помощью афферентных и эфферентных путей. • Афферентные пути идут к нему из спинного, продолговатого мозга, варолиевого моста, четверохолмия. • От клеток Пуркинье коры мозжечка начинаются пути к ядрам мозжечка - зубчатому, пробковидному, шарообразному. От этих ядер эфферентные волокна идут к среднему мозгу. • Через красное ядро мозжечок соединяется эфферентными путями с корой больших полушарий.
• Мозжечок связан с другими отделами мозга с помощью афферентных и эфферентных путей. • Афферентные пути идут к нему из спинного, продолговатого мозга, варолиевого моста, четверохолмия. • От клеток Пуркинье коры мозжечка начинаются пути к ядрам мозжечка - зубчатому, пробковидному, шарообразному. От этих ядер эфферентные волокна идут к среднему мозгу. • Через красное ядро мозжечок соединяется эфферентными путями с корой больших полушарий.
 Повреждение мозжечка • Одномоментное удаление мозжечка приводит к гибели человека, в то же время, если удаляется часть мозжечка, это вмешательство, как правило, не смертельно. • Несмотря на то, что деятельность мозжечка имеет ближайшее отношение к осуществлению произвольных движений, повреждение мозжечка не влечет за собой двигательных и сенсорных параличей.
Повреждение мозжечка • Одномоментное удаление мозжечка приводит к гибели человека, в то же время, если удаляется часть мозжечка, это вмешательство, как правило, не смертельно. • Несмотря на то, что деятельность мозжечка имеет ближайшее отношение к осуществлению произвольных движений, повреждение мозжечка не влечет за собой двигательных и сенсорных параличей.
 Последствия удаления мозжечка и выпадения его функции • Триада Лючиани: астазия - потеря способности мышц к слитному сокращению, атония - отсутствие тонуса скелетных мышц, астения - быстрая утомляемость мышц • Триада Шарко : нистагм - непроизвольные ритмические движения головы и глаз интенционное дрожание (тремор) – размашистое дрожание, возникающее при совершении движения дизартрия (скандированная речь) - нарушение плавности речи • атаксия - нарушение точности движений.
Последствия удаления мозжечка и выпадения его функции • Триада Лючиани: астазия - потеря способности мышц к слитному сокращению, атония - отсутствие тонуса скелетных мышц, астения - быстрая утомляемость мышц • Триада Шарко : нистагм - непроизвольные ритмические движения головы и глаз интенционное дрожание (тремор) – размашистое дрожание, возникающее при совершении движения дизартрия (скандированная речь) - нарушение плавности речи • атаксия - нарушение точности движений.
 Симптомы удаления мозжечка • Безмозжечковая собака стоит на широко расставленных лапах, совершая непрерывные качательные движения (астазия). У нее нарушено правильное распределение тонуса мышц сгибателей и разгибателей (дистония). Движения плохо координированы размашисты, несоразмерны, резки. При ходьбе лапы забрасываются за среднюю линию (атаксия), чего не бывает у нормальных животных. Атаксия объясняется тем, что нарушается контроль движений. Выпадает и анализ сигналов от проприорецепторов мышц и сухожилий. Собака не может попасть мордой в миску с едой. Наклон головы вниз или в сторону вызывает сильное противоположное движение. Движения очень утомляют, животное, пройдя несколько шагов, ложится и отдыхает (астения). С течением времени двигательные расстройства у безмозжечковой собаки сглаживаются. Она самостоятельно ест, походка ее почти нормальна. Только предвзятое наблюдение выявляет некоторые нарушения (фаза компенсации).
Симптомы удаления мозжечка • Безмозжечковая собака стоит на широко расставленных лапах, совершая непрерывные качательные движения (астазия). У нее нарушено правильное распределение тонуса мышц сгибателей и разгибателей (дистония). Движения плохо координированы размашисты, несоразмерны, резки. При ходьбе лапы забрасываются за среднюю линию (атаксия), чего не бывает у нормальных животных. Атаксия объясняется тем, что нарушается контроль движений. Выпадает и анализ сигналов от проприорецепторов мышц и сухожилий. Собака не может попасть мордой в миску с едой. Наклон головы вниз или в сторону вызывает сильное противоположное движение. Движения очень утомляют, животное, пройдя несколько шагов, ложится и отдыхает (астения). С течением времени двигательные расстройства у безмозжечковой собаки сглаживаются. Она самостоятельно ест, походка ее почти нормальна. Только предвзятое наблюдение выявляет некоторые нарушения (фаза компенсации).
 Мозжечковая атаксия позы и походки при опухоли левой части мозжечка А Б В • А – качание направо при удержании вертикальной позы • Б – удержание позы на правой ноге • В- неустойчивая поза на левой ноге • Г- некоординированная походка Г
Мозжечковая атаксия позы и походки при опухоли левой части мозжечка А Б В • А – качание направо при удержании вертикальной позы • Б – удержание позы на правой ноге • В- неустойчивая поза на левой ноге • Г- некоординированная походка Г
 Функции мозжечка • Двигательные функции: • 1. Регуляция мышечного тонуса и позы. • 2. Коррекция медленных целенаправленных движений в ходе их выполнения, а также координация этих движений с рефлексам и положения тела. • 3. Контроль за правильным выполнением быстрых движений, осуществляемых корой. (Часть импульсов, возникших в коре мозга, достигает противоположного полушария мозжечка, принося информацию не о произведённом, а лишь о намечаемом к выполнению активном движении. Получив такую информацию, мозжечок моментально высылает импульсы, корригирующие произвольное движение главным образом путём погашения инерции и наиболее рациональной регуляции тонуса мышц агонистов и антагонистов. В результате обеспечивается чёткость и отточенность произвольных движений, устраняются какие-либо нецелесообразные компоненты. ) • Адаптационно-трофические функции.
Функции мозжечка • Двигательные функции: • 1. Регуляция мышечного тонуса и позы. • 2. Коррекция медленных целенаправленных движений в ходе их выполнения, а также координация этих движений с рефлексам и положения тела. • 3. Контроль за правильным выполнением быстрых движений, осуществляемых корой. (Часть импульсов, возникших в коре мозга, достигает противоположного полушария мозжечка, принося информацию не о произведённом, а лишь о намечаемом к выполнению активном движении. Получив такую информацию, мозжечок моментально высылает импульсы, корригирующие произвольное движение главным образом путём погашения инерции и наиболее рациональной регуляции тонуса мышц агонистов и антагонистов. В результате обеспечивается чёткость и отточенность произвольных движений, устраняются какие-либо нецелесообразные компоненты. ) • Адаптационно-трофические функции.
 Адаптационно-трофическая функция • В исследованиях Л. А. Орбели установлено, что мозжечок влияет на различные вегетативные функции • Эти влияния могут, быть возбуждающими и тормозящими. Наступающие под влиянием мозжечка изменения кровообращения, состава крови, пищеварения, дыхания направлены на трофическое обеспечение деятельности скелетных мышц. • Раздражение мозжечка вызывает целый ряд вегетативных рефлексов (расширение зрачков, повышение или снижение артериального давления, изменение частоты сердцебиений, дыхания). • При удалении мозжечка отмечены нарушения дыхания, сердечнососудистой деятельности, моторики и секреторной функции желудочно-кишечного тракта, снижение резистентности (устойчивости) организма к действию повреждающих факторов. • Мозжечок оказывает адаптационно-трофическое влияние на все отделы мозга через симпатический отдел нервной системы, регулируя в мозге обмен веществ и способствуя приспособлению ЦНС к изменяющимся условиям среды.
Адаптационно-трофическая функция • В исследованиях Л. А. Орбели установлено, что мозжечок влияет на различные вегетативные функции • Эти влияния могут, быть возбуждающими и тормозящими. Наступающие под влиянием мозжечка изменения кровообращения, состава крови, пищеварения, дыхания направлены на трофическое обеспечение деятельности скелетных мышц. • Раздражение мозжечка вызывает целый ряд вегетативных рефлексов (расширение зрачков, повышение или снижение артериального давления, изменение частоты сердцебиений, дыхания). • При удалении мозжечка отмечены нарушения дыхания, сердечнососудистой деятельности, моторики и секреторной функции желудочно-кишечного тракта, снижение резистентности (устойчивости) организма к действию повреждающих факторов. • Мозжечок оказывает адаптационно-трофическое влияние на все отделы мозга через симпатический отдел нервной системы, регулируя в мозге обмен веществ и способствуя приспособлению ЦНС к изменяющимся условиям среды.
 Физиология промежуточного мозга В состав промежуточного мозга входят: таламус, эпиталамус, метаталамус гипоталамус.
Физиология промежуточного мозга В состав промежуточного мозга входят: таламус, эпиталамус, метаталамус гипоталамус.
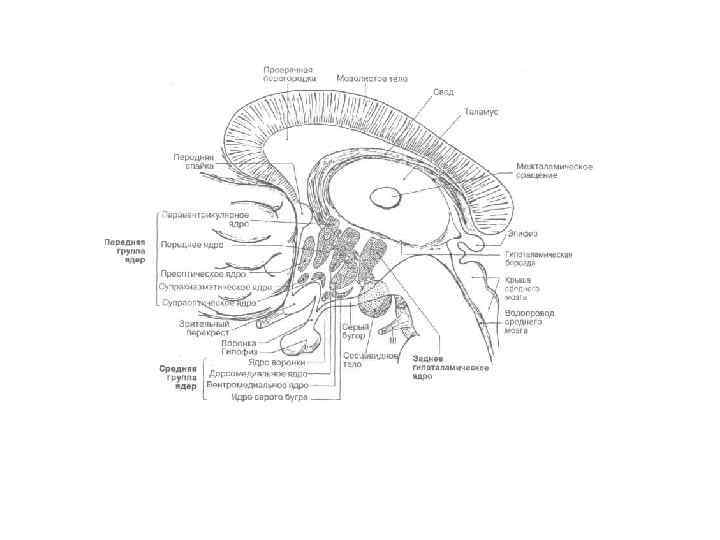
 • Таламус – наиболее крупное скопление серого вещества в промежуточном мозге. Таламус – коллектор всей афферентной информации, кроме обонятельных рецепторов, важнейший интегративный центр. • Топографически выделяют передние, средние, задние, медиальные и латеральные группы ядер. • По функции выделяют: • 1) специфические ядра: а) чувствительные, релейные ядра. Получают первичную информацию от различных рецепторов. Нервный импульс по таламокортикальному тракту идет в строго ограниченную зону коры головного мозга (первичные проекционные зоны), за счет этого возникают специфические ощущения. Ядра вентрабазального комплекса получают импульс от рецепторов кожи, проприорецепторов сухожилий, связок. Импульс направляется в сенсомоторную зону, происходит регуляция ориентировки тела в пространстве. Латеральные ядра переключают импульс от зрительных рецепторов в затылочную зрительную зону. Медиальные ядра реагируют на строго определенную длину звуковой волны и проводят импульс в височную зону; б) моторные ядра, участвующие в регуляции двигательной активности. Импульсы от мозжечка, базальных ядер идут в моторную зону, осуществляют взаимосвязь, согласованность, последовательность движений, пространственную ориентацию тела. • 2) неспецифические ядра. Это неспецифический путь передачи импульсов в кору головного мозга, изменяется частота биопотенциала (моделирующая функция); • 3) ассоциативные ядра. Первичный импульс идет от релейных ядер, перерабатывается (осуществляется интегративная функция), передается в ассоциативные зоны коры головного мозга;
• Таламус – наиболее крупное скопление серого вещества в промежуточном мозге. Таламус – коллектор всей афферентной информации, кроме обонятельных рецепторов, важнейший интегративный центр. • Топографически выделяют передние, средние, задние, медиальные и латеральные группы ядер. • По функции выделяют: • 1) специфические ядра: а) чувствительные, релейные ядра. Получают первичную информацию от различных рецепторов. Нервный импульс по таламокортикальному тракту идет в строго ограниченную зону коры головного мозга (первичные проекционные зоны), за счет этого возникают специфические ощущения. Ядра вентрабазального комплекса получают импульс от рецепторов кожи, проприорецепторов сухожилий, связок. Импульс направляется в сенсомоторную зону, происходит регуляция ориентировки тела в пространстве. Латеральные ядра переключают импульс от зрительных рецепторов в затылочную зрительную зону. Медиальные ядра реагируют на строго определенную длину звуковой волны и проводят импульс в височную зону; б) моторные ядра, участвующие в регуляции двигательной активности. Импульсы от мозжечка, базальных ядер идут в моторную зону, осуществляют взаимосвязь, согласованность, последовательность движений, пространственную ориентацию тела. • 2) неспецифические ядра. Это неспецифический путь передачи импульсов в кору головного мозга, изменяется частота биопотенциала (моделирующая функция); • 3) ассоциативные ядра. Первичный импульс идет от релейных ядер, перерабатывается (осуществляется интегративная функция), передается в ассоциативные зоны коры головного мозга;
 ядра таламуса делятся на сенсорные и несенсорные. Сенсорные специфические ядра таламуса переключают") Специфические (релейные) ядра таламуса делятся на сенсорные и несенсорные. Сенсорные специфические ядра таламуса переключают потоки афферентной импульсации в сенсорные зоны коры больших полушарий. В них также происходит перекодирование и обработка информации. К сенсорным ядрам относят вентробазальный комплекс, медиальное коленчатое тело и латеральное коленчатое тело. Вентробазальный комплекс является главным реле для переключения соматосенсорной афферентной системы, импульсы которой поступают по волокнам медиальной петли и других афферентных путей. В этом комплексе ядер переключается тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая чувствительность. В этих ядрах имеется топографическая проекция периферии. Импульсация из вентральнобазального комплекса проецируется в соматосенсорную кору постцентральной извилины. · Латеральное коленчатое тело способствует переключению зрительной импульсации в затылочную кору, где она используется в формировании зрительных ощущений. Кроме корковой проекции, часть зрительной импульсации направляется в верхние бугры четверохолмия. Эта информация используется для регуляции движения глаз и в зрительном ориентировочном рефлексе. · Медиальное коленчатое тело является реле для переключения слуховой импульсации в височную кору задней части сильвиевой борозды (извилины Гешля). В релейных ядрах обнаружены несколько типов нейронов. Нейроны, которые имеют длинный аксон, непосредственно обеспечивают переключение поступающей афферентной импульсации на нейроны сенсорной коры больших полушарий. Особенностью этого переключения является то, что информация без искажения поступает с входа на выход и идет далее в кору больших полушарий. Кроме этого имеются тормозные и возбуждающие вставочные нейроны, которые имеют короткие аксоны, и соответственно осуществляют: одни - возвратное торможение, другие возбуждение. Несенсорные специфические ядра таламуса переключают в кору несенсорную импульсацию. Различают переднее, вентролатеральное и переднецентральное ядра. · В передние ядра таламуса импульсация поступает из сосочковых тел гипоталамуса. Передние ядра рассматриваются как часть лимбической системы, так как нейроны этих ядер проецируются в лимбическую кору, откуда аксонные связи идут к гиппокампу и опять к гипоталамусу. В результате образуется так называемый круг возбуждения Пейпеца, где формируются эмоции. · Переднецентральное и вентролатеральное ядра участвуют в регуляции движения. В этих ядрах переключается импульсация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга в моторную и премоторную кору.
Специфические (релейные) ядра таламуса делятся на сенсорные и несенсорные. Сенсорные специфические ядра таламуса переключают потоки афферентной импульсации в сенсорные зоны коры больших полушарий. В них также происходит перекодирование и обработка информации. К сенсорным ядрам относят вентробазальный комплекс, медиальное коленчатое тело и латеральное коленчатое тело. Вентробазальный комплекс является главным реле для переключения соматосенсорной афферентной системы, импульсы которой поступают по волокнам медиальной петли и других афферентных путей. В этом комплексе ядер переключается тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая чувствительность. В этих ядрах имеется топографическая проекция периферии. Импульсация из вентральнобазального комплекса проецируется в соматосенсорную кору постцентральной извилины. · Латеральное коленчатое тело способствует переключению зрительной импульсации в затылочную кору, где она используется в формировании зрительных ощущений. Кроме корковой проекции, часть зрительной импульсации направляется в верхние бугры четверохолмия. Эта информация используется для регуляции движения глаз и в зрительном ориентировочном рефлексе. · Медиальное коленчатое тело является реле для переключения слуховой импульсации в височную кору задней части сильвиевой борозды (извилины Гешля). В релейных ядрах обнаружены несколько типов нейронов. Нейроны, которые имеют длинный аксон, непосредственно обеспечивают переключение поступающей афферентной импульсации на нейроны сенсорной коры больших полушарий. Особенностью этого переключения является то, что информация без искажения поступает с входа на выход и идет далее в кору больших полушарий. Кроме этого имеются тормозные и возбуждающие вставочные нейроны, которые имеют короткие аксоны, и соответственно осуществляют: одни - возвратное торможение, другие возбуждение. Несенсорные специфические ядра таламуса переключают в кору несенсорную импульсацию. Различают переднее, вентролатеральное и переднецентральное ядра. · В передние ядра таламуса импульсация поступает из сосочковых тел гипоталамуса. Передние ядра рассматриваются как часть лимбической системы, так как нейроны этих ядер проецируются в лимбическую кору, откуда аксонные связи идут к гиппокампу и опять к гипоталамусу. В результате образуется так называемый круг возбуждения Пейпеца, где формируются эмоции. · Переднецентральное и вентролатеральное ядра участвуют в регуляции движения. В этих ядрах переключается импульсация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга в моторную и премоторную кору.
 Неспецифические ядра составляют эволюционно более древнюю часть таламуса, включающую парные ретикулярные ядра и внутрипластинчатую ядерную группу. Неспецифические ядра имеют связи с моторными центрами ствола (красное ядро, черная субстанция), с мозжечком (ядро шатра и пробковое ядро), с базальными ганглиями, гиппокампом, с лобными долями полушарий. Благодаря этим связям неспецифические ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой и базальными ганглиями, с другой, объединяя их в единый функциональный комплекс. Главной функцией неспецифических ядер таламуса является тонкая настройка высшей нервной деятельности, за счет модулирующего влияния их на кору больших полушарий. Разрушение этих ядер ведет к нарушению тонкого регулирования поведения.
Неспецифические ядра составляют эволюционно более древнюю часть таламуса, включающую парные ретикулярные ядра и внутрипластинчатую ядерную группу. Неспецифические ядра имеют связи с моторными центрами ствола (красное ядро, черная субстанция), с мозжечком (ядро шатра и пробковое ядро), с базальными ганглиями, гиппокампом, с лобными долями полушарий. Благодаря этим связям неспецифические ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой и базальными ганглиями, с другой, объединяя их в единый функциональный комплекс. Главной функцией неспецифических ядер таламуса является тонкая настройка высшей нервной деятельности, за счет модулирующего влияния их на кору больших полушарий. Разрушение этих ядер ведет к нарушению тонкого регулирования поведения.
 Ассоциативные ядра таламуса принимают сигналы от других ядер таламуса. Эфферентные выходы от этих ядер направляются в ассоциативные поля коры больших полушарий. В свою очередь кора мозга посылает волокна к ассоциативным ядрам, регулируя их функцию. Главной функцией этих ядер является интегративная функция, которая выражается в объединении деятельности как таламических ядер, так и различных зон ассоциативной коры полушарий мозга. К ассоциативным ядрам относят подушку, медиодорсальное ядро и латеральное ядро. Связь ассоциативных ядер таламуса с другими образованиями ЦНС составляет основу ассоциативных систем таламуса. Ассоциативные системы таламуса 1. Таламопариетальная: от заднелатерального ядра и подушки к теменной и височной коре. Функции: центральный аппарат анализа и синтеза обстановочной афферентации, запуска ориентационных движений глаз и туловища; один из центральных аппаратов «схемы тела» и сенсорного контроля текущей двигательной активности; аппарат формирования полимодальных образов. 2. Таламофронтальная: от медиодорсального ядра к фронтальной и лимбической коре. Функции: корковый модулятор лимбической системы, программирование целенаправленных поведенческих актов на основе опыта и мотивации. Особенности работы ассоциативных систем таламуса: 1. Мультисенсорная конвергенция; 2. Пластическая перестройка при гетеромодальных сенсорных воздействиях: избирательное привыкание, сенситизация, экстраполяция. 3. Кратковременное хранение следов интеграции в виде реверберации импульсов.
Ассоциативные ядра таламуса принимают сигналы от других ядер таламуса. Эфферентные выходы от этих ядер направляются в ассоциативные поля коры больших полушарий. В свою очередь кора мозга посылает волокна к ассоциативным ядрам, регулируя их функцию. Главной функцией этих ядер является интегративная функция, которая выражается в объединении деятельности как таламических ядер, так и различных зон ассоциативной коры полушарий мозга. К ассоциативным ядрам относят подушку, медиодорсальное ядро и латеральное ядро. Связь ассоциативных ядер таламуса с другими образованиями ЦНС составляет основу ассоциативных систем таламуса. Ассоциативные системы таламуса 1. Таламопариетальная: от заднелатерального ядра и подушки к теменной и височной коре. Функции: центральный аппарат анализа и синтеза обстановочной афферентации, запуска ориентационных движений глаз и туловища; один из центральных аппаратов «схемы тела» и сенсорного контроля текущей двигательной активности; аппарат формирования полимодальных образов. 2. Таламофронтальная: от медиодорсального ядра к фронтальной и лимбической коре. Функции: корковый модулятор лимбической системы, программирование целенаправленных поведенческих актов на основе опыта и мотивации. Особенности работы ассоциативных систем таламуса: 1. Мультисенсорная конвергенция; 2. Пластическая перестройка при гетеромодальных сенсорных воздействиях: избирательное привыкание, сенситизация, экстраполяция. 3. Кратковременное хранение следов интеграции в виде реверберации импульсов.
 Функции таламуса • Таламус является коллектором и проводником всех видов чувствительности; • Участвует в регуляции функционального состояния организма; • Участвует в обеспечении двигательных реакций (сосание, жевание, глотание); • Таламус является центром болевой чувствительности, участвующим в формировании боли. • Участвует в формировании ощущений, влечений, различных эмоциональных состояний и др.
Функции таламуса • Таламус является коллектором и проводником всех видов чувствительности; • Участвует в регуляции функционального состояния организма; • Участвует в обеспечении двигательных реакций (сосание, жевание, глотание); • Таламус является центром болевой чувствительности, участвующим в формировании боли. • Участвует в формировании ощущений, влечений, различных эмоциональных состояний и др.
 Небольшая железа, расположенная между буграми четверохолмия головного мозга, входит в состав") Эпифиз (шишковидная железа) Небольшая железа, расположенная между буграми четверохолмия головного мозга, входит в состав эпиталамуса. Функции: • выделение гормонов; • регуляция суточного или циркадианного биологических ритмов, включающих периодичность сна и колебания температуры тела.
Эпифиз (шишковидная железа) Небольшая железа, расположенная между буграми четверохолмия головного мозга, входит в состав эпиталамуса. Функции: • выделение гормонов; • регуляция суточного или циркадианного биологических ритмов, включающих периодичность сна и колебания температуры тела.
 • Нейросекреция эпифиза зависит от освещенности. • Главным звеном в этой цепи является передний гипоталамус (супрахиазматическое ядро), которое получает прямой вход от волокон зрительного нерва. Далее от нейронов этого ядра образуется нисходящий путь к верхнему симпатическому узлу и затем в составе специального (пинеального) нерва поступает в эпифиз. На свету продукция мелатонина в эпифизе угнетается, тогда как в течение темной фазы суток она усиливается. Мелатонин влияет на функции многих отделов центральной нервной системы и некоторые поведенческие реакции. Например, у человека инъекция мелатонина вызывает сон.
• Нейросекреция эпифиза зависит от освещенности. • Главным звеном в этой цепи является передний гипоталамус (супрахиазматическое ядро), которое получает прямой вход от волокон зрительного нерва. Далее от нейронов этого ядра образуется нисходящий путь к верхнему симпатическому узлу и затем в составе специального (пинеального) нерва поступает в эпифиз. На свету продукция мелатонина в эпифизе угнетается, тогда как в течение темной фазы суток она усиливается. Мелатонин влияет на функции многих отделов центральной нервной системы и некоторые поведенческие реакции. Например, у человека инъекция мелатонина вызывает сон.
 Синтез мелатонина в зависимости от сезона
Синтез мелатонина в зависимости от сезона
 Функции мелатонина: • Регулирует деятельность эндокринной системы, кровяное давление, периодичность сна • Регулирует сезонную ритмику у многих животных • Замедляет процессы старения • Усиливает эффективность функционирования иммунной системы • Обладает антиоксидантными свойствами • Влияет на процессы адаптации при смене часовых поясов • Участвует в регуляции: • Кровяного давления • Функций пищеварительного тракта • Работы клеток головного мозга
Функции мелатонина: • Регулирует деятельность эндокринной системы, кровяное давление, периодичность сна • Регулирует сезонную ритмику у многих животных • Замедляет процессы старения • Усиливает эффективность функционирования иммунной системы • Обладает антиоксидантными свойствами • Влияет на процессы адаптации при смене часовых поясов • Участвует в регуляции: • Кровяного давления • Функций пищеварительного тракта • Работы клеток головного мозга
 Функции серотонина: играет важную роль в процессах свёртывания крови участвует в процессах аллергии и воспаления играет важную роль в регуляции моторики и секреции в желудочно -кишечном тракте • играет роль в паракринной регуляции сократимости матки, вовлечён в процесс овуляции • облегчает двигательную активность • играет важную роль в механизмах гипоталамической регуляции гормональной функции гипофиза. o • • •
Функции серотонина: играет важную роль в процессах свёртывания крови участвует в процессах аллергии и воспаления играет важную роль в регуляции моторики и секреции в желудочно -кишечном тракте • играет роль в паракринной регуляции сократимости матки, вовлечён в процесс овуляции • облегчает двигательную активность • играет важную роль в механизмах гипоталамической регуляции гормональной функции гипофиза. o • • •
 • Гипоталамус – это отдел промежуточного мозга, который играет роль поддержания гомеостатических констант организма, обеспечения интеграции функций вегетативной, эндокринной и соматической систем. • Он включает ряд анатомических образований: зрительный перекрест, зрительный тракт, сосцевидные тела, серый бугор с воронкой и гипофизом. • Нейроны гипоталамуса обладают особенностями: • 1) они чувствительны к составу омывающей их крови; • 2) отсутствие гематоэнцефалического барьера между кровью и нейронами; • 3) способность нейронов к нейросекреции пептидов и медиаторов. • 4) Ядра гипоталамуса имеют мощное кровоснабжение: на 1 мм² площади гипоталамуса приходится до 2600 капилляров (на той же площади V слоя предцентральной извилины (моторной коры) их 440, в гиппокампе 350, в бледном шаре 550, в затылочной доле коры большого мозга (зрительной коре) 900). • Гипоталамус содержит 32 пары ядер. В нем различают 3 группы скопления нервных клеток: переднюю, среднюю, заднюю группу.
• Гипоталамус – это отдел промежуточного мозга, который играет роль поддержания гомеостатических констант организма, обеспечения интеграции функций вегетативной, эндокринной и соматической систем. • Он включает ряд анатомических образований: зрительный перекрест, зрительный тракт, сосцевидные тела, серый бугор с воронкой и гипофизом. • Нейроны гипоталамуса обладают особенностями: • 1) они чувствительны к составу омывающей их крови; • 2) отсутствие гематоэнцефалического барьера между кровью и нейронами; • 3) способность нейронов к нейросекреции пептидов и медиаторов. • 4) Ядра гипоталамуса имеют мощное кровоснабжение: на 1 мм² площади гипоталамуса приходится до 2600 капилляров (на той же площади V слоя предцентральной извилины (моторной коры) их 440, в гиппокампе 350, в бледном шаре 550, в затылочной доле коры большого мозга (зрительной коре) 900). • Гипоталамус содержит 32 пары ядер. В нем различают 3 группы скопления нервных клеток: переднюю, среднюю, заднюю группу.
 Гипоталамус как высший центр интеграции вегетативных функций. Швейцарский физиолог В. Гесс в 1954 году показал, что в гипоталамусе имеются две функционально различные зоны: эрготропная, или симпатическая, расположенная в ядрах задней группы и области околоводопроводного серого вещества, и трофотропная, или парасимпатическая, локализованная в передних отделах гипоталамуса, преоптической области. Однако в настоящее время доказано, что имеет место мозаичное распределение и перекрытие участков, активизирующих симпатическую и парасимпатическую системы по всему гипоталамусу с некоторым преобладанием первых в задних отделах, а вторых – в передних. Вместе с тем на уровне гипоталамуса происходит не только интеграция деятельности различных вегетативных центров, но и включение их как компонента в более сложные физиологические системы различных форм биологического поведения, направленного на выживание организма, поддержание гомеостаза и сохранение вида.
Гипоталамус как высший центр интеграции вегетативных функций. Швейцарский физиолог В. Гесс в 1954 году показал, что в гипоталамусе имеются две функционально различные зоны: эрготропная, или симпатическая, расположенная в ядрах задней группы и области околоводопроводного серого вещества, и трофотропная, или парасимпатическая, локализованная в передних отделах гипоталамуса, преоптической области. Однако в настоящее время доказано, что имеет место мозаичное распределение и перекрытие участков, активизирующих симпатическую и парасимпатическую системы по всему гипоталамусу с некоторым преобладанием первых в задних отделах, а вторых – в передних. Вместе с тем на уровне гипоталамуса происходит не только интеграция деятельности различных вегетативных центров, но и включение их как компонента в более сложные физиологические системы различных форм биологического поведения, направленного на выживание организма, поддержание гомеостаза и сохранение вида.
 • Передняя группа включает супрахеазматическое ядро, супраоптическое и паравентрикулярное ядро. • Функции передней группы ядер гипоталамуса: • Возбуждение ядер передней группы приводит к реакции организма по парасимпатическому типу. Она направлена на восстановление и сохранение резервов организма. • Здесь находится центр сна (супрахеазматическое ядро). • Нейроны передней группы ядер гипоталамуса продуцируют гормоны окситоцин (паравентрикулярное ядро) и вазопрессин (супраоптическое ядро). Отростки этих нейронов идут к задней доле гипофиза.
• Передняя группа включает супрахеазматическое ядро, супраоптическое и паравентрикулярное ядро. • Функции передней группы ядер гипоталамуса: • Возбуждение ядер передней группы приводит к реакции организма по парасимпатическому типу. Она направлена на восстановление и сохранение резервов организма. • Здесь находится центр сна (супрахеазматическое ядро). • Нейроны передней группы ядер гипоталамуса продуцируют гормоны окситоцин (паравентрикулярное ядро) и вазопрессин (супраоптическое ядро). Отростки этих нейронов идут к задней доле гипофиза.
 • Средняя группа ядер гипоталамуса образована серобугорными ядрами, а также вентромедиальными и дорсомедиальными ядрами. • Функции средней группы ядер гипоталамуса: • Нейроны средней группы ядер гипоталамуса реагируют на изменение температуры крови, осмотического давления, количества и состава гормонов в крови. • Выработка релизинг-факторов. • Стимуляция ядер средней группы приводит к снижению влияний симпатической нервной системы.
• Средняя группа ядер гипоталамуса образована серобугорными ядрами, а также вентромедиальными и дорсомедиальными ядрами. • Функции средней группы ядер гипоталамуса: • Нейроны средней группы ядер гипоталамуса реагируют на изменение температуры крови, осмотического давления, количества и состава гормонов в крови. • Выработка релизинг-факторов. • Стимуляция ядер средней группы приводит к снижению влияний симпатической нервной системы.
 — класс пептидных гормонов гипоталамуса, общим свойством которых является") Рилизинг-гормоны • Рилизинг-факторы (либерины, статины) — класс пептидных гормонов гипоталамуса, общим свойством которых является реализация их эффектов через стимуляцию синтеза и секреции в кровь тропных гормонов передней доли гипофиза. К рилизинггормонам гипоталамуса относятся соматолиберин и соматостатин, пролактолиберин и пролактостатин, люлиберин, фоллиберин, кортиколиберин, тиролиберин, меланостатин.
Рилизинг-гормоны • Рилизинг-факторы (либерины, статины) — класс пептидных гормонов гипоталамуса, общим свойством которых является реализация их эффектов через стимуляцию синтеза и секреции в кровь тропных гормонов передней доли гипофиза. К рилизинггормонам гипоталамуса относятся соматолиберин и соматостатин, пролактолиберин и пролактостатин, люлиберин, фоллиберин, кортиколиберин, тиролиберин, меланостатин.
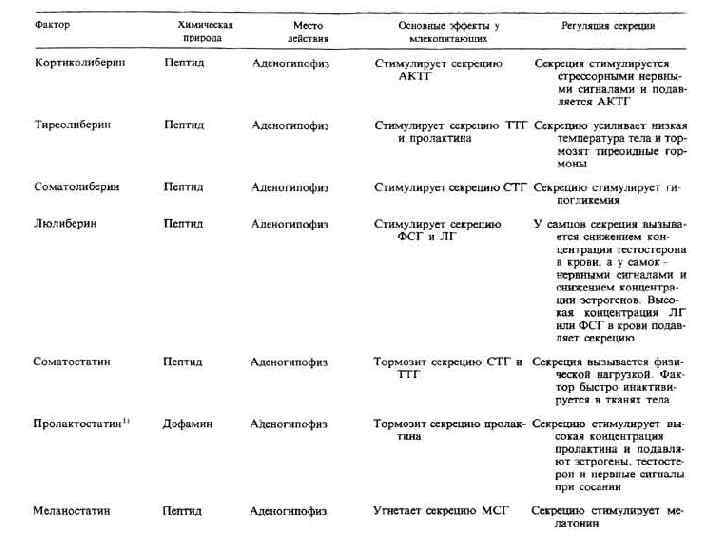
 • Задняя группа сформирована из заднего гипоталамического ядра, и сосцевидных ядер. • Функции задней группы ядер гипоталамуса: • Возбуждение ядер задней группы вызывает симпатический эффект работе органов, т. е. повышение АД; • Пищевое поведение наблюдается при стимуляции ядер заднего гипоталамуса. Повреждение латерального гипоталамуса ведет к полному отказу организма от пищи и воды, раздражение вентрамедиальных ядер гипоталамуса вызывает чрезмерное потребление пищи. • В заднем гипоталамусе обнаружены зоны, связанные с регуляцией полового поведения. • Гипоталамус участвует в регуляции агрессивного поведения. • Повреждение заднего гипоталамуса может вызвать летаргический сон.
• Задняя группа сформирована из заднего гипоталамического ядра, и сосцевидных ядер. • Функции задней группы ядер гипоталамуса: • Возбуждение ядер задней группы вызывает симпатический эффект работе органов, т. е. повышение АД; • Пищевое поведение наблюдается при стимуляции ядер заднего гипоталамуса. Повреждение латерального гипоталамуса ведет к полному отказу организма от пищи и воды, раздражение вентрамедиальных ядер гипоталамуса вызывает чрезмерное потребление пищи. • В заднем гипоталамусе обнаружены зоны, связанные с регуляцией полового поведения. • Гипоталамус участвует в регуляции агрессивного поведения. • Повреждение заднего гипоталамуса может вызвать летаргический сон.
 Центры гипоталамуса. • В гипоталамусе расположены важнейшие центры, организующие поддержание постоянства внутренней среды организма (гомеостаз) и приспособление внутренней среды к изменениям условий жизнедеятельности (гомеокинез).
Центры гипоталамуса. • В гипоталамусе расположены важнейшие центры, организующие поддержание постоянства внутренней среды организма (гомеостаз) и приспособление внутренней среды к изменениям условий жизнедеятельности (гомеокинез).
 • • • Гипоталамус – высший подкорковый интегративный центр вегетативной нервной системы. Регулирует гомеостаз организма. Гипоталамус оказывает действие на: 1) эндокринную систему, выделяя релизинг-факторы. Либерины способствуют секреции тропных гормонов передней доли гипофиза, статины тормозят ее; 2) терморегуляцию. Передние ядра – это центр теплоотдачи, где происходит регуляция процесса потоотделения, частоты дыхания и тонуса сосудов в ответ на повышение температуры окружающей среды. Задние ядра – центр теплопродукции и обеспечения сохранности тепла при понижении температуры; 3) жировой обмен. Раздражение латеральных (центра питания) ядер и вентромедиальных (центра насыщения) ядер ведет к ожирению, торможение – к кахексии; 4) углеводный обмен. Раздражение передних ядер ведет к гипогликемии, задних – к гипергликемии; 5) сердечно-сосудистую систему. Раздражение передних ядер оказывает тормозное влияние, задних – активирующее; 6) моторную и секреторную функции ЖКТ. Раздражение передних ядер повышает моторику и секреторную функцию ЖКТ, задних – тормозит; 7) половую функцию. Разрушение ядер ведет к нарушению овуляции, сперматогенеза, снижению половой функции; 8) поведенческие реакции. Раздражение передних ядер вызывает чувство радости, удовлетворения, эротические чувства. Раздражение задних ядер вызывает страх, чувство гнева, ярости.
• • • Гипоталамус – высший подкорковый интегративный центр вегетативной нервной системы. Регулирует гомеостаз организма. Гипоталамус оказывает действие на: 1) эндокринную систему, выделяя релизинг-факторы. Либерины способствуют секреции тропных гормонов передней доли гипофиза, статины тормозят ее; 2) терморегуляцию. Передние ядра – это центр теплоотдачи, где происходит регуляция процесса потоотделения, частоты дыхания и тонуса сосудов в ответ на повышение температуры окружающей среды. Задние ядра – центр теплопродукции и обеспечения сохранности тепла при понижении температуры; 3) жировой обмен. Раздражение латеральных (центра питания) ядер и вентромедиальных (центра насыщения) ядер ведет к ожирению, торможение – к кахексии; 4) углеводный обмен. Раздражение передних ядер ведет к гипогликемии, задних – к гипергликемии; 5) сердечно-сосудистую систему. Раздражение передних ядер оказывает тормозное влияние, задних – активирующее; 6) моторную и секреторную функции ЖКТ. Раздражение передних ядер повышает моторику и секреторную функцию ЖКТ, задних – тормозит; 7) половую функцию. Разрушение ядер ведет к нарушению овуляции, сперматогенеза, снижению половой функции; 8) поведенческие реакции. Раздражение передних ядер вызывает чувство радости, удовлетворения, эротические чувства. Раздражение задних ядер вызывает страх, чувство гнева, ярости.
 Центры теплорегуляции • • • Центры теплорегуляции включают в себя структуры, регулирующие теплообразование, и структуры, регулирующие теплоотдачу. Центр теплоотдачи локализован в передней и преоптической зонах гипоталамуса и включает в себя паравентрикулярные, супраоптические и медиальные преоптические ядра. Раздражение этих структур вызывает увеличение теплоотдачи в результате расширения сосудов кожи и повышения температуры ее поверхности, увеличения отделения и испарения пота, появления тепловой одышки. Разрушение центра теплоотдачи приводит к неспособности организма выдержать тепловую нагрузку. Центр теплопродукции локализован в заднем гипоталамусе и включает в себя медиальные, латеральные и промежуточные сосцевидные ядра. Его раздражение вызывает повышение температуры тела в результате усиления окислительных процессов, тонуса мышц и появления мышечной дрожи, сужения сосудов кожи. Разрушение этих ядер ведет к потере способности поддерживать температуру тела при охлаждении организма.
Центры теплорегуляции • • • Центры теплорегуляции включают в себя структуры, регулирующие теплообразование, и структуры, регулирующие теплоотдачу. Центр теплоотдачи локализован в передней и преоптической зонах гипоталамуса и включает в себя паравентрикулярные, супраоптические и медиальные преоптические ядра. Раздражение этих структур вызывает увеличение теплоотдачи в результате расширения сосудов кожи и повышения температуры ее поверхности, увеличения отделения и испарения пота, появления тепловой одышки. Разрушение центра теплоотдачи приводит к неспособности организма выдержать тепловую нагрузку. Центр теплопродукции локализован в заднем гипоталамусе и включает в себя медиальные, латеральные и промежуточные сосцевидные ядра. Его раздражение вызывает повышение температуры тела в результате усиления окислительных процессов, тонуса мышц и появления мышечной дрожи, сужения сосудов кожи. Разрушение этих ядер ведет к потере способности поддерживать температуру тела при охлаждении организма.
 Виды нейронов терморегуляторного центра гипоталамуса: • • • 1. Термочувствительные нейроны – термодетекторы – расположены в основном в центре теплоотдачи (от 30 до 70%). В центре теплопродукции их около 10%. Среди этих нейронов есть нейроны тепловые и холодовые. Холодовых нейронов в 3 раза меньше, чем тепловых. Чувствительность термодетекторов: от 0, 20 до 10 С. 2. Интегрирующие нейроны расположены преимущественно в центре теплоотдачи. Основной функцией этих нейронов является суммация температурных сигналов от терморецепторов гипоталамуса, среднего, продолговатого и спинного мозга, кожи, внутренних органов. Переработанная в них информация о тепловом состоянии организма поступает на следующий вид нейронов. 3. Эфферентные терморегуляторные нейроны расположены в основном в заднем гипоталамусе. Одни из них регулируют интенсивность механизмов теплопродукции, другие – механизмов теплоотдачи. 4. Тормозные интернейроны осуществляют реципрокные отношения между эфферентными нейронами теплоотдачи и теплопродукции. 5. Нейроны, воспринимающие нетемпературные стимулы. Эти нейроны осуществляют распознавание эндогенных и экзогенных влияний пирогенов. К экзогенным пирогенам относят вирусные и бактериальные интоксикации. К эндогенным пирогенам относят интерлейкин I, простагландин Е 1, фактор некроза опухоли.
Виды нейронов терморегуляторного центра гипоталамуса: • • • 1. Термочувствительные нейроны – термодетекторы – расположены в основном в центре теплоотдачи (от 30 до 70%). В центре теплопродукции их около 10%. Среди этих нейронов есть нейроны тепловые и холодовые. Холодовых нейронов в 3 раза меньше, чем тепловых. Чувствительность термодетекторов: от 0, 20 до 10 С. 2. Интегрирующие нейроны расположены преимущественно в центре теплоотдачи. Основной функцией этих нейронов является суммация температурных сигналов от терморецепторов гипоталамуса, среднего, продолговатого и спинного мозга, кожи, внутренних органов. Переработанная в них информация о тепловом состоянии организма поступает на следующий вид нейронов. 3. Эфферентные терморегуляторные нейроны расположены в основном в заднем гипоталамусе. Одни из них регулируют интенсивность механизмов теплопродукции, другие – механизмов теплоотдачи. 4. Тормозные интернейроны осуществляют реципрокные отношения между эфферентными нейронами теплоотдачи и теплопродукции. 5. Нейроны, воспринимающие нетемпературные стимулы. Эти нейроны осуществляют распознавание эндогенных и экзогенных влияний пирогенов. К экзогенным пирогенам относят вирусные и бактериальные интоксикации. К эндогенным пирогенам относят интерлейкин I, простагландин Е 1, фактор некроза опухоли.
 Центр пищевого поведения. • • В практической медицине давно было показано, что патологические процессы (опухоль, кровоизлияние, воспаление) в области гипоталамуса вызывают резкие изменения пищевого поведения. Экспериментальные исследования Б. Анада и Дж. Дробека (1951) показали, что разрушение небольшой зоны в области латерального гипоталамуса приводило к отказу от пищи, воды и гибели животного от истощения. Электрическая стимуляция этой зоны активизировала пищевое поведение и приводила к ожирению. Так был открыт центр голода и насыщения.
Центр пищевого поведения. • • В практической медицине давно было показано, что патологические процессы (опухоль, кровоизлияние, воспаление) в области гипоталамуса вызывают резкие изменения пищевого поведения. Экспериментальные исследования Б. Анада и Дж. Дробека (1951) показали, что разрушение небольшой зоны в области латерального гипоталамуса приводило к отказу от пищи, воды и гибели животного от истощения. Электрическая стимуляция этой зоны активизировала пищевое поведение и приводила к ожирению. Так был открыт центр голода и насыщения.
 Центр голода и насыщения • • • Центр голода и насыщения включает в себя две области гипоталамуса: центр голода расположен в латеральной зоне гипоталамуса, а центр насыщения располагается в зоне вентральных ядер. Нейроны центра голода возбуждаются при снижении содержания питательных веществ (глюкозы, аминокислот, жирных и органических кислот) в крови. Нейроны центра насыщения возбуждаются при достаточном повышении содержания в крови питательных веществ. Кроме этого на активность центров влияет уровень гормонов (инсулина, гастрина, адреналина и т. д. ). Нарушение работы центра голода приводит к отказу от приема пищи – афагии, и от приема воды – адипсии. Разрушение вентромедиальных ядер приводит и гиперфагии.
Центр голода и насыщения • • • Центр голода и насыщения включает в себя две области гипоталамуса: центр голода расположен в латеральной зоне гипоталамуса, а центр насыщения располагается в зоне вентральных ядер. Нейроны центра голода возбуждаются при снижении содержания питательных веществ (глюкозы, аминокислот, жирных и органических кислот) в крови. Нейроны центра насыщения возбуждаются при достаточном повышении содержания в крови питательных веществ. Кроме этого на активность центров влияет уровень гормонов (инсулина, гастрина, адреналина и т. д. ). Нарушение работы центра голода приводит к отказу от приема пищи – афагии, и от приема воды – адипсии. Разрушение вентромедиальных ядер приводит и гиперфагии.
 Центр жажды • • Центр жажды располагается в переднем гипотааламусе сзади и наружно от супраоптических ядер. Стимуляция этой области гипоталамуса ведет к резкому увеличению потребления воды, а разрушение ее – к адипсии – отказу от воды. Возбуждение нейронов центра связано с их чувствительностью к повышению осмотического давления крови, а также чувствительностью к концентрации антидиуретического (АДГ) гормона в крови. Центр жажды гипоталамуса был открыт в 1958 году Б. Андерсоном.
Центр жажды • • Центр жажды располагается в переднем гипотааламусе сзади и наружно от супраоптических ядер. Стимуляция этой области гипоталамуса ведет к резкому увеличению потребления воды, а разрушение ее – к адипсии – отказу от воды. Возбуждение нейронов центра связано с их чувствительностью к повышению осмотического давления крови, а также чувствительностью к концентрации антидиуретического (АДГ) гормона в крови. Центр жажды гипоталамуса был открыт в 1958 году Б. Андерсоном.
 Центры полового поведения • • Половая дифференцировка гипоталамуса по женскому или мужскому типу формируется в последние дни пренатального онтогенеза и первые дни после рождения. В гипоталамусе мужского организма функционирует «тонический» половой центр, расположенный в средней области (аркуатные и вентромедиальные ядра). Нейроны этих ядер выделяют гонадолиберины. Тем самым они оказывают стимулирующее влияние на секрецию лютеинезирующего (ЛГ) и фолликулостимулирющего (ФСГ) гормонов гипофиза. В свою очередь, концентрация половых гормонов крови сильно влияет на возбудимость секреторных центров гипоталамуса: повышение их концентрации тормозит секрецию гонадолиберинов и наоборот. В женском организме кроме «тонического» функционирует также «циклический» центр, расположенный в преоптической и передней областях гипоталамуса (особенно в супрахиазматическом и медиальном преоптическом ядрах). «Циклический» центр осуществляет регуляцию менструального цикла: предовуляторный пик секреции лютеинизирующего гормона В регуляции полового поведения участвуют зоны заднего гипоталамуса и медиальный пучок переднего мозга. Эти структуры названы «центром удовольствия» , так как при их раздражении формируется чувство радости и удовольствия, сопровождающееся эротическими переживаниями. Возбудимость сексуальных центров зависит от корковых и лимбических влияний, рефлекторной стимуляции с эрогенных генитальных и экстрагенитальных зон, уровня половых гормонов.
Центры полового поведения • • Половая дифференцировка гипоталамуса по женскому или мужскому типу формируется в последние дни пренатального онтогенеза и первые дни после рождения. В гипоталамусе мужского организма функционирует «тонический» половой центр, расположенный в средней области (аркуатные и вентромедиальные ядра). Нейроны этих ядер выделяют гонадолиберины. Тем самым они оказывают стимулирующее влияние на секрецию лютеинезирующего (ЛГ) и фолликулостимулирющего (ФСГ) гормонов гипофиза. В свою очередь, концентрация половых гормонов крови сильно влияет на возбудимость секреторных центров гипоталамуса: повышение их концентрации тормозит секрецию гонадолиберинов и наоборот. В женском организме кроме «тонического» функционирует также «циклический» центр, расположенный в преоптической и передней областях гипоталамуса (особенно в супрахиазматическом и медиальном преоптическом ядрах). «Циклический» центр осуществляет регуляцию менструального цикла: предовуляторный пик секреции лютеинизирующего гормона В регуляции полового поведения участвуют зоны заднего гипоталамуса и медиальный пучок переднего мозга. Эти структуры названы «центром удовольствия» , так как при их раздражении формируется чувство радости и удовольствия, сопровождающееся эротическими переживаниями. Возбудимость сексуальных центров зависит от корковых и лимбических влияний, рефлекторной стимуляции с эрогенных генитальных и экстрагенитальных зон, уровня половых гормонов.
 Центр агрессивно-оборонительного поведения. Агрессивные и оборонительные реакции были получены при раздражении следующих зон гипоталамуса: передней и задней, вентромедиальной и латеральной. При осуществлении агрессивно-оборонительных реакций гипоталамус взаимодействует с центральным серым веществом среднего мозга. В этой структуре были обнаружены «нейроны агрессии» , которые через гипоталамус запускают реакции агрессии и не возбуждаются при других реакциях. При повреждении этих нейронов резко увеличивается порог электрического раздражения гипоталамуса для вызова реакции ярости. На агрессивное поведение стимулирующее влияние оказывают андрогены, особенно тестостерон. В формировании «осмысленного» агрессивного поведения играют большую роль миндалевидный комплекс височной доли и орбитальная зона лобной коры.
Центр агрессивно-оборонительного поведения. Агрессивные и оборонительные реакции были получены при раздражении следующих зон гипоталамуса: передней и задней, вентромедиальной и латеральной. При осуществлении агрессивно-оборонительных реакций гипоталамус взаимодействует с центральным серым веществом среднего мозга. В этой структуре были обнаружены «нейроны агрессии» , которые через гипоталамус запускают реакции агрессии и не возбуждаются при других реакциях. При повреждении этих нейронов резко увеличивается порог электрического раздражения гипоталамуса для вызова реакции ярости. На агрессивное поведение стимулирующее влияние оказывают андрогены, особенно тестостерон. В формировании «осмысленного» агрессивного поведения играют большую роль миндалевидный комплекс височной доли и орбитальная зона лобной коры.
 Центры регуляции цикла «сонбодрствование» . • • • Центр сна расположен в переднем гипоталамусе, а центр бодрствования – в заднем. Электрическое раздражение преоптической области (тормозная система) гипоталамуса вызывало синхронизацию ЭЭГ и поведенческий сон. В это время активирующая ретикулярная формация ствола реципрокно заторможена. Стимуляция заднего гипоталамуса вызывала десинхронизацию ЭЭГ и пробуждение. Пробуждающий эффект заднего гипоталамуса увеличивается при возбуждении активирующей ретикулярной формации ствола мозга.
Центры регуляции цикла «сонбодрствование» . • • • Центр сна расположен в переднем гипоталамусе, а центр бодрствования – в заднем. Электрическое раздражение преоптической области (тормозная система) гипоталамуса вызывало синхронизацию ЭЭГ и поведенческий сон. В это время активирующая ретикулярная формация ствола реципрокно заторможена. Стимуляция заднего гипоталамуса вызывала десинхронизацию ЭЭГ и пробуждение. Пробуждающий эффект заднего гипоталамуса увеличивается при возбуждении активирующей ретикулярной формации ствола мозга.
 Центр циркадианных ритмов. Роль гипоталамуса не ограничивается только формированием механизмов сна и бодрствования. Пейсмекерами околосуточного ритма являются супрахиазматическое и вентромедиальное ядра. Регулируя околосуточные биоритмы, гипоталамус взаимодействует с эпифизом, с которым он имеет прямые связи.
Центр циркадианных ритмов. Роль гипоталамуса не ограничивается только формированием механизмов сна и бодрствования. Пейсмекерами околосуточного ритма являются супрахиазматическое и вентромедиальное ядра. Регулируя околосуточные биоритмы, гипоталамус взаимодействует с эпифизом, с которым он имеет прямые связи.
 Связь гипоталамуса с гипофизом Особая роль гипоталамуса в регуляции вегетативных функций организма определяется тем, что, наряду с регуляцией вегетативной системы, он регулирует работу гипофиза, а через него и всей эндокринной системы. Между гипоталамусом и гипофизом существуют тесные структурно-функциональные связи, объединяющие их в гипоталамо-гипофизарную систему. Задняя доля гипофиза (нейрогипофиз) связана с супраоптическим ядром гипоталамуса прямыми нервными связями: супрастино-гипофизарный тракт представляет собой аксоны клеток супраоптического ядра. Нейросекрет, вырабатываемый нейронами гипоталамуса, по аксонам транспортируется в заднюю долю гипофиза и выделяется в кровь на окончаниях аксонов при их возбуждении, как медиатор. Основными гормонами нейрогипофиза являются антидиуретический гормон (АДГ) – вазопрессин, регулирующий водный обмен, и окситоцин, регулирующий мускулатуру матки. С передней долей гипофиза (аденогипофизом) гипоталамус связан через общую воротную сосудистую систему. Попадающие в кровеносные сосуды продукты нейросекреции гипоталамуса с током крови – нейрогуморальным путем – регулируют работу аденогипофиза, который вырабатывает тропные гормоны, контролирующие работу других эндокринных желез. К основным тропным гормонам относятся: адренокортикотропный гормон (АКТГ), стимулирующий выделение гормонов коры надпочечников; тиреотропный гормон (ТТГ) стимулирует работу щитовидной железы; фолликулостимулирующий гормон (ФСГ) и лютеинезирующий гормон (ЛГ) регулируют работу половых желез; гормон роста – соматотропный гормон (СТГ); гормон пролактин, стимулирующий работу молочных желез. Регулируя работу гипофиза, гипоталамус может эффективно контролировать работу всех основных звеньев эндокринной системы и ее влияние на вегетативные функции. Так, например, путь гипоталамус – гипофиз – надпочечники является важнейшим для изменения вегетативных функций и состояния организма при стрессе.
Связь гипоталамуса с гипофизом Особая роль гипоталамуса в регуляции вегетативных функций организма определяется тем, что, наряду с регуляцией вегетативной системы, он регулирует работу гипофиза, а через него и всей эндокринной системы. Между гипоталамусом и гипофизом существуют тесные структурно-функциональные связи, объединяющие их в гипоталамо-гипофизарную систему. Задняя доля гипофиза (нейрогипофиз) связана с супраоптическим ядром гипоталамуса прямыми нервными связями: супрастино-гипофизарный тракт представляет собой аксоны клеток супраоптического ядра. Нейросекрет, вырабатываемый нейронами гипоталамуса, по аксонам транспортируется в заднюю долю гипофиза и выделяется в кровь на окончаниях аксонов при их возбуждении, как медиатор. Основными гормонами нейрогипофиза являются антидиуретический гормон (АДГ) – вазопрессин, регулирующий водный обмен, и окситоцин, регулирующий мускулатуру матки. С передней долей гипофиза (аденогипофизом) гипоталамус связан через общую воротную сосудистую систему. Попадающие в кровеносные сосуды продукты нейросекреции гипоталамуса с током крови – нейрогуморальным путем – регулируют работу аденогипофиза, который вырабатывает тропные гормоны, контролирующие работу других эндокринных желез. К основным тропным гормонам относятся: адренокортикотропный гормон (АКТГ), стимулирующий выделение гормонов коры надпочечников; тиреотропный гормон (ТТГ) стимулирует работу щитовидной железы; фолликулостимулирующий гормон (ФСГ) и лютеинезирующий гормон (ЛГ) регулируют работу половых желез; гормон роста – соматотропный гормон (СТГ); гормон пролактин, стимулирующий работу молочных желез. Регулируя работу гипофиза, гипоталамус может эффективно контролировать работу всех основных звеньев эндокринной системы и ее влияние на вегетативные функции. Так, например, путь гипоталамус – гипофиз – надпочечники является важнейшим для изменения вегетативных функций и состояния организма при стрессе.
 Сосцевидные тела • Сосцевидные тела— два маленьких округлых парных образования заднего гипоталамуса. Под поверхностным слоем белого вещества внутри каждого из сосцевидных тел находится по два серых ядра (медиальное и латеральное). Функции сосцевидных тел: • Относятся к лимбической системе. • Являются подкорковым центром обоняния. • Наряду с передним и медиальним ядрами таламуса, принимают участие в формирования памяти. • Также считается, что сосцевидные тела вовлечены в контроль эмоционального и сексуального поведения. • Повреждение сосцевидных тел из-за дефицита витамина В 1 может вызвать синдром Гайе — Вернике (болезнь Вернике). Классическая триада симптомов — офтальмоплегия (паралич мышц глаза), атаксия, спутанность сознания.
Сосцевидные тела • Сосцевидные тела— два маленьких округлых парных образования заднего гипоталамуса. Под поверхностным слоем белого вещества внутри каждого из сосцевидных тел находится по два серых ядра (медиальное и латеральное). Функции сосцевидных тел: • Относятся к лимбической системе. • Являются подкорковым центром обоняния. • Наряду с передним и медиальним ядрами таламуса, принимают участие в формирования памяти. • Также считается, что сосцевидные тела вовлечены в контроль эмоционального и сексуального поведения. • Повреждение сосцевидных тел из-за дефицита витамина В 1 может вызвать синдром Гайе — Вернике (болезнь Вернике). Классическая триада симптомов — офтальмоплегия (паралич мышц глаза), атаксия, спутанность сознания.
 НЕЙРОФИЗИОЛОГИЯ КОНЕЧНОГО МОЗГА. • • • Серое вещество конечного мозга представлено базальными ганглиями и корой больших полушарий. Базальные (подкорковые) ядра головного мозга располагаются под белым веществом внутри конечного мозга, преимущественно в лобных долях. К базальным ядрам относят хвостатое ядро, бледный шар, скорлупу, ограду и миндалину. Базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер.
НЕЙРОФИЗИОЛОГИЯ КОНЕЧНОГО МОЗГА. • • • Серое вещество конечного мозга представлено базальными ганглиями и корой больших полушарий. Базальные (подкорковые) ядра головного мозга располагаются под белым веществом внутри конечного мозга, преимущественно в лобных долях. К базальным ядрам относят хвостатое ядро, бледный шар, скорлупу, ограду и миндалину. Базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер.
. Отношения базальных ганглиев к желудочковой системе (Б). 1 - бледный") Положение базальных ганглиев (А). Отношения базальных ганглиев к желудочковой системе (Б). 1 - бледный шар; 2 - таламус; 3 скорлупа; 4 - хвостатое ядро; 5 - миндалевидное тело; 6 - головка хвостатого ядра; 7 - субталамическое ядро; 8 - хвостатого ядра; 9 - боковой желудочек.
Положение базальных ганглиев (А). Отношения базальных ганглиев к желудочковой системе (Б). 1 - бледный шар; 2 - таламус; 3 скорлупа; 4 - хвостатое ядро; 5 - миндалевидное тело; 6 - головка хвостатого ядра; 7 - субталамическое ядро; 8 - хвостатого ядра; 9 - боковой желудочек.
 Cтриопаллидарная система является центром экстрапирамидной системы. Ее основная функция – регуляция произвольных двигательных реакций. При ее участии создаются: · оптимальная для намеченного действия поза; · оптимальное соотношение тонуса между мышцами антагонистами и синергистами; · плавность и соразмерность движений во времени и пространстве.
Cтриопаллидарная система является центром экстрапирамидной системы. Ее основная функция – регуляция произвольных двигательных реакций. При ее участии создаются: · оптимальная для намеченного действия поза; · оптимальное соотношение тонуса между мышцами антагонистами и синергистами; · плавность и соразмерность движений во времени и пространстве.
 Поражение бледного шара. Поражение бледного шара характеризуется гипертоническигипокинетическим синдромом. Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи. При повреждении бледного шара наблюдается также миоклония — быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица. Поражение дает выпадение из произвольного двигательного акта сопутствующих неосновных компонентов. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно. Эти нарушения фиксируются при болезни Паркинсона (при повреждении нейронов в полосатом теле перестаёт вырабатываться дофамин).
Поражение бледного шара. Поражение бледного шара характеризуется гипертоническигипокинетическим синдромом. Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи. При повреждении бледного шара наблюдается также миоклония — быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица. Поражение дает выпадение из произвольного двигательного акта сопутствующих неосновных компонентов. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно. Эти нарушения фиксируются при болезни Паркинсона (при повреждении нейронов в полосатом теле перестаёт вырабатываться дофамин).
 Поражение бледного шара. Акинетико-ригидный синдром проявляется акинезом, пластической гипертонией мышц, дрожанием типа «катания пилюль» , флексорной позой головы и туловища, маскообразностью лица, немодулированной речью, отсутствием эмоциональных выражений. Начало произвольных движений затруднено; исчезают вспомогательные движения при вставании и ходьбе (ахейрокинез); походка мелкими шагами с пропульсией; повышаются постуральные рефлексы.
Поражение бледного шара. Акинетико-ригидный синдром проявляется акинезом, пластической гипертонией мышц, дрожанием типа «катания пилюль» , флексорной позой головы и туловища, маскообразностью лица, немодулированной речью, отсутствием эмоциональных выражений. Начало произвольных движений затруднено; исчезают вспомогательные движения при вставании и ходьбе (ахейрокинез); походка мелкими шагами с пропульсией; повышаются постуральные рефлексы.
 Поражение хвостатого ядра и скорлупы приводит к гипотоническигиперкинетическому синдрому. У больного наблюдаются разнообразные непроизвольные движения — гиперкинезы. Мышечный тонус снижается. Гиперкинезия (хорея) – сильные неправильные движения, совершающиеся без всякого порядка и последовательности, которые захватывают всю мускулатуру - «пляска святого Витта» . Причина: выпадение тормозного влияния стриарной системы на паллидарную систему.
Поражение хвостатого ядра и скорлупы приводит к гипотоническигиперкинетическому синдрому. У больного наблюдаются разнообразные непроизвольные движения — гиперкинезы. Мышечный тонус снижается. Гиперкинезия (хорея) – сильные неправильные движения, совершающиеся без всякого порядка и последовательности, которые захватывают всю мускулатуру - «пляска святого Витта» . Причина: выпадение тормозного влияния стриарной системы на паллидарную систему.
 Поражение хвостатого ядра и скорлупы • Хореический гиперкинез. При хорее бывают быстрые неритмические нелокализованные избыточные движения в мышцах лица, языка, конечностей, туловища, шеи • Атетотический гиперкинез пальцев и кисти.
Поражение хвостатого ядра и скорлупы • Хореический гиперкинез. При хорее бывают быстрые неритмические нелокализованные избыточные движения в мышцах лица, языка, конечностей, туловища, шеи • Атетотический гиперкинез пальцев и кисти.
 Атетоз
Атетоз
 Поражение хвостатого ядра и скорлупы • Торсионный спазм - подкорковый гиперкинез, характеризующийся тоническими спазмами различных мышечных групп преимущественно туловища, приводящий к медленным беспорядочным насильственным движениям. Движения туловища вычурные, нередко вращательные вокруг продольной оси (штопорообразные).
Поражение хвостатого ядра и скорлупы • Торсионный спазм - подкорковый гиперкинез, характеризующийся тоническими спазмами различных мышечных групп преимущественно туловища, приводящий к медленным беспорядочным насильственным движениям. Движения туловища вычурные, нередко вращательные вокруг продольной оси (штопорообразные).
 • Кора- продукт длительного") Высшим отделом ЦНС является кора большого мозга (кора больших полушарий) • Кора- продукт длительного эволюционного развития, в процессе котоpoгo сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. • С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано с совершенствованием и взаимодействием различных воспринимающих и двигательных систем. • У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. • Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты).
Высшим отделом ЦНС является кора большого мозга (кора больших полушарий) • Кора- продукт длительного эволюционного развития, в процессе котоpoгo сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. • С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано с совершенствованием и взаимодействием различных воспринимающих и двигательных систем. • У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. • Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты).
 включает в себя неструктурированную кору вокруг переднего продырявленного вещества:") • ДРЕВНЯЯ КОРА (paleocortecx) включает в себя неструктурированную кору вокруг переднего продырявленного вещества: околоконечную извилину, подмозолистое поле (расположено на внутренней стороне полушарий под коленом и клювом мозолистого тела). 0, 6% от всей коры. • СТАРАЯ КОРА (archicortex), двух- трехслойна, расположена в гиппокампе и зубчатой извилине. 2, 2% от объема коры. • СРЕДНЯЯ КОРА (mesocortex) занимает нижний отдел островковой доли, парагиппокампальную извилину и нижнюю лимбическую область, ее кора дифференцирована не полностью. 4 -5 слоев клеток 1, 6 %. • НОВАЯ КОРА (neocortex) составляет 95, 6 % от всей поверхности полушарий. По морфологическим особенностям в ней выделяют 6 основных слоев, однако в различных областях коры количество слоев варьирует.
• ДРЕВНЯЯ КОРА (paleocortecx) включает в себя неструктурированную кору вокруг переднего продырявленного вещества: околоконечную извилину, подмозолистое поле (расположено на внутренней стороне полушарий под коленом и клювом мозолистого тела). 0, 6% от всей коры. • СТАРАЯ КОРА (archicortex), двух- трехслойна, расположена в гиппокампе и зубчатой извилине. 2, 2% от объема коры. • СРЕДНЯЯ КОРА (mesocortex) занимает нижний отдел островковой доли, парагиппокампальную извилину и нижнюю лимбическую область, ее кора дифференцирована не полностью. 4 -5 слоев клеток 1, 6 %. • НОВАЯ КОРА (neocortex) составляет 95, 6 % от всей поверхности полушарий. По морфологическим особенностям в ней выделяют 6 основных слоев, однако в различных областях коры количество слоев варьирует.
 Специализация областей коры мозга
Специализация областей коры мозга
 Физиология коры больших полушарий • • Высшим отделом ЦНС является кора больших полушарий, ее площадь составляет 2200 см 2. Кора больших полушарий имеет шестислойное строение. Нейроны представлены сенсорными, моторными (клетками Бетца), интернейронами (тормозными и возбуждающими нейронами). Кора полушарий построена по колончатому принципу. Колонки – функциональные единицы коры, делятся на микромодули, которые имеют однородные нейроны. Согласно современным представлениям о локализации функций при прохождении импульса в коре головного мозга возникают три типа полей. 1. Первичная проекционная зона лежит в области центрального отдела ядеранализаторов, где впервые появился электрический ответ (вызванный потенциал), нарушения в области центральных ядер ведут к нарушению ощущений. 2. Вторичная зона лежит в окружении ядра, не связана с рецепторами, по вставочным нейронам импульс идет из первичной проекционной зоны. Здесь устанавливается взаимосвязь между явлениями и их качествами, нарушения ведут к нарушению восприятий (обобщенных отражений). 3. Третичная (ассоциативная) зона имеет мультисенсорные нейроны. Информация переработана до значимой. Система способна к пластической перестройке, длительному хранению следов сенсорного действия. При нарушении страдают форма абстрактного отражения действительности, речь, целенаправленное поведение.
Физиология коры больших полушарий • • Высшим отделом ЦНС является кора больших полушарий, ее площадь составляет 2200 см 2. Кора больших полушарий имеет шестислойное строение. Нейроны представлены сенсорными, моторными (клетками Бетца), интернейронами (тормозными и возбуждающими нейронами). Кора полушарий построена по колончатому принципу. Колонки – функциональные единицы коры, делятся на микромодули, которые имеют однородные нейроны. Согласно современным представлениям о локализации функций при прохождении импульса в коре головного мозга возникают три типа полей. 1. Первичная проекционная зона лежит в области центрального отдела ядеранализаторов, где впервые появился электрический ответ (вызванный потенциал), нарушения в области центральных ядер ведут к нарушению ощущений. 2. Вторичная зона лежит в окружении ядра, не связана с рецепторами, по вставочным нейронам импульс идет из первичной проекционной зоны. Здесь устанавливается взаимосвязь между явлениями и их качествами, нарушения ведут к нарушению восприятий (обобщенных отражений). 3. Третичная (ассоциативная) зона имеет мультисенсорные нейроны. Информация переработана до значимой. Система способна к пластической перестройке, длительному хранению следов сенсорного действия. При нарушении страдают форма абстрактного отражения действительности, речь, целенаправленное поведение.
 обеспечение связи организма с внешней средой,") Основные функции коры больших полушарий: • • 1) обеспечение связи организма с внешней средой, приспособление его к ее изменениям; 2) координация движений (возможность осуществлять произвольные движения, делать непроизвольные движения более точными, осуществлять двигательные задачи); 3) согласование работы разных систем внутри организма; 4) высшая нервная деятельность (мышление, сознание, речь и др. ).
Основные функции коры больших полушарий: • • 1) обеспечение связи организма с внешней средой, приспособление его к ее изменениям; 2) координация движений (возможность осуществлять произвольные движения, делать непроизвольные движения более точными, осуществлять двигательные задачи); 3) согласование работы разных систем внутри организма; 4) высшая нервная деятельность (мышление, сознание, речь и др. ).
 Сенсорные области коры. Каждому рецепторному аппарату в коре соответствует определенная область, которую Павлов назвал корковым ядром анализатора. Именно к корковому ядру анализатора по афферентным волокнам приходят сигналы от рецепторов органов чувств. В сенсорных зонах выделяют первичные и вторичные проекционные поля. Нейроны проекционных первичных полей выделяют отдельные признаки сигнала (например, контур, цвет, контраст). Вторичные – формируют их в целостный образ. Сенсорные зоны локализованы в определенных частях коры: зрительная – в затылочной области, слуховая – в височной, вкусовая – в нижней части теменных областей, соматосенсорная зона (анализирующая импульсацию с рецепторов мышц, суставов, сухожилий и кожи) располагается в области постцентральной извилины.
Сенсорные области коры. Каждому рецепторному аппарату в коре соответствует определенная область, которую Павлов назвал корковым ядром анализатора. Именно к корковому ядру анализатора по афферентным волокнам приходят сигналы от рецепторов органов чувств. В сенсорных зонах выделяют первичные и вторичные проекционные поля. Нейроны проекционных первичных полей выделяют отдельные признаки сигнала (например, контур, цвет, контраст). Вторичные – формируют их в целостный образ. Сенсорные зоны локализованы в определенных частях коры: зрительная – в затылочной области, слуховая – в височной, вкусовая – в нижней части теменных областей, соматосенсорная зона (анализирующая импульсацию с рецепторов мышц, суставов, сухожилий и кожи) располагается в области постцентральной извилины.
 Чувствительный гомункулус Показаны проекции частей тела человека на область коркового конца анализатора
Чувствительный гомункулус Показаны проекции частей тела человека на область коркового конца анализатора
 • Моторные – зоны, раздражение которых вызывает двигательную реакцию, расположены в предцентральной извилине. В моторной коре тело человека спроецировано как бы вверх ногами, то есть ближе к латеральной борозде находятся области, обеспечивают функционирование мышц головы, а у противоположного конца предцентральной извилины - мышц нижней конечности.
• Моторные – зоны, раздражение которых вызывает двигательную реакцию, расположены в предцентральной извилине. В моторной коре тело человека спроецировано как бы вверх ногами, то есть ближе к латеральной борозде находятся области, обеспечивают функционирование мышц головы, а у противоположного конца предцентральной извилины - мышц нижней конечности.
 Двигательный гомункулус Показаны проекции частей тела человека на область коркового конца двигательного анализатора
Двигательный гомункулус Показаны проекции частей тела человека на область коркового конца двигательного анализатора
 обработка и хранение поступающей информации Б) переход") Функции ассоциативных зон • • • А) обработка и хранение поступающей информации Б) переход от наглядного восприятия к абстрактным символическим процессам. В) Мышление (внутренняя речь) возможно только при совместной деятельности различных сенсорных систем, объединение информации от которых происходит в ассоциативных полях. Г) Целенаправленное поведение человека, формирование намерений и планов, программ произвольных движений Д)Отвечают за согласованную работу обоих полушарий мозга. Как правило, одно из полушарий является ведущим – доминантным. У большинства если ведущая рука – правая, доминантное полушарие – левое. Левое лучше снабжается кровью, в нем больше взаимосвязей нейронов, в нем находится моторный центр речи, отвечающий за произнесение слов и сенсорный центр речи, отвечающий за понимание слов. У человека есть три формы межполушарной функциональной асимметрии, т. е. неодинакового вклада полушарий: моторная, сенсорная и психическая. Моторная и сенсорная – это когда у человека с ведущей правой рукой, главным является левый глаз или левое ухо. Причем в каждом полушарии есть центры, которые контролируют оба уха, оба глаза и т. д. Это дает возможность совмещать функции двух полушарий в одном, при повреждении. Психическая асимметрия проявляется в виде специализации полушарий. Левое больше отвечает за аналитические процессы, абстрактное мышление, логическое мышление, предвосхищение событий. Правое обрабатывает информацию целиком, не расчленяя на детали, преобладает предметное мышление, художественное, а функции связаны с прошлым, т. е. обработка информации на основе прошлого опыта.
Функции ассоциативных зон • • • А) обработка и хранение поступающей информации Б) переход от наглядного восприятия к абстрактным символическим процессам. В) Мышление (внутренняя речь) возможно только при совместной деятельности различных сенсорных систем, объединение информации от которых происходит в ассоциативных полях. Г) Целенаправленное поведение человека, формирование намерений и планов, программ произвольных движений Д)Отвечают за согласованную работу обоих полушарий мозга. Как правило, одно из полушарий является ведущим – доминантным. У большинства если ведущая рука – правая, доминантное полушарие – левое. Левое лучше снабжается кровью, в нем больше взаимосвязей нейронов, в нем находится моторный центр речи, отвечающий за произнесение слов и сенсорный центр речи, отвечающий за понимание слов. У человека есть три формы межполушарной функциональной асимметрии, т. е. неодинакового вклада полушарий: моторная, сенсорная и психическая. Моторная и сенсорная – это когда у человека с ведущей правой рукой, главным является левый глаз или левое ухо. Причем в каждом полушарии есть центры, которые контролируют оба уха, оба глаза и т. д. Это дает возможность совмещать функции двух полушарий в одном, при повреждении. Психическая асимметрия проявляется в виде специализации полушарий. Левое больше отвечает за аналитические процессы, абстрактное мышление, логическое мышление, предвосхищение событий. Правое обрабатывает информацию целиком, не расчленяя на детали, преобладает предметное мышление, художественное, а функции связаны с прошлым, т. е. обработка информации на основе прошлого опыта.