Лекция 2. Физиология пищеварения. Часть 1..ppt
- Количество слайдов: 54



Физиология пищеварения

Пищеварение – процесс усвоения биологического материала после предварительной деградации полимерных молекул до сравнительно простых элементов, лишенных видовой специфичности и способных проникать во внутреннюю среду организма или клетки Весь комплекс ассимиляции пищевых веществ из окружающей среды может быть объединен термином «экзотрофия» , т. е. усвоение пищевых веществ, поступающих из внешней среды.
 и Рене Декарт")
История изучения физиологии пищеварения Аристотель (384 -322 до н. э. ) и Рене Декарт (1596 -1650) – задумывались над процессами пищеварения, но их размышления их носили отвлеченный характер и были чисто умозрительными. Ренье де Грааф (1641 -1673 гг. ) - изучал слюнную и панкреатическую секрецию у собаки, вводя в протоки околоушной и поджелудочной желез трубочки, однако не предпринял попыток исследования свойств этих секретов Спалланцани (1729 -1799) и Реомюр (1683 -1757) пытались получить желудочный сок, вводя в желудок привязанную на ниточке резиновую губку. Примитивность методики и неточность проводимого анализа не дали ясного и полного представления о секреторной функции желудка.

История изучения физиологии пищеварения В начале XIX в. Пру обнаружил в желудочном содержимом наличие соляной кислоты, но не объяснил ее значения для желудочного пищеварения и роли в механизме регуляции деятельности органов пищеварения. С 1825 по 1833 г. Бомон проводит наблюдения над желудочной секрецией у человека, имевшего после ранения свищ желудка. Не учитывал исходного функционального состояния железистого аппарата, не принимал во внимание роли влияний коры больших полушарий головного мозга в процессе возбуждения секреторных клеток и не дал точной характеристики количественного и качественного состава желудочного сока при действии тех или иных пищевых раздражителей. Тидеман и Гмелин, а впоследствии Браун с целью выяснения роли механического раздражения в процессе возбуждения желудочной секреции вводили в желудок животных мелкие камни и спустя несколько часов, убив животных, смотрели, имеется ли в желудке сок. Леуре и Лассажне давали собакам проглатывать кусочки губки и через 24 ч, убив животных, исследовали кишечное содержимое, выжатое из губок.

История изучения физиологии пищеварения Шульц пищеварительную функцию приписывал только слепой кишке, а Блондло, подробно описывая различные пищеварительные соки, отрицал какое-либо переваривающее действие кишечного и панкреатического соков на пищу. К отрицанию способности кишечного сока переваривать пищу (белок и крахмал) пришел на основании своих опытов и Функ. В 1842 г. русский хирург В. А. Басов, наложив фистульную трубку на желудок собаки, впервые предложил новый метод исследования желудочной секреции. В 1849 г. Барделебен впервые произвел на собаке операцию перерезки пищевода (эзофаготомию), но ни сам автор, ни последующие физиологи не использовали этот ценный оперативный прием для научно-исследовательских целей Ряд исследователей (Флюранс, Гаубер, Колен, Элленбергер) применяли фистульный метод на разных сельскохозяйственных животных (коза, овца, корова), но полученные экспериментальные данные были отрывочными и неполными. В 1852 г. физиологи Ф. Биддер и С. Шмидт описали психическое отделение желудочного сока у собак, не дав этому явлению объективного, строго научного толкования.
 1897 г. - классический труд")
История изучения физиологии пищеварения Иван Петрович Павлов (1849 -1936) 1897 г. - классический труд «Лекции о работе главных пищеварительных желез» 1902 - «Физиологическая хирургия пищеварительного тракта» 1904 г. - первым среди русских ученых и первым из физиологов был удостоен Нобелевской премии 1907 г. - был избран действительным членом Петербургской Академии наук. И. П. Павлов - основатель современной физиологии пищеварения

Основные достижения И. П. Павлова в области физиологии пищеварения разработал метод хронических фистул, что дало возможность изучать деятельность органов на неповрежденном животном – основа современной экспериментальной физиологии установил основные физиологические механизмы, регулирующие деятельность пищеварительных желез и пищеварительного тракта в целом экспериментально обосновал учение о пищевом центре, согласно которому регуляция двигательных и секреторных процессов, протекающих во время пищеварения, осуществляется при помощи коры больших полушарий и подкорковых ядер
 Крупнейший российский специалист в области")
История изучения физиологии пищеварения Уголев Александр Михайлович (1926 -1991) Крупнейший российский специалист в области физиологии, вегетативных функций и их регуляции. Академик АН СССР по отделению физиологии (1984). Впервые описал пристеночное пищеварение, исследовал эволюцию пищеварительной функции. Основные труды: 1961 - «Пищеварение и его приспособительная эволюция» 1963 - «Пристеночное (контактное) пищеварение» 1967 - «Физиология и патология пристеночного (контактного) пищеварения» 1972 - «Мембранное пищеварение. Полисубстратные процессы, организация и регуляция» 1987 - «Естественные технологии биологических систем» 1991 - «Теория адекватного питания и трофология» За свои работы был награждён золотой медалью им. И. И. Мечникова (1990).
 с")
А. М. Уголев охарактеризовал пищеварительный тракт как химический завод, обрабатывающий сырой материал (пищу) с использованием сложного технологического процесса, включающего: сортировку пищи по отделениям специальные реактивы, синтезированные на отдаленных небольших фабриках (железах) системы трубопроводов, связывающих этот комплекс в единое целое (ЖКТ, протоки желез) У человека 1010 кишечных клеток и 1015 соматических, значит 1 кишечная клетка обеспечивает 100000 соматических !!!

Специализированные регионы пищеварительной системы 1. Регион поглощения, механической обработки пищи и первичной обработки с помощью слюны 2. Передающий регион, который может также осуществлять функцию хранения. 3. Пищеварительный регион: а) регион начального пищеварения, б) регион конечного пищеварения и всасывания. 4. Регион реабсорбции - всасывания воды, солей и ряда эндогенных компонентов (например, конъюгированных желчных кислот и др. ).

Основные компоненты пищи*: Белки Жиры Углеводы Нуклеиновые кислоты * Исключение - эндопаразиты Этапы деградации пищи: Механическое измельчение Физико-химическое разрушение: кислый p. H, биологические детергенты Ферментативный гидролиз

Превращение исходных пищевых веществ во всасываемые продукты происходит под действием гидролитических ферментов. Для расщепления каждого пищевого биополимера (белков, жиров, углеводов) существует своя ферментная цепь. Общей закономерностью, справедливой для всех живых существ, является первоначальное переваривание в кислой среде и последующий гидролиз и всасывание в щелочной. У примитивных организмов это достигается изменением реакции в пищеварительных вакуолях. У высших животных отдельные звенья цепи реализуются в различных регионах ЖКТ.
 поразительно схожи")
Общая характеристика пищеварительных ферментов Ферментные системы различных организмов (от простейших до высших) поразительно схожи ! 1. Пептид-гидролазы (протеазы, протеиназы, пептидазы) - расщепляют пептидные связи в белках и полипептидах 2. Гликозидазы (карбогидразы) – гидролизуют гликозидные связи в углеводах 3. Липазы – действуют на эфирные связи в жирах 4. Нуклеазы - гидролиз фосфодиэфирных связей нуклеиновых кислот с образованием моно- и олигонуклеотидов
 экзопептидазы (атакуют концевые пептидные связи) эндопептидазы (специфичны к определенным")
1. Пептид-гидролазы (протеазы, протеиназы, пептидазы) экзопептидазы (атакуют концевые пептидные связи) эндопептидазы (специфичны к определенным внутренним пептидным связям) – расщепляют большую часть белков пищи Действуют как внутриклеточно (катепсины - в пищеварительных вакуолях и лизосомах – активны при слабокислом p. H), так и внеклеточно. Катепсины могут встречаться и в непищеварительных органах – выполняют функцию деградации и изменения соответствующих ферментативно активных белков.
 Эндопептидазы I. Кислые протеазы – у беспозвоночных практически не встречаются")
Пептид-гидролазы (протеазы, протеиназы, пептидазы) Эндопептидазы I. Кислые протеазы – у беспозвоночных практически не встречаются a) пепсин (пепсины) - оптимум при p. H=1. 5 -2. 0 - расщепляет белки до полипептидов - у позвоночных секретируется в виде профермента пепсиногена б) гастриксин (пепсин С, парапепсин II) - 20 -50% от пепсина - оптимум при p. H=3. 2 -3. 5 - у человека - секретируется в форме прогастриксина в) пепсин В (парапепсин, желатиназа) - выделяется в виде пепсиногена В - активируется ионами Са 2+ - расщепляет белки соединительной ткани г) химозин (реннин)- первый фермент выделенный химически в 1874 г - выделяется в виде профермента, активируется ионами Н+ - створаживает молоко – расщепляет казеиноген до казеина
 Эндопептидазы II. Сериновые протеазы – у позвоночных и человека, синтезируются")
Пептид-гидролазы (протеазы, протеиназы, пептидазы) Эндопептидазы II. Сериновые протеазы – у позвоночных и человека, синтезируются в поджелудочной железе (активны при p. H>7) а) трипсин - образуется из трипсиногена под действием энтерокиназы или аутолиза; - гидролизует пептидные связи между карбоксильными группами L-аргинина и L-лизина б) химотрипсин - образуется из химотрипсиногена под действием трипсина; - гидролизует связи между карбоксильными группами ароматических аминоксилот в) эластаза - секретируется в виде проэластазы, активируется трипсином - гидролизует связи между нейтральными аминокислотами (в эластине)
 Эндопептидазы Сопоставление последовательностей амикислотных остатков показало гомологичное сходство трипсина, химотрипсина")
Пептид-гидролазы (протеазы, протеиназы, пептидазы) Эндопептидазы Сопоставление последовательностей амикислотных остатков показало гомологичное сходство трипсина, химотрипсина и эластазы. Существует теория происхождения этих ферментов от общего «предка» нейтральной протеазы. Ферменты с трипсиновой и химотрипсиновой активностью обнаружены в кишечнике беспозвоночных животных (кишечнополостных и насекомых) – но в отличие от ферментов позвоночных они секретируются в активной форме. - кератиназа актиномицетов - коллагеназа (бактерии, крабы, яд некоторых змей) - фибринолизин (бактерии)
 Экзопептидазы I. Карбоксипептидазы А и B секретируются ПЖ в виде")
Пептид-гидролазы (протеазы, протеиназы, пептидазы) Экзопептидазы I. Карбоксипептидазы А и B секретируются ПЖ в виде прокарбоксипептидаз активируются трипсином и энтерокиназой также обнаружены в кишечнике беспозвоночных II. Аминопептидазы составляют основную часть пептидазной активности щеточной каймы клеток кишечного эпителия; расщепляют олигопептиды, трипептиды, дипептиды III. Дипептидазы завершают гидролиз белков; участвуют в мембранном гидролизе пептидов в тонк. кишечнике позвоночных и органах пищеварения ряда беспозвоночных; работают внутри клеток бактерий и тонкого кишечника позвоночных (катаболизм белков)
 I. Эндогликозидазы - расщепляют α– 1, 4 -глюкозидные связи в полисахаридах")
Гликозидазы (глюкозидазы, карбогидразы) I. Эндогликозидазы - расщепляют α– 1, 4 -глюкозидные связи в полисахаридах (крахмал, гликоген и т. п. ) до олиго- и дисахаридов. а) амилазы (не гидролизуют целлюлозу !!!) амилаза слюны млекопитающих, ряда хордовых и беспозвоночных панкреатическая амилаза млекопитающих кишечная гамма-амилаза хордовых и ряда беспозвоночных внутриклеточные амилазы кишечнополостых и простейших II. Экзогликозидазы (α- и β-гликозидазы) – гидролизуют олиго- и дисахариды (мальтозу, изомальтозу, сахарозу, лактозу и трегалозу) до моно-. Синтезируются в энтероцитах как хордовых, так и беспозвоночных. Сходны с внутриклеточными гликозидазами беспозвоночных а) мальтаза и изомальтаза б) сахараза (инвертаза) – сахарозу на глюкозу и фруктозу в) лактаза – лактозу на глюкозу и галактозу

Переваривание целлюлозы Ферментные системы для переваривание целлюлозы есть у небольшого числа видов: ряд брюхоногих и пластинчатожаберных моллюсков некоторые ракообразные некоторые рептилии и рыбы бактерии, простейшие, грибы, кольчатые черви, иглокожие (используют целлюлазу) Большинство травоядных животных для переваривания целлюлозы использует симбионтные организмы
 панкреатическая фосфолипаза секретируется в виде профосфолипазы А, активируется трипсином гидролизует эфирную связь")
Липазы а) панкреатическая фосфолипаза секретируется в виде профосфолипазы А, активируется трипсином гидролизует эфирную связь глицерина и ж. кислоты у 2 -го атома углерода б) кишечная моноглицеридлипаза гидролизует эфирные связи диглицеридов в) эстеразы в средней кишке ракообразных, у насекомых, двустворчатых моллюсков (как вне-, так и внутриклеточные эстеразы), у брюхоногих моллюсков, многощетинковых червей, иглокожих, простейших
 Щелочная фосфатаза - гидролизует моноэфиры ортофосфорной")
Другие пищеварительные ферменты (известны в основном у хордовых) Щелочная фосфатаза - гидролизует моноэфиры ортофосфорной кислоты Нуклеазы, нуклеотидазы, нуклеозидазы расщепляют полинуклеотиды и нуклеиновые кислоты до пуринов, пиримидинов, дезоксирибозы, рибозы и фосфата; продуцируются клетками поджелудочной железы и клетками кишечного эпителия хордовых.

Экзотрофия - это такой же древний процесс, как и сама жизнь…. . А. М. Уголев Основные типы пищеварительных процессов I. Внеклеточное (дистантное) п. - является основным у организмов, стоящих на более высоком этапе эволюционного развития, чем плоские черви, и преобладает у кольчатых червей, ракообразных, насекомых, головоногих, оболочников и хордовых ферменты выделяются во внеклеточную среду секреторные клетки расположены далеко от полостей, где реализуется действие ферментов дистантное пищеварение, происходящее в специальных полостях, обозначается как полостное может происходить и вне организма, продуцирующего ферменты (паукообразные и насекомые, бактерии*)

Основные типы пищеварительных процессов II. Внутриклеточное п. - простейшие, примитивные многоклеточные (например губки и плоские черви) Amoeba proteus Planaria spp. Porifera

Основные типы пищеварительных процессов Внутриклеточное пищеварение транспорт молекул через клеточные пищеварение с помощью лизосом мембраны; с набором гидролитических ферментов с оптимумом работы при переваривание в пищеварительных p. H 3. 5 -5. 5; вакуолях, образующихся при пино- и фагоцитозе, которые исчезают после пино- и фагоцитозные вакуоли расщепления пищи соединяются с лизосомами, образуя фагосомы; продукты гидролиза, всасываются в цитоплазму через мембрану фагосом; остатки фагосом выбрасываются путем экзоцитоза

Основные типы пищеварительных процессов III. Мембранное п. - осуществляется как адсорбированными из полости тонкой кишки ферментами: панкреатического происхождения, секретируемые кишечными железами, так и мембранными ферментами, синтезированными в кишечных клетках и встроенными в состав их апикальной мембраны. Мембранное пищеварение наблюдается у организмов на всех уровнях эволюционного развития (в т. ч. у дрожжей, бактерий и корней растений), т. е. является универсальным механизмом. Щеточная кайма кишечной клетки крысы (Уголев и др. , 1987)

Основные типы пищеварительных процессов Мембранное пищеварение Для правильной оценки мембранного пищеварения существенны следующие факторы: 1. Ферменты, реализующие этот процесс, относятся к структурированным. Структурирование ферментов приводит к изменению их свойств. 2. Возможна пространственная организация ферментных и транспортных систем, объединяющих заключительные этапы переваривания и начальные этапы всасывания 3. Зона мембранного пищеварения обладает особыми физико-химическими свойствами, в частности, такими как р. Н, концентрация органических и неорганических ионов, неперемешиваемый слой жидкости.

Основные типы пищеварительных процессов Мембранное пищеварение 4. Мембранное пищеварение осуществляется в стерильной зоне, недоступной для бактерий, что предотвращает поглощение последними низкомолекулярных пищевых веществ. 5. Благодаря ферментному аппарату гликокаликсный слой превращается в высокоспецифический фильтр. Через него проникают только вещества, для которых на поверхности и внутри гликокаликсного пространства имеются адекватные ферменты. 6. Микрофиламенты микроворсинок содержат актин и миозин.

Основные типы пищеварительных процессов Согласно современным представлениям, переваривание пищи у высокоорганизованных организмов происходит в 3 этапа: полостное п. мембранное п. всасывание + внутриклеточное п. Упрощенная схема аллостерических взаимодействий ферментной и транспортной частей комплекса. А — аллостерические влияния фермента на вход в транспортную систему; Б — аллостерические влияния транспортной системы на активность фермента. 1 — субстрат; 2 — продукт; 3 — фермент; 4 — транспортная система; 5 — мембрана

Основные типы пищеварительных процессов В зависимости от источника ферментов можно выделить следующие типы пищеварения: 1) собственное пищеварение, когда источником фермента служит сам организм; 2) симбионтное пищеварение, для которого характерна продукция ферментов симбионтами - бактериями и простейшими (характерно для жвачных животных); 3) аутолитическое пищеварение, происходящее за счет ферментов, содержащихся в пище.
")
Теории происхождения внешней и внутренней секреции Теория происхождения секреции Иордана I. Морфонекротическая с. (голокриновая) II. Морфокинетическая (апокриновая) III. Морфостатическая (истинная, мерокриновая)

Выводы на основе экскреторной теории 1. Истинная секреция возможна уже у простейших, при их переходе от свободного к паразитическому образу жизни. 2. Внешняя и внутренняя секреция могут формироваться многократно в процессе филогенеза в различных по своему происхождению структурах. 3. Морфонекротическая секреция является тупиковой ветвью эволюции пищеварительной системы или ее некоторых отделов. В ряде случаев она оказалась настолько эффективной, что сохранилась у высокоорганизованных животных. Кишечная секреция позвоночных происходит в основном по морфонекротическому типу.

Выводы на основе экскреторной теории 5. Внешняя и внутренняя секреция — родственные процессы, причем некоторые органы (поджелудочная железа, желудок, тонкая кишка) обладают как внешнесекреторной, так и внутрисекреторной функциями. 7. При каких условиях и под влиянием каких факторов некоторые метаболиты в процессе эволюции становятся жизненно важными регуляторами типа гормонов? Возможна первоначальная экскреция веществ-метаболитов не имеющих мишени, также как и существование мишеней (рецепторов), которые в течение длительного времени остаются без вещества-регулятора (у асцидий содержится гормон окситоцин, действующий лишь на структуры позвоночных животных). Время, разделяющее появление этих компонентов, может исчисляться миллионами лет.

Пищеварение у человека Для осуществления нормального пищеварения необходимо: -движение пищи по ЖКТ; - секреция пищеварительных соков и переваривание пищи; - всасывание продуктов переваривания пищи, воды и электролитов; - система кровоснабжения ЖКТ для транспорта мономеров; - контроль функций с помощью местных механизмов, нервной и эндокринной систем

Основные принципы функционирования ЖКТ: сократительная способность, нервная регуляция, кровоснабжение I. Основные принципы сократительной активности ЖКТ Слои стенки ЖКТ: 1. Серозный; 2. Слой продольной мускулатуры; 3. Слой кольцевой мускулатуры; 4. Подслизистый слой; 5. Слизистая * - в глубоких слоях слизистой есть скопления гладкомышечных клеток

I. Основные принципы сократительной активности ЖКТ Особенности гладкомышечной ткани в ЖКТ: - клетки длиной 200 -500 мкм, диаметром 2 -10 мкм, в составе пучка ~ 1000 волокон, идущих параллельно; -между отдельными пучками рыхлая соединительная ткань, соседние пучки имеют многочисленные точки соприкосновения; - клетки связаны щелевыми контактами; - гладкомышечные клетки функционируют как синцитий; - ПД распространяется от нескольких мм до десятков см (иногда по всему ЖКТ), в зависимости от возбудимости мышечной ткани; - существует связь между продольной и кольцевой мускулатурой

I. Основные принципы сократительной активности ЖКТ Электрическая активность гастроинтестинальной гладкой мускулатуры: 1. Медленные волны 2. Спайковая активность

I. Основные принципы сократительной активности ЖКТ Электрическая активность гастроинтестинальной гладкой мускулатуры: Медленные волны: - не являются ПД, медленное изменение мембранного потенциала засчет РNa+; - амплитуда 5 -15 м. В, частота в различных частях ЖКТ 3 -12/мин (3/мин в теле желудка, 12/мин. в 12 -перстной кишке, 8 -9/мин в подвздошной кишке); - причина возникновения до конца не ясна – результат взаимодействия гладкомышечных клеток и интерстициальных клеток Кахаля, которые предположительно являются пейсмекерами; - медленные волны не вызывают сокращений гладкой мускулатуры облегчают возникновение периодических спайковых потенциалов, вызывающих сокращение.

I. Основные принципы сократительной активности ЖКТ Электрическая активность гастроинтестинальной гладкой мускулатуры: Спайковые потенциалы: - являются настоящими потенциалами действия; - возникают автоматически, когда ПП гастроинтестинальной клетки (в норме – 50 -60 м. В) превышает – 40 м. В; - чем «выше» медленная волна – тем больше частота ПД на ее «гребне» (1 -10 Гц); эти ПД в 10 -40 раз длиннее, чем ПД крупных нервных волокон ~ 10 -20 мс; - ПД обеспечивается смешанным Ca-Na током; - входящие ионы Ca 2+ обеспечивают сокращение гладкомышечной клетки.

I. Основные принципы сократительной активности ЖКТ Электрическая активность гастроинтестинальной гладкой мускулатуры: Изменение уровня потенциала покоя (в норме – 56 м. В): I. Деполяризация: - растяжение клеточной мембраны; - воздействие АЦХ; - стимуляция парасимпатических нервов; - действие специфических гастроинтестинальных гормонов II. Гиперполяризация: - действие норадреналина или адреналина; - стимуляция симпатических нервов

Основные принципы сократительной активности ЖКТ Электрическая активность гастроинтестинальной гладкой мускулатуры: Тонические сокращения гастроинтестинальной мускулатуры: - продолжительность – минуты или даже часы; - часто являются следствием повторяющихся ПД – чем больше частота, тем больше амплитуда сокращения; - могут быть следствием влияния гуморальных агентов, вызывающих длительную деполяризацию мембраны; - в результате [Ca 2+] не связанного с изменением МП (механизм неизвестен)

I. Нервный контроль гастроинтестинальных функций Энтеральная нервная система ЭНС – лежит в стенке ЖКТ, начиная с пищевода до анального отверстия; ~ 100 млн. нейронов ~ числу нейронов в спинном мозге; функция – контроль перистальтики и секреции; образована миэнтеральным (ауэрбаховым) и подслизистым (мейснеровым) сплетениями; ЭНС может функционировать независимо от внешних регулирующих воздействий

I. Нервный контроль гастроинтестинальных функций Энтеральная нервная система Миэнтеральное сплетение: 1. состоит из цепочки связанных нейронов на протяжении всего ЖКТ; 2. стимуляция ведет к: - усиление тонического сокращения (тонуса); - усиление интенсивности ритмических сокращений; - небольшое учащение ритмических сокращений; - увеличение скорости проведение волн деполяризации, что ведет к ускорению перистальтических волн - сплетение не является полностью возбуждающим, ряд нейронов тормозные (из окончаний выделяется тормозный пептидный медиатор возможно ВИП). Важно для расслабления мышц сфинктеров пилорического, илеоцекального)

I. Нервный контроль гастроинтестинальных функций Энтеральная нервная система Мейснерово сплетение: - контроль за состоянием внутренней стенки каждого небольшого сегмента ЖКТ; - контроль за секрецией, всасыванием и локальной сократительной активностью подслизистого мышечного слоя
 - в")
I. Нервный контроль гастроинтестинальных функций Энтеральная нервная система Медиаторы ЭНС: ацетилхолин (АЦХ) - в основном стимулирует деятельность ЖКТ; норадреналин - почти всегда тормозит деятельность ЖКТ; АТФ (вазодилататор); серотонин; дофамин; холецистокинин (ХЦК); субстанция Р (sub P); вазоактивный интестинальный пептид (ВИП) (вазодилататор); лейэнкефалин (leu-enkephalin); метэнкефалин (met-enkephalin); бомбезин NO (вазодилататор)

I. Нервный контроль гастроинтестинальных функций Парасимпатическая иннервация Краниальный отдел - волокна блуждающего нерва (n. vagus, X пара), за исключением нескольких волокон, иннервирующих ротовую полость и глотку. Иннервирует пищевод, желудок, ПЖ, и кишечник до середины толстого кишечника. Сакральный отдел тазовые нервы дистальная часть толстого кишечника до анального отверстия. Сигмоидальная, ректальная и анальная области иннервированы лучше других. Эти волокна вовлечены в процесс дефекации. Постганглионарные нейроны расположены в миэнтеральном и мейснеровом сплетениях. Стимуляция PS активирует ЭНС и большинство функций ЖКТ

I. Нервный контроль гастроинтестинальных функций Симпатическая иннервация S волокна берут начало в спинном мозге (Th 5 -L 2); Часть преганглионарных волокон прерываются в S цепочке; Часть волокон пронизывает S цепочку и прерывается в превертебральных ганглиях (чревные и брыжеечные); Постганглионарные S нервы иннервируют все отделы ЖКТ; Из окончаний выделяется норадреналин и адреналин (очень немного) Стимуляция S нервов ведет к торможению функций ЖКТ, многие эффекты антагонистичны PS. Это достигается 2 -мя путями: 1) небольшое влияние через прямое торможение гладкой мускулатуры (кроме мускулатуры слизистой возбуждается); 2) мощное тормозное действие на нейроны ЭНС
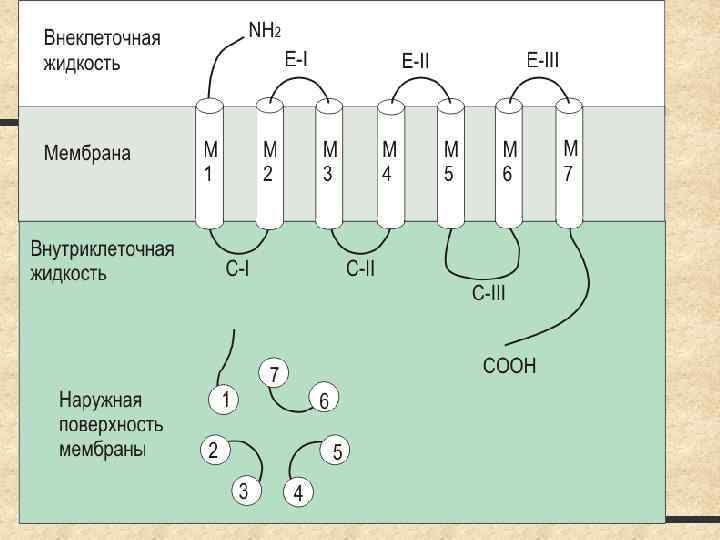
 нервная система M 1 и М 3")
I. Нервный контроль гастроинтестинальных функций Автономная (вегетативная) нервная система M 1 и М 3 холинорецепторы α- и β- адренорецепторы: α 2 –тормозные (на возб. холинергических нейронах)
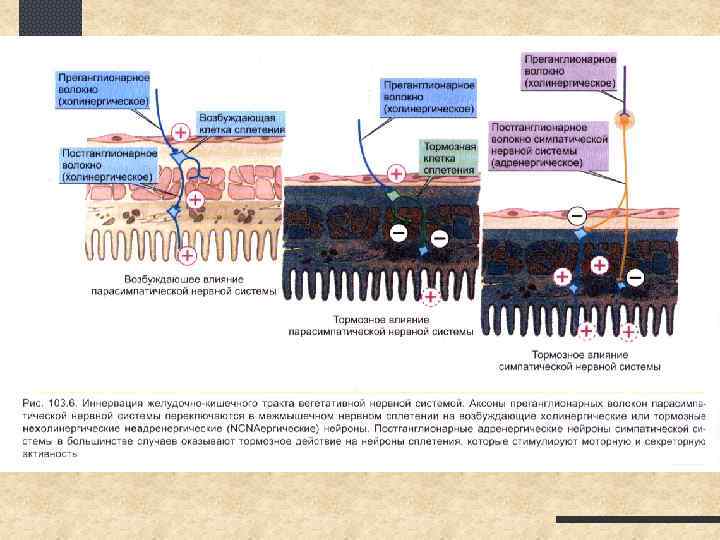

I. Нервный контроль гастроинтестинальных функций Афферентные сенсорные пути - тела сенсорных нейронов находятся в ЭНС или в ганглиях дорзальных корешков с. м. ; - афферентные нейроны могут активироваться при: 1. раздражении слизистой; 2. растяжении стенок ЖКТ; 3. действии специфических веществ, в результате - или активация или торможение перистальтики и секреции; - часть сенсорной информации поступает в спинной мозг и ствол (например 80% волокон вагуса - афферентные)

I. Нервный контроль гастроинтестинальных функций Гастроинтестинальные рефлексы I. Рефлексы замыкающиеся на уровне ЭНС - контролируют секрецию, перистальтику, локальные тормозные влияния II. Рефлексы от тракта к превертебральным S ганглиям и обратно к ЖКТ - гастроколический рефлекс - от желудка к ободочной кишке - опорожнение; - энтерогастральные рефлексы - сигналы от ободочной кишки и тонкого кишечника тормозят сокращения и секрецию желудка; - колоно-илеальный рефлекс - сигналы от ободочной кишки тормозят движение содержимого в подвздошную кишку. III. Рефлексы от ЖКТ в спинной мозг или ствол и обратно в ЖКТ - от желудка и 12 -перстной кишки обратно к желудку (n. vagus) - контроль за сокращением и секрецией желудка; - болевые рефлексы тормозящие деятельность ЖКТ; - дефекация - сигнал от ободочной и прямой кишки в спинной мозг и обратно к гладкой мускулатуре кишечника и скелетной мускулатуре туловища

II. Гуморальный контроль гастроинтестинальных функций 1. Гастрин - секретируется G-клетками антральной части желудка в ответ на: - растяжение стенок желудка; - воздействие продуктов расщепления белков; - действие гастрин-рилизинг-пептида (GLP) (выделяется в слизистой желудка в ответ на стимуляцию вагуса). Эффекты гастрина - 1) стимуляция секреции соляной кислоты в желудке; 2) рост и пролиферация клеток слизистой желудка 2. Холецистокинин - секретируется I-клетками 12 -перстной и тощей кишки в ответ на жирные кислоты, моноглицериды. Эффекты ХЦК - 1) сокращение желчного пузыря и секреция желчи в 12 -п. к. ; 2) умеренно тормозит сокращения желудка; 3) тормозит эвакуацию пищи из желудка в 12 -п. к.
 - выделяется Sклетками")
II. Гуморальный контроль гастроинтестинальных функций 3. Секретин (первый открытый гастроинтестинальный гормон) - выделяется Sклетками слизистой 12 -п. к. в ответ на кислый химус, поступающий из желудка. - оказывает мягкое стимулирующее влияние на моторные ф-ции ЖКТ; - стимулирует панкретическую секрецию бикарбонатов, для нейтрализации кислого р. Н. 4. Гастроингибирующий пептид (ГИП, GIP) секретируется слизистой верхних отделов тонкого кишечника в ответ на жирные кислоты, аминокислоты и, в меньшей степени, на углеводы. - мягкий тормозный эффект на моторику желудка; - тормозит эвакуацию химуса желудка, когда тонкий кишечник заполнен; 5. Мотилин - секретируется верхними отделами 12 -п. к. Единственная известная функция - усиление моторики ЖКТ. Секреция циклична, сопровождается волнами сокращений желудка и верхнего тонкого кишечника - раз в 90 мин. Секреция снижается после приема пищи. Механизм неясен.
Лекция 2. Физиология пищеварения. Часть 1..ppt