

Практика 1 Ф ЦНС студентам.ppt
- Количество слайдов: 113
 Физиология ЦНС. ПЗ № 1. Физиология нервов и синапсов
Физиология ЦНС. ПЗ № 1. Физиология нервов и синапсов
 Формируемые компетенции: • Актуальность темы: данная тема вносит вклад в формирование следующих компетенций: ОК 1, ПК 1, ПК 5, ПК 9, ПК 16, ПК 27, ПК 31. • Знание изучаемой темы необходимо для понимания процессов регуляции функций организма человека.
Формируемые компетенции: • Актуальность темы: данная тема вносит вклад в формирование следующих компетенций: ОК 1, ПК 1, ПК 5, ПК 9, ПК 16, ПК 27, ПК 31. • Знание изучаемой темы необходимо для понимания процессов регуляции функций организма человека.
. •") Цели занятия: • Учебная: • Изучить функции нервной системы (ПК 5, ПК 16). • Изучить особенности функционирования нервных клеток, нервных волокон (ПК 5, ПК 16). • Изучить законы проведения возбуждения в нервах (ПК 5, ПК 16). • Изучить функционирование синапсов (ПК 5, ПК 16). • Научиться применять терминологию по изучаемой теме (ПК 5, ПК 16, ПК 27, ПК 31).
Цели занятия: • Учебная: • Изучить функции нервной системы (ПК 5, ПК 16). • Изучить особенности функционирования нервных клеток, нервных волокон (ПК 5, ПК 16). • Изучить законы проведения возбуждения в нервах (ПК 5, ПК 16). • Изучить функционирование синапсов (ПК 5, ПК 16). • Научиться применять терминологию по изучаемой теме (ПК 5, ПК 16, ПК 27, ПК 31).
 Цели занятия: • Развивающая: • Формирование системного подхода к пониманию регуляции функций организма человека (ОК 1, ПК 5, ПК 9, ПК 16, ПК 27). • Воспитательная: • Формирование способности и готовности реализовать этические и деонтологические аспекты врачебной деятельности в общении с коллегами (ПК 1).
Цели занятия: • Развивающая: • Формирование системного подхода к пониманию регуляции функций организма человека (ОК 1, ПК 5, ПК 9, ПК 16, ПК 27). • Воспитательная: • Формирование способности и готовности реализовать этические и деонтологические аспекты врачебной деятельности в общении с коллегами (ПК 1).
 Межпредметные связи Фармакология Патологическая физиология Биохимия Неврология Физиология нервов и синапсов Гистология, цитология, эмбриология Нормальная физиология Клиническая патофизиология Анатомия Физика. Математика
Межпредметные связи Фармакология Патологическая физиология Биохимия Неврология Физиология нервов и синапсов Гистология, цитология, эмбриология Нормальная физиология Клиническая патофизиология Анатомия Физика. Математика
 Внутрипредметные связи ПЗ№ 2. Возбуждение в ЦНС. Торможение и координационная деятельность ЦНС. Рефлекс и функциональная система ПЗ№ 8. Методы исследования функций ЦНС ПЗ№ 7. Нервная регуляция висцеральных функций. Физиологияавтономной (вегетативной) нервной системы ПЗ№ 3. Частная физиология ЦНС. Спинной мозг Физиология нервов и синапсов ПЗ№ 6. Кора больших полушарий ПЗ№ 4. Частная физиология ЦНС. Продолговатый мозг, мост, мозжечок ПЗ№ 5. Частная физиология ЦНС. Ретикулярная формация. Промежуточный, передний мозг. Лимбическая система
Внутрипредметные связи ПЗ№ 2. Возбуждение в ЦНС. Торможение и координационная деятельность ЦНС. Рефлекс и функциональная система ПЗ№ 8. Методы исследования функций ЦНС ПЗ№ 7. Нервная регуляция висцеральных функций. Физиологияавтономной (вегетативной) нервной системы ПЗ№ 3. Частная физиология ЦНС. Спинной мозг Физиология нервов и синапсов ПЗ№ 6. Кора больших полушарий ПЗ№ 4. Частная физиология ЦНС. Продолговатый мозг, мост, мозжечок ПЗ№ 5. Частная физиология ЦНС. Ретикулярная формация. Промежуточный, передний мозг. Лимбическая система
 Задания входного контроля Подпишите структуры, обозначенные буквами и цифрами.
Задания входного контроля Подпишите структуры, обозначенные буквами и цифрами.
 1. Установите соответствие Состояние мембраны обусловлена во время возбуждения 1. Начальная А избыточным выходом деполяризация калия из клетки на ее (предспайк) поверхность. 2. Полная деполяризация В уменьшением натриевого мембраны тока в клетку и (приближение к увеличением калиевого вершине потенциала тока из клетки. действия) 3. Реполяризация С резким увеличением калиевого тока в клетку. 4. Гиперполяризация D резким увеличением натриевого тока в клетку. Е незначительным входом ионов натрия в клетку.
1. Установите соответствие Состояние мембраны обусловлена во время возбуждения 1. Начальная А избыточным выходом деполяризация калия из клетки на ее (предспайк) поверхность. 2. Полная деполяризация В уменьшением натриевого мембраны тока в клетку и (приближение к увеличением калиевого вершине потенциала тока из клетки. действия) 3. Реполяризация С резким увеличением калиевого тока в клетку. 4. Гиперполяризация D резким увеличением натриевого тока в клетку. Е незначительным входом ионов натрия в клетку.
") 2. Установите соответствие 1. 2. 3. 4. 5. Фаза потенциала действия Начальная деполяризация (предспайк) Восходящая часть спайка Нисходящая часть спайка Отрицательный следовый потенциал Положительный следовый потенциал соответствует А абсолютной рефрактерности. В относительной рефрактерности. субнормальной возбудимости. супернормальной возбудимости. фазе экзальтации. С D Е
2. Установите соответствие 1. 2. 3. 4. 5. Фаза потенциала действия Начальная деполяризация (предспайк) Восходящая часть спайка Нисходящая часть спайка Отрицательный следовый потенциал Положительный следовый потенциал соответствует А абсолютной рефрактерности. В относительной рефрактерности. субнормальной возбудимости. супернормальной возбудимости. фазе экзальтации. С D Е
 3. Установите соответствие Свойство нервных обусловлено волокон 1. Изолированное А наличием шванновской проведение возбуждения оболочки. по нервным волокнам 2. Сальтаторный принцип В наличием в миелиновой проведения возбуждения оболочке перехватов в нервных волокнах Ранвье. 3. Различная скорость С коротким периодом проведения возбуждения рефрактерности. в нервных волокнах D толщиной волокон и наличием или отсутствием миелиновой оболочки. Е синтезом медиатора в нервных окончаниях.
3. Установите соответствие Свойство нервных обусловлено волокон 1. Изолированное А наличием шванновской проведение возбуждения оболочки. по нервным волокнам 2. Сальтаторный принцип В наличием в миелиновой проведения возбуждения оболочке перехватов в нервных волокнах Ранвье. 3. Различная скорость С коротким периодом проведения возбуждения рефрактерности. в нервных волокнах D толщиной волокон и наличием или отсутствием миелиновой оболочки. Е синтезом медиатора в нервных окончаниях.
 4. Установите соответствие Свойство нервносвязано с мышечного синапса 1. Синаптическая А наличием хеморецепторов на задержка постсинаптической мембране. 2. Одностороннее В следовыми колебаниями проведение потенциала на постсинаптической мембране. 3. Низкая С нечувствительностью лабильность пресинаптической мембраны к медиатору. 4. Повышенная D накоплением медиатора на чувствительность к постсинаптической мембране. химическим веществам Е диффузией медиатора через синаптическую щель.
4. Установите соответствие Свойство нервносвязано с мышечного синапса 1. Синаптическая А наличием хеморецепторов на задержка постсинаптической мембране. 2. Одностороннее В следовыми колебаниями проведение потенциала на постсинаптической мембране. 3. Низкая С нечувствительностью лабильность пресинаптической мембраны к медиатору. 4. Повышенная D накоплением медиатора на чувствительность к постсинаптической мембране. химическим веществам Е диффузией медиатора через синаптическую щель.
") Установите правильную последовательность: • 1. смены состояний мембраны в одиночном цикле возбуждения: • 1) следовая гиперполяризация • 2) следовая деполяризация • 3) местная деполяризация • 4) реполяризация мембраны • 5) деполяризация мембраны.
Установите правильную последовательность: • 1. смены состояний мембраны в одиночном цикле возбуждения: • 1) следовая гиперполяризация • 2) следовая деполяризация • 3) местная деполяризация • 4) реполяризация мембраны • 5) деполяризация мембраны.
") Установите правильную последовательность • 2. смены фаз возбудимости при генерации потенциала действия: • 1) субнормальная возбудимость • 2) экзальтация • 3) относительная рефрактерность • 4) абсолютная рефрактерность • 5) латентное дополнение.
Установите правильную последовательность • 2. смены фаз возбудимости при генерации потенциала действия: • 1) субнормальная возбудимость • 2) экзальтация • 3) относительная рефрактерность • 4) абсолютная рефрактерность • 5) латентное дополнение.
 Функции НС Основная задача нервной системы — интеграция внешнего воздействия с соответствующей приспособительной реакцией организма. Основные функции нервной системы: получение, хранение, переработка информации из внешней и внутренней среды, регуляция и координация деятельности всех органов и органных систем.
Функции НС Основная задача нервной системы — интеграция внешнего воздействия с соответствующей приспособительной реакцией организма. Основные функции нервной системы: получение, хранение, переработка информации из внешней и внутренней среды, регуляция и координация деятельности всех органов и органных систем.
 Развитие нервной системы
Развитие нервной системы
 Классификация НС
Классификация НС
 Разновидности нейронов
Разновидности нейронов
 Разновидности нейронов
Разновидности нейронов
 Строение нейрона
Строение нейрона
 Нейрон • Нейрофибриллы формируют цитоскелет клетки. • Дендриты входная структура, получают сигналы, поступающие к нейрону, и проводят их к телу клетки. • Аксон – проводит сигнал от тела нервной клетки. • В аксоне белок не синтезируется. • В области аксонного холмика генерируется ПД. • Аксоны заканчиваются терминалями аксона (до 2000), в которых имеются синаптические пузырьки, содержащие медиатор.
Нейрон • Нейрофибриллы формируют цитоскелет клетки. • Дендриты входная структура, получают сигналы, поступающие к нейрону, и проводят их к телу клетки. • Аксон – проводит сигнал от тела нервной клетки. • В аксоне белок не синтезируется. • В области аксонного холмика генерируется ПД. • Аксоны заканчиваются терминалями аксона (до 2000), в которых имеются синаптические пузырьки, содержащие медиатор.
 Нейрон и его компоненты
Нейрон и его компоненты
 Особенности метаболизма нейронов ◘ высокое потребление О 2: полная гипоксия в течение 5 6 минут ведет к гибели клеток коры ◘ способность к альтернативным путям обмена ◘ способность к созданию крупных запасов веществ ◘ нервная клетка живет только вместе с глией ◘ способность к регенерации отростков (0, 5 4 мк/сут. )
Особенности метаболизма нейронов ◘ высокое потребление О 2: полная гипоксия в течение 5 6 минут ведет к гибели клеток коры ◘ способность к альтернативным путям обмена ◘ способность к созданию крупных запасов веществ ◘ нервная клетка живет только вместе с глией ◘ способность к регенерации отростков (0, 5 4 мк/сут. )
 Функции нейрона • Нейрон – функциональная единица нервной системы. – Тело клетки выполняет функцию сумматора – Аксон проводит нервный импульс к другим клеткам – Дендриты воспринимают возбуждающие и тормозные влияния от других нейронов
Функции нейрона • Нейрон – функциональная единица нервной системы. – Тело клетки выполняет функцию сумматора – Аксон проводит нервный импульс к другим клеткам – Дендриты воспринимают возбуждающие и тормозные влияния от других нейронов
 Виды активности нейронов • Нервные клетки ЦНС делятся на: • ФОНОВОАКТИВНЫЕ – разряжаются импульсами без раздражения, спонтанно (например, сенсорные нейроны вестибулярной системы); такая активность называется фоновой. • Фоновоактивные нейроны делятся на • тормозящиеся урежающие частоту разрядов в ответ на сигнал, • возбуждающиеся учащающие частоту разрядов в ответ на какое либо раздражение. • МОЛЧАЩИЕ это нейроны, разряжающиеся импульсами только в ответ на какое либо раздражение.
Виды активности нейронов • Нервные клетки ЦНС делятся на: • ФОНОВОАКТИВНЫЕ – разряжаются импульсами без раздражения, спонтанно (например, сенсорные нейроны вестибулярной системы); такая активность называется фоновой. • Фоновоактивные нейроны делятся на • тормозящиеся урежающие частоту разрядов в ответ на сигнал, • возбуждающиеся учащающие частоту разрядов в ответ на какое либо раздражение. • МОЛЧАЩИЕ это нейроны, разряжающиеся импульсами только в ответ на какое либо раздражение.
 Виды активности нейронов • Фоновоактивные нейроны участвуют в поддержании тонуса коры и других структур мозга, в повышении чувствительности нейронов к раздражению, так как даже самые слабые воздействия изменяют частоту следования импульсов. Таким способом нейрон сигнализирует о приходе к нему возбуждающих или тормозящих сигналов. • Частота нервных импульсов в фоновоактивных нейронах может увеличиваться или уменьшаться в зависимости от интенсивности действующего на вторичные рецепторы стимула; кроме того, она может определяться направлением, в котором отклоняются чувствительные волоски механорецепторов. • Например, отклонение волосков вторичных механорецепторов в одну сторону сопровождается повышением фоновой активности сенсорного нейрона, которому они принадлежат, а в противоположную сторону понижением его фоновой активности. • Этот способ рецепции позволяет получать информацию об интенсивности стимула и о направлении, в котором он действует.
Виды активности нейронов • Фоновоактивные нейроны участвуют в поддержании тонуса коры и других структур мозга, в повышении чувствительности нейронов к раздражению, так как даже самые слабые воздействия изменяют частоту следования импульсов. Таким способом нейрон сигнализирует о приходе к нему возбуждающих или тормозящих сигналов. • Частота нервных импульсов в фоновоактивных нейронах может увеличиваться или уменьшаться в зависимости от интенсивности действующего на вторичные рецепторы стимула; кроме того, она может определяться направлением, в котором отклоняются чувствительные волоски механорецепторов. • Например, отклонение волосков вторичных механорецепторов в одну сторону сопровождается повышением фоновой активности сенсорного нейрона, которому они принадлежат, а в противоположную сторону понижением его фоновой активности. • Этот способ рецепции позволяет получать информацию об интенсивности стимула и о направлении, в котором он действует.
 Виды фоновой активности • ПАЧЕЧНЫЙ характеризуется появлением группы импульсов с коротким межимпульсным интервалом, после чего наступает период молчания, и вновь возникает пачка импульсов. Межимпульсные интервалы в таких пачках приблизительно равны 1 3 мс. • ГРУППОВОЙ характеризуется апериодическим появлением в фоне группы импульсов, межимпульсные интервалы в этих группах могут быть разные от 3 до 30 мс. • НЕПРЕРЫВНО АРИТМИЧНЫЙ характеризуется наличием в фоне возникающих через близкие, но разные промежутки времени импульсных разрядов. • Нейроны могут реагировать: • на включение раздражения «оn» нейроны, • на выключение раздражения «оff» нейроны, • на включение и выключение раздражения «оn off» нейроны.
Виды фоновой активности • ПАЧЕЧНЫЙ характеризуется появлением группы импульсов с коротким межимпульсным интервалом, после чего наступает период молчания, и вновь возникает пачка импульсов. Межимпульсные интервалы в таких пачках приблизительно равны 1 3 мс. • ГРУППОВОЙ характеризуется апериодическим появлением в фоне группы импульсов, межимпульсные интервалы в этих группах могут быть разные от 3 до 30 мс. • НЕПРЕРЫВНО АРИТМИЧНЫЙ характеризуется наличием в фоне возникающих через близкие, но разные промежутки времени импульсных разрядов. • Нейроны могут реагировать: • на включение раздражения «оn» нейроны, • на выключение раздражения «оff» нейроны, • на включение и выключение раздражения «оn off» нейроны.
 НЕЙРОГЛИЯ: обеспечивает поддержку, питание нейронов и выполняет защитные функции. Шванновские клетки: формируют миелиновые оболочки в ПНС. Миелин изолирует нервные волокна, увеличивает скорость передачи импульсов по нерву. • Олигодендроциты: формируют миелиновые оболочки в ЦНС. • Астроциты: регулируют концентрацию К+. • Микроглия: макрофаги ЦНС. • Эпендимальные клетки
НЕЙРОГЛИЯ: обеспечивает поддержку, питание нейронов и выполняет защитные функции. Шванновские клетки: формируют миелиновые оболочки в ПНС. Миелин изолирует нервные волокна, увеличивает скорость передачи импульсов по нерву. • Олигодендроциты: формируют миелиновые оболочки в ЦНС. • Астроциты: регулируют концентрацию К+. • Микроглия: макрофаги ЦНС. • Эпендимальные клетки
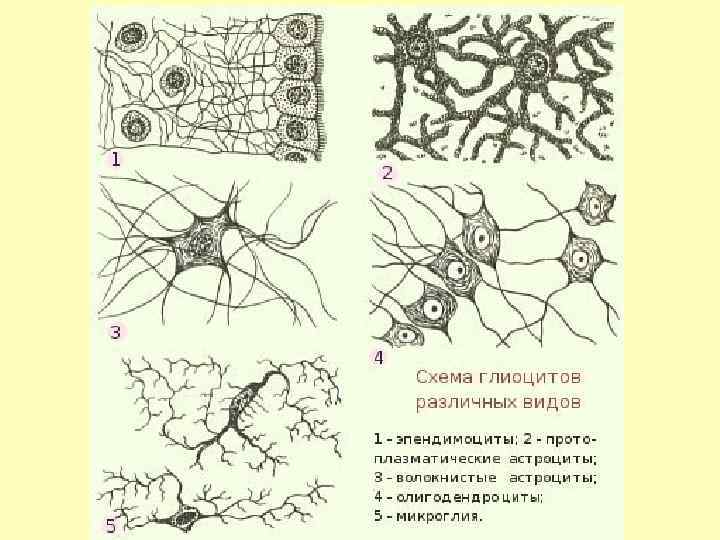
 • Распределение клеток нейроглии
• Распределение клеток нейроглии
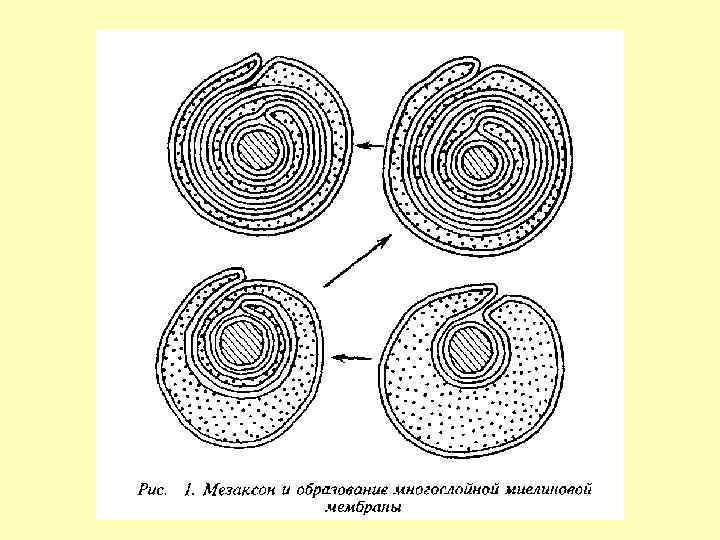
 Нейрон и миелиновая оболочка
Нейрон и миелиновая оболочка
 поступают") Аксональный транспорт • Многие ФАВ (медиаторы, факторы роста нервов, митохондрии, некоторые питательные вещества) поступают в нервные окончания путем аксонального транспорта. • Существует также ретроградный аксональный транспорт.
Аксональный транспорт • Многие ФАВ (медиаторы, факторы роста нервов, митохондрии, некоторые питательные вещества) поступают в нервные окончания путем аксонального транспорта. • Существует также ретроградный аксональный транспорт.
 Классификация нервных волокон по Гассеру Эрлангеру Тип волокна Миелиновая оболочка Средний диаметр, мкм Средняя скорость распространения возбуждения, м/с А альфа Есть 15 100 (двигательные во локна скелетных мышц) А бета Есть 8 50 (тактильная чувстви тельность) А гамма Есть 5 20 (двигательные волок на мышечных веретен) А А дельт а Есть <3 15 (ранний компонент боли) Группа В Есть <3 7 (преганглионарные симпатические волокна) Группа С Нет 1 1 Г Р У П П А (постганглионарные
Классификация нервных волокон по Гассеру Эрлангеру Тип волокна Миелиновая оболочка Средний диаметр, мкм Средняя скорость распространения возбуждения, м/с А альфа Есть 15 100 (двигательные во локна скелетных мышц) А бета Есть 8 50 (тактильная чувстви тельность) А гамма Есть 5 20 (двигательные волок на мышечных веретен) А А дельт а Есть <3 15 (ранний компонент боли) Группа В Есть <3 7 (преганглионарные симпатические волокна) Группа С Нет 1 1 Г Р У П П А (постганглионарные
 Проведение возбуждения в безмякотном нервном волокне
Проведение возбуждения в безмякотном нервном волокне
 Проведение возбуждения в мякотном нервном волокне
Проведение возбуждения в мякотном нервном волокне
 Законы проведения возбуждения по нервному волокну • Закон анатомо-физиологической целостности • Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. • При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Законы проведения возбуждения по нервному волокну • Закон анатомо-физиологической целостности • Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. • При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
 Законы проведения возбуждения по нервному волокну • Закон изолированного проведения возбуждения • В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе. • В мякотных НВ роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической ёмкости оболочки. • В безмякотных НВ возбуждение передается также изолированно, так как сопротивление жидкости в межклеточных щелях значительно ниже сопротивления мембраны НВ и ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние НВ.
Законы проведения возбуждения по нервному волокну • Закон изолированного проведения возбуждения • В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе. • В мякотных НВ роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической ёмкости оболочки. • В безмякотных НВ возбуждение передается также изолированно, так как сопротивление жидкости в межклеточных щелях значительно ниже сопротивления мембраны НВ и ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние НВ.
 Законы проведения возбуждения по нервному волокну • Закон двустороннего проведения возбуждения • Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно. • В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость НВ ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
Законы проведения возбуждения по нервному волокну • Закон двустороннего проведения возбуждения • Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно. • В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость НВ ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
 Конус роста При регенерации поврежденного отростка нейрона образуется конус роста волокна утолщение со множеством длинных и тонких отростков толщиной 0, 1 0, 2 мкм и длиной до 50 мкм, отходящих в разные стороны. Конус роста является зоной интенсивного экзо и эндоцитоза. Мембранный материал для регенерации образуется в теле нейрона и переносится быстрым транспортом в виде пузырьков к конусу роста и посредством экзоцитоза встраивается в клеточную мембрану, удлиняя ее. Для передвижения конуса роста необходимы актиновые филаменты, разрушение которых (например, цитохолазином В) прекращает рост.
Конус роста При регенерации поврежденного отростка нейрона образуется конус роста волокна утолщение со множеством длинных и тонких отростков толщиной 0, 1 0, 2 мкм и длиной до 50 мкм, отходящих в разные стороны. Конус роста является зоной интенсивного экзо и эндоцитоза. Мембранный материал для регенерации образуется в теле нейрона и переносится быстрым транспортом в виде пузырьков к конусу роста и посредством экзоцитоза встраивается в клеточную мембрану, удлиняя ее. Для передвижения конуса роста необходимы актиновые филаменты, разрушение которых (например, цитохолазином В) прекращает рост.
 Утомление нерва • Нервные волокна мало утомляемы из за низкого расхода энергии и интенсивного ресинтеза АТФ и креатинфосфата. • В опытах Н. Е. Введенского (1884) нерв подвергался непрерывному раздражению индукционным током многие часы без заметного ослабления действия нерва на мышцу. • В целом организме малой утомляемости нервных волокон способствует то, что частота импульсации из нервных центров (50 100 имп. /с) на периферию значительно ниже предельных возможностей волокон генерировать импульсы, т. е. лабильность нервных волокон значительно выше лабильности тела нервной клетки.
Утомление нерва • Нервные волокна мало утомляемы из за низкого расхода энергии и интенсивного ресинтеза АТФ и креатинфосфата. • В опытах Н. Е. Введенского (1884) нерв подвергался непрерывному раздражению индукционным током многие часы без заметного ослабления действия нерва на мышцу. • В целом организме малой утомляемости нервных волокон способствует то, что частота импульсации из нервных центров (50 100 имп. /с) на периферию значительно ниже предельных возможностей волокон генерировать импульсы, т. е. лабильность нервных волокон значительно выше лабильности тела нервной клетки.
 Виды взаимодействия нейронов • ДИСТАНТНОЕ взаимодействие обеспечивается двумя нейронами, расположенными в разных структурах организма, посредством синтезируемых ими и выделяемых в интерстициальную жидкость веществ. • Пример: в клетках ряда структур мозга образуются нейрогормоны, нейропептиды, которые способны воздействовать гуморально на нейроны других отделов.
Виды взаимодействия нейронов • ДИСТАНТНОЕ взаимодействие обеспечивается двумя нейронами, расположенными в разных структурах организма, посредством синтезируемых ими и выделяемых в интерстициальную жидкость веществ. • Пример: в клетках ряда структур мозга образуются нейрогормоны, нейропептиды, которые способны воздействовать гуморально на нейроны других отделов.
 Виды взаимодействия нейронов • СМЕЖНОЕ соединение – между нейронами, расположенными в непосредственной близости, разделенными межклеточным пространством (≈20 нм). • Это облегчает перемещение из одной клетки в межклеточную щель химических веществ (ионов, метаболитов нейронов), которые оказывают влияние как на ту же самую клетку, так и на отростки соседних нейронов, что обеспечивает участие соседних нейронов в выполнении единой функции. • Смежное взаимодействие может в ряде случаев обеспечивать передачу электрической информации от нейрона к нейрону.
Виды взаимодействия нейронов • СМЕЖНОЕ соединение – между нейронами, расположенными в непосредственной близости, разделенными межклеточным пространством (≈20 нм). • Это облегчает перемещение из одной клетки в межклеточную щель химических веществ (ионов, метаболитов нейронов), которые оказывают влияние как на ту же самую клетку, так и на отростки соседних нейронов, что обеспечивает участие соседних нейронов в выполнении единой функции. • Смежное взаимодействие может в ряде случаев обеспечивать передачу электрической информации от нейрона к нейрону.
 Виды взаимодействия нейронов • КОНТАКТНОЕ взаимодействие обусловлено специфическими контактами мембран нейронов, которые образуют электрические и химические синапсы. • химические синапсы - передача сигнала происходит с помощью химического вещества – медиатора): • между нейронами – межнейронные, • между нейроном и клеткой внутреннего органа – нейроэффекторные, • между нейроном и клеткой скелетной мышцы – нервно-мышечные); • электрические синапсы мембраны клеток тесно контактируют, передача сигнала происходит с помощью местных токов.
Виды взаимодействия нейронов • КОНТАКТНОЕ взаимодействие обусловлено специфическими контактами мембран нейронов, которые образуют электрические и химические синапсы. • химические синапсы - передача сигнала происходит с помощью химического вещества – медиатора): • между нейронами – межнейронные, • между нейроном и клеткой внутреннего органа – нейроэффекторные, • между нейроном и клеткой скелетной мышцы – нервно-мышечные); • электрические синапсы мембраны клеток тесно контактируют, передача сигнала происходит с помощью местных токов.
 Синапсы на теле нейрона Число синапсов на нейроне сильно варьирует: от нескольких в ядрах среднего мозга до 10 000 на мотонейронах спинного мозга. В сенсорной коре до 98 % синапсов локализованы на дендритах и только 2 % - на теле нейрона.
Синапсы на теле нейрона Число синапсов на нейроне сильно варьирует: от нескольких в ядрах среднего мозга до 10 000 на мотонейронах спинного мозга. В сенсорной коре до 98 % синапсов локализованы на дендритах и только 2 % - на теле нейрона.
 центральные синапсы; • лежат в пределах") Классификация синапсов • I. По локализации: • 1) центральные синапсы; • лежат в пределах ЦНС, а также находятся в ганглиях ВНС; это контакты между двумя нервными клетками. Различают синапсы: • аксосоматический, образованный аксоном одного нейрона и телом другого нейрона; • аксодендритный, образованный аксоном одного нейрона и дендритом другого; • аксональный (аксон первого нейрона образует синапс на аксоне второго нейрона); • дендродентритный (дендрит первого нейрона образует синапс на дендрите второго нейрона).
Классификация синапсов • I. По локализации: • 1) центральные синапсы; • лежат в пределах ЦНС, а также находятся в ганглиях ВНС; это контакты между двумя нервными клетками. Различают синапсы: • аксосоматический, образованный аксоном одного нейрона и телом другого нейрона; • аксодендритный, образованный аксоном одного нейрона и дендритом другого; • аксональный (аксон первого нейрона образует синапс на аксоне второго нейрона); • дендродентритный (дендрит первого нейрона образует синапс на дендрите второго нейрона).
 периферические синапсы: • мионевральный (нервно мышечный),") Классификация синапсов • 1. По локализации: • 2) периферические синапсы: • мионевральный (нервно мышечный), образованный аксоном мотонейрона и мышечной клеткой; • нервно эпителиальный, образованный аксоном нейрона и секреторной клеткой.
Классификация синапсов • 1. По локализации: • 2) периферические синапсы: • мионевральный (нервно мышечный), образованный аксоном мотонейрона и мышечной клеткой; • нервно эпителиальный, образованный аксоном нейрона и секреторной клеткой.
 Классификация синапсов • II. Функциональная классификация синапсов: • возбуждающие синапсы; • тормозящие синапсы. • III. По механизмам передачи возбуждения в синапсах: • химические; • электрические.
Классификация синапсов • II. Функциональная классификация синапсов: • возбуждающие синапсы; • тормозящие синапсы. • III. По механизмам передачи возбуждения в синапсах: • химические; • электрические.
 Виды синапсов • Электрический синапс • Химический синапс
Виды синапсов • Электрический синапс • Химический синапс
 Виды синапсов
Виды синапсов
 ХИМИЧЕСКИЙ СИНАПС • контакт между двумя нейронами • обязателен химический посредник • на одном нейроне может быть до 10 000 синапсов • на теле нейрона осуществляется временная и пространственная суммация • результат может быть возбуждающим или тормозным
ХИМИЧЕСКИЙ СИНАПС • контакт между двумя нейронами • обязателен химический посредник • на одном нейроне может быть до 10 000 синапсов • на теле нейрона осуществляется временная и пространственная суммация • результат может быть возбуждающим или тормозным
; • адренэргические (м. катехоламины); • дофаминэргические (м.") Классификация химических синапсов • холинэргические (м. ацетилхолин); • адренэргические (м. катехоламины); • дофаминэргические (м. дофамин); • гистаминэргические (м. гистамин); • ГАМКэргические (м. гаммааминомасляная тормозный синапс). ГАМК кислота;
Классификация химических синапсов • холинэргические (м. ацетилхолин); • адренэргические (м. катехоламины); • дофаминэргические (м. дофамин); • гистаминэргические (м. гистамин); • ГАМКэргические (м. гаммааминомасляная тормозный синапс). ГАМК кислота;
 Синапс в ЦНС
Синапс в ЦНС
 Схема работы химического синапса
Схема работы химического синапса
 Этапы передачи сигнала в химическом синапсе • 1. В пресинаптическое окончание поступает ПД. • 2. ПД вызывает открывание потенциалчувствительных кальциевых каналов в мембране пресинаптического окончания. • 3. Через кальциевые каналы в пресинаптическое окончание входит Са++.
Этапы передачи сигнала в химическом синапсе • 1. В пресинаптическое окончание поступает ПД. • 2. ПД вызывает открывание потенциалчувствительных кальциевых каналов в мембране пресинаптического окончания. • 3. Через кальциевые каналы в пресинаптическое окончание входит Са++.
 Этапы передачи сигнала в химическом синапсе • 4. Са++ активирует белки, отвечающие за слияние содержащих медиатор пузырьков с пресинаптической мембраной. • 5. Слившись с мембраной, пузырьки посредством экзоцитоза высвобождают медиатор в синаптическую щель. • 6. Молекулы медиатора взаимодействуют с постсинаптическими рецепторами и активируют их.
Этапы передачи сигнала в химическом синапсе • 4. Са++ активирует белки, отвечающие за слияние содержащих медиатор пузырьков с пресинаптической мембраной. • 5. Слившись с мембраной, пузырьки посредством экзоцитоза высвобождают медиатор в синаптическую щель. • 6. Молекулы медиатора взаимодействуют с постсинаптическими рецепторами и активируют их.
 Fast small molecules (e. g. acetylcholine) ligand gated ion") Химические синапсы • Chemical (neurotransmitter) Fast small molecules (e. g. acetylcholine) ligand gated ion channels Slow neurotransmitters synthesised from aa’s (e. g. noradrenaline) G protein signalling pathway Fig. 6 -11
Химические синапсы • Chemical (neurotransmitter) Fast small molecules (e. g. acetylcholine) ligand gated ion channels Slow neurotransmitters synthesised from aa’s (e. g. noradrenaline) G protein signalling pathway Fig. 6 -11
 Взаимодействие нейротрансмиттера с рецептором • Модель ключ-замок: • Нейротрансмиттер • связывается с рецептором • НТ = ключ • Рецептор = замок Рецептор изменяет структуру и открывает ионные каналы, что приводит к генерации ВПСП или ТПСП или активирует ферменты.
Взаимодействие нейротрансмиттера с рецептором • Модель ключ-замок: • Нейротрансмиттер • связывается с рецептором • НТ = ключ • Рецептор = замок Рецептор изменяет структуру и открывает ионные каналы, что приводит к генерации ВПСП или ТПСП или активирует ферменты.
 Быстрая и медленная химическая синаптическая передача • БЫСТРАЯ • МЕДЛЕННАЯ СИНАПТИЧЕСКАЯ ТРАНСМИССИЯ • Нейротрансмиттер выделяется и связывается с рецептором и хемочувствительным активирует ионным каналом. высвобождение вторичного • Ответ регистрируется мессенджера в через несколько постсинаптической миллисекунд. клетке. • Ответ регистрируется через несколько секунд или минут.
Быстрая и медленная химическая синаптическая передача • БЫСТРАЯ • МЕДЛЕННАЯ СИНАПТИЧЕСКАЯ ТРАНСМИССИЯ • Нейротрансмиттер выделяется и связывается с рецептором и хемочувствительным активирует ионным каналом. высвобождение вторичного • Ответ регистрируется мессенджера в через несколько постсинаптической миллисекунд. клетке. • Ответ регистрируется через несколько секунд или минут.
 • Быстрый химический синапс – НТ – ацетилхолин – Рецептор связан с лиганд управляемым ионным каналом (Na+, K+) – Происходит ферментативная инактивация НТ
• Быстрый химический синапс – НТ – ацетилхолин – Рецептор связан с лиганд управляемым ионным каналом (Na+, K+) – Происходит ферментативная инактивация НТ
 Медленная синаптическая передача li Нейротрансмиттер drenalin receptor Рецептор OUT MEMBRANE a IN Посредник G protein Усилитель adenylate cyclase c. AMP Вторичный мессенджер
Медленная синаптическая передача li Нейротрансмиттер drenalin receptor Рецептор OUT MEMBRANE a IN Посредник G protein Усилитель adenylate cyclase c. AMP Вторичный мессенджер
 Инактивация медиатора • Диффузия из синаптической щели в окружающие ткани. • Обратный захват пресинаптическим окончанием. • Ферментативный распад.
Инактивация медиатора • Диффузия из синаптической щели в окружающие ткани. • Обратный захват пресинаптическим окончанием. • Ферментативный распад.
 Диффузия • Самый простой способ, не требующий ферментов и белков переносчиков. • Самый медленный способ, позволяющий медиатору действовать долго, но тем самым делающий невозможным передачу частых сигналов. • Позволяет медиатору действовать сразу на много клеток, но тем самым делает невозможной точную прицельную регуляцию.
Диффузия • Самый простой способ, не требующий ферментов и белков переносчиков. • Самый медленный способ, позволяющий медиатору действовать долго, но тем самым делающий невозможным передачу частых сигналов. • Позволяет медиатору действовать сразу на много клеток, но тем самым делает невозможной точную прицельную регуляцию.
 Обратный захват • Требует специализированных белков, обеспечивающих перенос медиатора в пресинаптическое окончание, а затем – в пузырьки. • Существенно быстрее диффузии. • Экономичен – позволяет многократно использовать медиатор. • Может регулироваться, что позволяет изменять эффективность синаптической передачи.
Обратный захват • Требует специализированных белков, обеспечивающих перенос медиатора в пресинаптическое окончание, а затем – в пузырьки. • Существенно быстрее диффузии. • Экономичен – позволяет многократно использовать медиатор. • Может регулироваться, что позволяет изменять эффективность синаптической передачи.
 Ферментативный распад • Требует ферментов, отвечающих за разрушение медиатора. • Самый быстрый способ, обеспечивающий передачу импульсов высокой частоты. • Пример: передача возбуждения с нейрона на скелетную мышцу.
Ферментативный распад • Требует ферментов, отвечающих за разрушение медиатора. • Самый быстрый способ, обеспечивающий передачу импульсов высокой частоты. • Пример: передача возбуждения с нейрона на скелетную мышцу.
 Физиологические свойства синапсов • ОДНОСТОРОННЕЕ ПРОВЕДЕНИЕ: обусловлено направлением потока медиатора – он выделяется из пресинаптического окончания и действует на постсинаптические рецепторы. • СИНАПТИЧЕСКАЯ ЗАДЕРЖКА: обусловлена сравнительно длительным временем, необходимым для выделения медиатора, его диффузии к рецепторам, активации рецепторов и последующих постсинаптических процессов.
Физиологические свойства синапсов • ОДНОСТОРОННЕЕ ПРОВЕДЕНИЕ: обусловлено направлением потока медиатора – он выделяется из пресинаптического окончания и действует на постсинаптические рецепторы. • СИНАПТИЧЕСКАЯ ЗАДЕРЖКА: обусловлена сравнительно длительным временем, необходимым для выделения медиатора, его диффузии к рецепторам, активации рецепторов и последующих постсинаптических процессов.
 Физиологические свойства синапсов • НИЗКАЯ ЛАБИЛЬНОСТЬ • Мерой лабильности служит максимальная частота импульсов, которую может воспроизвести ткань. • В НВ лабильность ограничена временем периода рефрактерности; этот период короткий (около 1 мс), лабильность НВ высока. • Синапсы становятся готовы к проведению очередного сигнала лишь после того, как будет инактивирована очередная порция медиатора, на что требуется достаточно много времени. • Таким образом, лабильность синапса ограничена временем полного оборота медиатора (медиаторного цикла) и синапсы могут проводить лишь импульсы низкой частоты, то есть обладают низкой лабильностью.
Физиологические свойства синапсов • НИЗКАЯ ЛАБИЛЬНОСТЬ • Мерой лабильности служит максимальная частота импульсов, которую может воспроизвести ткань. • В НВ лабильность ограничена временем периода рефрактерности; этот период короткий (около 1 мс), лабильность НВ высока. • Синапсы становятся готовы к проведению очередного сигнала лишь после того, как будет инактивирована очередная порция медиатора, на что требуется достаточно много времени. • Таким образом, лабильность синапса ограничена временем полного оборота медиатора (медиаторного цикла) и синапсы могут проводить лишь импульсы низкой частоты, то есть обладают низкой лабильностью.
 Физиологические свойства синапсов • ВЫСОКАЯ УТОМЛЯЕМОСТЬ: заключается в снижении величины реакции клетки при длительном раздражении. • Причина – исчерпание ресурсов клетки, накопление метаболитов и пр. • В нервных клетках теоретически при чрезвычайно длительном раздражении могут выравниваться концентрации ионов во внутренней и наружной среде, но практически это невозможно благодаря работе ионных насосов. • В синапсе при длительном раздражении могут истощаться запасы медиатора, и тогда развивается утомление.
Физиологические свойства синапсов • ВЫСОКАЯ УТОМЛЯЕМОСТЬ: заключается в снижении величины реакции клетки при длительном раздражении. • Причина – исчерпание ресурсов клетки, накопление метаболитов и пр. • В нервных клетках теоретически при чрезвычайно длительном раздражении могут выравниваться концентрации ионов во внутренней и наружной среде, но практически это невозможно благодаря работе ионных насосов. • В синапсе при длительном раздражении могут истощаться запасы медиатора, и тогда развивается утомление.
 Регуляция синаптической передачи • на пресинаптическом уровне сводится к изменению: • количества медиатора в синаптической щели достигается за счет регуляции высвобождения медиатора; • времени пребывания медиатора в синаптической щели достигается за счет регуляции обратного захвата медиатора.
Регуляция синаптической передачи • на пресинаптическом уровне сводится к изменению: • количества медиатора в синаптической щели достигается за счет регуляции высвобождения медиатора; • времени пребывания медиатора в синаптической щели достигается за счет регуляции обратного захвата медиатора.
 Типы хемочувствительных ионных каналов на постсинаптической мембране • A. Контакт нейротрансмиттера с рецептором приводит к открытию хемочувствительного ионного канала B. Изучены 4 типа хемочувствительных ионных каналов: a. Na+ каналы b. Ca+ каналы • c. K+ каналы d. Cl каналы
Типы хемочувствительных ионных каналов на постсинаптической мембране • A. Контакт нейротрансмиттера с рецептором приводит к открытию хемочувствительного ионного канала B. Изучены 4 типа хемочувствительных ионных каналов: a. Na+ каналы b. Ca+ каналы • c. K+ каналы d. Cl каналы
 Постсинаптические потенциалы ТПСП Мембранный потенциал Время ВПСП
Постсинаптические потенциалы ТПСП Мембранный потенциал Время ВПСП
 Возбуждающий постсинаптический потенциал • Волна деполяризации, которая приближает МП к КУД, облегчая возможность генерации ПД.
Возбуждающий постсинаптический потенциал • Волна деполяризации, которая приближает МП к КУД, облегчая возможность генерации ПД.
 Тормозный постсинаптический потенциал • Волна гиперполяризации, которая увеличивает величину МП и уменьшает вероятность генерации ПД.
Тормозный постсинаптический потенциал • Волна гиперполяризации, которая увеличивает величину МП и уменьшает вероятность генерации ПД.
 Постсинаптическая передача сигнала • Постсинаптические рецепторы делятся на 2 группы: • рецепторы, сопряженные с ионными каналами – ионотропные рецепторы; • рецепторы, сопряженные с внутриклеточными ферментативными системами – метаботропные рецепторы.
Постсинаптическая передача сигнала • Постсинаптические рецепторы делятся на 2 группы: • рецепторы, сопряженные с ионными каналами – ионотропные рецепторы; • рецепторы, сопряженные с внутриклеточными ферментативными системами – метаботропные рецепторы.
 Нервно-мышечный синапс • Образован окончанием аксона двигательного нейрона на скелетной мышце. • Пресинаптическое окончание нейрона ветвится, образуя синапс очень больших размеров. • Поверхность постсинаптической мембраны дополнительно увеличена за счет многочисленных складок. • Медиатор – ацетилхолин. • Рецепторы АХ в нервно мышечном синапсе относятся к типу N холинорецепторов (чувствительных к никотину). Это ионотропные рецепторы; с ними связаны ионные каналы, пропускающие катионы.
Нервно-мышечный синапс • Образован окончанием аксона двигательного нейрона на скелетной мышце. • Пресинаптическое окончание нейрона ветвится, образуя синапс очень больших размеров. • Поверхность постсинаптической мембраны дополнительно увеличена за счет многочисленных складок. • Медиатор – ацетилхолин. • Рецепторы АХ в нервно мышечном синапсе относятся к типу N холинорецепторов (чувствительных к никотину). Это ионотропные рецепторы; с ними связаны ионные каналы, пропускающие катионы.
 Нервно-мышечный синапс • При активации N холинорецепторов каналы открываются, в мышечную клетку входит Nа+ (каналы пропускают все катионы, но из всех катионов наибольшим электрохимическим градиентом обладает Nа+ , и этот градиент направлен внутрь клетки) и возникает ВПСП, называемый в данном синапсе потенциалом концевой пластинки. • Потенциал концевой пластинки всегда сверхпороговый; это обусловлено большими размерами синапса и одновременным выбросом большого количества медиатора. • Т. о. , один ПД в нервном окончании всегда вызывает один ПД в скелетной мышце.
Нервно-мышечный синапс • При активации N холинорецепторов каналы открываются, в мышечную клетку входит Nа+ (каналы пропускают все катионы, но из всех катионов наибольшим электрохимическим градиентом обладает Nа+ , и этот градиент направлен внутрь клетки) и возникает ВПСП, называемый в данном синапсе потенциалом концевой пластинки. • Потенциал концевой пластинки всегда сверхпороговый; это обусловлено большими размерами синапса и одновременным выбросом большого количества медиатора. • Т. о. , один ПД в нервном окончании всегда вызывает один ПД в скелетной мышце.
 Нервно-мышечный синапс
Нервно-мышечный синапс
 Нервно-мышечный синапс • Инактивация ацетилхолина происходит в синаптической щели с помощью фермента ацетилхолинэстеразы. • Эта инактивация осуществляется чрезвычайно быстро, поэтому нервно мышечный синапс способен проводить импульсы высокой частоты.
Нервно-мышечный синапс • Инактивация ацетилхолина происходит в синаптической щели с помощью фермента ацетилхолинэстеразы. • Эта инактивация осуществляется чрезвычайно быстро, поэтому нервно мышечный синапс способен проводить импульсы высокой частоты.
 Нервно мышечная передача в гладких мышцах • Обязательное условие возникновения сокращение гладких мышц, как и скелетных, увеличение концентрации Са 2+ в миоплазме. • Считается, что процесс сокращения активизируется преимущественно внеклеточным Са 2+, поступающим в мышечные клетки через потенциалзависимые Са 2+ каналы. • Особенность в том, что иннервация осуществляется вегетативной нервной системой и она может оказывать как возбуждающее, так и тормозящее влияние.
Нервно мышечная передача в гладких мышцах • Обязательное условие возникновения сокращение гладких мышц, как и скелетных, увеличение концентрации Са 2+ в миоплазме. • Считается, что процесс сокращения активизируется преимущественно внеклеточным Са 2+, поступающим в мышечные клетки через потенциалзависимые Са 2+ каналы. • Особенность в том, что иннервация осуществляется вегетативной нервной системой и она может оказывать как возбуждающее, так и тормозящее влияние.
 и") Нервно мышечная передача в гладких мышцах • По типу различают холинергические (м. АХ) и адренергические (м. НА) синапсы. Первые обычно содержатся в мышцах пищеварительной системы, вторые в мышцах кровеносных сосудов. • Один и тот же медиатор в одних синапсах может быть возбуждающим, а в других тормозным (в зависимости от свойств циторецепторов). • Адренорецепторы делят на α и β. НА, воздействуя на α адренорецепторы, суживает кровеносные сосуды и тормозит моторику пищеварительного тракта, а воздействуя на β адренорецепторы, стимулирует деятельность сердца и расширяет кровеносные сосуды некоторых органов, расслабляет мышцы бронхов.
Нервно мышечная передача в гладких мышцах • По типу различают холинергические (м. АХ) и адренергические (м. НА) синапсы. Первые обычно содержатся в мышцах пищеварительной системы, вторые в мышцах кровеносных сосудов. • Один и тот же медиатор в одних синапсах может быть возбуждающим, а в других тормозным (в зависимости от свойств циторецепторов). • Адренорецепторы делят на α и β. НА, воздействуя на α адренорецепторы, суживает кровеносные сосуды и тормозит моторику пищеварительного тракта, а воздействуя на β адренорецепторы, стимулирует деятельность сердца и расширяет кровеносные сосуды некоторых органов, расслабляет мышцы бронхов.
 Нервно мышечная передача в гладких мышцах • В ответ на действие возбуждающего медиатора происходит деполяризация клеток гладких мышц, которая проявляется в виде возбуждающего синаптического потенциала (ВСП). Когда он достигает критического уровня, возникает ПД. Это происходит тогда, когда до нервного окончания друг за другом доходят несколько импульсов. • Возникновение ВСП является следствием увеличения проницаемости постсинаптической мембраны для Na+, Са 2+.
Нервно мышечная передача в гладких мышцах • В ответ на действие возбуждающего медиатора происходит деполяризация клеток гладких мышц, которая проявляется в виде возбуждающего синаптического потенциала (ВСП). Когда он достигает критического уровня, возникает ПД. Это происходит тогда, когда до нервного окончания друг за другом доходят несколько импульсов. • Возникновение ВСП является следствием увеличения проницаемости постсинаптической мембраны для Na+, Са 2+.
 Нервно мышечная передача в гладких мышцах • Тормозной медиатор вызывает гиперполяризацию постсинаптической мембраны, что проявляется в тормозном синаптическом потенциале (ТСП). • В основе гиперполяризации лежит повышение проницаемости мембраны в основном для К+. • Роль тормозного медиатора в гладких мышцах, возбуждаемых АХ (например, мышцы кишки, бронхов), играет НА, а в гладких мышцах, для которых возбуждающих медиатором является НА (например, мышцы мочевого пузыря), АХ.
Нервно мышечная передача в гладких мышцах • Тормозной медиатор вызывает гиперполяризацию постсинаптической мембраны, что проявляется в тормозном синаптическом потенциале (ТСП). • В основе гиперполяризации лежит повышение проницаемости мембраны в основном для К+. • Роль тормозного медиатора в гладких мышцах, возбуждаемых АХ (например, мышцы кишки, бронхов), играет НА, а в гладких мышцах, для которых возбуждающих медиатором является НА (например, мышцы мочевого пузыря), АХ.
 Причины нарушения проведения нервного импульса в синапсе • Истощение запасов медиатора • Инактивация рецепторов на постсинаптической мембране • Стойкая деполяризация постсинаптической мембраны
Причины нарушения проведения нервного импульса в синапсе • Истощение запасов медиатора • Инактивация рецепторов на постсинаптической мембране • Стойкая деполяризация постсинаптической мембраны
 Способы нарушения проведения нервного импульса в синапсе • Нарушение поступления ионов кальция в пресинаптическое окончание • Связывание медиатора в синаптической щели • Конкурентное связывание рецепторов на постсинаптической мембране • Стойкая деполяризация постсинаптической мембраны • Разрушение фермента, осуществляющего ферментативный распад медиатора
Способы нарушения проведения нервного импульса в синапсе • Нарушение поступления ионов кальция в пресинаптическое окончание • Связывание медиатора в синаптической щели • Конкурентное связывание рецепторов на постсинаптической мембране • Стойкая деполяризация постсинаптической мембраны • Разрушение фермента, осуществляющего ферментативный распад медиатора
 Механизм утомления синапса • связан главным образом с полным или частичным истощением запаса медиатора в пресинаптических терминалях. • Возбуждающие терминали на многих нейронах содержат только такое количество возбуждающего медиатора, которое необходимо для вызывания около 10000 ПД, поэтому при высокой частоте стимуляции медиатор может истощиться в течение нескольких секунд или минут. • ? частично процесс утомления связан также с действием двух других факторов: • (1) прогрессирующей инактивацией многих постсинаптических мембранных рецепторов; • (2) медленным изменением аномальных концентраций ионов в постсинаптической нервной клетке.
Механизм утомления синапса • связан главным образом с полным или частичным истощением запаса медиатора в пресинаптических терминалях. • Возбуждающие терминали на многих нейронах содержат только такое количество возбуждающего медиатора, которое необходимо для вызывания около 10000 ПД, поэтому при высокой частоте стимуляции медиатор может истощиться в течение нескольких секунд или минут. • ? частично процесс утомления связан также с действием двух других факторов: • (1) прогрессирующей инактивацией многих постсинаптических мембранных рецепторов; • (2) медленным изменением аномальных концентраций ионов в постсинаптической нервной клетке.
 Влияние ацидоза на синаптическую передачу • Большинство нейронов очень чувствительны к изменениям р. Н окружающей межклеточной жидкости. • Ацидоз очень угнетает активность нервной системы: • снижение р. Н от уровня 7, 4 до величин ниже 7, 0 обычно ведет к коматозному состоянию. • Например, при тяжелом диабетическом или уремическом ацидозе практически всегда развивается кома.
Влияние ацидоза на синаптическую передачу • Большинство нейронов очень чувствительны к изменениям р. Н окружающей межклеточной жидкости. • Ацидоз очень угнетает активность нервной системы: • снижение р. Н от уровня 7, 4 до величин ниже 7, 0 обычно ведет к коматозному состоянию. • Например, при тяжелом диабетическом или уремическом ацидозе практически всегда развивается кома.
 Влияние алкалоза на синаптическую передачу • Обычно алкалоз значительно увеличивает возбудимость нервной системы. Так, повышение р. Н артериальной крови от нормальной величины 7, 4 до 7, 8 8, 0 часто вызывает эпилептические приступы из за увеличенной возбудимости некоторых или всех мозговых нейронов. • Пример реакция человека, предрасположенного к эпилептическим приступам, на гипервентиляцию (усиленное дыхание в покое). • Гипервентиляция вымывает из крови СО 2, в результате на мгновение поднимается уровень р. Н крови, но даже это кратковременное изменение р. Н может спровоцировать эпилептический припадок.
Влияние алкалоза на синаптическую передачу • Обычно алкалоз значительно увеличивает возбудимость нервной системы. Так, повышение р. Н артериальной крови от нормальной величины 7, 4 до 7, 8 8, 0 часто вызывает эпилептические приступы из за увеличенной возбудимости некоторых или всех мозговых нейронов. • Пример реакция человека, предрасположенного к эпилептическим приступам, на гипервентиляцию (усиленное дыхание в покое). • Гипервентиляция вымывает из крови СО 2, в результате на мгновение поднимается уровень р. Н крови, но даже это кратковременное изменение р. Н может спровоцировать эпилептический припадок.
 Влияние гипоксии на синаптическую передачу • Возбудимость нервных структур во многом зависит от адекватного снабжения кислородом. • Прекращение доставки кислорода даже на несколько секунд может привести к полной потере возбудимости в некоторых нейронах. • Это наблюдают при временном прерывании мозгового кровотока, в этом случае через 3 7 с человек теряет сознание.
Влияние гипоксии на синаптическую передачу • Возбудимость нервных структур во многом зависит от адекватного снабжения кислородом. • Прекращение доставки кислорода даже на несколько секунд может привести к полной потере возбудимости в некоторых нейронах. • Это наблюдают при временном прерывании мозгового кровотока, в этом случае через 3 7 с человек теряет сознание.
 Влияние лекарств на синаптическую передачу • Среди лекарственных средств многие известны как повышающие возбудимость нейронов, а другие как снижающие их возбудимость. • Например, кофеин, теофиллин и теобромин, которые содержатся в кофе, чае и коке, соответственно, повышают нервную возбудимость, по видимому, путем снижения порога возбуждения нейронов.
Влияние лекарств на синаптическую передачу • Среди лекарственных средств многие известны как повышающие возбудимость нейронов, а другие как снижающие их возбудимость. • Например, кофеин, теофиллин и теобромин, которые содержатся в кофе, чае и коке, соответственно, повышают нервную возбудимость, по видимому, путем снижения порога возбуждения нейронов.
 Влияние лекарств на синаптическую передачу • Стрихнин один из самых известных агентов, увеличивающих возбудимость нейронов. • Стрихнин не уменьшает порог возбуждения нейронов, а подавляет действие некоторых тормозных медиаторов, особенно тормозной эффект глицина в спинном мозге. • При этом влияние возбуждающих медиаторов становится преобладающим, что резко повышает возбудимость нейронов, при этом они начинают генерировать частые разряды, сопровождаемые развитием сильных тонических мышечных спазмов.
Влияние лекарств на синаптическую передачу • Стрихнин один из самых известных агентов, увеличивающих возбудимость нейронов. • Стрихнин не уменьшает порог возбуждения нейронов, а подавляет действие некоторых тормозных медиаторов, особенно тормозной эффект глицина в спинном мозге. • При этом влияние возбуждающих медиаторов становится преобладающим, что резко повышает возбудимость нейронов, при этом они начинают генерировать частые разряды, сопровождаемые развитием сильных тонических мышечных спазмов.
 Влияние лекарств на синаптическую передачу • Большинство анестетиков увеличивают порог возбуждения мембраны нервных клеток, снижая таким образом эффективность синаптической передачи во многих участках нервной системы. • Поскольку многие из анестезирующих средств хорошо растворимы в липидах, предполагают, что некоторые из них могут изменять физические свойства мембран нейронов, делая их менее чувствительными к возбуждающим агентам.
Влияние лекарств на синаптическую передачу • Большинство анестетиков увеличивают порог возбуждения мембраны нервных клеток, снижая таким образом эффективность синаптической передачи во многих участках нервной системы. • Поскольку многие из анестезирующих средств хорошо растворимы в липидах, предполагают, что некоторые из них могут изменять физические свойства мембран нейронов, делая их менее чувствительными к возбуждающим агентам.
 Рецепторный потенциал • В результате действия адекватного раздражителя у большинства рецепторов увеличивается проницаемость клеточной мембраны для катионов, что приводит к ее деполяризации. • Исключение фоторецепторы, где после поглощения энергии квантов света в связи особенностями управления ионными каналами происходит гиперполяризация мембраны.
Рецепторный потенциал • В результате действия адекватного раздражителя у большинства рецепторов увеличивается проницаемость клеточной мембраны для катионов, что приводит к ее деполяризации. • Исключение фоторецепторы, где после поглощения энергии квантов света в связи особенностями управления ионными каналами происходит гиперполяризация мембраны.
 Рецепторный потенциал • Изменение величины мембранного потенциала рецепторов в ответ на действие стимула представляет собой рецепторный потенциал — входной сигнал первичных сенсорных нейронов. • Если величина рецепторного потенциала достигнет критического уровня деполяризации или превысит его, генерируются потенциалы действия (ПД), с помощью которых сенсорные нейроны передают в ЦНС информацию о действующих стимулах.
Рецепторный потенциал • Изменение величины мембранного потенциала рецепторов в ответ на действие стимула представляет собой рецепторный потенциал — входной сигнал первичных сенсорных нейронов. • Если величина рецепторного потенциала достигнет критического уровня деполяризации или превысит его, генерируются потенциалы действия (ПД), с помощью которых сенсорные нейроны передают в ЦНС информацию о действующих стимулах.
 Рецепторный потенциал • Генерация ПД происходит в ближайшем к рецепторам перехвате Ранвье миелинизированных волокон или ближайшей к рецепторам части мембраны безмиелинового волокна. • Минимальная сила адекватного стимула, достаточная для генерации потенциалов действия в первичном сенсорном нейроне, определяется как его абсолютный порог. • Минимальный прирост силы стимула, сопровождающийся значимым изменением реакции сенсорного нейрона, представляет собой дифференциальный порог его чувствительности.
Рецепторный потенциал • Генерация ПД происходит в ближайшем к рецепторам перехвате Ранвье миелинизированных волокон или ближайшей к рецепторам части мембраны безмиелинового волокна. • Минимальная сила адекватного стимула, достаточная для генерации потенциалов действия в первичном сенсорном нейроне, определяется как его абсолютный порог. • Минимальный прирост силы стимула, сопровождающийся значимым изменением реакции сенсорного нейрона, представляет собой дифференциальный порог его чувствительности.
 • Информация о силе действующего на рецепторы стимула кодируется двумя способами: • частотой потенциалов действия, возникающих в сенсорном нейроне (частотное кодирование), • числом сенсорных нейронов, возбудившихся в ответ на действие стимула.
• Информация о силе действующего на рецепторы стимула кодируется двумя способами: • частотой потенциалов действия, возникающих в сенсорном нейроне (частотное кодирование), • числом сенсорных нейронов, возбудившихся в ответ на действие стимула.
 • Длительность ощущения зависит от реального времени между началом и прекращением воздействия на рецепторы, а также от их способности уменьшать или даже прекращать генерацию нервных импульсов при продолжительном действии адекватного стимула. • При длительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. • Механизмы адаптации не одинаковы в рецепторах разных модальностей, среди них различают быстро адаптирующиеся (например, тактильные рецепторы кожи) и медленно адаптирующиеся рецепторы (например, проприоцепторы мышц и сухожилий).
• Длительность ощущения зависит от реального времени между началом и прекращением воздействия на рецепторы, а также от их способности уменьшать или даже прекращать генерацию нервных импульсов при продолжительном действии адекватного стимула. • При длительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. • Механизмы адаптации не одинаковы в рецепторах разных модальностей, среди них различают быстро адаптирующиеся (например, тактильные рецепторы кожи) и медленно адаптирующиеся рецепторы (например, проприоцепторы мышц и сухожилий).
 сильнее возбуждаются в ответ на быстрое нарастание интенсивности") • Быстро адаптирующиеся рецепторы (фазические) сильнее возбуждаются в ответ на быстрое нарастание интенсивности стимула (фазический ответ), быстро и коротко реагируют на стимуляцию группой импульсов. • Их быстрая адаптация способствует освобождению восприятия от биологически незначительной информации (например, контакт между кожей и одеждой). • Возбуждение медленно адаптирующихся рецепторов мало зависит от скорости изменения стимула и сохраняется при его длительном действии (тонический ответ), они длительное время генерируют ПД. Например, медленная адаптация проприоцепторов позволяет человеку получать нужную ему для сохранения позы информацию в течение всего необходимого времени.
• Быстро адаптирующиеся рецепторы (фазические) сильнее возбуждаются в ответ на быстрое нарастание интенсивности стимула (фазический ответ), быстро и коротко реагируют на стимуляцию группой импульсов. • Их быстрая адаптация способствует освобождению восприятия от биологически незначительной информации (например, контакт между кожей и одеждой). • Возбуждение медленно адаптирующихся рецепторов мало зависит от скорости изменения стимула и сохраняется при его длительном действии (тонический ответ), они длительное время генерируют ПД. Например, медленная адаптация проприоцепторов позволяет человеку получать нужную ему для сохранения позы информацию в течение всего необходимого времени.
 Рецепторный и генераторный потенциалы • Рецепторы специализированные образования, воспринимающие определенные виды раздражений, обладают наибольшей чувствительностью к адекватным для них раздражениям. • Рецепторы делят на четыре группы: механо , термо , хемо и фоторецепторы. • Каждую группу подразделяют на более узкие диапазоны рецепции. • Например, • Зрительные рецепторы делятся на воспринимающие освещенность, цвет, • слуховые определенный тон, • Вкусовые – определенные вкусовые раздражения (соленое, сладкое, горькое) и т. д.
Рецепторный и генераторный потенциалы • Рецепторы специализированные образования, воспринимающие определенные виды раздражений, обладают наибольшей чувствительностью к адекватным для них раздражениям. • Рецепторы делят на четыре группы: механо , термо , хемо и фоторецепторы. • Каждую группу подразделяют на более узкие диапазоны рецепции. • Например, • Зрительные рецепторы делятся на воспринимающие освещенность, цвет, • слуховые определенный тон, • Вкусовые – определенные вкусовые раздражения (соленое, сладкое, горькое) и т. д.
 Рецепторный и генераторный потенциалы • Рецепторный потенциал возникает при раздражении рецептора как результат деполяризации и повышения проводимости участка его мембраны, который называется рецептивным. • Рецептивный участок мембраны имеет специфические свойства, в том числе биохимические, отличающие его от мембраны тела и аксона. • Возникший в рецептивных участках мембраны рецепторный потенциал электротонически распространяется на аксонный холмик рецепторного нейрона, где возникает генераторный потенциал.
Рецепторный и генераторный потенциалы • Рецепторный потенциал возникает при раздражении рецептора как результат деполяризации и повышения проводимости участка его мембраны, который называется рецептивным. • Рецептивный участок мембраны имеет специфические свойства, в том числе биохимические, отличающие его от мембраны тела и аксона. • Возникший в рецептивных участках мембраны рецепторный потенциал электротонически распространяется на аксонный холмик рецепторного нейрона, где возникает генераторный потенциал.
 • Возникновение генераторного потенциала в области аксонного холмика объясняется тем, что этот участок нейрона имеет более низкие пороги возбуждения и ПД в нем развивается раньше, чем в других частях мембраны нейрона. • Чем выше генераторный потенциал, тем интенсивнее частота разрядов распространяющегося ПД от аксона к другим отделам нервной системы. • Следовательно, частота разрядов рецепторного нейрона зависит от амплитуды генераторного потенциала.
• Возникновение генераторного потенциала в области аксонного холмика объясняется тем, что этот участок нейрона имеет более низкие пороги возбуждения и ПД в нем развивается раньше, чем в других частях мембраны нейрона. • Чем выше генераторный потенциал, тем интенсивнее частота разрядов распространяющегося ПД от аксона к другим отделам нервной системы. • Следовательно, частота разрядов рецепторного нейрона зависит от амплитуды генераторного потенциала.
 • Таким образом, реакция рецепторного нейрона, предназначенного для передачи информации из области восприятия, имеет 5 стадий: • 1) преобразование сигнала внешнего раздражения; • 2) генерация рецепторного потенциала; • 3) распространение рецепторного потенциала по нейрону; • 4) возникновение генераторного потенциала; • 5) генерация нервного импульса.
• Таким образом, реакция рецепторного нейрона, предназначенного для передачи информации из области восприятия, имеет 5 стадий: • 1) преобразование сигнала внешнего раздражения; • 2) генерация рецепторного потенциала; • 3) распространение рецепторного потенциала по нейрону; • 4) возникновение генераторного потенциала; • 5) генерация нервного импульса.
 Поясните, как можно блокировать передачу нервного импульса: 1 в. – в области нейрона А; 2 в. – в области нейрона В.
Поясните, как можно блокировать передачу нервного импульса: 1 в. – в области нейрона А; 2 в. – в области нейрона В.
 Ионотропные рецепторы • Это ионные каналы, на наружной поверхности которых имеются собственно рецепторы медиаторов. • Медиатор активирует рецептор. • Активация рецептора приводит к открыванию (реже – закрыванию) ионного канала. • Меняется поток через канал ионов, для которых данный канал избирательно проницаем. • Поток ионов вызывает изменение заряда мембраны; возникает постсинаптический потенциал, обладающий всеми свойствами местных потенциалов (распространяется с затуханием, подчиняется закону силовых отношений; не обладает рефрактерностью; способен к суммации).
Ионотропные рецепторы • Это ионные каналы, на наружной поверхности которых имеются собственно рецепторы медиаторов. • Медиатор активирует рецептор. • Активация рецептора приводит к открыванию (реже – закрыванию) ионного канала. • Меняется поток через канал ионов, для которых данный канал избирательно проницаем. • Поток ионов вызывает изменение заряда мембраны; возникает постсинаптический потенциал, обладающий всеми свойствами местных потенциалов (распространяется с затуханием, подчиняется закону силовых отношений; не обладает рефрактерностью; способен к суммации).
 Метаботропные рецепторы • Эти рецепторы активируются не только медиаторами, но также и гормонами и другими биологически активными веществами. • Они сопряжены с внутриклеточными ферментативными системами: активация рецептора непосредственно или через промежуточные этапы приводит к активации фермента.
Метаботропные рецепторы • Эти рецепторы активируются не только медиаторами, но также и гормонами и другими биологически активными веществами. • Они сопряжены с внутриклеточными ферментативными системами: активация рецептора непосредственно или через промежуточные этапы приводит к активации фермента.
 Метаботропные рецепторы • Делятся на: • рецепторы с собственной ферментативной активностью (трансмембранные белки, внеклеточный участок которых является собственно рецептором, а внутриклеточный ферментом), • рецепторы, сопряженные с ферментами (внеклеточный участок является собственно рецептором, а внутриклеточный активирует определенный фермент), • рецепторы, сопряженные с G белками (внеклеточный участок является собственно рецептором, а внутриклеточный активирует так называемый G белок; последний, в свою очередь, активирует функционально важные внутриклеточные белки, многие из которых являются ферментами).
Метаботропные рецепторы • Делятся на: • рецепторы с собственной ферментативной активностью (трансмембранные белки, внеклеточный участок которых является собственно рецептором, а внутриклеточный ферментом), • рецепторы, сопряженные с ферментами (внеклеточный участок является собственно рецептором, а внутриклеточный активирует определенный фермент), • рецепторы, сопряженные с G белками (внеклеточный участок является собственно рецептором, а внутриклеточный активирует так называемый G белок; последний, в свою очередь, активирует функционально важные внутриклеточные белки, многие из которых являются ферментами).
 Рецептивное поле. Рефлексогенная зона • рецептивное поле рефлекса, область расположения рецепторов, раздражение которых вызывает специфический безусловный рефлекс. • Пример: раздражение поверхности роговицы глаза вызывает рефлекс мигания, слизистой оболочки носоглотки рефлекс чихания; при раздражении барорецепторов дуги аорты и каротидного синуса изменяется АД и ЧСС. • Формирование рефлексогенной зоны определено организацией нервных связей, но видоизменяется в процессе индивидуального развития организма. • Осуществление рефлекса при раздражении одной и той же РЗ зависит от функционального состояния организма и от интенсивности раздражения.
Рецептивное поле. Рефлексогенная зона • рецептивное поле рефлекса, область расположения рецепторов, раздражение которых вызывает специфический безусловный рефлекс. • Пример: раздражение поверхности роговицы глаза вызывает рефлекс мигания, слизистой оболочки носоглотки рефлекс чихания; при раздражении барорецепторов дуги аорты и каротидного синуса изменяется АД и ЧСС. • Формирование рефлексогенной зоны определено организацией нервных связей, но видоизменяется в процессе индивидуального развития организма. • Осуществление рефлекса при раздражении одной и той же РЗ зависит от функционального состояния организма и от интенсивности раздражения.
 Рефлексогенная зона • РЗ, находящиеся на поверхности кожи, могут перекрывать одна другую, вследствие чего раздражение, наносимое на определённый участок кожи, в зависимости от его силы и состояния ЦНС вызывает то один, то другой рефлекс. • Сложно организованные РЗ находятся во внутренних органах и участвуют в рефлекторной регуляции их функциональной деятельности. • Рефлекторный ответ на раздражение РЗ может иногда тормозиться под влиянием других нервных центров, не имеющих отношения к данному рефлексу (Доминанта). • РЗ относятся к структурной организации только безусловных рефлексов, так как условные рефлексы не имеют стабильных рефлекторных дуг.
Рефлексогенная зона • РЗ, находящиеся на поверхности кожи, могут перекрывать одна другую, вследствие чего раздражение, наносимое на определённый участок кожи, в зависимости от его силы и состояния ЦНС вызывает то один, то другой рефлекс. • Сложно организованные РЗ находятся во внутренних органах и участвуют в рефлекторной регуляции их функциональной деятельности. • Рефлекторный ответ на раздражение РЗ может иногда тормозиться под влиянием других нервных центров, не имеющих отношения к данному рефлексу (Доминанта). • РЗ относятся к структурной организации только безусловных рефлексов, так как условные рефлексы не имеют стабильных рефлекторных дуг.
 Функциональные свойства железистых клеток • Большинство гландулоцитов отличаются наличием секреторных включений в цитоплазме, развитыми эндоплазматической сетью и аппаратом Гольджи, а также полярным расположением органелл и секреторных гранул. • Гландулоциты лежат на базальной мембране. В цитоплазме гландулоцитов, которые вырабатывают секреты белкового характера (например, пищеварительные ферменты), хорошо развита гранулярная эндоплазматическая сеть. В клетках, синтезирующих небелковые секреты (липиды, стероиды), выражена агранулярная эндоплазматическая сеть. Многочисленные митохондрии накапливаются в местах наибольшей активности клеток, т. е. там, где образуется секрет. Число секреторных гранул в цитоплазме клеток колеблется в связи с фазами секреторного процесса.
Функциональные свойства железистых клеток • Большинство гландулоцитов отличаются наличием секреторных включений в цитоплазме, развитыми эндоплазматической сетью и аппаратом Гольджи, а также полярным расположением органелл и секреторных гранул. • Гландулоциты лежат на базальной мембране. В цитоплазме гландулоцитов, которые вырабатывают секреты белкового характера (например, пищеварительные ферменты), хорошо развита гранулярная эндоплазматическая сеть. В клетках, синтезирующих небелковые секреты (липиды, стероиды), выражена агранулярная эндоплазматическая сеть. Многочисленные митохондрии накапливаются в местах наибольшей активности клеток, т. е. там, где образуется секрет. Число секреторных гранул в цитоплазме клеток колеблется в связи с фазами секреторного процесса.
 Секреторный цикл • Периодические изменения железистой клетки, связанные с образованием, накоплением, выделением секрета и восстановлением ее для дальнейшей секреции, получили название секреторного цикла: поступление веществ синтез и накопление секрета выведение секрета. • Для образования секрета из крови и лимфы в железистые клетки поступают неорганические соединения, вода и низкомолекулярные органические вещества: АК, моносахариды, жирные кислоты. Путем пиноцитоза в клетку проникают более крупные молекулы органических веществ, например белки. • Из этих продуктов в эндоплазматической сети синтезируются секреты. Они перемещаются в зону аппарата Гольджи, где накапливаются, подвергаются химической перестройке и оформляются в виде гранул, которые выделяются из гландулоцитов.
Секреторный цикл • Периодические изменения железистой клетки, связанные с образованием, накоплением, выделением секрета и восстановлением ее для дальнейшей секреции, получили название секреторного цикла: поступление веществ синтез и накопление секрета выведение секрета. • Для образования секрета из крови и лимфы в железистые клетки поступают неорганические соединения, вода и низкомолекулярные органические вещества: АК, моносахариды, жирные кислоты. Путем пиноцитоза в клетку проникают более крупные молекулы органических веществ, например белки. • Из этих продуктов в эндоплазматической сети синтезируются секреты. Они перемещаются в зону аппарата Гольджи, где накапливаются, подвергаются химической перестройке и оформляются в виде гранул, которые выделяются из гландулоцитов.
 Секреторный цикл • В секреторном цикле выделяют несколько фаз: • поступление в клетку исходных веществ (ведущее значение в этом имеют диффузия, активный транспорт и эндоцитоз), • синтез и транспорт исходного секреторного продукта, • формирование секреторных гранул, • выделение секрета из клетки экзоцитоз. • Из клетки выделяются и негранулированные продукты секреции. • Существуют клетки с разными типами внутриклеточных процессов и видами выделения секретов. В зависимости от типа выделения секрета секрецию делят на голокриновую, апокриновую (макро и микро ) и мерокриновую двух видов в зависимости от механизма выхода секрета через апикальную мембрану: секрет покидает гландулоцит через отверстия, образующиеся при контакте с ней секреторной гранулы в апикальной мембране, или через мембрану, не меняющую свою структуру.
Секреторный цикл • В секреторном цикле выделяют несколько фаз: • поступление в клетку исходных веществ (ведущее значение в этом имеют диффузия, активный транспорт и эндоцитоз), • синтез и транспорт исходного секреторного продукта, • формирование секреторных гранул, • выделение секрета из клетки экзоцитоз. • Из клетки выделяются и негранулированные продукты секреции. • Существуют клетки с разными типами внутриклеточных процессов и видами выделения секретов. В зависимости от типа выделения секрета секрецию делят на голокриновую, апокриновую (макро и микро ) и мерокриновую двух видов в зависимости от механизма выхода секрета через апикальную мембрану: секрет покидает гландулоцит через отверстия, образующиеся при контакте с ней секреторной гранулы в апикальной мембране, или через мембрану, не меняющую свою структуру.
 Типы секреции • Мерокриновый тип – при этом типе секреции железистые клетки полностью сохраняют свою структуру (клетки слюнных желез). • Апокриновый тип – происходит частичное разрушение железистых клеток (клетки молочных желез), т. е. вместе с секреторными продуктами отделяются либо апикальная часть цитоплазмы железистых клеток, либо верхушки микроворсинок. • Голокриновый тип – сопровождается накоплением секрета в цитоплазме и полным разрушением железистых клеток (клетки сальных желез кожи). • Восстановление структуры железистых клеток происходит либо путем внутриклеточной регенерации, либо с помощью клеточной регенерации, т. е. деления и дифференцировки камбиальных клеток. • Нервные механизмы регуляции секреции действуют через высвобождение клеточного кальция, гуморальные преимущественно путем накопления ц. АМФ.
Типы секреции • Мерокриновый тип – при этом типе секреции железистые клетки полностью сохраняют свою структуру (клетки слюнных желез). • Апокриновый тип – происходит частичное разрушение железистых клеток (клетки молочных желез), т. е. вместе с секреторными продуктами отделяются либо апикальная часть цитоплазмы железистых клеток, либо верхушки микроворсинок. • Голокриновый тип – сопровождается накоплением секрета в цитоплазме и полным разрушением железистых клеток (клетки сальных желез кожи). • Восстановление структуры железистых клеток происходит либо путем внутриклеточной регенерации, либо с помощью клеточной регенерации, т. е. деления и дифференцировки камбиальных клеток. • Нервные механизмы регуляции секреции действуют через высвобождение клеточного кальция, гуморальные преимущественно путем накопления ц. АМФ.
 Биопотенциалы гландулоцитов • Биопотенциалы секреторных клеток имеют ряд особенностей в покое и при секреции: • низкую величину и скорость изменения, • градуальность, • различную поляризованность базальной и апикальной мембран, • гетерохронность изменения поляризованности мембраны при секреции и др. • МП гландулоцитов различных экзокринных желез в состоянии относительного покоя равен от 30 до 75 м. В. Стимуляция секреции меняет мембранный потенциал. Это изменение поляризованности мембраны называется секреторным потенциалом. У разных гландулоцитов он имеет существенные различия, характеризует секреторный процесс, влияет на секреторный цикл и сопряжение его фаз, синхронизацию активности гландулоцитов в составе данной железы (это не исключает химического взаимодействия их через межклеточные контакты). Оптимальной для возник новения секреторных потенциалов считается поляризованность мембран, равная 50 м. В.
Биопотенциалы гландулоцитов • Биопотенциалы секреторных клеток имеют ряд особенностей в покое и при секреции: • низкую величину и скорость изменения, • градуальность, • различную поляризованность базальной и апикальной мембран, • гетерохронность изменения поляризованности мембраны при секреции и др. • МП гландулоцитов различных экзокринных желез в состоянии относительного покоя равен от 30 до 75 м. В. Стимуляция секреции меняет мембранный потенциал. Это изменение поляризованности мембраны называется секреторным потенциалом. У разных гландулоцитов он имеет существенные различия, характеризует секреторный процесс, влияет на секреторный цикл и сопряжение его фаз, синхронизацию активности гландулоцитов в составе данной железы (это не исключает химического взаимодействия их через межклеточные контакты). Оптимальной для возник новения секреторных потенциалов считается поляризованность мембран, равная 50 м. В.
 Биопотенциалы гландулоцитов • Для возбуждения большинства видов гландулоцитов характерна деполяризация их мембран, но описаны гландулоциты, при возбуждении которых мембраны гиперполяризуются, формируя двухфазные потенциалы. • Деполяризация мембраны обусловлена потоком ионов Na+ в клетку и выходом из нее ионов К+. • Гиперполяризация мембраны обусловлена транспортом в клетку ионов Сl и выходом из нее ионов Na+ и К+. • Различие в поляризованности базальной и апикальной мембран составляет 2 3 м. В, что создает значительное электрическое поле (20 30 В/см). Его напряженность при возбуж дении секреторной клетки возрастает примерно вдвое, что способствует перемещению секреторных гранул к апикальному полюсу клетки и выходу секреторного материала из клетки.
Биопотенциалы гландулоцитов • Для возбуждения большинства видов гландулоцитов характерна деполяризация их мембран, но описаны гландулоциты, при возбуждении которых мембраны гиперполяризуются, формируя двухфазные потенциалы. • Деполяризация мембраны обусловлена потоком ионов Na+ в клетку и выходом из нее ионов К+. • Гиперполяризация мембраны обусловлена транспортом в клетку ионов Сl и выходом из нее ионов Na+ и К+. • Различие в поляризованности базальной и апикальной мембран составляет 2 3 м. В, что создает значительное электрическое поле (20 30 В/см). Его напряженность при возбуж дении секреторной клетки возрастает примерно вдвое, что способствует перемещению секреторных гранул к апикальному полюсу клетки и выходу секреторного материала из клетки.
 СПАСИБО ЗА ВНИМАНИЕ
СПАСИБО ЗА ВНИМАНИЕ