

ЦНС_3_2010.ppt
- Количество слайдов: 116
 Физиология ЦНС физиология ствола мозга
Физиология ЦНС физиология ствола мозга
 СПИНАЛЬНЫЙ ШОК Развивается в результате прерывания супраспинапьных влияний на мотонейроны спинного мозга при его травмах. Развиваются обратимые угнетения. Различают 4 стадии спинального шока: 1 - полная арефлексия (4 -6 недель) — мгновенно и окончательно выключаются все произвольные движения мышц, иннервируемых из сегментов, лежащих ниже места повреждения. - полный и окончательный потерей осозноваемой чувствительности в областях тела, соответствующих этим сегментам. - временной арефлексии, т. е. временным исчезновением рефлексов, дуги которых проходят ниже места травмы.
СПИНАЛЬНЫЙ ШОК Развивается в результате прерывания супраспинапьных влияний на мотонейроны спинного мозга при его травмах. Развиваются обратимые угнетения. Различают 4 стадии спинального шока: 1 - полная арефлексия (4 -6 недель) — мгновенно и окончательно выключаются все произвольные движения мышц, иннервируемых из сегментов, лежащих ниже места повреждения. - полный и окончательный потерей осозноваемой чувствительности в областях тела, соответствующих этим сегментам. - временной арефлексии, т. е. временным исчезновением рефлексов, дуги которых проходят ниже места травмы.
 • 2. Период появления небольших рефлекторных движений пальцев ног и особенно в первую очередь большого пальца (от 2 недель до несколько месяцев) • 3. Стадия постепенного усиления сгибательных рефлексов. В эту стадию сначала появляются рефлекторные реакции большого пальца ноги (рефлекс Бабинского) и стопы, а в дальнейшем рефлекторное сгибание колена и бедра. Иногда сопровождаются перекрестными расгибательными рефлексами. • 4. Хроническая стадия (спустя шесть и более месяцев) характеризуется преобладанием сгибательных рефлексов, однако могут наблюдать и выраженные расгибательные рефлексы, переходящие в длительные расгибательные спазмы (спинальное стояние). Отклонения от подобной клинической картины, особенно наличие расгибательных рефлексов и повешенного мышечного тонуса, обычно служит симптомом неполного перерыва СМ.
• 2. Период появления небольших рефлекторных движений пальцев ног и особенно в первую очередь большого пальца (от 2 недель до несколько месяцев) • 3. Стадия постепенного усиления сгибательных рефлексов. В эту стадию сначала появляются рефлекторные реакции большого пальца ноги (рефлекс Бабинского) и стопы, а в дальнейшем рефлекторное сгибание колена и бедра. Иногда сопровождаются перекрестными расгибательными рефлексами. • 4. Хроническая стадия (спустя шесть и более месяцев) характеризуется преобладанием сгибательных рефлексов, однако могут наблюдать и выраженные расгибательные рефлексы, переходящие в длительные расгибательные спазмы (спинальное стояние). Отклонения от подобной клинической картины, особенно наличие расгибательных рефлексов и повешенного мышечного тонуса, обычно служит симптомом неполного перерыва СМ.
 Механизм гипертонуса мышц после спинального шока • После исчезновения спинального шока, тонус мышц, ниже повреждения резко повышается, а сгибательные и разгибательные рефлексы нижних конечностей усиливаются: • Причинами гипертонуса яв-ся: - спонтанная активность мышечных рецепторов, что обеспечивает обильную импульсацию от них к αмотонейронам, а их возбуждение приводит к сокращению мышц. - спонтанная активность γ-мотонейронов приводит к сокращению мышечных веретен и как следствие к их возбуждению, которое по Ia – волокнам поступает к α -мотонейронам, вызывая их возбуждение, что приводит к постоянному (тоническому) сокращению мышц и развивается гипертонус у спинального организма.
Механизм гипертонуса мышц после спинального шока • После исчезновения спинального шока, тонус мышц, ниже повреждения резко повышается, а сгибательные и разгибательные рефлексы нижних конечностей усиливаются: • Причинами гипертонуса яв-ся: - спонтанная активность мышечных рецепторов, что обеспечивает обильную импульсацию от них к αмотонейронам, а их возбуждение приводит к сокращению мышц. - спонтанная активность γ-мотонейронов приводит к сокращению мышечных веретен и как следствие к их возбуждению, которое по Ia – волокнам поступает к α -мотонейронам, вызывая их возбуждение, что приводит к постоянному (тоническому) сокращению мышц и развивается гипертонус у спинального организма.
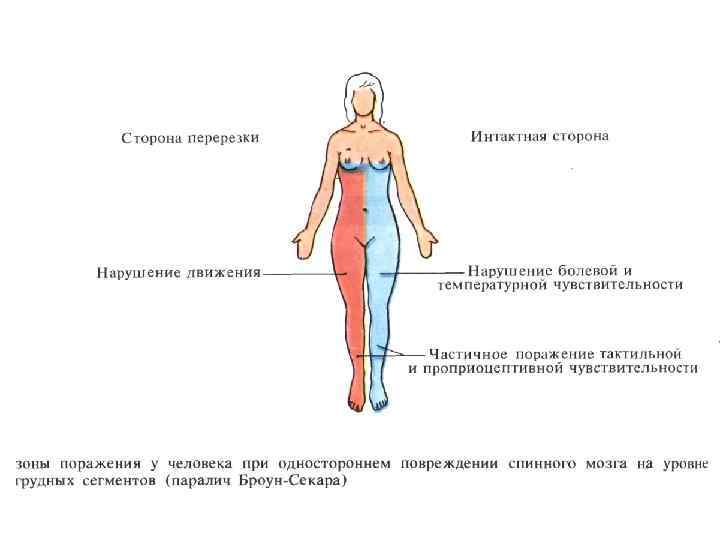
 СИНДРОМ БРОУН-СЕКАРА Односторонний перерыв спинного мозга, характеризуется на стороне поражения двигательным параличем и потерей осязания, а на противоположной - потерей болевой и температурной чувствительности. Причины: - блокада нисходящих путей (пирамидный тракт) — паралич произвольных движений - повреждение заднего столба (потеря осязания) - повреждение переднебокового канатика — потеря болевой, температурной чувствительной на противоположной стороне.
СИНДРОМ БРОУН-СЕКАРА Односторонний перерыв спинного мозга, характеризуется на стороне поражения двигательным параличем и потерей осязания, а на противоположной - потерей болевой и температурной чувствительности. Причины: - блокада нисходящих путей (пирамидный тракт) — паралич произвольных движений - повреждение заднего столба (потеря осязания) - повреждение переднебокового канатика — потеря болевой, температурной чувствительной на противоположной стороне.
 ПАТОЛОГИЧЕСКИЕ РАЗГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА - - - РЕФЛЕКС БАБИНСКОГО - при раздражении подошвы стопы штриховым движением от пятки до носка происходит разгибание большого пальца в сторону тыла. В норме этот рефлекс имеется у детей примерно до двух лет. Потом тормозиться и выявляется при нарушении супраспинальных рефлексов. РЕФЛЕКС ОПЕНГЕЙМА - при надавливании на поверхность голени в области ТИБИА от колена до голеностопного сустава - происходит тыльное расгибание большого пальца. РЕФЛЕКС ГОРДОНА - при сдавливании икраножной мышцы происходит тыльное расгибание большого пальца РЕФЛЕКС ШЕФЕРА - то же самое, при сдавливании ахиллового сухожилия.
ПАТОЛОГИЧЕСКИЕ РАЗГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА - - - РЕФЛЕКС БАБИНСКОГО - при раздражении подошвы стопы штриховым движением от пятки до носка происходит разгибание большого пальца в сторону тыла. В норме этот рефлекс имеется у детей примерно до двух лет. Потом тормозиться и выявляется при нарушении супраспинальных рефлексов. РЕФЛЕКС ОПЕНГЕЙМА - при надавливании на поверхность голени в области ТИБИА от колена до голеностопного сустава - происходит тыльное расгибание большого пальца. РЕФЛЕКС ГОРДОНА - при сдавливании икраножной мышцы происходит тыльное расгибание большого пальца РЕФЛЕКС ШЕФЕРА - то же самое, при сдавливании ахиллового сухожилия.
 ПАТОЛОГИЧЕСКИЕ СГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА • ХВАТАТЕЛЬНЫЙ РЕФЛЕКС ЯНУШКЕВИЧА - возникает при касании пальцев руки или стопы каким либо предметом и проявляется в непроизвольном захватавании этого предмета. У детей этот рефлекс проявляется в возрасте до 2 - 4 месяцев, при этом удается приподнять ребенка над опорой — хватательный рефлекс РОБИНСОНА • РЕФЛЕКС РОССОЛИНО - сгибание пальцев стопы, в том числе и большого при быстром касательном ударе по подушечкам пальцев стопы • РЕФЛЕКС БЕХТЕРЕВА - МЕНДЕЛЯ - подошвенное сгибание большого пальца при ударе молоточком по тылу стопы • РЕФЛЕКС ЖУКОВСКОГО - то же самое при ударе по подошвенной поверхности под пальцами стопы • РЕФЛЕКС БЕХТЕРЕВА - то же самое при ударе по пятке
ПАТОЛОГИЧЕСКИЕ СГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА • ХВАТАТЕЛЬНЫЙ РЕФЛЕКС ЯНУШКЕВИЧА - возникает при касании пальцев руки или стопы каким либо предметом и проявляется в непроизвольном захватавании этого предмета. У детей этот рефлекс проявляется в возрасте до 2 - 4 месяцев, при этом удается приподнять ребенка над опорой — хватательный рефлекс РОБИНСОНА • РЕФЛЕКС РОССОЛИНО - сгибание пальцев стопы, в том числе и большого при быстром касательном ударе по подушечкам пальцев стопы • РЕФЛЕКС БЕХТЕРЕВА - МЕНДЕЛЯ - подошвенное сгибание большого пальца при ударе молоточком по тылу стопы • РЕФЛЕКС ЖУКОВСКОГО - то же самое при ударе по подошвенной поверхности под пальцами стопы • РЕФЛЕКС БЕХТЕРЕВА - то же самое при ударе по пятке
 ПАТОЛОГИЧЕСКИЕ РЕФЛЕКСЫ, ВЫЗЫВАЕМЫЕ ПРИ НАНЕСЕНИИ РАЗДРАЖЕНИЯ НА ВЕРХНИЕ КОНЕЧНОСТИ: • РЕФЛЕКС РОССОЛИНО - сгибание пальцев кисти в ответ на быстрое касание к пальцам • РЕФЛЕКС ЖУКОВСКОГО - то же самое при ударе молоточком по ладонной поверхости кисти • РЕФЛЕКС БЕХТЕРЕВА - то же самое при ударе по тылу кисти
ПАТОЛОГИЧЕСКИЕ РЕФЛЕКСЫ, ВЫЗЫВАЕМЫЕ ПРИ НАНЕСЕНИИ РАЗДРАЖЕНИЯ НА ВЕРХНИЕ КОНЕЧНОСТИ: • РЕФЛЕКС РОССОЛИНО - сгибание пальцев кисти в ответ на быстрое касание к пальцам • РЕФЛЕКС ЖУКОВСКОГО - то же самое при ударе молоточком по ладонной поверхости кисти • РЕФЛЕКС БЕХТЕРЕВА - то же самое при ударе по тылу кисти
 ФИЗИОЛОГИЯ СТВОЛА МОЗГА
ФИЗИОЛОГИЯ СТВОЛА МОЗГА
 Функции продолговатого мозга • Рефлекторная • Проводниковая • Тоническая
Функции продолговатого мозга • Рефлекторная • Проводниковая • Тоническая
 СРЕЗ ПРОДОЛГОВАТОГО МОЗГА
СРЕЗ ПРОДОЛГОВАТОГО МОЗГА
 Расположение ядер черепных нервов в стволовой части мозга 1 — добавочный нерв, 2 — подъязычный, 3 — преддверно—улитковый, 4 — отводящий, 5 — блоковый, 6 — глазодвигательный, 7 — тройничный, 8 — лицевой, 9 — языкоглоточный, 10 — блуждающий нерв.
Расположение ядер черепных нервов в стволовой части мозга 1 — добавочный нерв, 2 — подъязычный, 3 — преддверно—улитковый, 4 — отводящий, 5 — блоковый, 6 — глазодвигательный, 7 — тройничный, 8 — лицевой, 9 — языкоглоточный, 10 — блуждающий нерв.
. Нарушая") Схема расположения двигательных центров в стволе мозга (продолговатом мозгу, мосте и среднем мозгу). Нарушая связь с отделами мозга, расположенными выше каждой из трех указанных линий перерезки, получают соответственно децеребрированное (1), мезенцефальное (2) и таламическое (3) животное
Схема расположения двигательных центров в стволе мозга (продолговатом мозгу, мосте и среднем мозгу). Нарушая связь с отделами мозга, расположенными выше каждой из трех указанных линий перерезки, получают соответственно децеребрированное (1), мезенцефальное (2) и таламическое (3) животное
 Ядра продолговатого мозга 1. ЯДРА ЧЕРЕПН 0 М 03 Г 0 ВЫх НЕРВОВ XII пара - ПОДЪЯЗЫЧНОГО НЕРВА - n. hypoglossus - двигательные ядра XI пара - ДОБАВОЧНОГО НЕРВА - n. accessorius двигательные ядра X пара - БЛУЖДАЮЩЕГО НЕРВА n. vagus: • 1) вегетативное ядро • 2) чувствительнее ядро одиночного пучка • 3) обоюдное ЯДРО - двигательное тки и гортани
Ядра продолговатого мозга 1. ЯДРА ЧЕРЕПН 0 М 03 Г 0 ВЫх НЕРВОВ XII пара - ПОДЪЯЗЫЧНОГО НЕРВА - n. hypoglossus - двигательные ядра XI пара - ДОБАВОЧНОГО НЕРВА - n. accessorius двигательные ядра X пара - БЛУЖДАЮЩЕГО НЕРВА n. vagus: • 1) вегетативное ядро • 2) чувствительнее ядро одиночного пучка • 3) обоюдное ЯДРО - двигательное тки и гортани
 Ядра продолговатого мозга - 2 2. ЯДРА ЧЁРЁПНОМОЗГОВЫХ НЕРВОВ: IX пара - языкоглоточного НЕРВА: • 1) двигательное ядро - рот и глотка • 2) чувствительное ядро - вкус задней трети языка • 3) вегетативное ядро - слюнные железы • На границе с мостом: YIII пара - ВЕСТИБУЛОКОХЛЕАРНОГО НЕРВА • 1) кохлеарные ядра • 2) вестибулярные ядра - медиальное Швальбе, латеральное Дейтерса, верхнее Бехтерева
Ядра продолговатого мозга - 2 2. ЯДРА ЧЁРЁПНОМОЗГОВЫХ НЕРВОВ: IX пара - языкоглоточного НЕРВА: • 1) двигательное ядро - рот и глотка • 2) чувствительное ядро - вкус задней трети языка • 3) вегетативное ядро - слюнные железы • На границе с мостом: YIII пара - ВЕСТИБУЛОКОХЛЕАРНОГО НЕРВА • 1) кохлеарные ядра • 2) вестибулярные ядра - медиальное Швальбе, латеральное Дейтерса, верхнее Бехтерева
 Ядра продолговатого мозга - 3 ПЕРЕКЛЮЧАЮЩИЕ ЯДРА: • Голля и Бурдаха – к таламусу • Ретикулярной формации- от коры и подкорковых ганглиев к спинному мозгу • Оливарные ядра - от коры, подкорковым ядер и мозжечка к спинному мозгу и от спинного мозга к мозжечку, таламусу и коре; от слуховым ядер в средний мозг и четверохолмие.
Ядра продолговатого мозга - 3 ПЕРЕКЛЮЧАЮЩИЕ ЯДРА: • Голля и Бурдаха – к таламусу • Ретикулярной формации- от коры и подкорковых ганглиев к спинному мозгу • Оливарные ядра - от коры, подкорковым ядер и мозжечка к спинному мозгу и от спинного мозга к мозжечку, таламусу и коре; от слуховым ядер в средний мозг и четверохолмие.
 Дыхательный 7) Мигания 2) Сердечно-сосудистый 8) Рвоты 3) Слюноотделения 9)") Центры продолговатого мозга 1) Дыхательный 7) Мигания 2) Сердечно-сосудистый 8) Рвоты 3) Слюноотделения 9) Сосания 4) Слезоотделения 10) Жевания 5) Кашля 11) Глотания 6) Чихания 12) Рефлексов поддержания позы
Центры продолговатого мозга 1) Дыхательный 7) Мигания 2) Сердечно-сосудистый 8) Рвоты 3) Слюноотделения 9) Сосания 4) Слезоотделения 10) Жевания 5) Кашля 11) Глотания 6) Чихания 12) Рефлексов поддержания позы
 РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА • • • Жизненно-важные рефлексы Защитные рефлексы Рефлексы пищевого поведения Рефлексы поддержания позы Вегетативные рефлексы Вестибуло-вегетативные рефлексы
РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА • • • Жизненно-важные рефлексы Защитные рефлексы Рефлексы пищевого поведения Рефлексы поддержания позы Вегетативные рефлексы Вестибуло-вегетативные рефлексы
![Постуральные рефлексы СТАТИЧЕСКИЕ - от рецепторов преддверия • рефлексы положения • рефлексы выпрямления (установочные]](https://present5.com/presentation/134069999_112202509/image-20.jpg "Постуральные рефлексы СТАТИЧЕСКИЕ - от рецепторов преддверия • рефлексы положения • рефлексы выпрямления (установочные]") Постуральные рефлексы СТАТИЧЕСКИЕ - от рецепторов преддверия • рефлексы положения • рефлексы выпрямления (установочные] СТАТОКИНЕТИЧЕСКИЕ - от рецепторов полукружных каналов • рефлексы прямолинейного ускорения • рефлексы углового ускорения
Постуральные рефлексы СТАТИЧЕСКИЕ - от рецепторов преддверия • рефлексы положения • рефлексы выпрямления (установочные] СТАТОКИНЕТИЧЕСКИЕ - от рецепторов полукружных каналов • рефлексы прямолинейного ускорения • рефлексы углового ускорения
 ВЕСТИБУЛЯРНЫЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА • ВЕРХНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Бехтерева • НИЖНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Роллера (нисходящие влияния) • МЕДИАЛЬНОЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Швальбе • ЛАТЕРАЛЬНОЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Дейтерса (начинается вестибулоспинальный путь) • Также проходит транзитом в мозжечёк, непрерываясь в продолговатом мозге, вестибулоцеребеллярный путь от афферентных нейронов вестибулярного ганглия
ВЕСТИБУЛЯРНЫЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА • ВЕРХНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Бехтерева • НИЖНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Роллера (нисходящие влияния) • МЕДИАЛЬНОЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Швальбе • ЛАТЕРАЛЬНОЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Дейтерса (начинается вестибулоспинальный путь) • Также проходит транзитом в мозжечёк, непрерываясь в продолговатом мозге, вестибулоцеребеллярный путь от афферентных нейронов вестибулярного ганглия
 ОСНОВНЫЕ ПРОВОДЯЩИЕ ПУТИ ОТ ВЕСТИБУЛЯРНЫХ ЯДЕР • • ВЕСТИБУЛОСПИНАЛЬНЫЙ ПУТЬ — от ядра Дейтерса передает информацию на альфа - и гамма - мотонейроны СМ, активируя мышцы разгибатели и тормозя - сгибатели. ВЕСТИБУЛООКУЛЯРНЫИ ПУТЬ — передает информацию на мышцы глаза при движении, благодаря этому на сетчатке глаза сохраняется объект наблюдения, несмотря на всевозможные перемещения. -ВЕСТИБУЛОМОЗЖЕЧКОВЫЙ ПУТЬ — передает информацию к мозжечку и характеризует о положении тела в пространстве. Совместно с вестибулослинальным трактом обеспечивает регуляцию мышечного тонуса и других перемещений тела. ВЕСТИБУЛОТАЛАМИЧЕСКИЙ ПУТЬ - обеспечивает передачу информации к: 1. Специфическим ядрам таламуса по Лемнисковому пути, а от них к сенсорной зоне коры, расположенной в прецентральной извилине в области проекции лица. 2. Колатерали идут к ретикулярной формации, а от нее к неспецифическим ядрам таламуса, от которых диффузно ко многим участкам коры, вызываю их активацию.
ОСНОВНЫЕ ПРОВОДЯЩИЕ ПУТИ ОТ ВЕСТИБУЛЯРНЫХ ЯДЕР • • ВЕСТИБУЛОСПИНАЛЬНЫЙ ПУТЬ — от ядра Дейтерса передает информацию на альфа - и гамма - мотонейроны СМ, активируя мышцы разгибатели и тормозя - сгибатели. ВЕСТИБУЛООКУЛЯРНЫИ ПУТЬ — передает информацию на мышцы глаза при движении, благодаря этому на сетчатке глаза сохраняется объект наблюдения, несмотря на всевозможные перемещения. -ВЕСТИБУЛОМОЗЖЕЧКОВЫЙ ПУТЬ — передает информацию к мозжечку и характеризует о положении тела в пространстве. Совместно с вестибулослинальным трактом обеспечивает регуляцию мышечного тонуса и других перемещений тела. ВЕСТИБУЛОТАЛАМИЧЕСКИЙ ПУТЬ - обеспечивает передачу информации к: 1. Специфическим ядрам таламуса по Лемнисковому пути, а от них к сенсорной зоне коры, расположенной в прецентральной извилине в области проекции лица. 2. Колатерали идут к ретикулярной формации, а от нее к неспецифическим ядрам таламуса, от которых диффузно ко многим участкам коры, вызываю их активацию.
 ОБЩАЯ СХЕМА ОБРАБОТКИ И ПЕРЕДАЧИ ИНФОРМАЦИИ ВЕСТИБУЛЯРНЫМИ ЯДРАМИ • 1. От вестибулярного ганглия информация передается в верхнее вестибулярное ядро (ядро Бехтерева), где происходит ее обработка. • 2. Из ядра Бехтерева, переработанная информация оценки необходимости изменения позы, посылается к латеральному вестибулярному ядру (ядро Дейтерса) и медиальному вестибулярному ядру (ядро Швальбе) • 3. От медиального (ядро Швальбе) и латерального ядер (ядро Дейтерса) информация по вестибулоспинальному трактупоступает в передним рогам соответствующим сегментам СМ, обеспечивая иннервацию альфа — мотонейронов мышц, которые изменяют позу в данный момент времени.
ОБЩАЯ СХЕМА ОБРАБОТКИ И ПЕРЕДАЧИ ИНФОРМАЦИИ ВЕСТИБУЛЯРНЫМИ ЯДРАМИ • 1. От вестибулярного ганглия информация передается в верхнее вестибулярное ядро (ядро Бехтерева), где происходит ее обработка. • 2. Из ядра Бехтерева, переработанная информация оценки необходимости изменения позы, посылается к латеральному вестибулярному ядру (ядро Дейтерса) и медиальному вестибулярному ядру (ядро Швальбе) • 3. От медиального (ядро Швальбе) и латерального ядер (ядро Дейтерса) информация по вестибулоспинальному трактупоступает в передним рогам соответствующим сегментам СМ, обеспечивая иннервацию альфа — мотонейронов мышц, которые изменяют позу в данный момент времени.
 • - VIII пара ЧМН имеет две веточки: • • вестибулярная и слуховая — являющихся чувствительными (афферентными). Слуховая ветвь образована афферентными волокнами от кортиевого органа улитки и передает информацию в нижнее двухолмие. Вестибулярная ветвь — в значительной мере образована афферентными волокнами идущими от полукружных каналов вестибулярного анализатора, заканчивающихся в вестибулярном ганглии, а от него к вестибулярным ядрам продолговатого мозга.
• - VIII пара ЧМН имеет две веточки: • • вестибулярная и слуховая — являющихся чувствительными (афферентными). Слуховая ветвь образована афферентными волокнами от кортиевого органа улитки и передает информацию в нижнее двухолмие. Вестибулярная ветвь — в значительной мере образована афферентными волокнами идущими от полукружных каналов вестибулярного анализатора, заканчивающихся в вестибулярном ганглии, а от него к вестибулярным ядрам продолговатого мозга.
 поступает:") ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ ОТ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА Информация от вестибулярного анализатора (VIII пара ЧМН) поступает: • от волосковых клеток макулы (расположенных в маточке и мешочках — линейное ускорение, а также наклон головы вперед или назад) • от волосковых клеток гребешков (расположенных в расширенной части полукружных каналов — угловое ускорение (ускорение Кориолиса), а также начало и конец движения) • передается на дендриты аффентных нейронов вестибулярного ганглия, а от него к вестибулярным ядрам продолговатого мозга.
ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ ОТ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА Информация от вестибулярного анализатора (VIII пара ЧМН) поступает: • от волосковых клеток макулы (расположенных в маточке и мешочках — линейное ускорение, а также наклон головы вперед или назад) • от волосковых клеток гребешков (расположенных в расширенной части полукружных каналов — угловое ускорение (ускорение Кориолиса), а также начало и конец движения) • передается на дендриты аффентных нейронов вестибулярного ганглия, а от него к вестибулярным ядрам продолговатого мозга.
, определяя состояние активности его активности и") ЯДРО ДЕЙТЕРСА находится под контролем мозжечка {ядро шатра), определяя состояние активности его активности и контролирует тонус мышц сгибателей и разгибателей. Патология мозжечка проявляется примерно сходными симптомами, что и патология вестибулярного аппарата и вестибулярных ядер продолговатого мозга. От ядра Дейтерса по вестибулоспинальному пути информация поступает к альфа-мотонейронам СМ, вызывая возбуждение альфа-мотонейронов разгибателей и одновременно торможению альфамотонейронов мышц сгибателей. Благодаря этому меняется тонус мышц верхних и нижних конечностей — и сохраняется равновесие
ЯДРО ДЕЙТЕРСА находится под контролем мозжечка {ядро шатра), определяя состояние активности его активности и контролирует тонус мышц сгибателей и разгибателей. Патология мозжечка проявляется примерно сходными симптомами, что и патология вестибулярного аппарата и вестибулярных ядер продолговатого мозга. От ядра Дейтерса по вестибулоспинальному пути информация поступает к альфа-мотонейронам СМ, вызывая возбуждение альфа-мотонейронов разгибателей и одновременно торможению альфамотонейронов мышц сгибателей. Благодаря этому меняется тонус мышц верхних и нижних конечностей — и сохраняется равновесие
 От вестибулярных ядер продолговатого мозга информация передается к медиальному продольному пучку, который начинается от ядра Даршкевича и промежуточного ядра среднего мозга и ведет в сторону спинного мозга, соединяя в единый функциональный ансамбль все ядра нервов, участвующих в регуляции активности мышц глазного яблока (III пара) глазодвигательных, IV пара ( блоковый), VI пара (отводящий) ЧМН. Благодаря этому, движение обоих глаз происходит синхронно и в норме у большенства людей отсутствует косоглазие. С участием продольного медиального пучка осуществляется явление НИСТАГМА глазных яблок — т. е. движения глаз, которые способствуют поддержанию взора на точке фиксации — на сетчатку, изображение падает примерно на одно и тоже место, не смотря на то, что человек совершает движения.
От вестибулярных ядер продолговатого мозга информация передается к медиальному продольному пучку, который начинается от ядра Даршкевича и промежуточного ядра среднего мозга и ведет в сторону спинного мозга, соединяя в единый функциональный ансамбль все ядра нервов, участвующих в регуляции активности мышц глазного яблока (III пара) глазодвигательных, IV пара ( блоковый), VI пара (отводящий) ЧМН. Благодаря этому, движение обоих глаз происходит синхронно и в норме у большенства людей отсутствует косоглазие. С участием продольного медиального пучка осуществляется явление НИСТАГМА глазных яблок — т. е. движения глаз, которые способствуют поддержанию взора на точке фиксации — на сетчатку, изображение падает примерно на одно и тоже место, не смотря на то, что человек совершает движения.
 При патологии вестибулярного аппарата или вестибулярных ядер и вестибулярного тракта, возникают такие симптомы, как • головокружение (субъективное осущение вращение предметов окружающего мира, которое возникает вследствие «НЕ ВЕРНОЙ ИНФОРМАЦИИ, ПОСТУПАЮЩЕЙ В АССОЦИАТИВНЫЕ УЧАСТКИ КОРЫ» ), • нарушения равновесия (атаксия статическая и статокинетическая) возникающий спонтанно НИСТАГМ глаз: - горизотальный - вертикальный - или ротационный
При патологии вестибулярного аппарата или вестибулярных ядер и вестибулярного тракта, возникают такие симптомы, как • головокружение (субъективное осущение вращение предметов окружающего мира, которое возникает вследствие «НЕ ВЕРНОЙ ИНФОРМАЦИИ, ПОСТУПАЮЩЕЙ В АССОЦИАТИВНЫЕ УЧАСТКИ КОРЫ» ), • нарушения равновесия (атаксия статическая и статокинетическая) возникающий спонтанно НИСТАГМ глаз: - горизотальный - вертикальный - или ротационный
, участвующих в регуляции двигательной активности: •") ЯДРА РЕТИКУЛЯРНОЙ ФОРМАЦИИ ЗАДНЕГО МОЗГА (ПМ И МОСТ), участвующих в регуляции двигательной активности: • 1. Нейроны продолговатого мозга — активируют альфа-мотонейроны сгибателей и тормозят альфа-мотонейроны разгибателей, т. е. ведут себя как нейроны Красного ядра. • 2. Нейроны ретикулярной формации моста активируют — альфа-мотонейроны разгибателей и тормозят альфа-мотонейроны сгибателей, т. е. действуют как нейроны вестибулярных ядер (ядро Дейтерса)
ЯДРА РЕТИКУЛЯРНОЙ ФОРМАЦИИ ЗАДНЕГО МОЗГА (ПМ И МОСТ), участвующих в регуляции двигательной активности: • 1. Нейроны продолговатого мозга — активируют альфа-мотонейроны сгибателей и тормозят альфа-мотонейроны разгибателей, т. е. ведут себя как нейроны Красного ядра. • 2. Нейроны ретикулярной формации моста активируют — альфа-мотонейроны разгибателей и тормозят альфа-мотонейроны сгибателей, т. е. действуют как нейроны вестибулярных ядер (ядро Дейтерса)
 Роль продолговатого мозга в двигательных функциях
Роль продолговатого мозга в двигательных функциях
 ФИЗИОЛОГИЯ СРЕДНЕГО И ПРОМЕЖУТОЧНОГО МОЗГА
ФИЗИОЛОГИЯ СРЕДНЕГО И ПРОМЕЖУТОЧНОГО МОЗГА
 ПРОДОЛГОВАТЫЙ МОЗГ, МОСТ И СРЕДНИЙ МОЗГ
ПРОДОЛГОВАТЫЙ МОЗГ, МОСТ И СРЕДНИЙ МОЗГ
 Средний мозг
Средний мозг
 ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
 Основные ядра среднего мозга • Ядра черепномозговых нервов: • --- III пара - глазодвигательного нерва --- IV пара - блокового нерва --- Ядро Даркшевича - продольный пучок среднего мозга, связывающий ядра глазодвигательного, блокового и отводящего нерва в единую систему • Непарное вегетативное ядро Якубовича-Эдингера - через цилиарный ганглий к мышцам радужки и ресничного тела • Ядра тектальной области: верхнее или переднее двухолмие зрительные рефлексы; нижнее или заднее двухолмие - слуховые рефлексы - четверохолмие • Черная субстанция • Красные ядра
Основные ядра среднего мозга • Ядра черепномозговых нервов: • --- III пара - глазодвигательного нерва --- IV пара - блокового нерва --- Ядро Даркшевича - продольный пучок среднего мозга, связывающий ядра глазодвигательного, блокового и отводящего нерва в единую систему • Непарное вегетативное ядро Якубовича-Эдингера - через цилиарный ганглий к мышцам радужки и ресничного тела • Ядра тектальной области: верхнее или переднее двухолмие зрительные рефлексы; нижнее или заднее двухолмие - слуховые рефлексы - четверохолмие • Черная субстанция • Красные ядра
 ДВИГАТЕЛЬНЫЕ ЯДРА СТВОЛА МОЗГА Можно условно разделить на два класса по их влиянию на мышцы: • флексорные системы — повышающие активность сгибателей — нейроны Красного ядра и ретикулярной формации ПМ. • экстензорные системы — повышающие активность разгибателей — нейроны вестибулярных ядер и ретикулярной формации моста
ДВИГАТЕЛЬНЫЕ ЯДРА СТВОЛА МОЗГА Можно условно разделить на два класса по их влиянию на мышцы: • флексорные системы — повышающие активность сгибателей — нейроны Красного ядра и ретикулярной формации ПМ. • экстензорные системы — повышающие активность разгибателей — нейроны вестибулярных ядер и ретикулярной формации моста
 Стволовые рефлексы тонуса мышц, позы и равновесия тела. • Осуществляется с вестибулярных, проприоцентивных, тактильных и зрительных рецепторов через двигательные центры ствола, включая центры ретикулярной формации (РФ). • Эти эффекторные влияния из моторных центров ствола распространяются на мускулатуру конечностей и туловища по четырем нисходящим путям: 1. вестибулярному 2. медиальному ретикулоспинальному [1 и 2 – состовляют медиальную нисходящую экстензорную (разгибательную) систему] 3. руброспинальному 4. латеральному ретикулоспинальному [состовляют латеральную нисходящую флексорную (сгибательную) систему].
Стволовые рефлексы тонуса мышц, позы и равновесия тела. • Осуществляется с вестибулярных, проприоцентивных, тактильных и зрительных рецепторов через двигательные центры ствола, включая центры ретикулярной формации (РФ). • Эти эффекторные влияния из моторных центров ствола распространяются на мускулатуру конечностей и туловища по четырем нисходящим путям: 1. вестибулярному 2. медиальному ретикулоспинальному [1 и 2 – состовляют медиальную нисходящую экстензорную (разгибательную) систему] 3. руброспинальному 4. латеральному ретикулоспинальному [состовляют латеральную нисходящую флексорную (сгибательную) систему].
 Медиальная разгибательная система Вестибулоспинальный путь – от латеральных вестибулярных ядер, которые получают афферентную импульсацию от рецепторов отолитового аппарата и ампул полукружных каналов. Волокна этого пути возбуждают спинальные α- и γ- мотонейроны мышцразгибателей конечностей, туловища, шеи и одновременно, через тормозные интернейроны тормозят сгибатели. Медиальный ретиколоспинальный путь – от ретикулярных ядер моста. Через них осуществляется стимуляция α- и γ- мотонейронов мышц-разгибателей осевой мускулатуры тела (туловища и проксимальных отделов конечностей), и торможение сгибателей.
Медиальная разгибательная система Вестибулоспинальный путь – от латеральных вестибулярных ядер, которые получают афферентную импульсацию от рецепторов отолитового аппарата и ампул полукружных каналов. Волокна этого пути возбуждают спинальные α- и γ- мотонейроны мышцразгибателей конечностей, туловища, шеи и одновременно, через тормозные интернейроны тормозят сгибатели. Медиальный ретиколоспинальный путь – от ретикулярных ядер моста. Через них осуществляется стимуляция α- и γ- мотонейронов мышц-разгибателей осевой мускулатуры тела (туловища и проксимальных отделов конечностей), и торможение сгибателей.
 Латеральная сгибательная система Руброспинальный путь – от красного ядра покрышки мозга, возбуждает α- и γ- мотонейроны сгибателей и реципрокно тормозят мотонейроны разгибателей. Латеральный ретикулоспинальный путь – от гигантоклеточного ретикулярного ядра продолговатого мозга, его волокна возбуждают α- и γ- мотонейроны мышц-сгибателей и реципрокно тормозят мышцы-разбибатели. Вместе с тем раздражение нейронов вентральной части РФ продолговатого мозга, вызывает подавление всех спинальных рефлексов (сгибательных и разгибательных) и ликвидацию децеребрационной ригидности. Поэтому механизму осуществляется центральное торможение рефлексов СМ, открытое И. М. Сеченовым (1862 г. )
Латеральная сгибательная система Руброспинальный путь – от красного ядра покрышки мозга, возбуждает α- и γ- мотонейроны сгибателей и реципрокно тормозят мотонейроны разгибателей. Латеральный ретикулоспинальный путь – от гигантоклеточного ретикулярного ядра продолговатого мозга, его волокна возбуждают α- и γ- мотонейроны мышц-сгибателей и реципрокно тормозят мышцы-разбибатели. Вместе с тем раздражение нейронов вентральной части РФ продолговатого мозга, вызывает подавление всех спинальных рефлексов (сгибательных и разгибательных) и ликвидацию децеребрационной ригидности. Поэтому механизму осуществляется центральное торможение рефлексов СМ, открытое И. М. Сеченовым (1862 г. )
 КРАСНОЕ ЯДРО Расположено в области среднего мозга, его нейроны получают информацию из моторной зоны коры больших полушарий, промежуточных ядер мозжечка ( пробковидного и шаровидного) и от нейронов черной субстанции, т. о. имеют всю информацию о положении тела в прострастве, о состоянии мышечной системы и кожи. По рубраспинальному трактупередает информацию в спинальные центры, контролирующие мускулатуру верхних и нижних конечностей.
КРАСНОЕ ЯДРО Расположено в области среднего мозга, его нейроны получают информацию из моторной зоны коры больших полушарий, промежуточных ядер мозжечка ( пробковидного и шаровидного) и от нейронов черной субстанции, т. о. имеют всю информацию о положении тела в прострастве, о состоянии мышечной системы и кожи. По рубраспинальному трактупередает информацию в спинальные центры, контролирующие мускулатуру верхних и нижних конечностей.
 НЕЙРОНЫ КРАСНОГО ЯДРА Через руброспинальный тракт оказывает активирующее влияние на альфа — мотонейрона мышц сгибателей и тормозят активность альфа-мотонейронов разгибателей, т. о. уравновешивают активность вестибулярного ядра Дейтерса, а совместно с ним участвуют в регуляции позы. Отделение красного ядра от нижележащих структур (перерезка выше моста) ведет к повышению тонуса мышц разгибателей (экстензоров) и развитию ДЕЦЕРЕБРАЦИОННОЙ ригидности -в этих условиях разрушение ядра Дейтерса или перерезка вестибулоспинального тракта, ведет к исчезновению ригидности. В развитии ДЕЦЕРЕБРАЦИОННОЙ ригидности участвует гамма - петля, т. к. ДЕАФФЕРЕНТАЦИЯ конечности устраняет децеребрационную ригидность.
НЕЙРОНЫ КРАСНОГО ЯДРА Через руброспинальный тракт оказывает активирующее влияние на альфа — мотонейрона мышц сгибателей и тормозят активность альфа-мотонейронов разгибателей, т. о. уравновешивают активность вестибулярного ядра Дейтерса, а совместно с ним участвуют в регуляции позы. Отделение красного ядра от нижележащих структур (перерезка выше моста) ведет к повышению тонуса мышц разгибателей (экстензоров) и развитию ДЕЦЕРЕБРАЦИОННОЙ ригидности -в этих условиях разрушение ядра Дейтерса или перерезка вестибулоспинального тракта, ведет к исчезновению ригидности. В развитии ДЕЦЕРЕБРАЦИОННОЙ ригидности участвует гамма - петля, т. к. ДЕАФФЕРЕНТАЦИЯ конечности устраняет децеребрационную ригидность.
 ЧЕРНАЯ СУБСТАНЦИЯ Получает обильную информацию от базалальных ганглиев и участвует в координации движений и эмоций. При разрушении развивается болезнь ПАРКИНСОНА -нарушается координация тонких движений пальцев рук, мышечной ригидности, тремору. От ядра Дейтерса по вестибулоспинальному пути информация поступает к альфа-мотонейронам СМ, вызывая возбуждение альфа-мотонейронов разгибателей и одновременно торможению альфамотонейронов мышц сгибателей. Благодаря этому меняется тонус мышц верхних и нижних конечностей — и сохраняется равновесие
ЧЕРНАЯ СУБСТАНЦИЯ Получает обильную информацию от базалальных ганглиев и участвует в координации движений и эмоций. При разрушении развивается болезнь ПАРКИНСОНА -нарушается координация тонких движений пальцев рук, мышечной ригидности, тремору. От ядра Дейтерса по вестибулоспинальному пути информация поступает к альфа-мотонейронам СМ, вызывая возбуждение альфа-мотонейронов разгибателей и одновременно торможению альфамотонейронов мышц сгибателей. Благодаря этому меняется тонус мышц верхних и нижних конечностей — и сохраняется равновесие
; б —") Децеребрационная ригидность. а — кошка с выключенными большими полушариями (путем перевязки сосудов); б — то же при одновременном выключении мозжечка (по Дэвису и Поллоку, из И. С. Беритова).
Децеребрационная ригидность. а — кошка с выключенными большими полушариями (путем перевязки сосудов); б — то же при одновременном выключении мозжечка (по Дэвису и Поллоку, из И. С. Беритова).
 Статические рефлексы Их ролью яв-ся сохранение естественной позы человека и животных - Это тонические рефлексы в покое (человек стоит и сидит) - В том числе и при поворотах и наклонах головы и движения рук, а сидя – и ног, вызывающих смещение центра тяжести. Осуществляются с участием: - Сегментарных и надсегментарных (шейных тонических рефлексов СМ. ) - Рефлексов ствола мозга, возникающих с вестибулорецепторов Р. Магнус: - для выполнения роли шейных рефлексов разрушал у животного вестибулярный аппарат, чтобы устранить вестибулярные рефлексы. - Для выяснения роли вестибулярных рефлексов, на шею животного, расположенную на одной линии с туловищем, накладывали гипсовую повязку, чтобы прекратить шейные надсегментарные рефлексы. Вестибулярные рефлексы выявляются при поворачивании животного через ось висков. - При горизонтальном расположении животного спиной и затылком вниз поворот головы от 0 до 45 градусов выше горизонтали приводит к максимальному повышению тонуса мышц-разгибателей. - При дальнейшем повороте, если оно располагается спиной и затылком вверх по горизонтале и до -45 градусов ниже горизонтали, то тонус разгибателей становиться минимальным. У человека, в положении стоя (голова вверх, шея наклонена несколько вперед, руки вдоль туловища) шейные и лабиринтные рефлексы усиливают друга, и происходит повышение тонуса разгибателей нижних конечностей и сгибателей верхних конечностей.
Статические рефлексы Их ролью яв-ся сохранение естественной позы человека и животных - Это тонические рефлексы в покое (человек стоит и сидит) - В том числе и при поворотах и наклонах головы и движения рук, а сидя – и ног, вызывающих смещение центра тяжести. Осуществляются с участием: - Сегментарных и надсегментарных (шейных тонических рефлексов СМ. ) - Рефлексов ствола мозга, возникающих с вестибулорецепторов Р. Магнус: - для выполнения роли шейных рефлексов разрушал у животного вестибулярный аппарат, чтобы устранить вестибулярные рефлексы. - Для выяснения роли вестибулярных рефлексов, на шею животного, расположенную на одной линии с туловищем, накладывали гипсовую повязку, чтобы прекратить шейные надсегментарные рефлексы. Вестибулярные рефлексы выявляются при поворачивании животного через ось висков. - При горизонтальном расположении животного спиной и затылком вниз поворот головы от 0 до 45 градусов выше горизонтали приводит к максимальному повышению тонуса мышц-разгибателей. - При дальнейшем повороте, если оно располагается спиной и затылком вверх по горизонтале и до -45 градусов ниже горизонтали, то тонус разгибателей становиться минимальным. У человека, в положении стоя (голова вверх, шея наклонена несколько вперед, руки вдоль туловища) шейные и лабиринтные рефлексы усиливают друга, и происходит повышение тонуса разгибателей нижних конечностей и сгибателей верхних конечностей.
 и максимум (б) экстензорного тонуса (по Магнусу).") Положения головы, вызывающие минимум (а) и максимум (б) экстензорного тонуса (по Магнусу).
Положения головы, вызывающие минимум (а) и максимум (б) экстензорного тонуса (по Магнусу).
 При положении головы, отметенном 360 и 180°, тонус разгибателей минимален; он дохо дитдо. максимума при положениях головы от 65 до 100° и от 290 до 235° (цифры указывают градусы по отношению к воображаемому диаметру круга, соединяющего цифры 360 и 180°) (по Радемакеру, из И. С. Беритова).
При положении головы, отметенном 360 и 180°, тонус разгибателей минимален; он дохо дитдо. максимума при положениях головы от 65 до 100° и от 290 до 235° (цифры указывают градусы по отношению к воображаемому диаметру круга, соединяющего цифры 360 и 180°) (по Радемакеру, из И. С. Беритова).
 Статокинетические рефлексы • Их роль – восстановление нарушенной позы Это рефлекторное перераспределение мышечного тонуса, возникающее с: Кожных Проприо – (шея) Вестибулорецепторов При нарушении позы (организм находится в горизонтальном положении). К этим рефлексам относятся выпрямительные (установочные) рефлексы - выпрямление головы и туловища Выпрямление головы запускается с: Вестибулярных и кожных рецепторов Выпрямление туловища запускается с: Проприорецепторов шеи и кожных рецепторов. Т. е. имется две рефлесогенных зоны, поэтому выключение одной из зон (или рецепторов кожи или вестибулярного аппарата) у мезэнцефального организма не нарушает выпрямительные рефлексы. Выключение обеих названных рефлексогенных зон приводит к потере выпрямительных рефлексов. Это надсегментарные рефлексы ствола мозга и спинного мозга. Особо важную роль играют Красные ядра – при их разрушении нарушенная поза не восстанавливается. У человека и обезьян выпрямительные рефлексы осуществляются при обязательном участии КБП – ее выключение приводит к потере этих рефлексов. Р. Магнус – относил эти рефлексы – к статическим, однако они имеют две состовляющие – статическую (нет перемещения тела в пространстве или вращения вокруг своей оси на месте) и кинетическую – животное поднимается и занимает естественную позу.
Статокинетические рефлексы • Их роль – восстановление нарушенной позы Это рефлекторное перераспределение мышечного тонуса, возникающее с: Кожных Проприо – (шея) Вестибулорецепторов При нарушении позы (организм находится в горизонтальном положении). К этим рефлексам относятся выпрямительные (установочные) рефлексы - выпрямление головы и туловища Выпрямление головы запускается с: Вестибулярных и кожных рецепторов Выпрямление туловища запускается с: Проприорецепторов шеи и кожных рецепторов. Т. е. имется две рефлесогенных зоны, поэтому выключение одной из зон (или рецепторов кожи или вестибулярного аппарата) у мезэнцефального организма не нарушает выпрямительные рефлексы. Выключение обеих названных рефлексогенных зон приводит к потере выпрямительных рефлексов. Это надсегментарные рефлексы ствола мозга и спинного мозга. Особо важную роль играют Красные ядра – при их разрушении нарушенная поза не восстанавливается. У человека и обезьян выпрямительные рефлексы осуществляются при обязательном участии КБП – ее выключение приводит к потере этих рефлексов. Р. Магнус – относил эти рефлексы – к статическим, однако они имеют две состовляющие – статическую (нет перемещения тела в пространстве или вращения вокруг своей оси на месте) и кинетическую – животное поднимается и занимает естественную позу.
 Поза лежащей на левом боку обезьяны, у которой кора мозга удалена при сохранении зрительных бугров и части стриопаллидарной системы. Если животное переложить на правую сторону, то разогнуты будут правые конечности, а согнуты левые. В сгибании кисти правой руки ясно виден «хватательный» рефлекс (по Фултоиу)
Поза лежащей на левом боку обезьяны, у которой кора мозга удалена при сохранении зрительных бугров и части стриопаллидарной системы. Если животное переложить на правую сторону, то разогнуты будут правые конечности, а согнуты левые. В сгибании кисти правой руки ясно виден «хватательный» рефлекс (по Фултоиу)
 Кинетические рефлексы Рефлекторное перераспределение мышечного тонуса при ходьбе и беге, вращательном движении на месте, при изменении скорости движения в транспорте и лифте Возникают с: • Проприорецепторов • Вестибулорецепторов Назначение этих рецепторов – сохранение естественной позы при движении, а сокращения мышц направлены на преодоление сил гравитации, изменяющихся в следствии ускорения или замедления движения Важную роль играет система зрения – в ходе вращения наблюдается поворот головы и нистагм глаз (пилообразные движения глаз)
Кинетические рефлексы Рефлекторное перераспределение мышечного тонуса при ходьбе и беге, вращательном движении на месте, при изменении скорости движения в транспорте и лифте Возникают с: • Проприорецепторов • Вестибулорецепторов Назначение этих рецепторов – сохранение естественной позы при движении, а сокращения мышц направлены на преодоление сил гравитации, изменяющихся в следствии ускорения или замедления движения Важную роль играет система зрения – в ходе вращения наблюдается поворот головы и нистагм глаз (пилообразные движения глаз)
 Рефлексы вращения – 1 Возникают с ампул полукружных каналов и включают в перераспределение тонуса мускулатуру тела и глазные мышцы. Важным для практической медицины является нистагм – непроизвольные ритмические двухфазные движения глаз, который имеет две фазы: • При ускорении вращения сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению (сохранение имеющегося поля зрения) • После этого они быстро отклоняются обратно – по ходу вращения (захват нового поля зрения) Медленный компонент вызывается с рецепторов полукружных каналов Быстрый компонент связан с влиянием центров зрительной (поле 19) и лобной (поле 8) коры, действующих через верхнее двухолмие, моторные центры III, IV, VI нервов, и ретикулярную формацию ствола (при наркозе он исчезает). Во врачебной практике направление нистагма определяют по быстрому компоненту. При замедлении вращения, напротив, медленное движение глаз направлено в сторону вращения, и быстрое – против направления вращения. Вид нистагма (горизонтальный, вертикальный, ротационный) – зависит от полукружного канала, находящегося в плоскости вращения и служит для оценки функционального состояния вестибулярного аппарата. У здорового человека глазной нистагм может возникать и без вращения: при длительной фиксации взгляда на близкорасположенном предмете, а также при крайнем отведении глазных яблок (более 45 градусов)
Рефлексы вращения – 1 Возникают с ампул полукружных каналов и включают в перераспределение тонуса мускулатуру тела и глазные мышцы. Важным для практической медицины является нистагм – непроизвольные ритмические двухфазные движения глаз, который имеет две фазы: • При ускорении вращения сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению (сохранение имеющегося поля зрения) • После этого они быстро отклоняются обратно – по ходу вращения (захват нового поля зрения) Медленный компонент вызывается с рецепторов полукружных каналов Быстрый компонент связан с влиянием центров зрительной (поле 19) и лобной (поле 8) коры, действующих через верхнее двухолмие, моторные центры III, IV, VI нервов, и ретикулярную формацию ствола (при наркозе он исчезает). Во врачебной практике направление нистагма определяют по быстрому компоненту. При замедлении вращения, напротив, медленное движение глаз направлено в сторону вращения, и быстрое – против направления вращения. Вид нистагма (горизонтальный, вертикальный, ротационный) – зависит от полукружного канала, находящегося в плоскости вращения и служит для оценки функционального состояния вестибулярного аппарата. У здорового человека глазной нистагм может возникать и без вращения: при длительной фиксации взгляда на близкорасположенном предмете, а также при крайнем отведении глазных яблок (более 45 градусов)
 Рефлексы вращения – 2 В реализации рефлексов вращательного движения, кроме зрительного, участвуют волосковые клетки полукружных каналов вестибулярного аппарата: - При вращении – эндолимфа по инерции «отстает» от движения костных стенок канала и в результате усиливается ее давление на тот конец канала, который противоположен направлению вращения. - При остановке ее давление возрастает в области другого конца канала. • Этот сигнал от вестибулярного анализатора идет через вестибулярные ядра на медиальный продольный пучок к ядрам III, IV и VI пары ЧМН и приводит к изменению положения в пространстве глазных яблок – что сохраняет ориентацию в пространстве. • Одновременно – через вестибулярный тракт меняется тонус мышц конечностей и туловища, что сохраняет равновесие.
Рефлексы вращения – 2 В реализации рефлексов вращательного движения, кроме зрительного, участвуют волосковые клетки полукружных каналов вестибулярного аппарата: - При вращении – эндолимфа по инерции «отстает» от движения костных стенок канала и в результате усиливается ее давление на тот конец канала, который противоположен направлению вращения. - При остановке ее давление возрастает в области другого конца канала. • Этот сигнал от вестибулярного анализатора идет через вестибулярные ядра на медиальный продольный пучок к ядрам III, IV и VI пары ЧМН и приводит к изменению положения в пространстве глазных яблок – что сохраняет ориентацию в пространстве. • Одновременно – через вестибулярный тракт меняется тонус мышц конечностей и туловища, что сохраняет равновесие.
 Лифтные рефлексы возникают при подъеме вверх – повышается тонус сгибателей, и животное приседает, а при опускании вниз – возрастает тонус разгибателей и животное как бы приподнимается. Кроме зрительных рецепторов, в этом рефлексе участвуют волосковые клетки преддверия вестибулярного аппарата, т. к. рефлексы сохраняются и при закрытых глазах – подъем в самолете. При подъеме тела – отолиты более обычного давят на волосковые клетки, а в конце подъема – их давление снижается.
Лифтные рефлексы возникают при подъеме вверх – повышается тонус сгибателей, и животное приседает, а при опускании вниз – возрастает тонус разгибателей и животное как бы приподнимается. Кроме зрительных рецепторов, в этом рефлексе участвуют волосковые клетки преддверия вестибулярного аппарата, т. к. рефлексы сохраняются и при закрытых глазах – подъем в самолете. При подъеме тела – отолиты более обычного давят на волосковые клетки, а в конце подъема – их давление снижается.
 Локомоторная функция ствола мозга • лежит в основе совокупности согласованных движений, с помощью которых организм перемещается в пространстве (ходьба). • Большая роль отводится клинообразному ретикулярному ядру в задненижнем отделе среднего мозга, т. н. локомоторную область среднего мозга, имеющую эффекторный выход к ядрам моста. • Так же в эту систему входит субмамиллярная зона гипоталамуса. У человека локомоция без участия КБП – невозможна.
Локомоторная функция ствола мозга • лежит в основе совокупности согласованных движений, с помощью которых организм перемещается в пространстве (ходьба). • Большая роль отводится клинообразному ретикулярному ядру в задненижнем отделе среднего мозга, т. н. локомоторную область среднего мозга, имеющую эффекторный выход к ядрам моста. • Так же в эту систему входит субмамиллярная зона гипоталамуса. У человека локомоция без участия КБП – невозможна.
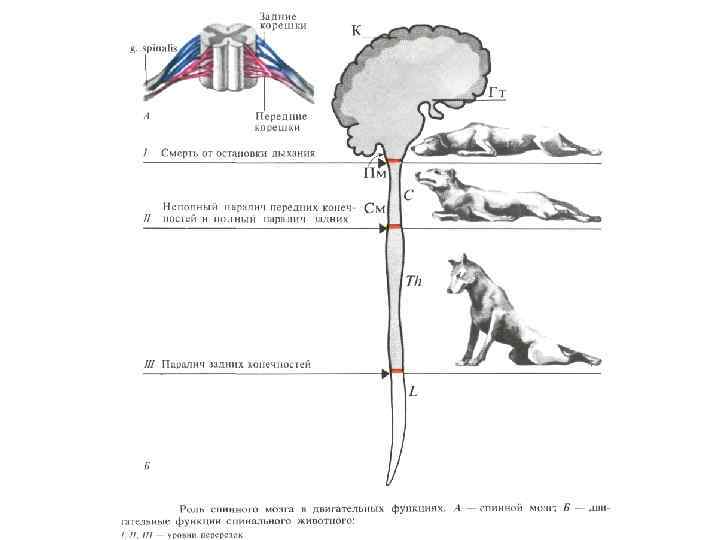
 ЖИВОТНЫЕ С ПЕРЕЗКОЙ НА РАЗЛИЧНЫХ УРОВНЯХ ЦНС Бульбарный организм (перерезка между мостом и средним мозгом) – развивается децеребрационная регидность, хотя имеются статические позно-тонические рефлексы шейные, лабиринтные), но отсутствуют выпрямительные рефлексы, локомоция, произвольные движения. Мезэнцефальный организм – имеется нормальный мышечный тонус, но у него отсутствуют произвольные движения.
ЖИВОТНЫЕ С ПЕРЕЗКОЙ НА РАЗЛИЧНЫХ УРОВНЯХ ЦНС Бульбарный организм (перерезка между мостом и средним мозгом) – развивается децеребрационная регидность, хотя имеются статические позно-тонические рефлексы шейные, лабиринтные), но отсутствуют выпрямительные рефлексы, локомоция, произвольные движения. Мезэнцефальный организм – имеется нормальный мышечный тонус, но у него отсутствуют произвольные движения.

 Характерные тонические рефлексы четырехлетнего ребенка, лишенного больших полушарий головного мозга
Характерные тонические рефлексы четырехлетнего ребенка, лишенного больших полушарий головного мозга
 и активную разгрузку. Вверху: испытуемый поддерживает рукой груз; угол") Сравнение реакций на пассивную (А) и активную разгрузку. Вверху: испытуемый поддерживает рукой груз; угол его локтевого сустава и нагрузка записываются. Внизу (кривые сверху вниз): электромиограммы (ЭМГ) левой и правой двуглавых мышц плеча; положение предплечья (угол сустава); нагрузка на него. Груз приподнимали последовательно 10 раз и усредняли данные. Красной стрелкой показан период снижения электромиографической активности. При пассивной разгрузке (А) оно рефлекторное, а при активной (6)– опережающее, т. е. происходит почти одновременно с сокращением левой двуглавой мышцы
Сравнение реакций на пассивную (А) и активную разгрузку. Вверху: испытуемый поддерживает рукой груз; угол его локтевого сустава и нагрузка записываются. Внизу (кривые сверху вниз): электромиограммы (ЭМГ) левой и правой двуглавых мышц плеча; положение предплечья (угол сустава); нагрузка на него. Груз приподнимали последовательно 10 раз и усредняли данные. Красной стрелкой показан период снижения электромиографической активности. При пассивной разгрузке (А) оно рефлекторное, а при активной (6)– опережающее, т. е. происходит почти одновременно с сокращением левой двуглавой мышцы
 Позно-тонические рефлексы у человека ярко выражены в первое полугодие жизни, так у грудных детей: • Лабиринтный тонический рефлекс – у ребенка лежащего на спине, повышен тонус разгибателей шеи, спины, ног. Если ребенок лежит на животе, то повышен тонус сгибателей шеи, спины, конечностей. • Симметричный шейный тонический рефлекс – при пассивном сгибании головы ребенка, лежащего на спине, происходит повышение тонуса сгибателей рук и повышение тонуса разгибателей ног. При разгибании головы наблюдаются противоположные процессы. • Ассиметричный шейный тонический рефлекс – у ребенка лежащего на спине голову поворачивают в сторону так, чтобы подбородок касался плеча, при этом снижается тонус сгибателей конечностей в сторону поворота и повышается тонус разгибателей. На противоположной стороне, на руке и ноге, возрастает тонус сгибателей. Все эти рефлексы тормозятся у детей ко 2 -3 месяцу, если они сохраняются в возрасте одного года, это может свидетельствовать о замедлении развития моторных центров головы.
Позно-тонические рефлексы у человека ярко выражены в первое полугодие жизни, так у грудных детей: • Лабиринтный тонический рефлекс – у ребенка лежащего на спине, повышен тонус разгибателей шеи, спины, ног. Если ребенок лежит на животе, то повышен тонус сгибателей шеи, спины, конечностей. • Симметричный шейный тонический рефлекс – при пассивном сгибании головы ребенка, лежащего на спине, происходит повышение тонуса сгибателей рук и повышение тонуса разгибателей ног. При разгибании головы наблюдаются противоположные процессы. • Ассиметричный шейный тонический рефлекс – у ребенка лежащего на спине голову поворачивают в сторону так, чтобы подбородок касался плеча, при этом снижается тонус сгибателей конечностей в сторону поворота и повышается тонус разгибателей. На противоположной стороне, на руке и ноге, возрастает тонус сгибателей. Все эти рефлексы тормозятся у детей ко 2 -3 месяцу, если они сохраняются в возрасте одного года, это может свидетельствовать о замедлении развития моторных центров головы.
 - Определяющий адекватную позу у плода,") В онтогенезе преобладает тонус мышц сгибателей (спинальный) - Определяющий адекватную позу у плода, т. е. максимальную сгибательную активность в суставах позвоночника и конечностей. - У новорожденного наличие примитивных сгибательных рефлексов (рефлекс Моро – симметрические обхватывающие движения ручек при действии различных стимулов) Появление разгибательного тонуса – обусловлено более поздним созреванием структур продолговатого мозга. - У детей, только на второй месяц появляется рефлекс Ландау – ребенок лежа на животе, приподнимает голову, опираясь на предплечья, а позже и на кисти. - на 5 -6 месяц, лежа на животе, ребенок разгибает ноги и приподнимает таз, что создает готовность к ползанью. Так постепенно флексорная поза, сменяется – экстензорной, при этом увеличивается пространство видимое глазами и обогащающая жизненный двигательный опыт ребенка.
В онтогенезе преобладает тонус мышц сгибателей (спинальный) - Определяющий адекватную позу у плода, т. е. максимальную сгибательную активность в суставах позвоночника и конечностей. - У новорожденного наличие примитивных сгибательных рефлексов (рефлекс Моро – симметрические обхватывающие движения ручек при действии различных стимулов) Появление разгибательного тонуса – обусловлено более поздним созреванием структур продолговатого мозга. - У детей, только на второй месяц появляется рефлекс Ландау – ребенок лежа на животе, приподнимает голову, опираясь на предплечья, а позже и на кисти. - на 5 -6 месяц, лежа на животе, ребенок разгибает ноги и приподнимает таз, что создает готовность к ползанью. Так постепенно флексорная поза, сменяется – экстензорной, при этом увеличивается пространство видимое глазами и обогащающая жизненный двигательный опыт ребенка.
") Сторожевой и ориентировочный рефлексы осуществляются с участием четвертохолмия, в ответ на зрительный сигнал (световой) или на звук происходит активация соответствующих рецепторов, сигнал достигает первичных центров (зрительные – передних или верхних бугров, слуховые – задние или нижние бугры), переключаются на тектоспинальный тракт – возникают ориентировочные рефлексы: поворот головы в сторону, откуда поступает сигнал и одновременно происходит перераспределение тонуса мышц, в том числе за счет изменении положения головы, благодаря которому сохраняется равновесие и возникают условия для совершения рефлекса «бег или борьба» .
Сторожевой и ориентировочный рефлексы осуществляются с участием четвертохолмия, в ответ на зрительный сигнал (световой) или на звук происходит активация соответствующих рецепторов, сигнал достигает первичных центров (зрительные – передних или верхних бугров, слуховые – задние или нижние бугры), переключаются на тектоспинальный тракт – возникают ориентировочные рефлексы: поворот головы в сторону, откуда поступает сигнал и одновременно происходит перераспределение тонуса мышц, в том числе за счет изменении положения головы, благодаря которому сохраняется равновесие и возникают условия для совершения рефлекса «бег или борьба» .
 Роль ЧМН ствола • С участием ядер V, IX, X, XI и XII пар осуществляется произвольные движения типа жевания (V пара) и глотания (последовательное возбуждение αмотонейронов языкоглоточного, вагуса, добавочного и подъязычного нервов). Повреждение – нарушаются акты жевания и глотания. • С участием нейронов IX и X пары идет управление голосовыми связками, а с помощью XII пары – языком. Повреждение этих структур – нарушается фонация и артикуляция (речь).
Роль ЧМН ствола • С участием ядер V, IX, X, XI и XII пар осуществляется произвольные движения типа жевания (V пара) и глотания (последовательное возбуждение αмотонейронов языкоглоточного, вагуса, добавочного и подъязычного нервов). Повреждение – нарушаются акты жевания и глотания. • С участием нейронов IX и X пары идет управление голосовыми связками, а с помощью XII пары – языком. Повреждение этих структур – нарушается фонация и артикуляция (речь).
 пути являются частью проводникового отдела анализаторов передающих информацию") Проводниковая функция ствола мозга Восходящие (афферентные) пути являются частью проводникового отдела анализаторов передающих информацию в сенсорную зону КБП. Выделяют: • Специфическую • Неспецифическую Специфическая – составляет лемнискоталамический путь, в котором выделяют: • Медиальную петлю • Латеральную петлю • Функционально-проводящие пути (т. к. не входит в состав лемнисков) зрительного анализатора – переключаются в латеральном коленчатом теле таламуса.
Проводниковая функция ствола мозга Восходящие (афферентные) пути являются частью проводникового отдела анализаторов передающих информацию в сенсорную зону КБП. Выделяют: • Специфическую • Неспецифическую Специфическая – составляет лемнискоталамический путь, в котором выделяют: • Медиальную петлю • Латеральную петлю • Функционально-проводящие пути (т. к. не входит в состав лемнисков) зрительного анализатора – переключаются в латеральном коленчатом теле таламуса.
Тонкого (Голля) ядра 2) Клиновидного (Бурдаха)") Медиальная петля Преимущественно образуется из аксонов нейронов ядер: 1)Тонкого (Голля) ядра 2) Клиновидного (Бурдаха) ядра • (1+2 - проводят информацию от рецепторов конечностей, туловища и шеи, тактильную и висцеральную чувствительность) 3)Тергеминоталамический путь (от ядер V нерва) проводит проприоцептивную и тактильную чувствительность от головы 4) Одиночного пути на подходе к таламусу – проводят вкусовую и висцеральную чувствительность 5) Бокового спиноталамического пути (небольшая часть) – проводят острую (быструю) локализованную боль. Волокна этой петли – переключаются преимущественно в вентральных задних специфических ядрах таламуса.
Медиальная петля Преимущественно образуется из аксонов нейронов ядер: 1)Тонкого (Голля) ядра 2) Клиновидного (Бурдаха) ядра • (1+2 - проводят информацию от рецепторов конечностей, туловища и шеи, тактильную и висцеральную чувствительность) 3)Тергеминоталамический путь (от ядер V нерва) проводит проприоцептивную и тактильную чувствительность от головы 4) Одиночного пути на подходе к таламусу – проводят вкусовую и висцеральную чувствительность 5) Бокового спиноталамического пути (небольшая часть) – проводят острую (быструю) локализованную боль. Волокна этой петли – переключаются преимущественно в вентральных задних специфических ядрах таламуса.
 Сформирована аксонами он нейронов ядер: • Трапецевидного тела • Верхней") Латеральная петля (слуховая петля) Сформирована аксонами он нейронов ядер: • Трапецевидного тела • Верхней оливы моста Входящих в проводниковый отдел слухового анализатора, переключается в медиальном коленчатом теле таламуса. Функционально – обеспечивает быструю передачу информации с четким различением свойств раздражителя, с формированием в КБП первичного ответа вызванных потенциалов соответствующего анализатора
Латеральная петля (слуховая петля) Сформирована аксонами он нейронов ядер: • Трапецевидного тела • Верхней оливы моста Входящих в проводниковый отдел слухового анализатора, переключается в медиальном коленчатом теле таламуса. Функционально – обеспечивает быструю передачу информации с четким различением свойств раздражителя, с формированием в КБП первичного ответа вызванных потенциалов соответствующего анализатора
 – преимущественно волокна: • Бокового спиноталомического пути • Спиноретикулярного пути (переключаются в") Неспецифическая (экстралемнисковая) – преимущественно волокна: • Бокового спиноталомического пути • Спиноретикулярного пути (переключаются в ядрах РФ) • Часть волокон тригеминоталомического пути Проводят температурную и болевую чувствительность. Переключаются в неспецифических (интраламинарных и ретикулярном) ядрах таламуса, прое 5 цируются в различные области КБП, особенно в лобную орбитальную кору. Функционально – обеспечивают медленную передачу информации нечеткой локализацией раздражителя и плохим анализом его свойств, с формированием в КБП вторичного ответа вызванных потенциалов. Неспецифическая система – получает коллатерали от специфической системы, что обеспечивает тесную связь этих двух афферентных систем.
Неспецифическая (экстралемнисковая) – преимущественно волокна: • Бокового спиноталомического пути • Спиноретикулярного пути (переключаются в ядрах РФ) • Часть волокон тригеминоталомического пути Проводят температурную и болевую чувствительность. Переключаются в неспецифических (интраламинарных и ретикулярном) ядрах таламуса, прое 5 цируются в различные области КБП, особенно в лобную орбитальную кору. Функционально – обеспечивают медленную передачу информации нечеткой локализацией раздражителя и плохим анализом его свойств, с формированием в КБП вторичного ответа вызванных потенциалов. Неспецифическая система – получает коллатерали от специфической системы, что обеспечивает тесную связь этих двух афферентных систем.
 системе ствола также относятся пути передающие сенсорную информацию в кору мозжечка:") К афферентной (проводящей) системе ствола также относятся пути передающие сенсорную информацию в кору мозжечка: • Дорсальный спиномозжечковый путь – от рецепторов мышц и связок • Вентральный спиномозжечковый путь – от рецепторов сухожилий, кожи и внутренних органов • Вестибуломозжечковый путь – от вестибулярных рецепторов Из коры мозжечка – к его зубчатым и промежуточным ядрам, далее по церебеллоталамическому пути к двигательным вентральным ядрам таламуса и после переключения в них в соматосенсорную, моторную и премоторные зоны КБП.
К афферентной (проводящей) системе ствола также относятся пути передающие сенсорную информацию в кору мозжечка: • Дорсальный спиномозжечковый путь – от рецепторов мышц и связок • Вентральный спиномозжечковый путь – от рецепторов сухожилий, кожи и внутренних органов • Вестибуломозжечковый путь – от вестибулярных рецепторов Из коры мозжечка – к его зубчатым и промежуточным ядрам, далее по церебеллоталамическому пути к двигательным вентральным ядрам таламуса и после переключения в них в соматосенсорную, моторную и премоторные зоны КБП.
 путь – от клеток") Нисходящие проводниковые пути ствола мозга Функционально объединяются: • Пирамидный (двигательный) путь – от клеток Беца КПБ прецентральной извилины – переключаются на мотонейроны передних рогов спинного мозга (кортико-спинальный) путь или двигательных ядер ЧМН (кортикобульбарный путь) – обеспечивают произвольные сокращения мышц конечностей, туловища, шеи и головы. • Экстрапирамидные – кортико-рубральный и кортико-ретикулярный – оканчиваются на моторных центрах ствола – от которых идут руброспинальный и ретикулоспинальный пути. Основная функция – произвольная регуляция мышечного тонуса, позы и равновесия. • Кортико-мостомозжечковый путь – в мозжечок поступает информация от двигательной и других областей КБП – обработанная в коре мозжечка и его ядрах информация поступает на моторные ядра ствола (красное, вестибулярные, ретикулярные), что выключает мозжечок в функции экстрапирамидной системы. • Тектоспинальный путь – от ядер четвертохолмия – обеспечивает двигательные реакции организма в ориентировочных, зрительных и слуховых рефлексах.
Нисходящие проводниковые пути ствола мозга Функционально объединяются: • Пирамидный (двигательный) путь – от клеток Беца КПБ прецентральной извилины – переключаются на мотонейроны передних рогов спинного мозга (кортико-спинальный) путь или двигательных ядер ЧМН (кортикобульбарный путь) – обеспечивают произвольные сокращения мышц конечностей, туловища, шеи и головы. • Экстрапирамидные – кортико-рубральный и кортико-ретикулярный – оканчиваются на моторных центрах ствола – от которых идут руброспинальный и ретикулоспинальный пути. Основная функция – произвольная регуляция мышечного тонуса, позы и равновесия. • Кортико-мостомозжечковый путь – в мозжечок поступает информация от двигательной и других областей КБП – обработанная в коре мозжечка и его ядрах информация поступает на моторные ядра ствола (красное, вестибулярные, ретикулярные), что выключает мозжечок в функции экстрапирамидной системы. • Тектоспинальный путь – от ядер четвертохолмия – обеспечивает двигательные реакции организма в ориентировочных, зрительных и слуховых рефлексах.
 Нейрогуморальная функция ствола мозга осуществляется за счет выработки нейронами нейромодуляторов, которые действуют как «регуляторы нейромодуляторов» • Норадренергическая система ствола мозга – медиатор норадреналин, имеется в двух ядрах: 1. Голубом пятне среднего мозга 2. Вентральное ядро ретикулярной формации продолговатого мозга Главной структурой этой системы является голубое пятно, содержащее более 50 % всех норадренергических нейронов ствола, имеющих обширные афферентные и эфферентные связи со всеми отделами ЦНС. Учувствуют в контроле проведения болевой и неболевой чувствительности, активности моторных систем, в регуляции деятельности висцеральных систем, эмоционального состояния тоски и страха, механизмов памяти и внимания, фаз цикла «сон – бодрствование» , индуцируя фазу быстрого сна, в формировании (через гипоталамус) центрального звена стрессформирующей системы, участвуя в развитии общего адаптационного синдрома.
Нейрогуморальная функция ствола мозга осуществляется за счет выработки нейронами нейромодуляторов, которые действуют как «регуляторы нейромодуляторов» • Норадренергическая система ствола мозга – медиатор норадреналин, имеется в двух ядрах: 1. Голубом пятне среднего мозга 2. Вентральное ядро ретикулярной формации продолговатого мозга Главной структурой этой системы является голубое пятно, содержащее более 50 % всех норадренергических нейронов ствола, имеющих обширные афферентные и эфферентные связи со всеми отделами ЦНС. Учувствуют в контроле проведения болевой и неболевой чувствительности, активности моторных систем, в регуляции деятельности висцеральных систем, эмоционального состояния тоски и страха, механизмов памяти и внимания, фаз цикла «сон – бодрствование» , индуцируя фазу быстрого сна, в формировании (через гипоталамус) центрального звена стрессформирующей системы, участвуя в развитии общего адаптационного синдрома.
 Серотонинергическая система ствола мозга • образована скоплением серотонинергических нейронов, образующих ядра шва, центральное серое вещество и ретикулярные магноцеллюлярные ядра. • участвует в контроле проведения болевой чувствительности, в регуляции цикла «сон – бодрствование» , потребления пищи и воды, тормозит сексуальное и агрессивное поведение. Дофаминергическая система ствола мозга • Представлена - черным веществом среднего мозга, в функциональном плане относится к стриопаллидарной системе. Нейроны черного вещества (медиатор дофамин) оказывают влияние на полосатое тело приводя к двигательным расстройствам типа Паркинсонизм.
Серотонинергическая система ствола мозга • образована скоплением серотонинергических нейронов, образующих ядра шва, центральное серое вещество и ретикулярные магноцеллюлярные ядра. • участвует в контроле проведения болевой чувствительности, в регуляции цикла «сон – бодрствование» , потребления пищи и воды, тормозит сексуальное и агрессивное поведение. Дофаминергическая система ствола мозга • Представлена - черным веществом среднего мозга, в функциональном плане относится к стриопаллидарной системе. Нейроны черного вещества (медиатор дофамин) оказывают влияние на полосатое тело приводя к двигательным расстройствам типа Паркинсонизм.
, нейроны которой расположены диффузно или") Интегративная функция ствола мозга Осуществляется его ретикулярной формацией (РФ), нейроны которой расположены диффузно или в виде ядер в центральных отделах ствола мозга: • Нейроны РФ имеют длинные, мало ветвящиеся дендриты и хорошо ветвящиеся аксоны, которые могут устанавливать контакты с 25 тыс. других нейронов • Нейроны РФ принимают информацию от различных сенсорных путей, они полисенсорные, имеют большие рецепторные поля • Имеют длительный латентный период на периферическую стимуляцию, за счет многочисленных синапсов • Фоновоактивны (5 -10 Гц), высоковозбудимы и лабильны • Чувствительны к веществам крови – адреналину, СО 2 и лекарственным препаратам – барбитураты и аминазин
Интегративная функция ствола мозга Осуществляется его ретикулярной формацией (РФ), нейроны которой расположены диффузно или в виде ядер в центральных отделах ствола мозга: • Нейроны РФ имеют длинные, мало ветвящиеся дендриты и хорошо ветвящиеся аксоны, которые могут устанавливать контакты с 25 тыс. других нейронов • Нейроны РФ принимают информацию от различных сенсорных путей, они полисенсорные, имеют большие рецепторные поля • Имеют длительный латентный период на периферическую стимуляцию, за счет многочисленных синапсов • Фоновоактивны (5 -10 Гц), высоковозбудимы и лабильны • Чувствительны к веществам крови – адреналину, СО 2 и лекарственным препаратам – барбитураты и аминазин
 Ретикулярная формация среднего мозга
Ретикулярная формация среднего мозга
 Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
 • • • Основные системы ретикулярной формации АФФЕРЕНТНЫЕ СИСТЕМЫ: от спинного мозга, от мозжечка, от четверохолмия, от коры, от чувствительных ядер черепномозговых нервов ЭФФЕРЕНТНЫЕ СИСТЕМЫ: Восходящая активирующая система неспецифическое тонизирование через синапсы на дентритах нейронов I и II слоев коры • Нисходящие ретикулоспинальные системы: - облегчающая - тормозящая
• • • Основные системы ретикулярной формации АФФЕРЕНТНЫЕ СИСТЕМЫ: от спинного мозга, от мозжечка, от четверохолмия, от коры, от чувствительных ядер черепномозговых нервов ЭФФЕРЕНТНЫЕ СИСТЕМЫ: Восходящая активирующая система неспецифическое тонизирование через синапсы на дентритах нейронов I и II слоев коры • Нисходящие ретикулоспинальные системы: - облегчающая - тормозящая
 Афферентные входы РФ Преимущественно образованы от 3 источников: • От температурных и болевых рецепторов по волокнам спиноретикулярного пути и тройничного нерва. • От моторных и сенсорных областей КБП по кортикоретикулярным путям импульсация идет в ядра, дающие начало ретикулоспинальным путям, а так же проецируется в мозжечок • От ядер мозжечка по мозжечковоретикулярным путям
Афферентные входы РФ Преимущественно образованы от 3 источников: • От температурных и болевых рецепторов по волокнам спиноретикулярного пути и тройничного нерва. • От моторных и сенсорных областей КБП по кортикоретикулярным путям импульсация идет в ядра, дающие начало ретикулоспинальным путям, а так же проецируется в мозжечок • От ядер мозжечка по мозжечковоретикулярным путям
 Эфферентные выходы РФ проецируются: • В спинной мозг по латеральному и медиальному ретикулоспинальным путям. • К неспецифическим ядрам таламуса, заднему гипоталамусу, полосатому телу по восходящим путям, которые начинаются в ядрах РФ продолговатого мозга и моста. • К мозжечку, по путям, которые начинаются в латеральном и парамедиальном ретикулярных ядрах и в ядре покрышки моста.
Эфферентные выходы РФ проецируются: • В спинной мозг по латеральному и медиальному ретикулоспинальным путям. • К неспецифическим ядрам таламуса, заднему гипоталамусу, полосатому телу по восходящим путям, которые начинаются в ядрах РФ продолговатого мозга и моста. • К мозжечку, по путям, которые начинаются в латеральном и парамедиальном ретикулярных ядрах и в ядре покрышки моста.
 Интеграция соматических функций: • Проявляется в координирующем влиянии РФ, посредством которой отдельные рефлексы ствола мозга объединяются в сложные двигательные акты. • Имеет специализированные ядра, участвующие в осуществлении сложных форм двигательного поведения: жевания, глотания, содружественного движения глаз в горизонтальном и вертикальном направлениях, локомоции, пения и др. (генератор ритма жевания) • Осуществляют контроль передачи сенсорной информации на различных уровнях ЦНС, так: • На уровне первичных афферентов нейронов V (тройничного) ядра передача информации может облегчатся или тормозиться при раздражении ретикулярного гигантоклеточного ядра продолговатого мозга • В ядрах задних столбов СМ сенсорный поток контролируются тормозными влияниями из РФ продолговатого мозга. • Восходящая РФ контролирует передачу информации в сенсорных специфических ядрах таламуса, определяя объем и качество передаваемой в головной мозг информации
Интеграция соматических функций: • Проявляется в координирующем влиянии РФ, посредством которой отдельные рефлексы ствола мозга объединяются в сложные двигательные акты. • Имеет специализированные ядра, участвующие в осуществлении сложных форм двигательного поведения: жевания, глотания, содружественного движения глаз в горизонтальном и вертикальном направлениях, локомоции, пения и др. (генератор ритма жевания) • Осуществляют контроль передачи сенсорной информации на различных уровнях ЦНС, так: • На уровне первичных афферентов нейронов V (тройничного) ядра передача информации может облегчатся или тормозиться при раздражении ретикулярного гигантоклеточного ядра продолговатого мозга • В ядрах задних столбов СМ сенсорный поток контролируются тормозными влияниями из РФ продолговатого мозга. • Восходящая РФ контролирует передачу информации в сенсорных специфических ядрах таламуса, определяя объем и качество передаваемой в головной мозг информации
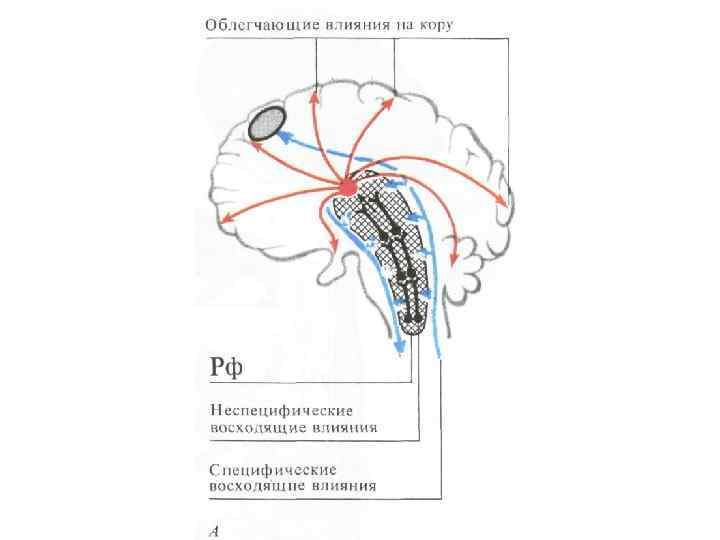
 Восходящие влияния РФ на большой мозг Может быть активирующим и тормозным, реализуется 5 -17% нейронов РФ, формирующими ретиколоталамические пути, а после переключения в таламусе сенсорный поток проектируется в различные области коры. - кроме таламуса восходящее влияния идут в задний гипоталамус, полосатое тело, прозрачную перегородку. Выделяют две части - ростральную – образуемую нейронами РФ среднего мозга - каудальную – образуемую нейронами РФ продолговатого мозга и моста
Восходящие влияния РФ на большой мозг Может быть активирующим и тормозным, реализуется 5 -17% нейронов РФ, формирующими ретиколоталамические пути, а после переключения в таламусе сенсорный поток проектируется в различные области коры. - кроме таламуса восходящее влияния идут в задний гипоталамус, полосатое тело, прозрачную перегородку. Выделяют две части - ростральную – образуемую нейронами РФ среднего мозга - каудальную – образуемую нейронами РФ продолговатого мозга и моста
 и тормозящих (-) зон РФ и ее связи с корой и") Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи с корой и подкоркой ОБЛЕГЧАЮЩИЕ ПУТИ: 5 – спиноретикулярный, 6 -вестибулоспинальный ТОРМОЗЯЩИЕ ПУТИ: 1 – кортикоретикулярный, 2 - каудатоспинальный, 3 – мозжечковретикулярный, 4 -ретикулоспинальный
Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи с корой и подкоркой ОБЛЕГЧАЮЩИЕ ПУТИ: 5 – спиноретикулярный, 6 -вестибулоспинальный ТОРМОЗЯЩИЕ ПУТИ: 1 – кортикоретикулярный, 2 - каудатоспинальный, 3 – мозжечковретикулярный, 4 -ретикулоспинальный
 Схема восходящих влияний РФ на кору мозга Восходящая активирующая система РФ Первичный ответ специфической проекционной зоны Вторичный ответ в зонах неспецифической стимуляции
Схема восходящих влияний РФ на кору мозга Восходящая активирующая система РФ Первичный ответ специфической проекционной зоны Вторичный ответ в зонах неспецифической стимуляции
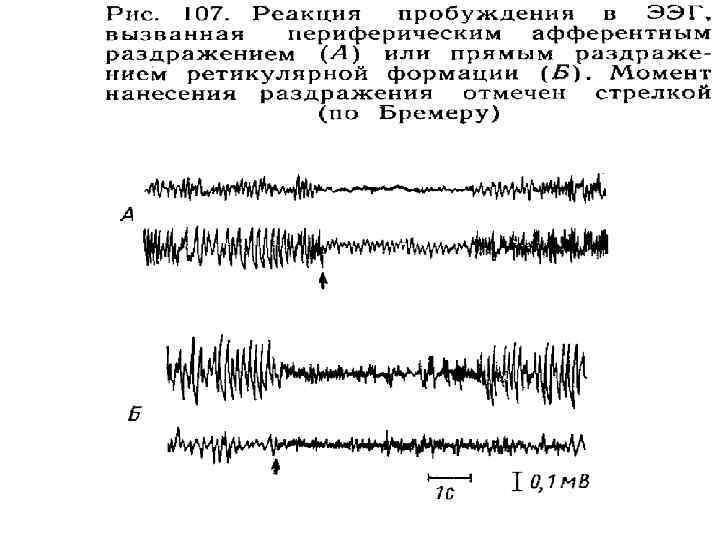
 –") Активирующее влияние на КБП оказывает ростральная часть РФ Опыт Бренера (1935 г. ) – перерезка ствола мозга между верхним и нижним холмиками четверохолмия вызывает снопоподобное состояние – нарушен контакт с внешним миром, отсутствие реакции на свет и звук ( «спящий мозг» ) Х. Мегун и Дж. Моруцци (1949 г. ) – раздражение РФ у спящих животных вызывало их пробуждение, появление ориентировочной реакции и появление высокочастотного β – ритма ЭЭГ (реакция десинхронизации), что доказывает генералзованное активирующее действие РФ на КБП. Важнейшая функция активирующей возходящей РФ яв-ся регуляция цикла «сон - бодрствование» и уровня сознания.
Активирующее влияние на КБП оказывает ростральная часть РФ Опыт Бренера (1935 г. ) – перерезка ствола мозга между верхним и нижним холмиками четверохолмия вызывает снопоподобное состояние – нарушен контакт с внешним миром, отсутствие реакции на свет и звук ( «спящий мозг» ) Х. Мегун и Дж. Моруцци (1949 г. ) – раздражение РФ у спящих животных вызывало их пробуждение, появление ориентировочной реакции и появление высокочастотного β – ритма ЭЭГ (реакция десинхронизации), что доказывает генералзованное активирующее действие РФ на КБП. Важнейшая функция активирующей возходящей РФ яв-ся регуляция цикла «сон - бодрствование» и уровня сознания.
 Тормозные влияния РФ на КБП оказывает её каудальный отдел, стимуляция его приводит к формированию на корковых нейронах ТПСП и появляется синхронизирующий эффект на ЭЭГ. Тормозное влияние РФ вызывает синхронизацию ЭЭГ в фазе «медленного» сна и атонию мышц во время «быстрого» сна. У бодрствующего животного – угнетает локомоторную функцию
Тормозные влияния РФ на КБП оказывает её каудальный отдел, стимуляция его приводит к формированию на корковых нейронах ТПСП и появляется синхронизирующий эффект на ЭЭГ. Тормозное влияние РФ вызывает синхронизацию ЭЭГ в фазе «медленного» сна и атонию мышц во время «быстрого» сна. У бодрствующего животного – угнетает локомоторную функцию
 Нисходящее влияния РФ на СМ - облегчающие активность моторных систем СМ, берут начало от всех отделов РФ. Пути, идущие от моста – тормозят активность мотонейронов мышц-сгибателей и активируют мотонейроны разгибателей. Пути, идущие от продолговатого мозга – тормозят разгибатели и активируют сгибатели. Раздражение РФ – вызывает тремор и повышение тонуса мышц, которое сохраняется длительное время (последействие).
Нисходящее влияния РФ на СМ - облегчающие активность моторных систем СМ, берут начало от всех отделов РФ. Пути, идущие от моста – тормозят активность мотонейронов мышц-сгибателей и активируют мотонейроны разгибателей. Пути, идущие от продолговатого мозга – тормозят разгибатели и активируют сгибатели. Раздражение РФ – вызывает тремор и повышение тонуса мышц, которое сохраняется длительное время (последействие).
 Схема нисходящих влияний РФ на мотонейроны РФ Ретикуло-спинальные пути Альфа-мотонейрон Активация Торможение Гамма мотонейрон
Схема нисходящих влияний РФ на мотонейроны РФ Ретикуло-спинальные пути Альфа-мотонейрон Активация Торможение Гамма мотонейрон
 Промежуточный мозг Состоит из таламической области и гипоталамуса. Таламическая область – включает таламус, коленчатые тела и эпиталамус (эпифиз) Таламус (зрительный бугор) 1. Парный ядерный комплекс (до 60 ядер) 2. Ядра подразделяются на: релейные (специфические), ассоциативные, неспецифические 3. Выполняет 3 функции: переключающую, интегративную, модулирующую.
Промежуточный мозг Состоит из таламической области и гипоталамуса. Таламическая область – включает таламус, коленчатые тела и эпиталамус (эпифиз) Таламус (зрительный бугор) 1. Парный ядерный комплекс (до 60 ядер) 2. Ядра подразделяются на: релейные (специфические), ассоциативные, неспецифические 3. Выполняет 3 функции: переключающую, интегративную, модулирующую.
 Основные ядра таламуса Специфические ядра • переключающие ассоциативные моторные Неспецифические ядра • срединные ядра, надколенное ядро, пограничное ядро, парафасцикулярное ядро, ретикулярное ядро (проекция к полосатому телу и V -VI слоям всех областей коры больших полушарий)
Основные ядра таламуса Специфические ядра • переключающие ассоциативные моторные Неспецифические ядра • срединные ядра, надколенное ядро, пограничное ядро, парафасцикулярное ядро, ретикулярное ядро (проекция к полосатому телу и V -VI слоям всех областей коры больших полушарий)
 и неспецифических (б) ядер таламуса в кору больших полушарий 1 —") Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших полушарий 1 — сенсорные афферентные пути, 2 — ретикулярно— таламические пути, 3 — проекционная зона коры, 4 — ассоциативная зона коры
Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших полушарий 1 — сенсорные афферентные пути, 2 — ретикулярно— таламические пути, 3 — проекционная зона коры, 4 — ассоциативная зона коры
 Сенсорные – переключают афферентную импульсацию в сенсорные") • • • Релейные ядра (специфические) Сенсорные – переключают афферентную импульсацию в сенсорные зоны КБП, перекодируют и обрабатывают информацию Несенсорные – переключают в КБП несенсорную импульсацию поступающую из разных отделов головного мозга Сенсорные ядра Выделяют 3 основных структуры: Вентральные задние ядра (вентробазальный комплекс) Латеральное коленчатое тело Медиальное коленчатое тело
• • • Релейные ядра (специфические) Сенсорные – переключают афферентную импульсацию в сенсорные зоны КБП, перекодируют и обрабатывают информацию Несенсорные – переключают в КБП несенсорную импульсацию поступающую из разных отделов головного мозга Сенсорные ядра Выделяют 3 основных структуры: Вентральные задние ядра (вентробазальный комплекс) Латеральное коленчатое тело Медиальное коленчатое тело
 Специфические ядра таламуса ПЕРЕКЛЮЧАЮЩИЕ АССОЦИАТИВНЫЕ МОТОРНЫЕ Вентробазальный комплекс (тактильная, проприоцептивная, температурная болевая, вкусовая информация в соматосенсорную тону кору больших полушарий) Внутреннее коленчатое ядро тело (слуховая информация в слуховые зоны коры Наружное коленчатое тело (зрительная информа ция в зрительные зоны коры) Медиодорсальное ядро (Проекция в лобные доли) Подушка (Проекция в теменную и височную кору) Заднелатеральное (Проекция в темен ную кору) переднее ядро (Проекция в лимбнчвскую кору) Переднецентральное и вентролатеральное ядра (Переключение) от мозжечка и базальных ганглиев в моторную коры больших полушарий)
Специфические ядра таламуса ПЕРЕКЛЮЧАЮЩИЕ АССОЦИАТИВНЫЕ МОТОРНЫЕ Вентробазальный комплекс (тактильная, проприоцептивная, температурная болевая, вкусовая информация в соматосенсорную тону кору больших полушарий) Внутреннее коленчатое ядро тело (слуховая информация в слуховые зоны коры Наружное коленчатое тело (зрительная информа ция в зрительные зоны коры) Медиодорсальное ядро (Проекция в лобные доли) Подушка (Проекция в теменную и височную кору) Заднелатеральное (Проекция в темен ную кору) переднее ядро (Проекция в лимбнчвскую кору) Переднецентральное и вентролатеральное ядра (Переключение) от мозжечка и базальных ганглиев в моторную коры больших полушарий)
 Ассоциативные системы таламуса • ТАЛАМОПАРИЕТАЛЬНАЯ: ОТ ЗАДНЕЛАТЕРАЛЬНОГО ЯДРА И ПОДУШКИ ТАЛАМУСА К ТЕМЕННОЙ И ВИСОЧНОЙ КОРЕ • ТАЛАМОФРОНТАЛЬНАЯ: ОТ МЕДИОДОРСАЛЬНОГО И ПЕРЕДНЕГО ЯДЕР ТАЛАМУСА К ФРОНТАЛЬНОЙ И ЛИМБИЧЕСКОЙ КОРЕ • Механизмы работы: • 1) Мультисенсорная конвергенция • 2) Пластическая перестройка при гетеромодальных сенсорных воздействиях: избирательное привыкание, сенситизация и эстраполяция • 3) Кратковременное хранение следов интеграции в виде реверберации импульсов
Ассоциативные системы таламуса • ТАЛАМОПАРИЕТАЛЬНАЯ: ОТ ЗАДНЕЛАТЕРАЛЬНОГО ЯДРА И ПОДУШКИ ТАЛАМУСА К ТЕМЕННОЙ И ВИСОЧНОЙ КОРЕ • ТАЛАМОФРОНТАЛЬНАЯ: ОТ МЕДИОДОРСАЛЬНОГО И ПЕРЕДНЕГО ЯДЕР ТАЛАМУСА К ФРОНТАЛЬНОЙ И ЛИМБИЧЕСКОЙ КОРЕ • Механизмы работы: • 1) Мультисенсорная конвергенция • 2) Пластическая перестройка при гетеромодальных сенсорных воздействиях: избирательное привыкание, сенситизация и эстраполяция • 3) Кратковременное хранение следов интеграции в виде реверберации импульсов
 Центральный аппарат анализа и") Основные функции ассоциативных систем таламуса • ТАЛАМОПАРИЕТАЛЬНАЯ СИСТЕМА • 1) Центральный аппарат анализа и синтеза обстановочной афферентации, запуска ориентационных движений глаз и туловища • 2) Один из центральных аппаратов «схемы тела» и сенсорного контроля текущей двигательной активности • 3) Аппарат формирования полимодальных образов • ТАЛАМОФРОНТАЛЬНАЯ СИСТЕМА • Корковый модулятор лимбической системы, программирование целенаправленных поведенческих актов на основе опыта и мотивации
Основные функции ассоциативных систем таламуса • ТАЛАМОПАРИЕТАЛЬНАЯ СИСТЕМА • 1) Центральный аппарат анализа и синтеза обстановочной афферентации, запуска ориентационных движений глаз и туловища • 2) Один из центральных аппаратов «схемы тела» и сенсорного контроля текущей двигательной активности • 3) Аппарат формирования полимодальных образов • ТАЛАМОФРОНТАЛЬНАЯ СИСТЕМА • Корковый модулятор лимбической системы, программирование целенаправленных поведенческих актов на основе опыта и мотивации
 Вентральные задние ядра • • • являются главными реле переключения соматосенсорноафферентной системы в составе волокон медиальной петли и примыкающей к ней волокнам других афферентных путей. Передается в соматосенсорную кору постцентральной извилины (поля 1 -3 по Бродману), где формируются соответствующие ощущения. В них переключаются: Тактильная Проприоцептивная Вкусовая Висцеральная Частично температурная и болевая В ядрах имеется топографическая проекция периферии, при этом функционально более тонко организованные части тела (язык, лицо) имеют большую зону представительства. Стимуляция ядер вызывает парастезии (ложные ощущения) в разных частях тела и нарушение «схемы тела» (искаженное восприятие частей тела в пространстве и относительно друга) Особенностью сенсорных ядер является точность передачи информации с минимальным искажением в КБП, по принципу «точка в точку»
Вентральные задние ядра • • • являются главными реле переключения соматосенсорноафферентной системы в составе волокон медиальной петли и примыкающей к ней волокнам других афферентных путей. Передается в соматосенсорную кору постцентральной извилины (поля 1 -3 по Бродману), где формируются соответствующие ощущения. В них переключаются: Тактильная Проприоцептивная Вкусовая Висцеральная Частично температурная и болевая В ядрах имеется топографическая проекция периферии, при этом функционально более тонко организованные части тела (язык, лицо) имеют большую зону представительства. Стимуляция ядер вызывает парастезии (ложные ощущения) в разных частях тела и нарушение «схемы тела» (искаженное восприятие частей тела в пространстве и относительно друга) Особенностью сенсорных ядер является точность передачи информации с минимальным искажением в КБП, по принципу «точка в точку»
 Реле для переключения зрительной импульсации в затылочную кору (поле 17),") Латеральное коленчатое тело (наружное) Реле для переключения зрительной импульсации в затылочную кору (поле 17), где используется для формирования зрительных ощущений, далее в лобные и теменные доли – ассоциативные зоны коры, где формируется ответ на вопрос «Что такое» , т. е. окончательное формирование образа. Медиальное коленчатое тело (внутреннее) • Реле для переключения слуховой и, возможно, вестибулярной импульсации в верхнюю часть височной коры (поля 41 и 42)
Латеральное коленчатое тело (наружное) Реле для переключения зрительной импульсации в затылочную кору (поле 17), где используется для формирования зрительных ощущений, далее в лобные и теменные доли – ассоциативные зоны коры, где формируется ответ на вопрос «Что такое» , т. е. окончательное формирование образа. Медиальное коленчатое тело (внутреннее) • Реле для переключения слуховой и, возможно, вестибулярной импульсации в верхнюю часть височной коры (поля 41 и 42)
 Несенсорные релейные ядра Передние и вентральные Передние ядра - Афферентные – поступают из мамиллярных тел гипоталамуса - Эффекторные – проектируются в лимбическую кору, а от туда в гиппокамп и опять к гипоталамусу, формируя нейронный круг, лежащий в основе возникновения эмоций (так называемые «лимбические» ядра таламуса) Вентральные ядра Участвуют в регуляции движения, выполняя моторную функцию. В них переключается информация от - базальных ганглиев - зубчатого ядра мозжечка - красного ядра которая затем проецируется в моторную и промоторную кору (поля 4 и 6), обеспечивая выполнение сложных двигательных программ, образованных в мозжечке и базальных ганглиях. Специфические ядра получают также нисходящие корковые волокна из своих проекционных зон, что лежит в основе кольцевых взаиморегулирующих отношений между корой и таламусом.
Несенсорные релейные ядра Передние и вентральные Передние ядра - Афферентные – поступают из мамиллярных тел гипоталамуса - Эффекторные – проектируются в лимбическую кору, а от туда в гиппокамп и опять к гипоталамусу, формируя нейронный круг, лежащий в основе возникновения эмоций (так называемые «лимбические» ядра таламуса) Вентральные ядра Участвуют в регуляции движения, выполняя моторную функцию. В них переключается информация от - базальных ганглиев - зубчатого ядра мозжечка - красного ядра которая затем проецируется в моторную и промоторную кору (поля 4 и 6), обеспечивая выполнение сложных двигательных программ, образованных в мозжечке и базальных ганглиях. Специфические ядра получают также нисходящие корковые волокна из своих проекционных зон, что лежит в основе кольцевых взаиморегулирующих отношений между корой и таламусом.
 Ассоциативные ядра таламуса Относится: - подушка - медиодорсальное ядро - латеральные ядра их функция интегрирующая, в объединении деятельности таламических ядер и различных зон ассоциативной КБП. Подушка - получает входы от коленчатых тел и неспецифических ядер - эфференты – в височно – теменно – затылочные зоны КБП, которые учавствуют в гностических (узнавание предметов, явлений), речевых и зрительных функций (интеграция слова и зрительного образа), а также в восприятии «схемы тела» . Медиодорсальное ядро - получает информацию от гипоталамуса, амигдалы, гипокампа, таламических ядер, центрального серого в-ва ствола. - эфференты – на ассоциативную лобную и лимбическую кору. Участвует в формировании эмоциональной и поведенческой двигательной активности и в запоминании. Разрушение – устраняет страх, тревогу, напряженность, страдание от боли, но возникает лобный синдром – снижение инициативы, безразличие и гипокинезия. Латеральные ядра Получают зрительную и слуховую импульсацию от коленчатых тел и от вентрикулярного ядра (соматосенсорная импульсация), далее она передается в ассоциативную теменную кору и используется в функции гнозиса, праксиса, «схемы тела» (т. н. таламотеменная система)
Ассоциативные ядра таламуса Относится: - подушка - медиодорсальное ядро - латеральные ядра их функция интегрирующая, в объединении деятельности таламических ядер и различных зон ассоциативной КБП. Подушка - получает входы от коленчатых тел и неспецифических ядер - эфференты – в височно – теменно – затылочные зоны КБП, которые учавствуют в гностических (узнавание предметов, явлений), речевых и зрительных функций (интеграция слова и зрительного образа), а также в восприятии «схемы тела» . Медиодорсальное ядро - получает информацию от гипоталамуса, амигдалы, гипокампа, таламических ядер, центрального серого в-ва ствола. - эфференты – на ассоциативную лобную и лимбическую кору. Участвует в формировании эмоциональной и поведенческой двигательной активности и в запоминании. Разрушение – устраняет страх, тревогу, напряженность, страдание от боли, но возникает лобный синдром – снижение инициативы, безразличие и гипокинезия. Латеральные ядра Получают зрительную и слуховую импульсацию от коленчатых тел и от вентрикулярного ядра (соматосенсорная импульсация), далее она передается в ассоциативную теменную кору и используется в функции гнозиса, праксиса, «схемы тела» (т. н. таламотеменная система)
 ФИЗИОЛОГИЯ МОЗЖЕЧКА
ФИЗИОЛОГИЯ МОЗЖЕЧКА
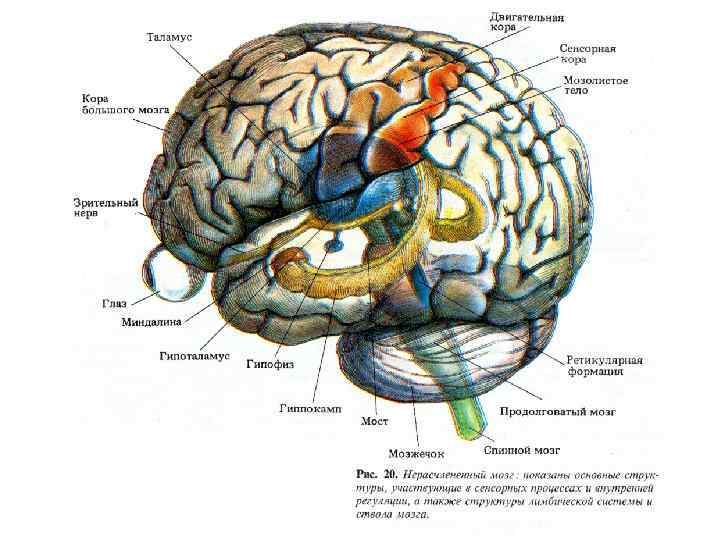
 СТРОЕНИЕ МОЗЖЕЧКА
СТРОЕНИЕ МОЗЖЕЧКА
 Схема областей коры мозжечка
Схема областей коры мозжечка
 Фазы последствия удаления мозжечка • 1. Фаза раздражения — связана с повреждением мозжечка, • У человека — длиться несколько суток и характеризуется параличом двигательных систем. • 2. Фаза выпадения функций — характеризуется резким нарушением координированности, пластичности движений и их точности. Сопровождается потерей способности к выполнению сложных двигательных актов (катание на коньках, выполнение спортивных движений и т. д. ) Длиться у человека до нескольких лет. • 3. Стадия компенсации — наступает через 3 — 5 лет и сопровождается постепенным исчезновением симптомов мозжечковой недостаточности второй фазы. Декортикация устраняет эту стадию, так разрушение лобных долей ведет к возникновению мозжечковых изменений (лобно — мозжечковый путь).
Фазы последствия удаления мозжечка • 1. Фаза раздражения — связана с повреждением мозжечка, • У человека — длиться несколько суток и характеризуется параличом двигательных систем. • 2. Фаза выпадения функций — характеризуется резким нарушением координированности, пластичности движений и их точности. Сопровождается потерей способности к выполнению сложных двигательных актов (катание на коньках, выполнение спортивных движений и т. д. ) Длиться у человека до нескольких лет. • 3. Стадия компенсации — наступает через 3 — 5 лет и сопровождается постепенным исчезновением симптомов мозжечковой недостаточности второй фазы. Декортикация устраняет эту стадию, так разрушение лобных долей ведет к возникновению мозжечковых изменений (лобно — мозжечковый путь).
 Симптомы фазы выпадения функций: • • • • • • 1. атония — снижение тонуса мышц 2. aстения — повышенная утомляемость при выполнении движений (причина нарушения трофики мышц) З. астезия — нарушение способности к слитному тетаническому сокращению (качанию головь, туловища, конечностей, затруднение стояния или сидения) 4. атаксия — нарушение точности и координированности движений. Тремор действия может переходить в размашистые колебания конечностей, а поставленная цель не достигается. Наблюдается неуверенная походка, с широко расставленными ногами и избыточными движениями, при которых больного как бы «бросает» из «стороны в сторону» ( «пьяная походка» ). Больной не может выполнить пальце-носовую, указательную или пяточно-коленную пробы. 5. асинергия — неспособность сесть из положения лежа в положение сидя без помощи рук. 6. дистония — непроизвольное повышение или понижение тонуса мышц. 7. дисметрия — расстройство равномерности движения выражающееся в излишнем, либо недостаточном движении. Больной пытается взять предмет со стола и проносит руку (гиперметрия), либо не доносит до предмета (гипометрия) 8. адиадохокинез — расстройства при работе антагонистических групп мышц (невозможным становится быстрое сгибание и разгибание пальцев, вращение ладоней вниз-вверх), неспособность выполнения быстрой последовательности движений. 9. деэквилибраиия —- нарушение равновесия (выявляется пробой Ронберта) 10. дизартрия — расстройства речевой моторики (скандированная речь)
Симптомы фазы выпадения функций: • • • • • • 1. атония — снижение тонуса мышц 2. aстения — повышенная утомляемость при выполнении движений (причина нарушения трофики мышц) З. астезия — нарушение способности к слитному тетаническому сокращению (качанию головь, туловища, конечностей, затруднение стояния или сидения) 4. атаксия — нарушение точности и координированности движений. Тремор действия может переходить в размашистые колебания конечностей, а поставленная цель не достигается. Наблюдается неуверенная походка, с широко расставленными ногами и избыточными движениями, при которых больного как бы «бросает» из «стороны в сторону» ( «пьяная походка» ). Больной не может выполнить пальце-носовую, указательную или пяточно-коленную пробы. 5. асинергия — неспособность сесть из положения лежа в положение сидя без помощи рук. 6. дистония — непроизвольное повышение или понижение тонуса мышц. 7. дисметрия — расстройство равномерности движения выражающееся в излишнем, либо недостаточном движении. Больной пытается взять предмет со стола и проносит руку (гиперметрия), либо не доносит до предмета (гипометрия) 8. адиадохокинез — расстройства при работе антагонистических групп мышц (невозможным становится быстрое сгибание и разгибание пальцев, вращение ладоней вниз-вверх), неспособность выполнения быстрой последовательности движений. 9. деэквилибраиия —- нарушение равновесия (выявляется пробой Ронберта) 10. дизартрия — расстройства речевой моторики (скандированная речь)
 СОМАТИЧЕСКАЯ ПРОЕКЦИЯ В КОРЕ ЧЕРВЯ И ПОЛУШАРИЯХ МОЗЖЕЧКА
СОМАТИЧЕСКАЯ ПРОЕКЦИЯ В КОРЕ ЧЕРВЯ И ПОЛУШАРИЯХ МОЗЖЕЧКА
 Нейронная организация коры мозжечка Вверху слева: схема поперечного среза через листок Внизу, главные синаптические контакты в листке
Нейронная организация коры мозжечка Вверху слева: схема поперечного среза через листок Внизу, главные синаптические контакты в листке
 КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ КОРЫ МОЗЖЕЧКА
КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ КОРЫ МОЗЖЕЧКА
 Мелкие рецептивные поля шести клеток Пуркинье, активность которых регистрировалас ь двумя микроэлектрода ми, введенными в дольки V и ПVI
Мелкие рецептивные поля шести клеток Пуркинье, активность которых регистрировалас ь двумя микроэлектрода ми, введенными в дольки V и ПVI
 Вестибулярных ядер") СВЯЗИ КОРЫ МОЗЖЕЧКА • АФФЕРЕНТНЫЕ СВЯЗИ - МОХОВИДНЫЕ ВОЛОКНА: от • 1) Вестибулярных ядер - вестибулоцеребеллярные тракты • 2) Спинного мозга - спиноцеребеллярные тракты • 3) Ретикулярной формации - ретикулоцеребеллярные тракты • 4) Коры больших полушарий - кортикоцеребеллярные тракты - ЛИАНОВИДНЫЕ ВОЛОКНА: от • Нижней оливы - клетки Пуркинье (1 волокно-1 клетка) • ЭФФЕРЕНТНЫЕ СВЯЗИ - К ПОДКОРКОВЫМ ЯДРАМ
СВЯЗИ КОРЫ МОЗЖЕЧКА • АФФЕРЕНТНЫЕ СВЯЗИ - МОХОВИДНЫЕ ВОЛОКНА: от • 1) Вестибулярных ядер - вестибулоцеребеллярные тракты • 2) Спинного мозга - спиноцеребеллярные тракты • 3) Ретикулярной формации - ретикулоцеребеллярные тракты • 4) Коры больших полушарий - кортикоцеребеллярные тракты - ЛИАНОВИДНЫЕ ВОЛОКНА: от • Нижней оливы - клетки Пуркинье (1 волокно-1 клетка) • ЭФФЕРЕНТНЫЕ СВЯЗИ - К ПОДКОРКОВЫМ ЯДРАМ
 Связи ядер мозжечка • АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА - ЗУБЧАТЫЕ ЯДРА: от коры полушарий - ВСТАВОЧНЫЕ ЯДРА (ПРОБКОВОЕ И ШАРОВИДНОЕ): от средней части коры - ЯДРО ШАТРА: от коры червя • ЭФФЕРЕНТНЫЕ СВЯЗИ ЯДЕР: - ЗУБЧАТЫЕ ЯДРА: к моторным ядрам таламуса и затем к двигательной зоне коры больших полушарий - ВСТАВОЧНЫЕ ЯДРА: к красным ядрам - ЯДРО ШАТРА: к ретикулярной формации и вестибуляр-ному ядру Дейтерса
Связи ядер мозжечка • АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА - ЗУБЧАТЫЕ ЯДРА: от коры полушарий - ВСТАВОЧНЫЕ ЯДРА (ПРОБКОВОЕ И ШАРОВИДНОЕ): от средней части коры - ЯДРО ШАТРА: от коры червя • ЭФФЕРЕНТНЫЕ СВЯЗИ ЯДЕР: - ЗУБЧАТЫЕ ЯДРА: к моторным ядрам таламуса и затем к двигательной зоне коры больших полушарий - ВСТАВОЧНЫЕ ЯДРА: к красным ядрам - ЯДРО ШАТРА: к ретикулярной формации и вестибуляр-ному ядру Дейтерса
 ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА • РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА, РАВНОВЕСИЯ, ПОДДЕРЖИВАЮЩИХ ДВИЖЕНИЙ. • КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВ-ЛЕННЫХ ДВИЖЕНИЙ И ИХ КООРДИНАЦИЯ С РЕФЛЕКСАМИ ПОДДЕРЖАНИЯ ПОЗЫ • ПРАВИЛЬНОЕ ВЫПОЛНЕНИЕ БЫСТРЫХ ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ ПО КОМАНДАМ КБП В СТРУКТУРЕ ОБЩЕЙ ПРОГРАММЫ ДВИЖЕНИЙ • Т. О. МОЗЖЕЧОК НЕ ТОЛЬКО МОДУЛИРУЕТ УЖЕ НАЧАТЫЕ ДВИГАТЕЛЬНЫЕ АКТЫ, НО И ОБЕСПЕЧИВАЕТ СОЗДАНИЕ И ХРАНЕНИЕ ИНДИВИДУАЛЬНЫХ ПРОГРАММ БЫСТРЫХ, ВЫСОКОКООРДИНИРОВАННЫХ ДВИЖЕНИЙ.
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА • РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА, РАВНОВЕСИЯ, ПОДДЕРЖИВАЮЩИХ ДВИЖЕНИЙ. • КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВ-ЛЕННЫХ ДВИЖЕНИЙ И ИХ КООРДИНАЦИЯ С РЕФЛЕКСАМИ ПОДДЕРЖАНИЯ ПОЗЫ • ПРАВИЛЬНОЕ ВЫПОЛНЕНИЕ БЫСТРЫХ ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ ПО КОМАНДАМ КБП В СТРУКТУРЕ ОБЩЕЙ ПРОГРАММЫ ДВИЖЕНИЙ • Т. О. МОЗЖЕЧОК НЕ ТОЛЬКО МОДУЛИРУЕТ УЖЕ НАЧАТЫЕ ДВИГАТЕЛЬНЫЕ АКТЫ, НО И ОБЕСПЕЧИВАЕТ СОЗДАНИЕ И ХРАНЕНИЕ ИНДИВИДУАЛЬНЫХ ПРОГРАММ БЫСТРЫХ, ВЫСОКОКООРДИНИРОВАННЫХ ДВИЖЕНИЙ.
 Упрощенная функциональная схема медиальных структур мозжечка
Упрощенная функциональная схема медиальных структур мозжечка
 и латеральной (справа) частей мозжечка") Главные связи медиальной (слева) и латеральной (справа) частей мозжечка
Главные связи медиальной (слева) и латеральной (справа) частей мозжечка
 Характер движений после удаления мозжечка У животных с удаленным или разрушенным мозжечком наблюдается: атония, астазия, астения, атаксия, абазия
Характер движений после удаления мозжечка У животных с удаленным или разрушенным мозжечком наблюдается: атония, астазия, астения, атаксия, абазия
 ЦИТАТА • «Как скульптор избирательно удаляет резцом все лишнее из первоначально бес-форменного камня, так и мозжечок, подавляя торможением лишние возбуждения, добивается четкой формы двигательной реакции» • Экклз, 1969
ЦИТАТА • «Как скульптор избирательно удаляет резцом все лишнее из первоначально бес-форменного камня, так и мозжечок, подавляя торможением лишние возбуждения, добивается четкой формы двигательной реакции» • Экклз, 1969
 ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА • ТРИАДА ЛЮЧИАНИ: атония, астазия, астения • ТРИАДА ШАРКО: нистагм, • тремор, • скандированная речь • АТАКСИЯ (пьяная походка) • ДИСМЕТРИЯ (избыточность) • ДИЗАРТРИЯ • ДИЗЭКВИЛИБРАЦИЯ • АДИАДОХОКИНЕЗ
ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА • ТРИАДА ЛЮЧИАНИ: атония, астазия, астения • ТРИАДА ШАРКО: нистагм, • тремор, • скандированная речь • АТАКСИЯ (пьяная походка) • ДИСМЕТРИЯ (избыточность) • ДИЗАРТРИЯ • ДИЗЭКВИЛИБРАЦИЯ • АДИАДОХОКИНЕЗ
 БЛАГОДАРИМ ЗА ВНИМАНИЕ!
БЛАГОДАРИМ ЗА ВНИМАНИЕ!