

 Физиология ЦНС Часть 1
Физиология ЦНС Часть 1
 План лекции: 1. Нейрон, как структурная и функциональная единица ЦНС 2. Рефлекторная теория 3. Понятие о нервном центре и его свойства 4. Торможение в ЦНС 5. Принципы координации в ЦНС
План лекции: 1. Нейрон, как структурная и функциональная единица ЦНС 2. Рефлекторная теория 3. Понятие о нервном центре и его свойства 4. Торможение в ЦНС 5. Принципы координации в ЦНС
 считал мозг") ВЕХИ ИСТОРИИ VI в. до н. э. — Пифагор Самосский (Др. Греция) считал мозг средоточием психической деятельности. V— IV вв. до н. э. — Гиппократ (Др. Греция) описал четыре типа темперамента. V—I вв. до н. э. — Демокрит (Др. Греция) и Лукреций (Др. Рим) считали мозг седалищем души. IV в. до н. э. — Аристотель (Др. Греция) создал известный труд по психологии — трактат «О душе» . 1012— 1024 — Авиценна (Абу Али ибн Сина, Центральная Азия, Иран) в «Каноне врачебной науки» дал описание нервов черепномозговых и спинномозговых, чувствительных и двигательных. Считал мозг местом пребывания душевной силы и источником ее действия. 1637 — Р. Декарт (R. Descartes, Франция) дал первое описание двигательного рефлекса. 1784 — И. Прохаска (J. Prochaska, Чехия) развил представление о рефлексе и предложил этот термин. Описал рефлекторную дугу. 1811 и 1822 — Ч. Белл (Ch. Bell, Великобритания) и Ф. Мажанди (F. Magendie, Франция) открыли порядок распределения чувствительных и двигательных нервных волокон между дорсальными и вентральными корешками спинного мозга (закон Белла—Мажанди). 1832 — М. Голл и И. Мюллер (М. Goll, J. Muller, Германия) изложили учение о рефлексе. 1838 — Т. Шванн (Th. Schwann, Германия) описал строение оболочки нервного волокна (шванновская оболочка). 1856 — Р. Вирхов (R. Virchow, Германия) — открыл нейроглию. 1863 — И. М. Сеченов (Россия) в работе «Рефлексы головного мозга» описал явление центрального торможения и распространил принцип рефлекторной реакции на психическую деятельность и поведение человека. 1865 — О. Дейтерс (О. Deiters, Германия) описал отростки нейронов — аксон и дендриты. 1874 — В. А. Бец (Россия) описал гигантские пирамидные клетки в коре больших полушарий головного мозга, непосредственно управляющие мотонейронами скелетных мышц. 1874 — В. Вундт (W. Wundt, Германия) основал первую лабораторию экспериментальной психологии, ввел психометрию — количественный метод изучения психики. 1885 — П. Эрлих (Р. Ehrlich, Германия) впервые высказал мысль о существовании барьера между кровью и мозгом. 1891 — Г. Вальдейер (Н. Waldeyer, Германия) заявил о применимости клеточной теории к нервной системе и предложил термин «нейрон» .
ВЕХИ ИСТОРИИ VI в. до н. э. — Пифагор Самосский (Др. Греция) считал мозг средоточием психической деятельности. V— IV вв. до н. э. — Гиппократ (Др. Греция) описал четыре типа темперамента. V—I вв. до н. э. — Демокрит (Др. Греция) и Лукреций (Др. Рим) считали мозг седалищем души. IV в. до н. э. — Аристотель (Др. Греция) создал известный труд по психологии — трактат «О душе» . 1012— 1024 — Авиценна (Абу Али ибн Сина, Центральная Азия, Иран) в «Каноне врачебной науки» дал описание нервов черепномозговых и спинномозговых, чувствительных и двигательных. Считал мозг местом пребывания душевной силы и источником ее действия. 1637 — Р. Декарт (R. Descartes, Франция) дал первое описание двигательного рефлекса. 1784 — И. Прохаска (J. Prochaska, Чехия) развил представление о рефлексе и предложил этот термин. Описал рефлекторную дугу. 1811 и 1822 — Ч. Белл (Ch. Bell, Великобритания) и Ф. Мажанди (F. Magendie, Франция) открыли порядок распределения чувствительных и двигательных нервных волокон между дорсальными и вентральными корешками спинного мозга (закон Белла—Мажанди). 1832 — М. Голл и И. Мюллер (М. Goll, J. Muller, Германия) изложили учение о рефлексе. 1838 — Т. Шванн (Th. Schwann, Германия) описал строение оболочки нервного волокна (шванновская оболочка). 1856 — Р. Вирхов (R. Virchow, Германия) — открыл нейроглию. 1863 — И. М. Сеченов (Россия) в работе «Рефлексы головного мозга» описал явление центрального торможения и распространил принцип рефлекторной реакции на психическую деятельность и поведение человека. 1865 — О. Дейтерс (О. Deiters, Германия) описал отростки нейронов — аксон и дендриты. 1874 — В. А. Бец (Россия) описал гигантские пирамидные клетки в коре больших полушарий головного мозга, непосредственно управляющие мотонейронами скелетных мышц. 1874 — В. Вундт (W. Wundt, Германия) основал первую лабораторию экспериментальной психологии, ввел психометрию — количественный метод изучения психики. 1885 — П. Эрлих (Р. Ehrlich, Германия) впервые высказал мысль о существовании барьера между кровью и мозгом. 1891 — Г. Вальдейер (Н. Waldeyer, Германия) заявил о применимости клеточной теории к нервной системе и предложил термин «нейрон» .
 заложил основы психоанализа. 1897 — Ч. Шеррингтон") 1895 — 3. Фрейд (Z. Freud, Австро—Венгрия) заложил основы психоанализа. 1897 — Ч. Шеррингтон (Ch. Sherrington, Великобритания) ввел понятие «синапс» . 1902 — в докладе из лаборатории И. П. Павлова впервые заявлено об открытии условных рефлексов. 1905 — А. Бине и Т. Симон (A. Binet, Th. Simon, Франция) создали первый тест для оценки интеллекта (впоследствии IQ — «коэффициент интеллектуальности» ). 1906 — К. Гольджи (С. Golgi, Италия) и С. Рамон—и—Кахаль (S. Ramon у Cajal, Испания) — Нобелевская премия за доказательство клеточного строения нервной системы. 1909 — К. Бродман (К. Brodmann, Германия) описал в коре больших полушарий головного мозга 11 областей, включающих в себя 52 цитоархитектонических поля. 1921 — К. Юнг (К. Jung, Швейцария) в основу типологии характеров положил доминирующую психическую функцию индивида. Ввел понятия об интра— и экстраверсии. 1923 — А. А. Ухтомский (СССР) создал учение о доминанте. 1924 — Р. Магнус и А. Де Клейн (R. Magnus, A. De Kleijn, Нидерланды) ввели понятие о статических и статокинетических рефлексах ствола мозга. 1925 — В. Правдич—Неминский (СССР) зарегистрировал электрические процессы в мозгу животного. 1929 — Л. С. Штерн (СССР) ввела понятие о гистогематических барьерах. 1929 — Г. Бергер (Н. Berger, Германия) зарегистрировал электрические процессы в мозгу человека. 1920— 1930—е — Л. С. Выготский (СССР) исследовал развитие мышления и речи. 1932 — Ч. Шеррингтон и Э. Эдриан (Е. Adrian, Великобритания) — Нобелевская премия за открытие роли рецепторов нервов, передачи информации в виде электрических импульсов. 1936 — О. Лёви (О. Loewi, Австрия) и Г. Дейл (Н. Dale, Великобритания) — Нобелевская премия за открытие механизма синаптической передачи. 1937 — Дж. Папес (J. Papez, США) описал функции «круга Папеса» , включающего в себя гиппокамп, маммилярное тело, часть таламуса, кору поясной извилины и другие структуры мозга. 1938 — К. Хейманс (С. Heymans, Бельгия) — Нобелевская премия за открытие роли синусного и аортального механизмов в регуляции дыхания. 1930—е — У. Пенфилд (W. Penfield, Канада) обнаружил правильную пространственную проекцию скелетных мышц на двигательную область коры больших полушарий головного мозга. 1938 — Б. Скиннер (В. Skinner, США) предложил камеру для выработки инструментальных рефлексов у крыс (камера Скиннера).
1895 — 3. Фрейд (Z. Freud, Австро—Венгрия) заложил основы психоанализа. 1897 — Ч. Шеррингтон (Ch. Sherrington, Великобритания) ввел понятие «синапс» . 1902 — в докладе из лаборатории И. П. Павлова впервые заявлено об открытии условных рефлексов. 1905 — А. Бине и Т. Симон (A. Binet, Th. Simon, Франция) создали первый тест для оценки интеллекта (впоследствии IQ — «коэффициент интеллектуальности» ). 1906 — К. Гольджи (С. Golgi, Италия) и С. Рамон—и—Кахаль (S. Ramon у Cajal, Испания) — Нобелевская премия за доказательство клеточного строения нервной системы. 1909 — К. Бродман (К. Brodmann, Германия) описал в коре больших полушарий головного мозга 11 областей, включающих в себя 52 цитоархитектонических поля. 1921 — К. Юнг (К. Jung, Швейцария) в основу типологии характеров положил доминирующую психическую функцию индивида. Ввел понятия об интра— и экстраверсии. 1923 — А. А. Ухтомский (СССР) создал учение о доминанте. 1924 — Р. Магнус и А. Де Клейн (R. Magnus, A. De Kleijn, Нидерланды) ввели понятие о статических и статокинетических рефлексах ствола мозга. 1925 — В. Правдич—Неминский (СССР) зарегистрировал электрические процессы в мозгу животного. 1929 — Л. С. Штерн (СССР) ввела понятие о гистогематических барьерах. 1929 — Г. Бергер (Н. Berger, Германия) зарегистрировал электрические процессы в мозгу человека. 1920— 1930—е — Л. С. Выготский (СССР) исследовал развитие мышления и речи. 1932 — Ч. Шеррингтон и Э. Эдриан (Е. Adrian, Великобритания) — Нобелевская премия за открытие роли рецепторов нервов, передачи информации в виде электрических импульсов. 1936 — О. Лёви (О. Loewi, Австрия) и Г. Дейл (Н. Dale, Великобритания) — Нобелевская премия за открытие механизма синаптической передачи. 1937 — Дж. Папес (J. Papez, США) описал функции «круга Папеса» , включающего в себя гиппокамп, маммилярное тело, часть таламуса, кору поясной извилины и другие структуры мозга. 1938 — К. Хейманс (С. Heymans, Бельгия) — Нобелевская премия за открытие роли синусного и аортального механизмов в регуляции дыхания. 1930—е — У. Пенфилд (W. Penfield, Канада) обнаружил правильную пространственную проекцию скелетных мышц на двигательную область коры больших полушарий головного мозга. 1938 — Б. Скиннер (В. Skinner, США) предложил камеру для выработки инструментальных рефлексов у крыс (камера Скиннера).
 в трудах по физиологии движений создал схему рефлекторного") 1941 — Н. А. Бернштейн (СССР) в трудах по физиологии движений создал схему рефлекторного кольца. 1943 — Д. Ллойд (D. Lloyd, Великобритания) установил, что рефлекс на растяжение скелетной мышцы является моносинаптическим. 1944 — Г. Мэгун (Н. Magoun, США) установил, что электрическое раздражение ретикулярной формации ствола мозга может либо активировать, либо тормозить двигательные рефлексы. 1944 — Дж. Эрлангер и Г. Гассер (J. Erianger, Н. Gasser, США) — Нобелевская премия за открытие функциональных различий между нервными волокнами различных типов. 1949— В. Хесс (W. Hess, Швейцария)— Нобелевская премия за открытие гипоталамических центров регуляции вегетативных функций. 1950 — У. Пенфилд обнаружил соматотопическую локализацию моторных функций в коре больших полушарий головного мозга. 1952 — П. Маклин (Р. Mac. Lean, США) развил идеи Папеса и ввел понятие «лимбическая система» . 1957 — В. Маунткасл (V. Mountcastle, США) показал, что соматосенсорная кора больших полушарий головного мозга организована в элементарные функциональные единицы — колонки. 1957 — Ч. Осгуд (Ch. Osgood, США) разработал метод семантического дифференциала. 1963 — Дж. Эклс (J. Eccles, Австралия), А. Ходжкин (A. Hodgkin, Великобритания) и А. Хаксли (A. Huxley, Великобритания) — Нобелевская премия за открытие ионных механизмов передачи возбуждения по нервному волокну. 1965 — Э. Хеннеман (Е. Henneman, ФРГ) описал законы вовлечения в возбуждение мотонейронов двигательных ядер в зависимости от их размеров и возбудимости. 1960—е — А. Р. Лурия (СССР) применил психологические методы для анализа поражений мозга. 1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) — Нобелевская премия за открытие роли норадреналина в синаптической передаче. 1970 — Р. Гранит (R. Granit, Швеция) опубликовал классическую монографию о физиологических механизмах регуляции движений. 1973 — К. фон Фриш (К. v. Frisch, ФРГ), К. Лоренц (К. Lorenz, Австрия) и Н. Тинберген (N. Tinbergen, Великобритания) — Нобелевская премия за создание этологии (науки о поведении). 1975 — Дж. Хьюз и Г. Костерлиц (J. Huges, H. Kosterlitz, Великобритания) обнаружили в мозгу энкефалины — медиаторы опиоидной системы.
1941 — Н. А. Бернштейн (СССР) в трудах по физиологии движений создал схему рефлекторного кольца. 1943 — Д. Ллойд (D. Lloyd, Великобритания) установил, что рефлекс на растяжение скелетной мышцы является моносинаптическим. 1944 — Г. Мэгун (Н. Magoun, США) установил, что электрическое раздражение ретикулярной формации ствола мозга может либо активировать, либо тормозить двигательные рефлексы. 1944 — Дж. Эрлангер и Г. Гассер (J. Erianger, Н. Gasser, США) — Нобелевская премия за открытие функциональных различий между нервными волокнами различных типов. 1949— В. Хесс (W. Hess, Швейцария)— Нобелевская премия за открытие гипоталамических центров регуляции вегетативных функций. 1950 — У. Пенфилд обнаружил соматотопическую локализацию моторных функций в коре больших полушарий головного мозга. 1952 — П. Маклин (Р. Mac. Lean, США) развил идеи Папеса и ввел понятие «лимбическая система» . 1957 — В. Маунткасл (V. Mountcastle, США) показал, что соматосенсорная кора больших полушарий головного мозга организована в элементарные функциональные единицы — колонки. 1957 — Ч. Осгуд (Ch. Osgood, США) разработал метод семантического дифференциала. 1963 — Дж. Эклс (J. Eccles, Австралия), А. Ходжкин (A. Hodgkin, Великобритания) и А. Хаксли (A. Huxley, Великобритания) — Нобелевская премия за открытие ионных механизмов передачи возбуждения по нервному волокну. 1965 — Э. Хеннеман (Е. Henneman, ФРГ) описал законы вовлечения в возбуждение мотонейронов двигательных ядер в зависимости от их размеров и возбудимости. 1960—е — А. Р. Лурия (СССР) применил психологические методы для анализа поражений мозга. 1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) — Нобелевская премия за открытие роли норадреналина в синаптической передаче. 1970 — Р. Гранит (R. Granit, Швеция) опубликовал классическую монографию о физиологических механизмах регуляции движений. 1973 — К. фон Фриш (К. v. Frisch, ФРГ), К. Лоренц (К. Lorenz, Австрия) и Н. Тинберген (N. Tinbergen, Великобритания) — Нобелевская премия за создание этологии (науки о поведении). 1975 — Дж. Хьюз и Г. Костерлиц (J. Huges, H. Kosterlitz, Великобритания) обнаружили в мозгу энкефалины — медиаторы опиоидной системы.
 — Нобелевская") 1977 — Р. Гиймен и Э. Шалли (R. Guillemin, A. Schally, США) — Нобелевская премия за открытие тиролиберина — первого из рилизинг—факторов гипоталамуса. 1977 — Р. Ялоу ( R. Yalow, США) — Нобелевская премия за развитие методов радиоиммунологического исследования (RIA) пептидных гормонов. 1979 — А. Кормак (A. Cormack, США) и Г. Хаунсфилд (G. Hounsfield, Великобритания) — Нобелевская премия за разработку метода компьютерной томографии. 1981 — Р. Сперри (R. Sperry, США) — Нобелевская премия за открытие функциональной специализации полушарий мозга. 2000 — А. Карлссон (A. Carlsson, Швеция), П. Грингард и Э. Кэндел (Р. Greengard, Е. Kandel, СИТА) — Нобелевская премия за открытия в области передачи сигналов в нервной системе.
1977 — Р. Гиймен и Э. Шалли (R. Guillemin, A. Schally, США) — Нобелевская премия за открытие тиролиберина — первого из рилизинг—факторов гипоталамуса. 1977 — Р. Ялоу ( R. Yalow, США) — Нобелевская премия за развитие методов радиоиммунологического исследования (RIA) пептидных гормонов. 1979 — А. Кормак (A. Cormack, США) и Г. Хаунсфилд (G. Hounsfield, Великобритания) — Нобелевская премия за разработку метода компьютерной томографии. 1981 — Р. Сперри (R. Sperry, США) — Нобелевская премия за открытие функциональной специализации полушарий мозга. 2000 — А. Карлссон (A. Carlsson, Швеция), П. Грингард и Э. Кэндел (Р. Greengard, Е. Kandel, СИТА) — Нобелевская премия за открытия в области передачи сигналов в нервной системе.
 Глиальные клетки • • Нейроглия – составляет до 50% от объема мозга, их число в ЦНС достигает 140 млр (в 10 раз больше, чем нейронов), размеры в 3 -4 раза меньше нервных, способны к делению. Различают: Астроциты – многооторосчатые, крупные (7 -25 мкм), распологаются преимущественно в сером веществе мозга, отростки формируют «ножки» , окутывающие каппиляры. Олигодендроциты – клетки имеющие малое количество отростков, меньше астроцитов, участвуют в миелинизации аксонов. Микроглия - многоотросчатые мелкие клетки, относятся к блуждающим клеткам, способны к фагоцитозу, источником является мезодерма. ПП глиальных клеток – 70 – 90 м. В, электроневозбудимы, формируют локальные ответы, их электрические поля взаимодействуют с ВПСП и ПД нейронов.
Глиальные клетки • • Нейроглия – составляет до 50% от объема мозга, их число в ЦНС достигает 140 млр (в 10 раз больше, чем нейронов), размеры в 3 -4 раза меньше нервных, способны к делению. Различают: Астроциты – многооторосчатые, крупные (7 -25 мкм), распологаются преимущественно в сером веществе мозга, отростки формируют «ножки» , окутывающие каппиляры. Олигодендроциты – клетки имеющие малое количество отростков, меньше астроцитов, участвуют в миелинизации аксонов. Микроглия - многоотросчатые мелкие клетки, относятся к блуждающим клеткам, способны к фагоцитозу, источником является мезодерма. ПП глиальных клеток – 70 – 90 м. В, электроневозбудимы, формируют локальные ответы, их электрические поля взаимодействуют с ВПСП и ПД нейронов.
 Функции глии: • • • Электроизолирующая – обеспечивает возбудимость и проводимость нервных волокон, так как окутывают нейроциты в виде своеобразного футляра и продуцируют миелин. У новорожденных – восходящие пути покрыты миелином, а нисходящие не покрыты. Опорная. Защитная – выработка цитокинов, фагоцитоз, при повреждении нейронов образуют глиальный рубец. Обменная – синтез белков памяти, снабжение нейронов питательными веществами, поддержание оптимальной концентрации К+ в интерстиции. «Пульсация» - способность к ритмичному изменению объема (период «сокращения» - 1, 5 мин, «расслабления 2 – 4 мин), периодичность через каждые 2 -24 часа, способствует продвижению аксоплазмы и влияет на ток жидкости в межклеточном пространстве. НА – усиливает, а сератонин – угнетает пульсацию глиоцитов. Участвует в формировании гематоэнцефалического барьера.
Функции глии: • • • Электроизолирующая – обеспечивает возбудимость и проводимость нервных волокон, так как окутывают нейроциты в виде своеобразного футляра и продуцируют миелин. У новорожденных – восходящие пути покрыты миелином, а нисходящие не покрыты. Опорная. Защитная – выработка цитокинов, фагоцитоз, при повреждении нейронов образуют глиальный рубец. Обменная – синтез белков памяти, снабжение нейронов питательными веществами, поддержание оптимальной концентрации К+ в интерстиции. «Пульсация» - способность к ритмичному изменению объема (период «сокращения» - 1, 5 мин, «расслабления 2 – 4 мин), периодичность через каждые 2 -24 часа, способствует продвижению аксоплазмы и влияет на ток жидкости в межклеточном пространстве. НА – усиливает, а сератонин – угнетает пульсацию глиоцитов. Участвует в формировании гематоэнцефалического барьера.
 Нейрон и его компоненты
Нейрон и его компоненты
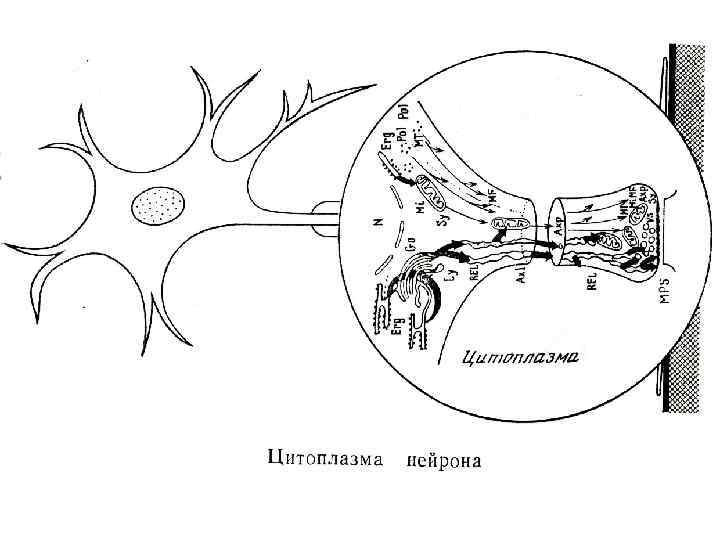
 Нейрон в электронном микроскопе
Нейрон в электронном микроскопе
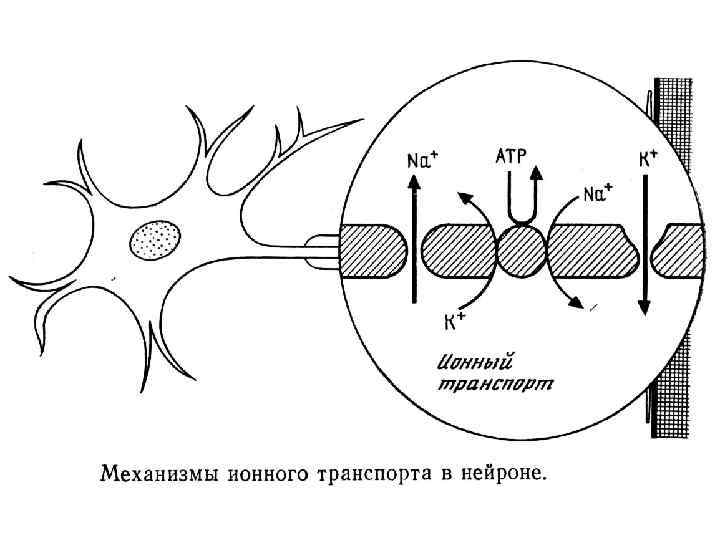
 Особенности метаболизма нейронов • Высокое потребление О 2. Полная гипоксия в течение 5 -6 минут ведет к гибели клеток коры. • Способность к альтернативным путям обмена . • Способность к созданию крупный запасов веществ. • Нервная клетка живет только вместе с глией. • Способность к регенерации отростков (0, 5 -4 мк/сут).
Особенности метаболизма нейронов • Высокое потребление О 2. Полная гипоксия в течение 5 -6 минут ведет к гибели клеток коры. • Способность к альтернативным путям обмена . • Способность к созданию крупный запасов веществ. • Нервная клетка живет только вместе с глией. • Способность к регенерации отростков (0, 5 -4 мк/сут).
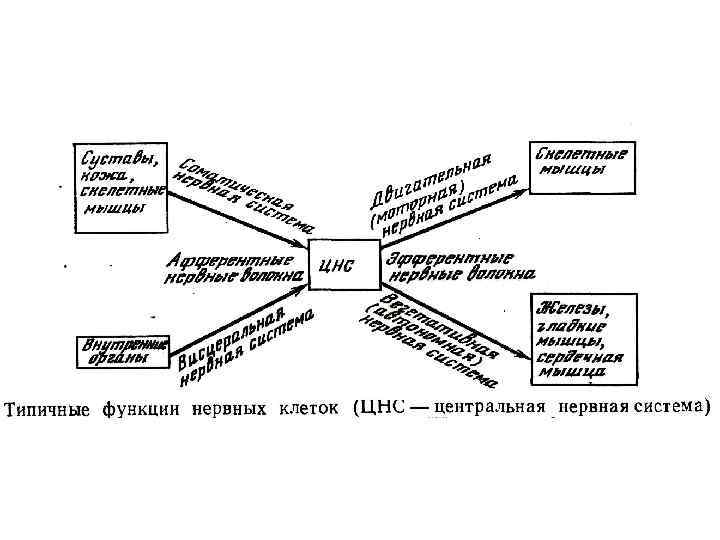
 КЛАССИФИКАЦИИ НЕЙРОНОВ Афферентные нейроны проводят возбуждение от рецепторов периферических органов в структуры ЦНС Эфферентные нейроны осуществляют передачу сигналов от ЦНС к органам-эффекторам (мышцам и железам) Ассоциативные нейроны проводят возбуждение между нейронами
КЛАССИФИКАЦИИ НЕЙРОНОВ Афферентные нейроны проводят возбуждение от рецепторов периферических органов в структуры ЦНС Эфферентные нейроны осуществляют передачу сигналов от ЦНС к органам-эффекторам (мышцам и железам) Ассоциативные нейроны проводят возбуждение между нейронами
 Классификация нейронов Эфферентный, эффекторный, моторный Афферентный, чувствительный Ассоциативный, вставочный рецептор мышца
Классификация нейронов Эфферентный, эффекторный, моторный Афферентный, чувствительный Ассоциативный, вставочный рецептор мышца
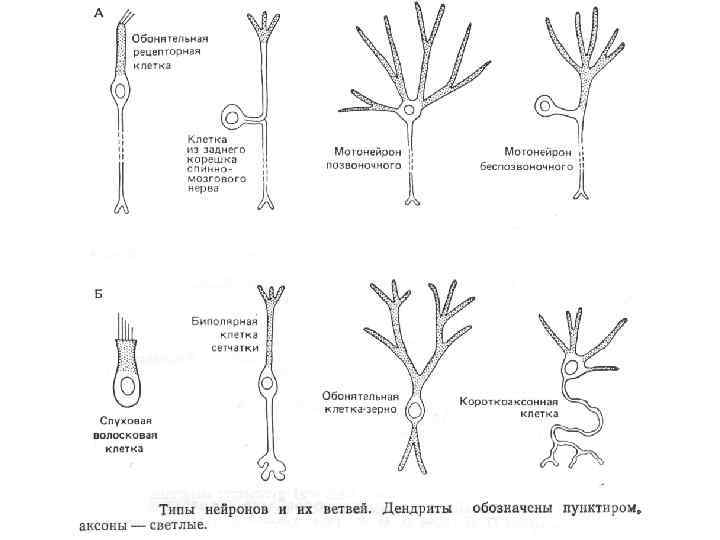
 Морфологическая классификация Псевдоуниполярные нейроны имеют один короткий отросток, который разделяется на некотором расстоянии от сомы на два длинных — дендрит и аксон. К псевдоуниполярным относятся нейроны сенсорных ганглиев спинного мозга. Биполярные нейроны имеют один дендрит и один аксон. Этот вид нейронов встречается в периферическом отделе зрительного, обонятельного и слухового анализаторов. Мультиполярные нейроны имеют один аксон и несколько дендритов, это наиболее распространённый вид нейронов. К ним относятся мотонейроны спинного мозга.
Морфологическая классификация Псевдоуниполярные нейроны имеют один короткий отросток, который разделяется на некотором расстоянии от сомы на два длинных — дендрит и аксон. К псевдоуниполярным относятся нейроны сенсорных ганглиев спинного мозга. Биполярные нейроны имеют один дендрит и один аксон. Этот вид нейронов встречается в периферическом отделе зрительного, обонятельного и слухового анализаторов. Мультиполярные нейроны имеют один аксон и несколько дендритов, это наиболее распространённый вид нейронов. К ним относятся мотонейроны спинного мозга.
 Основные типы нейронов в ЦНС млекопитающих
Основные типы нейронов в ЦНС млекопитающих
 Мотонейрон спинного мозга позвоночных Указаны функции разных его частей
Мотонейрон спинного мозга позвоночных Указаны функции разных его частей
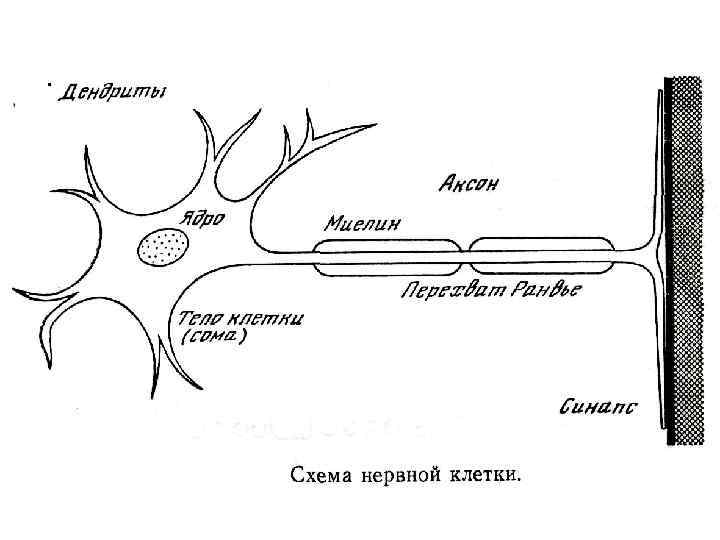
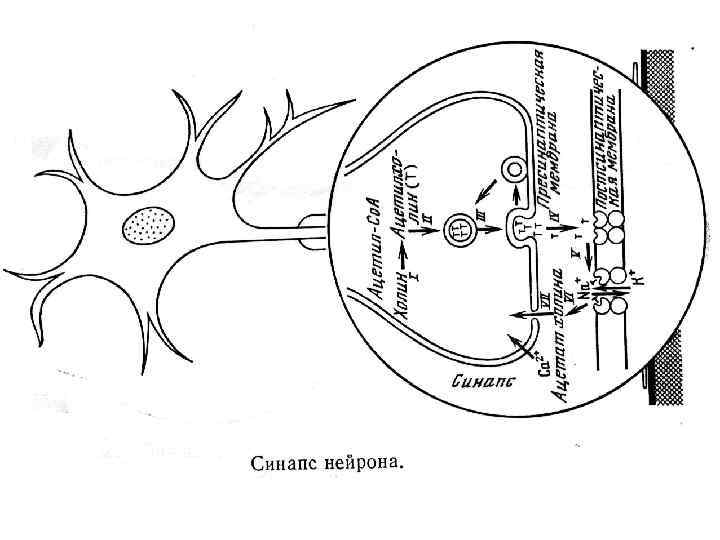
 содержит ядро. От перикариона отходят отростки. Один из них") Многоотростчатый нейрон Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них — аксон, все другие — дендриты. Справа и сверху вниз: функциональные зоны нервной клетки — рецептивная (дендритная), аксон (область проведения ПД), концевых разветвлений аксона (пресинаптическая).
Многоотростчатый нейрон Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них — аксон, все другие — дендриты. Справа и сверху вниз: функциональные зоны нервной клетки — рецептивная (дендритная), аксон (область проведения ПД), концевых разветвлений аксона (пресинаптическая).
 Универсальная классификация частей различных типов нейронов Дендриты — рецепторная зона — возбуждающий или тормозящий вход. Перикарион с дендритами — интегративная зона. Аксон — проводящая возбуждение (ПД) часть нейрона, его концевые разветвления образуют синапсы (выход) с соседними клетками. А–Г — афферентные (чувствительные) нейроны; Д — мотонейрон; Е — интернейроны
Универсальная классификация частей различных типов нейронов Дендриты — рецепторная зона — возбуждающий или тормозящий вход. Перикарион с дендритами — интегративная зона. Аксон — проводящая возбуждение (ПД) часть нейрона, его концевые разветвления образуют синапсы (выход) с соседними клетками. А–Г — афферентные (чувствительные) нейроны; Д — мотонейрон; Е — интернейроны
 Виды активности нейронов Фоновая активность Вызванная активность
Виды активности нейронов Фоновая активность Вызванная активность
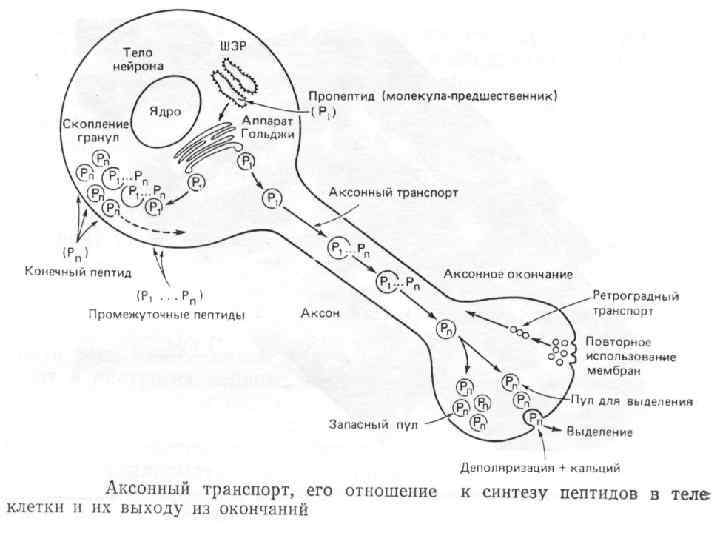
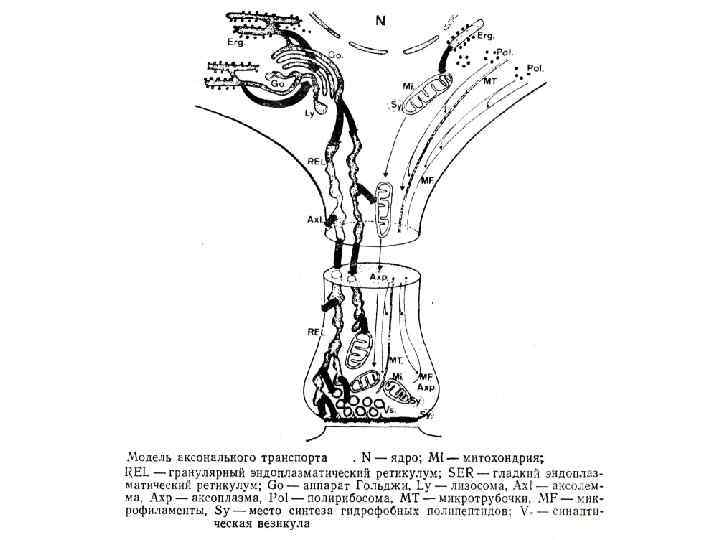
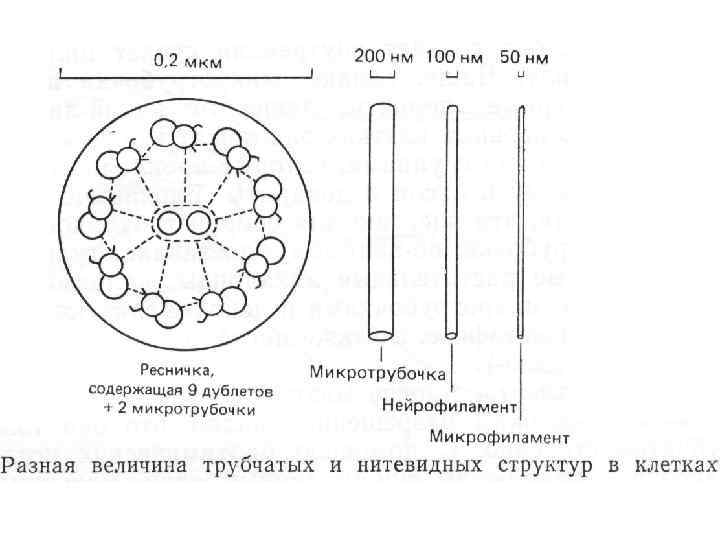
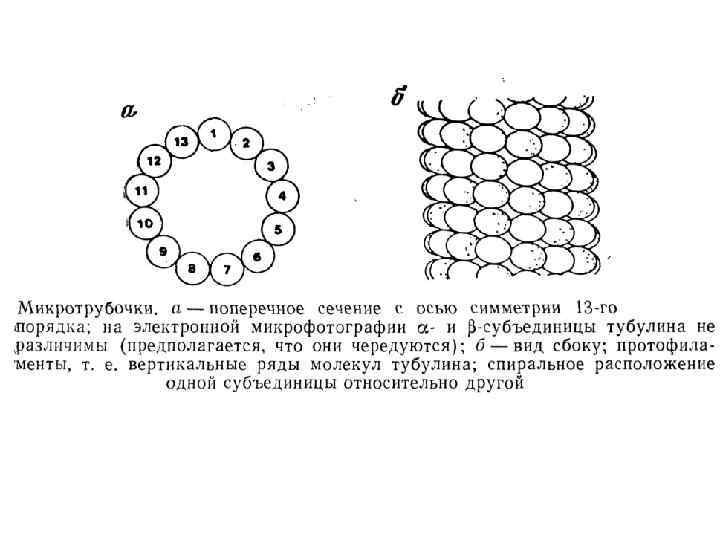
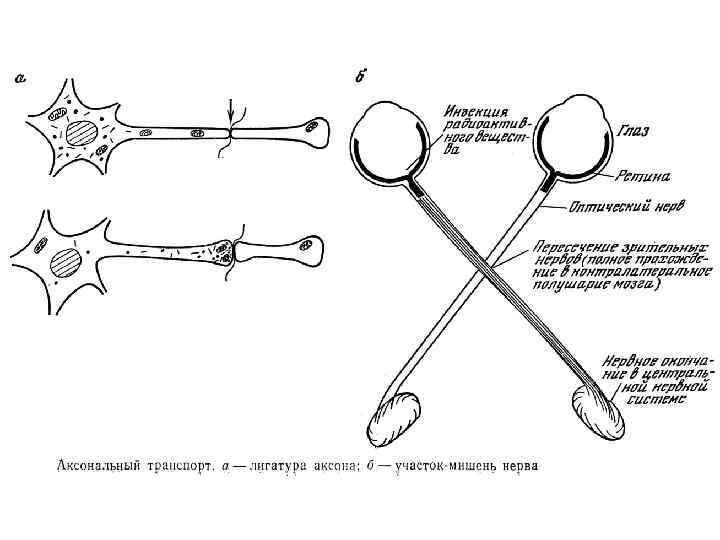
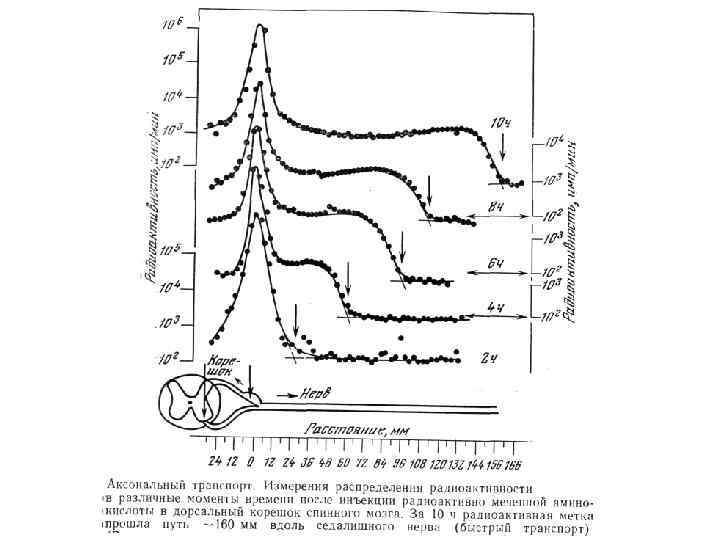
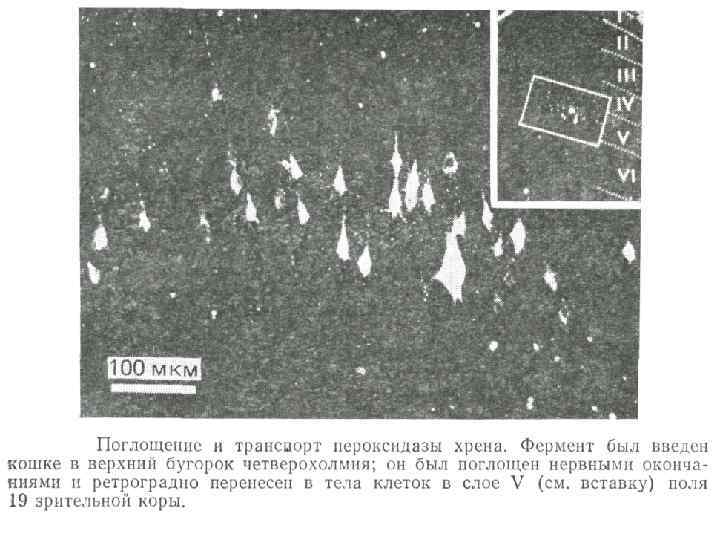

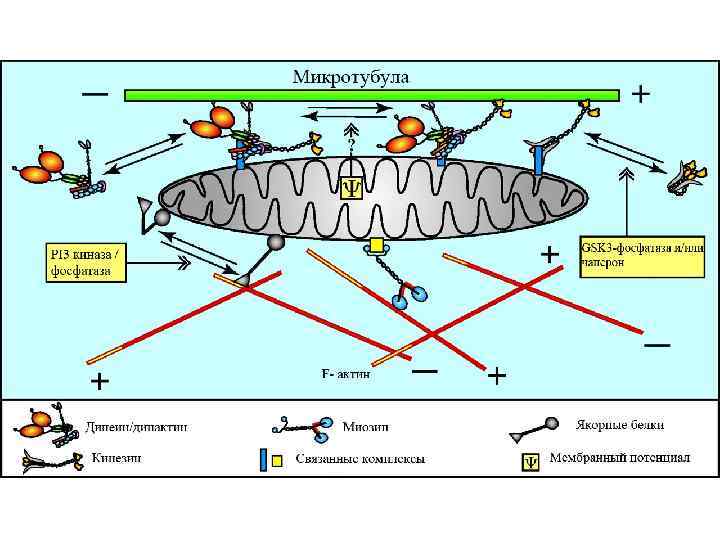
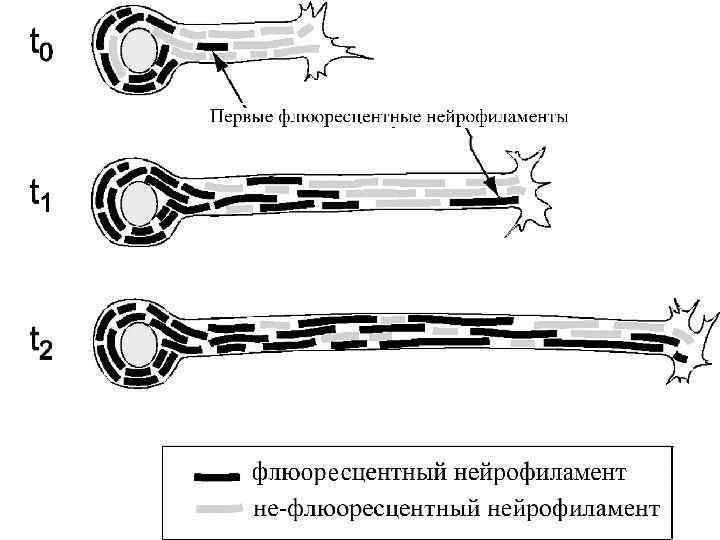
 Направление Мощность") Аксональный транспорт различных компонентов Структура груза Максимальная скорость Средняя скорость (световая микроскопия) Направление Мощность транспорта Везикулы-дериваты аппарата Гольджи 200 -400 мм/сут (2 -5 мкм/сек) 1 -5 мм/сек Антероградно е Высокоя Везикулы эндоцитов, лизосомы аутофагосомы 100 -250 мм/сут (1 -3 мм/сек) 1 -3 мм/сек Ретроградное Высокоя Митохондрия <70 мм/сут (<0. 8 мм/сек) 0. 3 -0. 7 мм/сек Двунаправлен ное Промежуточная Микрофиламенты, цитозольные белковые комплексы (медленный компонент b) 2 -8 мм/сут (0. 02 -0. 09 мм/сек) Неизвестна Неизвестно Микротубулы нейрофиламенты (медленный компонент а) 0. 2 -1 мм/сут (0. 0020. 01 мм/сек) 0. 3 -1 мм сек Двунаправлен ное Низкая
Аксональный транспорт различных компонентов Структура груза Максимальная скорость Средняя скорость (световая микроскопия) Направление Мощность транспорта Везикулы-дериваты аппарата Гольджи 200 -400 мм/сут (2 -5 мкм/сек) 1 -5 мм/сек Антероградно е Высокоя Везикулы эндоцитов, лизосомы аутофагосомы 100 -250 мм/сут (1 -3 мм/сек) 1 -3 мм/сек Ретроградное Высокоя Митохондрия <70 мм/сут (<0. 8 мм/сек) 0. 3 -0. 7 мм/сек Двунаправлен ное Промежуточная Микрофиламенты, цитозольные белковые комплексы (медленный компонент b) 2 -8 мм/сут (0. 02 -0. 09 мм/сек) Неизвестна Неизвестно Микротубулы нейрофиламенты (медленный компонент а) 0. 2 -1 мм/сут (0. 0020. 01 мм/сек) 0. 3 -1 мм сек Двунаправлен ное Низкая
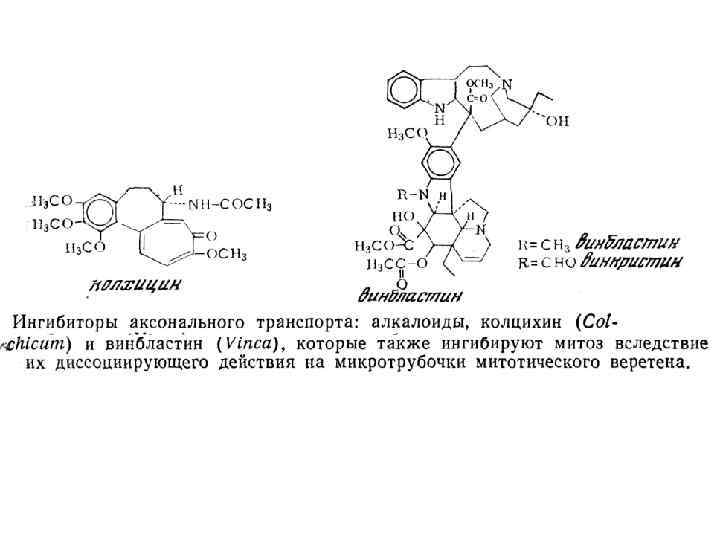
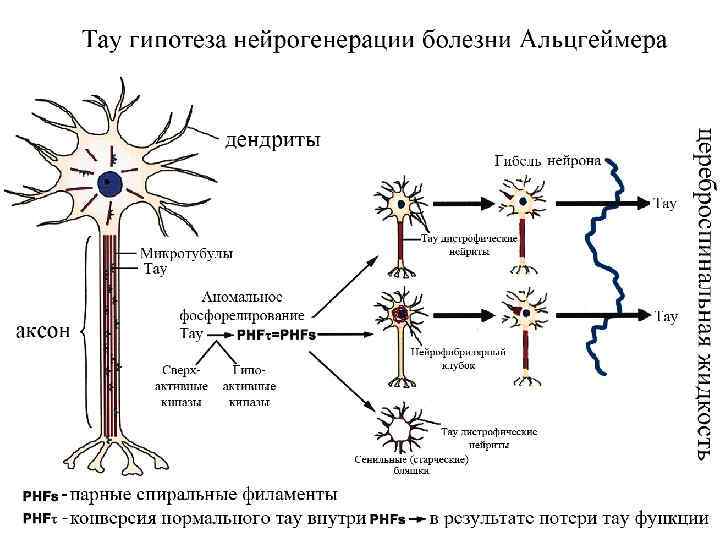
 Нейродегенеративные заболевания, связанные с ослаблением аксонального транспорта Заболевания Мутантный ген / нарушения синтеза белка Наследственная спастическая параплегия Тяжелые цепи кинезина Семейная моторнонейрональная болезнь Субъединица p 150 динактина Болезнь Шарко-Мари-Тута типа 2 А (мышечная атрофия перонеального типа) b-субъединица кинезина Kif 1 B Болезнь Хантингтона Хантингтин Семейные амиотрофические латеральные склерозы Супероксид дисмутаза I Болезнь Паркинсона и связанные синуклеинопатии a-синуклин Болезнь Альцгеймера АРР, призенилины, Тау
Нейродегенеративные заболевания, связанные с ослаблением аксонального транспорта Заболевания Мутантный ген / нарушения синтеза белка Наследственная спастическая параплегия Тяжелые цепи кинезина Семейная моторнонейрональная болезнь Субъединица p 150 динактина Болезнь Шарко-Мари-Тута типа 2 А (мышечная атрофия перонеального типа) b-субъединица кинезина Kif 1 B Болезнь Хантингтона Хантингтин Семейные амиотрофические латеральные склерозы Супероксид дисмутаза I Болезнь Паркинсона и связанные синуклеинопатии a-синуклин Болезнь Альцгеймера АРР, призенилины, Тау
 Синапсы в ЦНС химический электрический
Синапсы в ЦНС химический электрический
 Синапсы на нейроне
Синапсы на нейроне
 СТРУКТУРА И МЕХАНИЗМ ДЕЙСТВИЯ СИНАПСА
СТРУКТУРА И МЕХАНИЗМ ДЕЙСТВИЯ СИНАПСА
 Синаптические процессы в возбужденном и невозбужденном синапсе
Синаптические процессы в возбужденном и невозбужденном синапсе
 Генерация ПД при возникновении ВПСП
Генерация ПД при возникновении ВПСП
 мв ДЕПОЛЯРИЗАЦИЯ Na+ - 70 - 85 - 90") Возбуждающий постсинаптический потенциал ( ВПСП) мв ДЕПОЛЯРИЗАЦИЯ Na+ - 70 - 85 - 90 мс 0 4 8 12
Возбуждающий постсинаптический потенциал ( ВПСП) мв ДЕПОЛЯРИЗАЦИЯ Na+ - 70 - 85 - 90 мс 0 4 8 12
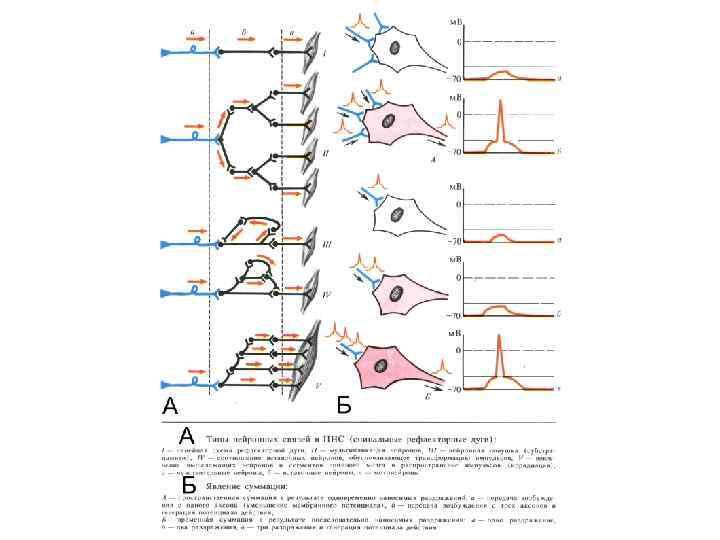
 Виды связей между нейронами
Виды связей между нейронами
 ФУНКЦИОНАЛЬНЫЕ МЕЖНЕЙРОННЫЕ ОБРАЗОВАНИЯ В ЦНС • НЕРВНЫЕ ЦЕПИ • ЛОКАЛЬНЫЕ СЕТИ • НЕЙРОННЫЕ АНСАМБЛИ • НЕРВНЫЕ ЦЕНТРЫ
ФУНКЦИОНАЛЬНЫЕ МЕЖНЕЙРОННЫЕ ОБРАЗОВАНИЯ В ЦНС • НЕРВНЫЕ ЦЕПИ • ЛОКАЛЬНЫЕ СЕТИ • НЕЙРОННЫЕ АНСАМБЛИ • НЕРВНЫЕ ЦЕНТРЫ
 В ЦНС • Односторонний характер проведения в нейронной") РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ ( ВОЗБУЖДЕНИЯ ) В ЦНС • Односторонний характер проведения в нейронной цепи • Дивергенция, мультипликация, иррадиация • Конвергенция и суммация • Реверберация возбуждения в нейронных цепях Лоренто-де-Но
РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ ( ВОЗБУЖДЕНИЯ ) В ЦНС • Односторонний характер проведения в нейронной цепи • Дивергенция, мультипликация, иррадиация • Конвергенция и суммация • Реверберация возбуждения в нейронных цепях Лоренто-де-Но
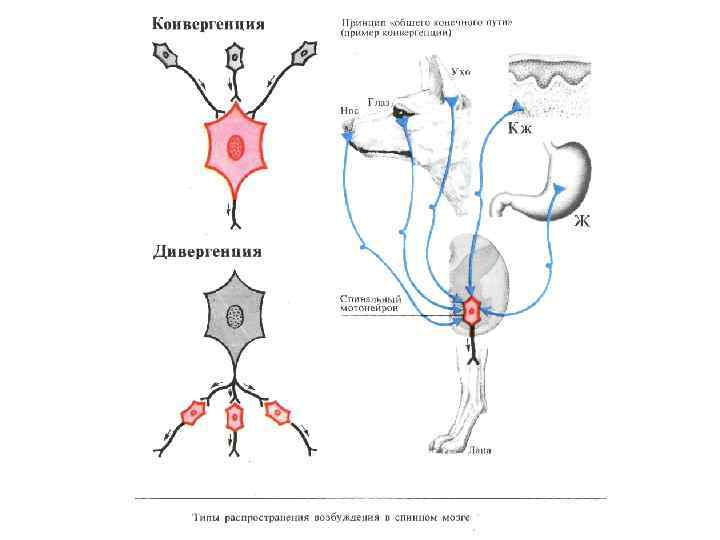
 РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
 РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС КОНВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ
РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС КОНВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ
 РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС РЕВЕРБЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СЕТИ по Лоренто-де-Но
РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС РЕВЕРБЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СЕТИ по Лоренто-де-Но
 КОДИРОВАНИЕ ИНФОРМАЦИИ В НЕЙРОННОЙ ЦЕПИ • ИМПУЛЬСНОЕ: НЕПРЕРЫВНОЕ, ПАЧЕЧНОЕ, ЧАСТОТНОЕ, ИНТЕРВАЛЬНОЕ, ДЛИТЕЛЬНОСТЬЮ АКТИВНОСТИ, ВАРИАБЕЛЬНОСТЬЮ АКТИВНОСТИ • НЕИМПУЛЬСНОЕ: СОСТАВ И СКОРОСТЬ ТОКА АКСОПЛАЗМЫ
КОДИРОВАНИЕ ИНФОРМАЦИИ В НЕЙРОННОЙ ЦЕПИ • ИМПУЛЬСНОЕ: НЕПРЕРЫВНОЕ, ПАЧЕЧНОЕ, ЧАСТОТНОЕ, ИНТЕРВАЛЬНОЕ, ДЛИТЕЛЬНОСТЬЮ АКТИВНОСТИ, ВАРИАБЕЛЬНОСТЬЮ АКТИВНОСТИ • НЕИМПУЛЬСНОЕ: СОСТАВ И СКОРОСТЬ ТОКА АКСОПЛАЗМЫ
 НЕРВНЫЙ ЦЕНТР 1. Нервный центр - совокупность нейронов, обеспечивающих реализацию определенного рефлекса 2. Нервный центр - функционально связанная совокупность нейронных ансамблей разных этажей нервной системы, обеспечивающих регуляцию определенных функций организма
НЕРВНЫЙ ЦЕНТР 1. Нервный центр - совокупность нейронов, обеспечивающих реализацию определенного рефлекса 2. Нервный центр - функционально связанная совокупность нейронных ансамблей разных этажей нервной системы, обеспечивающих регуляцию определенных функций организма
 Свойства нервных центров 1. 2. 3. 4. Односторонность проведения возбуждения Иррадиация возбуждения Суммация возбуждения Наличие синаптической задержки (центральная задержка рефлекса) 5. Высокая утомляемость. 6. Тонус 7. Пластичность 8. Конвергенция 9. Интеграция в нервных центрах 10. Свойство доминанты 11. Цефализация нервной системы 12. Трансформация 13. Явление последействия 14. Явление центрального облегчения и окклюзия
Свойства нервных центров 1. 2. 3. 4. Односторонность проведения возбуждения Иррадиация возбуждения Суммация возбуждения Наличие синаптической задержки (центральная задержка рефлекса) 5. Высокая утомляемость. 6. Тонус 7. Пластичность 8. Конвергенция 9. Интеграция в нервных центрах 10. Свойство доминанты 11. Цефализация нервной системы 12. Трансформация 13. Явление последействия 14. Явление центрального облегчения и окклюзия
 и мышечного (коленного) (Б) рефлексов млекопитающих А — трехнейронная, дисинаптическая") Рефлекторные дуги кожного (А) и мышечного (коленного) (Б) рефлексов млекопитающих А — трехнейронная, дисинаптическая дуга кожного рефлекса; Б — двухнейронная, моносинаптическая дуга коленного рефлекса: 1 — кожные рецепторы, 2 — мышечные веретена (рецепторы растяжения мышцы, реагирующие на удар по подколенному сухожилию)
Рефлекторные дуги кожного (А) и мышечного (коленного) (Б) рефлексов млекопитающих А — трехнейронная, дисинаптическая дуга кожного рефлекса; Б — двухнейронная, моносинаптическая дуга коленного рефлекса: 1 — кожные рецепторы, 2 — мышечные веретена (рецепторы растяжения мышцы, реагирующие на удар по подколенному сухожилию)
, конвергенция (Б), окклюзия (В), временная (Г) и пространственная (Д) суммация в локальных") Дивергенция (А), конвергенция (Б), окклюзия (В), временная (Г) и пространственная (Д) суммация в локальных нейронных цепях
Дивергенция (А), конвергенция (Б), окклюзия (В), временная (Г) и пространственная (Д) суммация в локальных нейронных цепях
") Центральная окклюзия А 1 При раздражении А возбуждаются 4 нейрона (1, 2, 3, 4) 2 3 При раздражении В возбуждаются 4 нейрона (3, 4, 5, 6) В 4 5 6 Клетки центральной части нейронного пула При раздражении А + В возбуждаются 6 нейронов (1, 2, 3, 4, 5, 6)
Центральная окклюзия А 1 При раздражении А возбуждаются 4 нейрона (1, 2, 3, 4) 2 3 При раздражении В возбуждаются 4 нейрона (3, 4, 5, 6) В 4 5 6 Клетки центральной части нейронного пула При раздражении А + В возбуждаются 6 нейронов (1, 2, 3, 4, 5, 6)
 Трансформация ритма 50 А ? 50 В Фазовые соотношения входящих импульсов В А 100 В А В 1 (следующие попадают в рефрактерность предыдущего
Трансформация ритма 50 А ? 50 В Фазовые соотношения входящих импульсов В А 100 В А В 1 (следующие попадают в рефрактерность предыдущего
 Трансформация ритма Триггерные свойства аксонного холмика Порог 30 мв Порог 10 мв Тело нейрона Ек Ео Аксонный холмик Ек Ео «На ружейный выстрел нейрон отвечает пулеметной очередью»
Трансформация ритма Триггерные свойства аксонного холмика Порог 30 мв Порог 10 мв Тело нейрона Ек Ео Аксонный холмик Ек Ео «На ружейный выстрел нейрон отвечает пулеметной очередью»
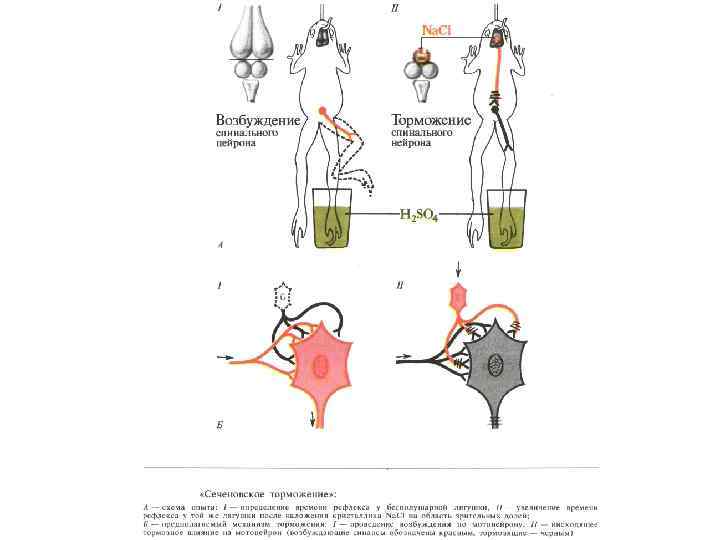
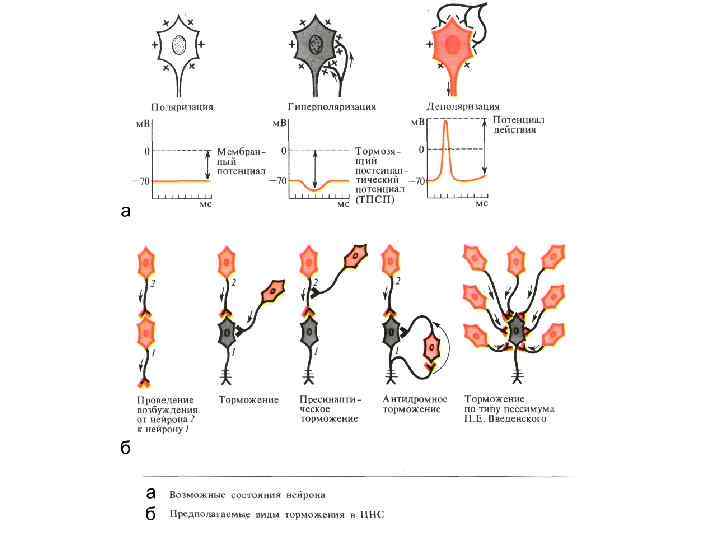
 ТОРМОЖЕНИЯ • П Е Р В И Ч Н О Е: А)") ВИДЫ (СПОСОБЫ) ТОРМОЖЕНИЯ • П Е Р В И Ч Н О Е: А) ПОСТСИНАПТИЧЕСКОЕ Б) ПРЕСИНАПТИЧЕСКОЕ • В Т О Р И Ч Н О Е: А) ПЕССИМАЛЬНОЕ по Н. Введенскому Б) СЛЕДОВОЕ (при следовой гиперполяризации)
ВИДЫ (СПОСОБЫ) ТОРМОЖЕНИЯ • П Е Р В И Ч Н О Е: А) ПОСТСИНАПТИЧЕСКОЕ Б) ПРЕСИНАПТИЧЕСКОЕ • В Т О Р И Ч Н О Е: А) ПЕССИМАЛЬНОЕ по Н. Введенскому Б) СЛЕДОВОЕ (при следовой гиперполяризации)
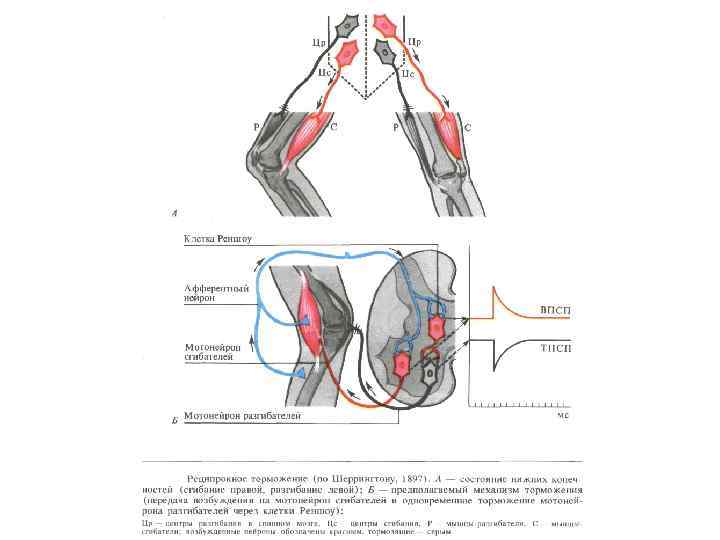
") Типы торможения • ВОЗВРАТНОЕ • ЛАТЕРАЛЬНОЕ • СОПРЯЖЕННОЕ (РЕЦИПРОКНОЕ)
Типы торможения • ВОЗВРАТНОЕ • ЛАТЕРАЛЬНОЕ • СОПРЯЖЕННОЕ (РЕЦИПРОКНОЕ)
 ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ 1 - аксон тормозного нейрона; 2 - аксон возбуждающего нейрона; 3 постсинаптическая мембрана альфа-мотонейрона Катодическая депрессия в аксо-аксональном синапсе при длительной деполяризации его постсинаптической мембраны
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ 1 - аксон тормозного нейрона; 2 - аксон возбуждающего нейрона; 3 постсинаптическая мембрана альфа-мотонейрона Катодическая депрессия в аксо-аксональном синапсе при длительной деполяризации его постсинаптической мембраны
 Посттетаническое торможение • Суммация следовой гиперполяризации после серии возбуждений
Посттетаническое торможение • Суммация следовой гиперполяризации после серии возбуждений
 Посттетаническое торможение • Суммация следовой гиперполяризации после серии возбуждений
Посттетаническое торможение • Суммация следовой гиперполяризации после серии возбуждений
 Пессимальное торможение • Развитие катодической депрессии в результате длительной деполяризации при длительном частом раздражении
Пессимальное торможение • Развитие катодической депрессии в результате длительной деполяризации при длительном частом раздражении
 Возвратное торможение по Реншоу • б) Латеральное торможение •") ТОРМОЖЕНИЕ В ЦНС • а) Возвратное торможение по Реншоу • б) Латеральное торможение • г) Сопряженное торможение • В - возбуждение • Т - торможение
ТОРМОЖЕНИЕ В ЦНС • а) Возвратное торможение по Реншоу • б) Латеральное торможение • г) Сопряженное торможение • В - возбуждение • Т - торможение
 Примеры нарушения торможения в ЦНС • НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: • СТРИХНИН - БЛОКАДА РЕЦЕПТОРОВ ТОРМОЗНЫХ СИНАПСОВ • СТОЛБНЯЧНЫЙ ТОКСИН - НАРУШЕНИЕ ОСВОБОЖДЕНИЯ ТОРМОЗНОГО МЕДИАТОРА • НАРУШЕНИЕ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: • ПИКРОТОКСИН - БЛОКАДА ПРЕСИНАПТИЧЕСКИХ СИНАПСОВ
Примеры нарушения торможения в ЦНС • НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: • СТРИХНИН - БЛОКАДА РЕЦЕПТОРОВ ТОРМОЗНЫХ СИНАПСОВ • СТОЛБНЯЧНЫЙ ТОКСИН - НАРУШЕНИЕ ОСВОБОЖДЕНИЯ ТОРМОЗНОГО МЕДИАТОРА • НАРУШЕНИЕ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: • ПИКРОТОКСИН - БЛОКАДА ПРЕСИНАПТИЧЕСКИХ СИНАПСОВ
 ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ • 1. РЕЦИПРОКНОСТИ • 2. ДОМИНАНТЫ • 3. ОБЩЕГО КОНЕЧНОГО ПУТИ (по Шеррингтону) • 4. СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ • 5. ОБРАТНОЙ АФФЕРЕНТАЦИИ
ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ • 1. РЕЦИПРОКНОСТИ • 2. ДОМИНАНТЫ • 3. ОБЩЕГО КОНЕЧНОГО ПУТИ (по Шеррингтону) • 4. СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ • 5. ОБРАТНОЙ АФФЕРЕНТАЦИИ
 ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ ОБРАТНАЯ АФФЕРЕНТАЦИЯ ДВИГАТЕЛЬНАЯ КОМАНДА
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ ОБРАТНАЯ АФФЕРЕНТАЦИЯ ДВИГАТЕЛЬНАЯ КОМАНДА
 + + + - -") ПРИНЦИП РЕЦИПРОКНОСТИ ( СОПРЯЖЕННОГО ТОРМОЖЕНИЯ ) + + + - -
ПРИНЦИП РЕЦИПРОКНОСТИ ( СОПРЯЖЕННОГО ТОРМОЖЕНИЯ ) + + + - -
 Торможение в ЦНС Виды торможения Нейроны Рецепторы Ионный механизм Блокаторы Пресинаптические ГАМКергические ГАМК 1 Cl- Бикукулин, столбнячный токсин Постсинаптические ГАМКергические, Глицинергические ГАМК 1 ГАМК 2 глицинорецептор ы Cl. K+ Cl- Стрихнин, столбнячный токсин
Торможение в ЦНС Виды торможения Нейроны Рецепторы Ионный механизм Блокаторы Пресинаптические ГАМКергические ГАМК 1 Cl- Бикукулин, столбнячный токсин Постсинаптические ГАМКергические, Глицинергические ГАМК 1 ГАМК 2 глицинорецептор ы Cl. K+ Cl- Стрихнин, столбнячный токсин
 Принцип общего конечного пути в спинном мозге
Принцип общего конечного пути в спинном мозге
 ПРИНЦИП ДОМИНАНТЫ Раздражители Нервные центры Рефлексы
ПРИНЦИП ДОМИНАНТЫ Раздражители Нервные центры Рефлексы
 ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ КОРА ПОДКОРКА СТВОЛ СПИННОЙ МОЗГ
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ КОРА ПОДКОРКА СТВОЛ СПИННОЙ МОЗГ
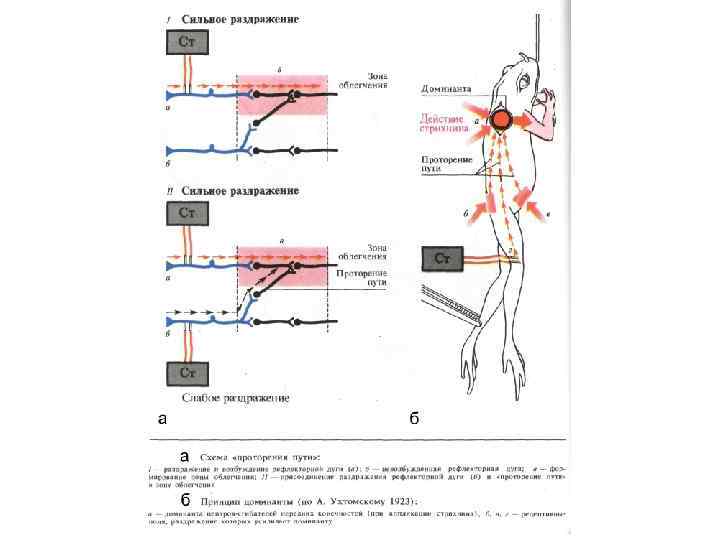
 • Доминанта - временно господствующий рефлекс") ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А. А. Ухтомскому, 1931) • Доминанта - временно господствующий рефлекс или поведенческий акт, которым трансформируется и направляется для данного времени прочих равных условиях работа прочих рефлекторных дуг, рефлекторного аппарата и поведения в целом
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А. А. Ухтомскому, 1931) • Доминанта - временно господствующий рефлекс или поведенческий акт, которым трансформируется и направляется для данного времени прочих равных условиях работа прочих рефлекторных дуг, рефлекторного аппарата и поведения в целом
 • 1. Повышенная возбудимость доминантного центра") Основные признаки доминанты ( по А. А. Ухтомскому) • 1. Повышенная возбудимость доминантного центра • 2. Стойкость возбуждения в доминантном центре • 3. Способность подкреплять свое возбуждение посторонними импульсами • 4. Способность тормозить другие текущие рефлексы на общем конечном пути
Основные признаки доминанты ( по А. А. Ухтомскому) • 1. Повышенная возбудимость доминантного центра • 2. Стойкость возбуждения в доминантном центре • 3. Способность подкреплять свое возбуждение посторонними импульсами • 4. Способность тормозить другие текущие рефлексы на общем конечном пути
 • «След однажды пережитой доминанты, а подчас и вся пережитая доминанта, могут быть вызваны вновь в поле внимания, как только возобновится, хотя бы частично раздражитель, ставший для нее адекватным. Старый и дряхлый боевой конь весь преображается и попрежнему мчится в строй при звуке сигнальной трубы» А. А. Ухтомский, 1923
• «След однажды пережитой доминанты, а подчас и вся пережитая доминанта, могут быть вызваны вновь в поле внимания, как только возобновится, хотя бы частично раздражитель, ставший для нее адекватным. Старый и дряхлый боевой конь весь преображается и попрежнему мчится в строй при звуке сигнальной трубы» А. А. Ухтомский, 1923
 ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
 РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
 ЗВЕНЬЯ РЕФЛЕКТОРНОЙ РЕГУЛЯЦИИ
ЗВЕНЬЯ РЕФЛЕКТОРНОЙ РЕГУЛЯЦИИ
 Простая и сложные рефлекторные дуги
Простая и сложные рефлекторные дуги

 Принципы классификации рефлексов 1. 2. 3. 4. 5. 6. 7. 8. 9. По происхождению – безусловные и условные. По биологическому значению. По расположению рецепторов. По виду рецепторов. По месту расположения центра. По длительности ответной реакции. По характеру ответной реакции. По принадлежности к системе органов. По характеру внешнего проявления реакции.
Принципы классификации рефлексов 1. 2. 3. 4. 5. 6. 7. 8. 9. По происхождению – безусловные и условные. По биологическому значению. По расположению рецепторов. По виду рецепторов. По месту расположения центра. По длительности ответной реакции. По характеру ответной реакции. По принадлежности к системе органов. По характеру внешнего проявления реакции.
 Благодарим за внимание
Благодарим за внимание