

Физиология анализаторов_ФФ_2015.ppt
- Количество слайдов: 113
 Физиология анализаторов Сенсорные системы
Физиология анализаторов Сенсорные системы
 Анализатор - по И. П. Павлову совокупность нервных образований у высших позвоночных животных, обеспечивающая разложение и анализ в нервной системе раздражителей, воздействующих на организм.
Анализатор - по И. П. Павлову совокупность нервных образований у высших позвоночных животных, обеспечивающая разложение и анализ в нервной системе раздражителей, воздействующих на организм.
 1. 2. 3. 4. 5. 6. Зрительный. Слуховой. Обонятельная. Вкусовая. Интероцептивная.") Анализаторы (сенсорные системы) 1. 2. 3. 4. 5. 6. Зрительный. Слуховой. Обонятельная. Вкусовая. Интероцептивная. Болевая (ноцицептивная).
Анализаторы (сенсорные системы) 1. 2. 3. 4. 5. 6. Зрительный. Слуховой. Обонятельная. Вкусовая. Интероцептивная. Болевая (ноцицептивная).
 - осуществляет восприятие энергии раздражителя и трансформацию ее в специфический") ОТДЕЛЫ АНАЛИЗАТОРА Периферический (рецепторный) - осуществляет восприятие энергии раздражителя и трансформацию ее в специфический процесс возбуждения. Проводниковый - осуществляет передачу возникшего возбуждения в кору головного мозга (представленный афферентными нервами и подкорковыми центрами). Центральный (корковый) - осуществляет высший анализ и синтез возбуждений и формирование соответствующего ощущения (представлен соответствующими зонами коры головного мозга).
ОТДЕЛЫ АНАЛИЗАТОРА Периферический (рецепторный) - осуществляет восприятие энергии раздражителя и трансформацию ее в специфический процесс возбуждения. Проводниковый - осуществляет передачу возникшего возбуждения в кору головного мозга (представленный афферентными нервами и подкорковыми центрами). Центральный (корковый) - осуществляет высший анализ и синтез возбуждений и формирование соответствующего ощущения (представлен соответствующими зонами коры головного мозга).
 Структура анализаторной системы
Структура анализаторной системы
 Этапы деятельности анализатора
Этапы деятельности анализатора
 КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ : ПО • • • Психофизиологический характер ощущения: тепловые, холодовые, болевые и др. Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осморецепторы и др. Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы. Отношение к одной или нескольким модальностям: моно- и полимодальные. Способность воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним: контактные и дистантные. Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы). Скорость адаптации: быстроадаптирующиеся, (тактильные), медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы). Отношение к различным моментам действия раздражителя. Морфофункциональная организация и механизм возникновения возбуждения: первичночувствующие и вторичночувствующие.
КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ : ПО • • • Психофизиологический характер ощущения: тепловые, холодовые, болевые и др. Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осморецепторы и др. Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы. Отношение к одной или нескольким модальностям: моно- и полимодальные. Способность воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним: контактные и дистантные. Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы). Скорость адаптации: быстроадаптирующиеся, (тактильные), медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы). Отношение к различным моментам действия раздражителя. Морфофункциональная организация и механизм возникновения возбуждения: первичночувствующие и вторичночувствующие.
 • В первичночувствующих рецепторах стимул действует на воспринимающий субстрат, заложенный в самом сенсорном нейроне, который при этом возбуждается непосредственно (первично) раздражителем. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена. • К вторичночувствующим относятся те рецепторы, у которых между действующим стимулом и сенсорным нейроном располагаются дополнительные рецептирующие клетки, при этом сенсорный нейрон возбуждается не непосредственно стимулом, а опосредовано (вторично) — потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
• В первичночувствующих рецепторах стимул действует на воспринимающий субстрат, заложенный в самом сенсорном нейроне, который при этом возбуждается непосредственно (первично) раздражителем. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена. • К вторичночувствующим относятся те рецепторы, у которых между действующим стимулом и сенсорным нейроном располагаются дополнительные рецептирующие клетки, при этом сенсорный нейрон возбуждается не непосредственно стимулом, а опосредовано (вторично) — потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
 МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ВОЗБУЖДЕНИЯ В ПЕРВИЧНОЧУВСТВУЮЩИХ РЕЦЕПТОРАХ I этап — специфическое взаимодействие раздражителя с мембраной рецептора; II этап — возникновение рецепторного потенциала в месте взаимодействия раздражителя с рецептором; III этап — электротоническое распространение рецепторного потенциала к аксону сенсорного нейрона); IV этап — генерация потенциала действия; V этап — проведение потенциала действия по нервному волокну в ортодромном направлении.
МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ВОЗБУЖДЕНИЯ В ПЕРВИЧНОЧУВСТВУЮЩИХ РЕЦЕПТОРАХ I этап — специфическое взаимодействие раздражителя с мембраной рецептора; II этап — возникновение рецепторного потенциала в месте взаимодействия раздражителя с рецептором; III этап — электротоническое распространение рецепторного потенциала к аксону сенсорного нейрона); IV этап — генерация потенциала действия; V этап — проведение потенциала действия по нервному волокну в ортодромном направлении.
 МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ВОЗБУЖДЕНИЯ ВО ВТОРИЧНОЧУВСТВУЮЩИХ РЕЦЕПТОРАХ I-III этапы - совпадают с такими же этапами первичночувствующих рецепторов, но протекают они в специализированной рецептирующей клетке и заканчиваются на ее пресинаптической мембране; IV этап — выделение медиатора пресинаптическими структурами рецептирующей клетки; V этап — возникновение генераторного потенциала на постсинаптической мембране нервного волокна; VI этап — электротоническое распространение генераторного потенциала по нервному волокну; VII этап — генерация потенциала действия электрогенными участками нервного волокна; VIII этап — проведение потенциала действия по нервному волокну в ортодромном направлении.
МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ВОЗБУЖДЕНИЯ ВО ВТОРИЧНОЧУВСТВУЮЩИХ РЕЦЕПТОРАХ I-III этапы - совпадают с такими же этапами первичночувствующих рецепторов, но протекают они в специализированной рецептирующей клетке и заканчиваются на ее пресинаптической мембране; IV этап — выделение медиатора пресинаптическими структурами рецептирующей клетки; V этап — возникновение генераторного потенциала на постсинаптической мембране нервного волокна; VI этап — электротоническое распространение генераторного потенциала по нервному волокну; VII этап — генерация потенциала действия электрогенными участками нервного волокна; VIII этап — проведение потенциала действия по нервному волокну в ортодромном направлении.
 СВОЙСТВА ПЕРИФЕРИЧЕСКОГО ОТДЕЛА АНАЛИЗАТОРА 1. Специфичность. 2. Высокая чувствительность. 3. Способность к ритмической генерации импульсов возбуждения в ответ на однократное действие раздражителя. 4. Способность к адаптации. 5. Функциональная мобильность. 6. Низкая способность к аккомодации. 7. Специализация рецепторов. 8. Способность к элементарному первичному анализу. 9. Кодирование информации.
СВОЙСТВА ПЕРИФЕРИЧЕСКОГО ОТДЕЛА АНАЛИЗАТОРА 1. Специфичность. 2. Высокая чувствительность. 3. Способность к ритмической генерации импульсов возбуждения в ответ на однократное действие раздражителя. 4. Способность к адаптации. 5. Функциональная мобильность. 6. Низкая способность к аккомодации. 7. Специализация рецепторов. 8. Способность к элементарному первичному анализу. 9. Кодирование информации.
; • медленноадаптирующиеся (терморецепторы); • неадаптирующиеся") В зависимости от скорости адаптации различают: • быстроадаптирующиеся (тактильные); • медленноадаптирующиеся (терморецепторы); • неадаптирующиеся (вестибулярные и проприорецепторы). Выделяют несколько видов адаптации • изменение возбудимости рецептора в сторону снижения — десенсибилизация; • изменение возбудимости в сторону повышения — сенсибилизация.
В зависимости от скорости адаптации различают: • быстроадаптирующиеся (тактильные); • медленноадаптирующиеся (терморецепторы); • неадаптирующиеся (вестибулярные и проприорецепторы). Выделяют несколько видов адаптации • изменение возбудимости рецептора в сторону снижения — десенсибилизация; • изменение возбудимости в сторону повышения — сенсибилизация.
 Свойства коркового отдела анализаторов • Имеет центральную и окружающую ее ассоциативную зону (по представлению И. П. Павлова — “ядро” и рассеянные элементы). • Принцип двойственной проекции сенсорных систем в кору больших полушарий, которые полушарий можно разделить на первичные и вторичные проекции. • Взаимодействие анализаторов. Осуществляется на различных уровнях — спинальном, ретикулярном и таламо-кортикальном, кортикальном.
Свойства коркового отдела анализаторов • Имеет центральную и окружающую ее ассоциативную зону (по представлению И. П. Павлова — “ядро” и рассеянные элементы). • Принцип двойственной проекции сенсорных систем в кору больших полушарий, которые полушарий можно разделить на первичные и вторичные проекции. • Взаимодействие анализаторов. Осуществляется на различных уровнях — спинальном, ретикулярном и таламо-кортикальном, кортикальном.
 Кодирование информации в различных отделах анализатора • Кодирование информации. Информация преобразуется рецепторами в универсальные для мозга сигналы — нервные импульсы. • Кодирование качества достигается избирательной чувствительностью рецепторов к определенному виду энергии и очень низкими порогами возбуждения. • Кодирование интенсивности. • Пространственное кодирование. • Временное кодирование.
Кодирование информации в различных отделах анализатора • Кодирование информации. Информация преобразуется рецепторами в универсальные для мозга сигналы — нервные импульсы. • Кодирование качества достигается избирательной чувствительностью рецепторов к определенному виду энергии и очень низкими порогами возбуждения. • Кодирование интенсивности. • Пространственное кодирование. • Временное кодирование.
 Кодирование качества • Принцип меченой линии или топической организации. (Пространственно упорядоченное расположение нейронов на различных уровнях сенсорных систем соответственно характеристикам их рецептивных полей. ) • Теория “структуры ответа” - рецепторы кодируют качественные особенности раздражителей структурой импульсного ответа. (Эта теория предполагала отсутствие жестких связей между рецепторами и центральными нейронами. )
Кодирование качества • Принцип меченой линии или топической организации. (Пространственно упорядоченное расположение нейронов на различных уровнях сенсорных систем соответственно характеристикам их рецептивных полей. ) • Теория “структуры ответа” - рецепторы кодируют качественные особенности раздражителей структурой импульсного ответа. (Эта теория предполагала отсутствие жестких связей между рецепторами и центральными нейронами. )
 ЧУВСТВИТЕЛЬНОСТЬ СЕНСОРНЫХ СИСТЕМ • Абсолютная чувствительность измеряется порогом реакции. Существуют субсенсорные неосознаваемые реакции. • Дифференциальная чувствительность измеряется порогом различения d. I. • Отношение Вебера / I = const d. I , где I – сила раздражения, d. I – едва ощущаемый ее прирост (порог различения)
ЧУВСТВИТЕЛЬНОСТЬ СЕНСОРНЫХ СИСТЕМ • Абсолютная чувствительность измеряется порогом реакции. Существуют субсенсорные неосознаваемые реакции. • Дифференциальная чувствительность измеряется порогом различения d. I. • Отношение Вебера / I = const d. I , где I – сила раздражения, d. I – едва ощущаемый ее прирост (порог различения)
 1/60 Кинестезия (поднятие тяжести)") Константа Вебера для разных сенсорных модальностей Зрение (яркий белый свет) 1/60 Кинестезия (поднятие тяжести) 1/50 Боль (термическая на коже) 1/30 Слух (звук средней высоты и громкости) 1/10 • Давление на кожу 1/7 • Обоняние 1/4 • Вкус поваренной соли 1/3 • •
Константа Вебера для разных сенсорных модальностей Зрение (яркий белый свет) 1/60 Кинестезия (поднятие тяжести) 1/50 Боль (термическая на коже) 1/30 Слух (звук средней высоты и громкости) 1/10 • Давление на кожу 1/7 • Обоняние 1/4 • Вкус поваренной соли 1/3 • •
 Закон Вебера-Фехнера Между интенсивностью стимула и частотой потенциалов действия существует логарифмическая зависимость — ощущение увеличивается пропорционально логарифму интенсивности раздражения. Эта зависимость получила название закона Вебера-Фехнера E = a log I + b E – величина ощущения I – сила раздражения a и b – константы для разных модальностей
Закон Вебера-Фехнера Между интенсивностью стимула и частотой потенциалов действия существует логарифмическая зависимость — ощущение увеличивается пропорционально логарифму интенсивности раздражения. Эта зависимость получила название закона Вебера-Фехнера E = a log I + b E – величина ощущения I – сила раздражения a и b – константы для разных модальностей
 Ни на одном уровне сенсорной системы не происходит восстановления стимула в его первоначальной форме, т. е. декодирование. Это основное отличие физиологического кодирования от большинства технических систем связи, где сообщение, как правило, восстанавливается в первоначальном, декодированном виде.
Ни на одном уровне сенсорной системы не происходит восстановления стимула в его первоначальной форме, т. е. декодирование. Это основное отличие физиологического кодирования от большинства технических систем связи, где сообщение, как правило, восстанавливается в первоначальном, декодированном виде.
 Физиология зрительного анализатора
Физиология зрительного анализатора
 Мышцы глаза. А – вид спереди, Б – вид сверху
Мышцы глаза. А – вид спереди, Б – вид сверху
 Зрительные пути
Зрительные пути
 ОПТИКА ГЛАЗА Глаз имеет систему линз с различной кривизной и различными показателями преломления световых лучей, включающую четыре преломляющих среды между: 1. воздухом и передней поверхностью роговицы; 2. задней поверхностью роговицы и водянистой влагой передней камеры; 3. водянистой влагой передней камеры и хрусталиком; 4. задней поверхностью хрусталика и стекловидным телом.
ОПТИКА ГЛАЗА Глаз имеет систему линз с различной кривизной и различными показателями преломления световых лучей, включающую четыре преломляющих среды между: 1. воздухом и передней поверхностью роговицы; 2. задней поверхностью роговицы и водянистой влагой передней камеры; 3. водянистой влагой передней камеры и хрусталиком; 4. задней поверхностью хрусталика и стекловидным телом.
 Аккомодация приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии. • Основная роль в процессе аккомодации принадлежит хрусталику, способному изменять свою кривизну. • У молодых людей преломляющая сила хрусталика может увеличиваться от 20 до 34 диоптрий.
Аккомодация приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии. • Основная роль в процессе аккомодации принадлежит хрусталику, способному изменять свою кривизну. • У молодых людей преломляющая сила хрусталика может увеличиваться от 20 до 34 диоптрий.
 Механизм аккомодации глаза
Механизм аккомодации глаза
 1 диоптрия равна преломляющей силе") Преломляющая сила любых оптических систем выражается в диоптриях (D) 1 диоптрия равна преломляющей силе линзы с фокусным расстоянием в 1 метр. Если сферическая (двояковыпуклая) линза сводит параллельные лучи в фокусной точке на 1 м позади линзы, то её преломляющая сила равна +1 диоптрии.
Преломляющая сила любых оптических систем выражается в диоптриях (D) 1 диоптрия равна преломляющей силе линзы с фокусным расстоянием в 1 метр. Если сферическая (двояковыпуклая) линза сводит параллельные лучи в фокусной точке на 1 м позади линзы, то её преломляющая сила равна +1 диоптрии.
 Миопия
Миопия
 Гиперметропия
Гиперметропия
 Схема строения сетчатки
Схема строения сетчатки
 2 вида фоторецепторов
2 вида фоторецепторов
 Распределение палочек и колбочек в сетчатке
Распределение палочек и колбочек в сетчатке
 ФОТОРЕЦЕПЦИЯ В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин состоит из белковой части (опсин) и хромофора — 11 -цис-ретиналя, под действием фотонов переходящего в транс-ретиналь (фотоизомеризация). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события: 1. активация родопсина в результате фотоизомеризации 2. каталитическая активация G белка (Gt, трансдуцин) родопсином 3. активация фосфодиэстеразы при связывании с Gt 4. гидролиз ц. ГМФ–фосфодиэстеразой 5. переход ц. ГМФ–зависимых Na+ каналов из открытого состояния в закрытое 6. гиперполяризация плазмолеммы фоторецепторной клетки 7. передача сигнала на биполярные клетки.
ФОТОРЕЦЕПЦИЯ В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин состоит из белковой части (опсин) и хромофора — 11 -цис-ретиналя, под действием фотонов переходящего в транс-ретиналь (фотоизомеризация). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события: 1. активация родопсина в результате фотоизомеризации 2. каталитическая активация G белка (Gt, трансдуцин) родопсином 3. активация фосфодиэстеразы при связывании с Gt 4. гидролиз ц. ГМФ–фосфодиэстеразой 5. переход ц. ГМФ–зависимых Na+ каналов из открытого состояния в закрытое 6. гиперполяризация плазмолеммы фоторецепторной клетки 7. передача сигнала на биполярные клетки.
 Схема образования и обесцвечивания родопсина
Схема образования и обесцвечивания родопсина
 Потенциалы фоторецептора
Потенциалы фоторецептора
 Потенциалы клеток сетчатки
Потенциалы клеток сетчатки
 ИОННЫЕ ОСНОВЫ ФОТОРЕЦЕПТОРНЫХ ПОТЕНЦИАЛОВ В темноте Na+ каналы мембраны наружных сегментов палочек и колбочек открыты, и ток течёт из цитоплазмы внутренних сегментов в мембраны наружных сегментов. Ток течёт также в синаптическое окончание фоторецептора, вызывая постоянное выделение нейромедиатора. Na+, K+– насос, находящийся во внутреннем сегменте, поддерживает ионное равновесие, компенсируя выход Na+ входом K+. Таким образом, в темноте ионные каналы поддерживаются в открытом состоянии и потоки внутрь клетки Na+ и Ca 2+ через открытые каналы обеспечивают появление тока (темновой ток).
ИОННЫЕ ОСНОВЫ ФОТОРЕЦЕПТОРНЫХ ПОТЕНЦИАЛОВ В темноте Na+ каналы мембраны наружных сегментов палочек и колбочек открыты, и ток течёт из цитоплазмы внутренних сегментов в мембраны наружных сегментов. Ток течёт также в синаптическое окончание фоторецептора, вызывая постоянное выделение нейромедиатора. Na+, K+– насос, находящийся во внутреннем сегменте, поддерживает ионное равновесие, компенсируя выход Na+ входом K+. Таким образом, в темноте ионные каналы поддерживаются в открытом состоянии и потоки внутрь клетки Na+ и Ca 2+ через открытые каналы обеспечивают появление тока (темновой ток).
 Механизм реакции палочек на фотон света МРН-2 - метародопсин-2; Т - трансдуцин ФДЭ - фосфодиэстераза
Механизм реакции палочек на фотон света МРН-2 - метародопсин-2; Т - трансдуцин ФДЭ - фосфодиэстераза
 ИОННЫЕ ОСНОВЫ ФОТОРЕЦЕПТОРНЫХ ПОТЕНЦИАЛОВ На свету т. е. когда свет возбуждает наружный сегмент, Na+ , каналы закрываются и возникает гиперполяризационный рецепторный потенциал. Этот потенциал, появившийся на мембране наружного сегмента, распространяется до синаптического окончания фоторецептора и уменьшает выделение синаптического медиатора — глутамата. Это немедленно приводит к появлению ПД в аксонах ганглиозных клетках. Таким образом, гиперполяризация плазмолеммы — следствие закрытия ионных каналов. Вышеописанный каскад реакций усиливает световой сигнал и объясняет удивительный факт, каким образом один фотон света — наименьшая возможная квантовая единица световой энергии — может вызывать измеряемый рецепторный потенциал величиной в 1 м. В, а 30 фотонов света могут повышать функциональную готовность палочкового аппарата сетчатки на 50%.
ИОННЫЕ ОСНОВЫ ФОТОРЕЦЕПТОРНЫХ ПОТЕНЦИАЛОВ На свету т. е. когда свет возбуждает наружный сегмент, Na+ , каналы закрываются и возникает гиперполяризационный рецепторный потенциал. Этот потенциал, появившийся на мембране наружного сегмента, распространяется до синаптического окончания фоторецептора и уменьшает выделение синаптического медиатора — глутамата. Это немедленно приводит к появлению ПД в аксонах ганглиозных клетках. Таким образом, гиперполяризация плазмолеммы — следствие закрытия ионных каналов. Вышеописанный каскад реакций усиливает световой сигнал и объясняет удивительный факт, каким образом один фотон света — наименьшая возможная квантовая единица световой энергии — может вызывать измеряемый рецепторный потенциал величиной в 1 м. В, а 30 фотонов света могут повышать функциональную готовность палочкового аппарата сетчатки на 50%.
 Перекрытие рецептивных полей соседних нейронов в зрительной коре
Перекрытие рецептивных полей соседних нейронов в зрительной коре
 Цветовосприятие — функция колбочек. Существует три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов. Трихромазия — возможность различать любые цвета — определяется присутствием в сетчатке всех трёх зрительных пигментов (для красного, зелёного и синего — первичные цвета). Эти основы теории цветного зрения предложил Томас Янг (1802) и развил Герман Гельмгольц.
Цветовосприятие — функция колбочек. Существует три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов. Трихромазия — возможность различать любые цвета — определяется присутствием в сетчатке всех трёх зрительных пигментов (для красного, зелёного и синего — первичные цвета). Эти основы теории цветного зрения предложил Томас Янг (1802) и развил Герман Гельмгольц.
 Поле зрения для объектов разного цвета. Пунктир – белый цвет
Поле зрения для объектов разного цвета. Пунктир – белый цвет
 Восприятие цвета
Восприятие цвета
 БИНОКУЛЯРНОЕ ЗРЕНИЕ
БИНОКУЛЯРНОЕ ЗРЕНИЕ
— дефекты цветового восприятия (преимущественно у мужчин; например, в Европе") Дихромазии (цветовая слепота, или дальтонизм)— дефекты цветового восприятия (преимущественно у мужчин; например, в Европе разные дефекты у мужчин составляют 8% общей популяции) по одному из первичных цветов — подразделяют на протанопии, дейтанопии и тританопии (от греч. первый, второй и третий [имеются в виду порядковые номера первичных цветов: соответственно красный, зелёный, синий]). Протанопия (страдает восприятие красного, примерно 25% случаев цветовой слепоты) развивается при связанном с хромосомой Х наследовании генного дефекта. Дейтанопия (цветовая слепота по восприятию зелёного, около 75% всех случаев; связанное с хромосомой Х наследование, полиморфизм гена). Тританопия (страдает преимущественно восприятие фиолетового цвета, дефектное зрение по синему и жёлтому). Аутосомное доминантное наследование дефектного гена. Полная цветовая слепота — ахромазия — встречается крайне редко. При ахромазии поражается весь колбочковый аппарат, и человек видит все предметы в разных оттенках серого.
Дихромазии (цветовая слепота, или дальтонизм)— дефекты цветового восприятия (преимущественно у мужчин; например, в Европе разные дефекты у мужчин составляют 8% общей популяции) по одному из первичных цветов — подразделяют на протанопии, дейтанопии и тританопии (от греч. первый, второй и третий [имеются в виду порядковые номера первичных цветов: соответственно красный, зелёный, синий]). Протанопия (страдает восприятие красного, примерно 25% случаев цветовой слепоты) развивается при связанном с хромосомой Х наследовании генного дефекта. Дейтанопия (цветовая слепота по восприятию зелёного, около 75% всех случаев; связанное с хромосомой Х наследование, полиморфизм гена). Тританопия (страдает преимущественно восприятие фиолетового цвета, дефектное зрение по синему и жёлтому). Аутосомное доминантное наследование дефектного гена. Полная цветовая слепота — ахромазия — встречается крайне редко. При ахромазии поражается весь колбочковый аппарат, и человек видит все предметы в разных оттенках серого.
 Слуховая система
Слуховая система
 Слуховой анализатор
Слуховой анализатор
 Среднее и внутреннее ухо в разрезе
Среднее и внутреннее ухо в разрезе
 Разрез улиткового хода
Разрез улиткового хода
 Кортиев орган
Кортиев орган
 Функция клеток органа Корти Глутамат
Функция клеток органа Корти Глутамат
 Потенциал улитки
Потенциал улитки
 Микрофонный потенциал улитки и потенциал действия, зарегистрированный у овального окна в ответ на щелчок
Микрофонный потенциал улитки и потенциал действия, зарегистрированный у овального окна в ответ на щелчок
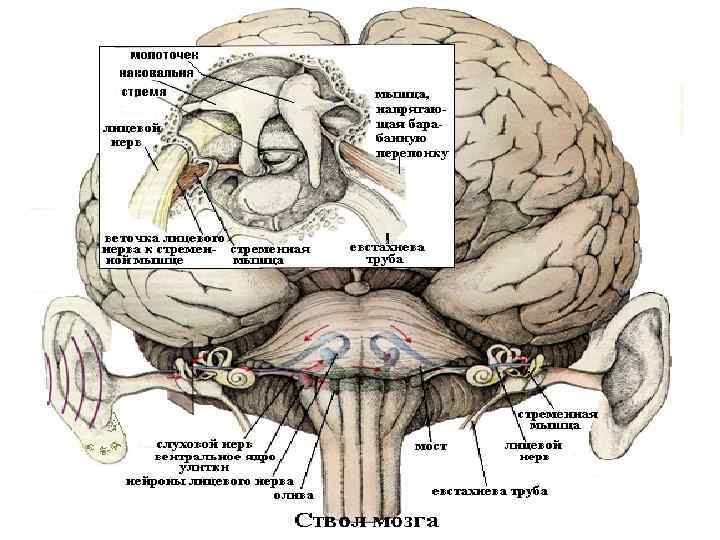
 Блок-схема слухового анализатора
Блок-схема слухового анализатора
 Слуховое поле
Слуховое поле
 Вестибулярный аппарат
Вестибулярный аппарат
 Система равновесия
Система равновесия
 И МАКУЛА (Б)") КУПУЛА (А) И МАКУЛА (Б)
КУПУЛА (А) И МАКУЛА (Б)
") Макулы органов преддверия (утрикулус и саккулус)
Макулы органов преддверия (утрикулус и саккулус)
 ФУНКЦИИ ВОЛОСКОВЫХ КЛЕТОК ВЕСТИБУЛЯРНОГО АППАРАТА
ФУНКЦИИ ВОЛОСКОВЫХ КЛЕТОК ВЕСТИБУЛЯРНОГО АППАРАТА
 ПОЛУКРУЖНЫЕ КАНАЛЫ
ПОЛУКРУЖНЫЕ КАНАЛЫ
 Афферентные и эфферентные связи вестибулярного аппарата
Афферентные и эфферентные связи вестибулярного аппарата
 Физиология боли
Физиология боли
 “Боль - это неприятное ощущение и эмоциональное переживание, возникающее в связи с настоящей или потенциальной угрозой повреждения тканей“. Международная Ассоциация по изучению боли (1994)
“Боль - это неприятное ощущение и эмоциональное переживание, возникающее в связи с настоящей или потенциальной угрозой повреждения тканей“. Международная Ассоциация по изучению боли (1994)
 Классификации боли: 1. По причине возникновения болевых ощущений: физиологическая - возникает как адекватная ответная реакция на действия повреждающего фактора. патологическая - возникает при поражении нервной системы или на действие неповреждающего фактора (каузалгия). 2. По времени возникновения и продолжительности болевых ощущений: острая - кратковременная, в виде приступов. хроническая - более длительная.
Классификации боли: 1. По причине возникновения болевых ощущений: физиологическая - возникает как адекватная ответная реакция на действия повреждающего фактора. патологическая - возникает при поражении нервной системы или на действие неповреждающего фактора (каузалгия). 2. По времени возникновения и продолжительности болевых ощущений: острая - кратковременная, в виде приступов. хроническая - более длительная.
 3. По локализации болевых ощущений: местная - в месте действия повреждающего фактора; проекционная - возникает в зоне иннервации повреждённого волокна. 4. По источнику: Висцеральная боль возникает при максимальном растяжении полых органов, действии химических веществ, из-за нарушения гемодинамики. Соматическая боль подразделяется на: а) поверхностную - возникает при раздражении экстерорецепторов (боль локализованная); б) глубокую - возникает при раздражении проприорецепторов (размытая боль).
3. По локализации болевых ощущений: местная - в месте действия повреждающего фактора; проекционная - возникает в зоне иннервации повреждённого волокна. 4. По источнику: Висцеральная боль возникает при максимальном растяжении полых органов, действии химических веществ, из-за нарушения гемодинамики. Соматическая боль подразделяется на: а) поверхностную - возникает при раздражении экстерорецепторов (боль локализованная); б) глубокую - возникает при раздражении проприорецепторов (размытая боль).
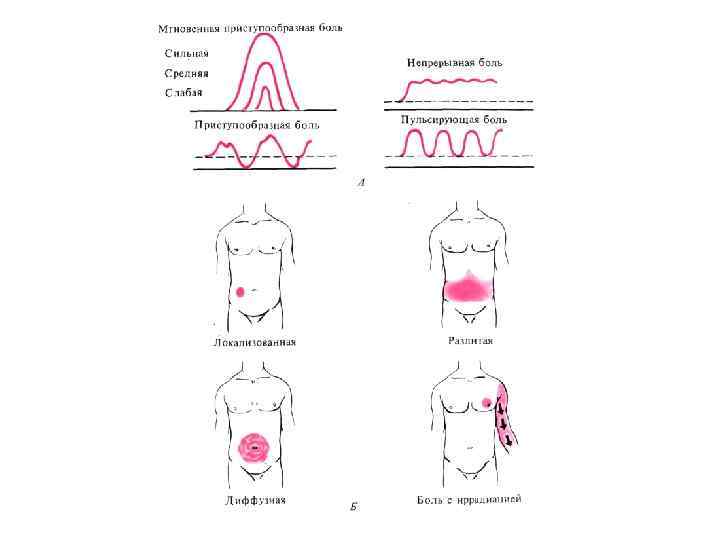
 и области происхождения каждого из них (серый фон) с примерами") Типы боли (розовый фон) и области происхождения каждого из них (серый фон) с примерами специфических форм боли
Типы боли (розовый фон) и области происхождения каждого из них (серый фон) с примерами специфических форм боли
 Боль состоит из 3 -х процессов: 1. возбуждение рецепторов; 2. проведение импульсов в центральную нервную систему и возбуждение центральных структур; 3. комплекс эффективных реакций, направленных на избавление организма от вредного фактора.
Боль состоит из 3 -х процессов: 1. возбуждение рецепторов; 2. проведение импульсов в центральную нервную систему и возбуждение центральных структур; 3. комплекс эффективных реакций, направленных на избавление организма от вредного фактора.
 - теория специфичности") Существует несколько теорий механизма возникновения болевых ощущений: теория Фрея (1895 г) - теория специфичности - болевые ощущения возникают при возбуждении специфических рецепторов (ноцицепторов). теория Гольдшейдера (1894 г) - теория интенсивности - болевые ощущения могут возникать в любых рецепторах, но при действии на них очень сильных раздражителей. Обе тории справедливы.
Существует несколько теорий механизма возникновения болевых ощущений: теория Фрея (1895 г) - теория специфичности - болевые ощущения возникают при возбуждении специфических рецепторов (ноцицепторов). теория Гольдшейдера (1894 г) - теория интенсивности - болевые ощущения могут возникать в любых рецепторах, но при действии на них очень сильных раздражителей. Обе тории справедливы.
 Структура и локализация кожных рецепторов Ноцицепторы кожи и слизистых оболочек представлены свободными неинкапсулированными нервными окончаниями, которые могут иметь самую разнообразную форму (спиралей, пластинок, волосков и др. ).
Структура и локализация кожных рецепторов Ноцицепторы кожи и слизистых оболочек представлены свободными неинкапсулированными нервными окончаниями, которые могут иметь самую разнообразную форму (спиралей, пластинок, волосков и др. ).
 По механизму возбуждения ноцицепторы делятся на: • механоноцицепторы, • хемоноцицепторы.
По механизму возбуждения ноцицепторы делятся на: • механоноцицепторы, • хемоноцицепторы.
 Механоноцицепторы реагируют на механические повреждения открытием Na+- каналов. Этот тип ноцицепторов реагирует не только на механические повреждения, но и на чрезмерные тепловые и холодовые раздражители. Возбуждение от механоноцицепторов проводится по дельта волокнам со скоростью 12 -30 м/с. Эти возбуждения обеспечивают ощущение быстрой, острой, хорошо локализованной боли — эпикритической боли. Трансмиттер дельта волокон остаётся неизвестным.
Механоноцицепторы реагируют на механические повреждения открытием Na+- каналов. Этот тип ноцицепторов реагирует не только на механические повреждения, но и на чрезмерные тепловые и холодовые раздражители. Возбуждение от механоноцицепторов проводится по дельта волокнам со скоростью 12 -30 м/с. Эти возбуждения обеспечивают ощущение быстрой, острой, хорошо локализованной боли — эпикритической боли. Трансмиттер дельта волокон остаётся неизвестным.
, под воздействием которых их постсинаптическая мембрана деполяризуется. Возбуждение") Хемоноцицепторы реагируют на химические вещества (алгогены), под воздействием которых их постсинаптическая мембрана деполяризуется. Возбуждение от хемоноцицепторов проводится по С -волокнам со скоростью 0, 5 -3 м/с и формирует ощущение медленной неприятной, плохо локализованной боли — протопатической боли. Нейротрансмиттером является субстанция P, а также связанный пептид, а волокна из внутренних органов – ВИП.
Хемоноцицепторы реагируют на химические вещества (алгогены), под воздействием которых их постсинаптическая мембрана деполяризуется. Возбуждение от хемоноцицепторов проводится по С -волокнам со скоростью 0, 5 -3 м/с и формирует ощущение медленной неприятной, плохо локализованной боли — протопатической боли. Нейротрансмиттером является субстанция P, а также связанный пептид, а волокна из внутренних органов – ВИП.
 медиаторы воспаления: брадикинин, гистамин, серотонин, ацетилхолин, простагландины, протеолитические ферменты, некоторые пептиды; б)") Алгогены а) медиаторы воспаления: брадикинин, гистамин, серотонин, ацетилхолин, простагландины, протеолитические ферменты, некоторые пептиды; б) ионы К+ (при концентрации выше 20 ммоль/л); в) ионы Н+ (при р. Н ниже 6), накапливающиеся при дефиците кислорода в клетке.
Алгогены а) медиаторы воспаления: брадикинин, гистамин, серотонин, ацетилхолин, простагландины, протеолитические ферменты, некоторые пептиды; б) ионы К+ (при концентрации выше 20 ммоль/л); в) ионы Н+ (при р. Н ниже 6), накапливающиеся при дефиците кислорода в клетке.
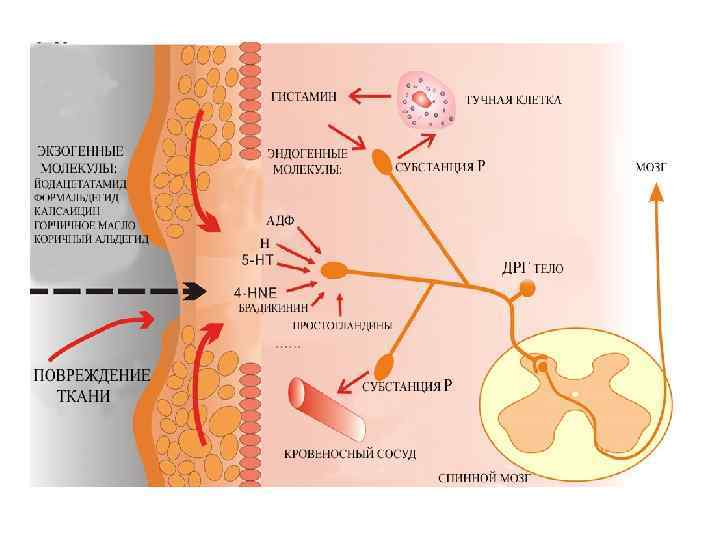
 Рис. Мембранно-ионные механизмы сенсинга боли при действии химических и физических раздражителей. Ноцицепторы используют различные ионные каналы для формирования рецепторного потенциала, где основная роль принадлежит семейству ваниллоид-зависимых TRP – каналов (transient receptor potential – кратковременные рецепторные потенциалы), обеспечивающих восприятие различных по природе стимулов.
Рис. Мембранно-ионные механизмы сенсинга боли при действии химических и физических раздражителей. Ноцицепторы используют различные ионные каналы для формирования рецепторного потенциала, где основная роль принадлежит семейству ваниллоид-зависимых TRP – каналов (transient receptor potential – кратковременные рецепторные потенциалы), обеспечивающих восприятие различных по природе стимулов.
 Ноцицептивная система 1. Ретикулярная формация 2. Таламус 3. Кора Первичное восприятия боли осуществляется соматосенсорной и фронто-орбитальной областями коры. Другие области коры участвуют в качественной ее оценке и в формировании ответных реакций на боль.
Ноцицептивная система 1. Ретикулярная формация 2. Таламус 3. Кора Первичное восприятия боли осуществляется соматосенсорной и фронто-орбитальной областями коры. Другие области коры участвуют в качественной ее оценке и в формировании ответных реакций на боль.
 Блок-схема проводящих путей болевой чувствительности
Блок-схема проводящих путей болевой чувствительности
 ► Ретикулярная формация - В РФ") Структуры и механизмы интеграции боли (ноцицептивная система) ► Ретикулярная формация - В РФ продолговатого мозга существуют нейроны, активирующиеся исключительно ноцицептивными стимулами. Наибольшее их количество (40 -60%) выявлено в медиальных ретикулярных ядрах. ► Таламус - 3 основных ядерных комплекса, имеющих непосредственное отношение к интеграции боли: вентро-базальный комплекс, задняя группа ядер, медиальные и интраламинарные ядра. ► Кора головного мозга - Процесс первичного восприятия боли осуществляется соматосенсорной и фронто-орбитальной областями коры. Другие области участвуют в качественной ее оценке, в формировании мотивационно-аффективных и психодинамических процессов, обеспечивающих переживание боли и реализацию ответных реакций на боль.
Структуры и механизмы интеграции боли (ноцицептивная система) ► Ретикулярная формация - В РФ продолговатого мозга существуют нейроны, активирующиеся исключительно ноцицептивными стимулами. Наибольшее их количество (40 -60%) выявлено в медиальных ретикулярных ядрах. ► Таламус - 3 основных ядерных комплекса, имеющих непосредственное отношение к интеграции боли: вентро-базальный комплекс, задняя группа ядер, медиальные и интраламинарные ядра. ► Кора головного мозга - Процесс первичного восприятия боли осуществляется соматосенсорной и фронто-орбитальной областями коры. Другие области участвуют в качественной ее оценке, в формировании мотивационно-аффективных и психодинамических процессов, обеспечивающих переживание боли и реализацию ответных реакций на боль.
 и передним (экстралемнисковым) трактами идущими") Восходящие пути болевой чувствительности представлены спиноталамическим латеральным (лемнисковым) и передним (экстралемнисковым) трактами идущими контрлатерально стороне вхождения болевых стимулов.
Восходящие пути болевой чувствительности представлены спиноталамическим латеральным (лемнисковым) и передним (экстралемнисковым) трактами идущими контрлатерально стороне вхождения болевых стимулов.
 Лемнисковый путь: через специфические ядра таламуса обеспечивает передачу сигналов о прикосновении к коже и давлении на нее. По этому пути передается быстрая наиболее тонкая информация, дифференцированная по силе и месту воздействия.
Лемнисковый путь: через специфические ядра таламуса обеспечивает передачу сигналов о прикосновении к коже и давлении на нее. По этому пути передается быстрая наиболее тонкая информация, дифференцированная по силе и месту воздействия.
 Экстралемнисковый путь: через неспецифические таламические ядра передают остальную ноцицептивную информацию по медленным нервным волокнам. На своём пути к таламусу тракт направляет часть волокон к РФ.
Экстралемнисковый путь: через неспецифические таламические ядра передают остальную ноцицептивную информацию по медленным нервным волокнам. На своём пути к таламусу тракт направляет часть волокон к РФ.
 Антиноцицептивная система
Антиноцицептивная система
 Структура антиноцицептивной системы 1. нервные механизмы, 2. гуморальные системы: - опиоидная, - моноаминергическая (норадреналин, дофамин, серотонин), - холинергическая, - ГАМКергическая.
Структура антиноцицептивной системы 1. нервные механизмы, 2. гуморальные системы: - опиоидная, - моноаминергическая (норадреналин, дофамин, серотонин), - холинергическая, - ГАМКергическая.
 В спинном мозге имеется желатинозная субстанция, ее функция") Теория “воротного контроля” (Мельзак, Уолл, 1965) В спинном мозге имеется желатинозная субстанция, ее функция – ограничивать сигналы идущие от тела к мозгу. Импульсы от тактильных рецепторов по быстрым волокнам проскакивают через нее (только часть) и активируют. Следующая сразу за этим импульсация не проходит. Сильные болевые сигналы способны затормозить желатинозную субстанцию – «открыть ворота» и прорваться в головной мозг.
Теория “воротного контроля” (Мельзак, Уолл, 1965) В спинном мозге имеется желатинозная субстанция, ее функция – ограничивать сигналы идущие от тела к мозгу. Импульсы от тактильных рецепторов по быстрым волокнам проскакивают через нее (только часть) и активируют. Следующая сразу за этим импульсация не проходит. Сильные болевые сигналы способны затормозить желатинозную субстанцию – «открыть ворота» и прорваться в головной мозг.
 Два обстоятельства могут закрыть ворота: 1. импульсы, проходящие по толстым “тактильным” волокнам (глубокая боль уменьшается контрраздражением) активируют желатинозную субстанцию 2. нисходящее (от ствола, Гт-Гф-системы, симпато-адреналовой системы) активирование желатинозных клеток предотвращает передачу информации выше (закрытие ворот изнутри).
Два обстоятельства могут закрыть ворота: 1. импульсы, проходящие по толстым “тактильным” волокнам (глубокая боль уменьшается контрраздражением) активируют желатинозную субстанцию 2. нисходящее (от ствола, Гт-Гф-системы, симпато-адреналовой системы) активирование желатинозных клеток предотвращает передачу информации выше (закрытие ворот изнутри).
 Уровни активации антиноцицептивной системы 1. На уровне ствола мозга включается нисходящая аналгетическая система (ядра шва через серотонин-, норадреналин-, энкефалинергические механизмы оказывает нисходящие влияния на желатинозную субстанцию спинного мозга).
Уровни активации антиноцицептивной системы 1. На уровне ствола мозга включается нисходящая аналгетическая система (ядра шва через серотонин-, норадреналин-, энкефалинергические механизмы оказывает нисходящие влияния на желатинозную субстанцию спинного мозга).
 2. Симпато-адреналовая система через ликвор и αадренорецепторы: - тормозит образование субстанции Р нейронами таламуса и центрального серого вещества мозга, обеспечивая глубокую анальгезию, - усиливает образование серотонина, который тормозит реализацию эффектов субстанции Р, - тормозит передачу болевой информации за счет образования эндогенных опиатов.
2. Симпато-адреналовая система через ликвор и αадренорецепторы: - тормозит образование субстанции Р нейронами таламуса и центрального серого вещества мозга, обеспечивая глубокую анальгезию, - усиливает образование серотонина, который тормозит реализацию эффектов субстанции Р, - тормозит передачу болевой информации за счет образования эндогенных опиатов.
 3. Гипоталамо-гипофизарная система - является основным местом синтеза и секреции эндогенных опиатов, - усиливает прямое влияние нейронов гипоталамуса на задние рога спинного мозга.
3. Гипоталамо-гипофизарная система - является основным местом синтеза и секреции эндогенных опиатов, - усиливает прямое влияние нейронов гипоталамуса на задние рога спинного мозга.
, 2. динорфины (а") Выделяют четыре семейства эндогенных опиатов: 1. эндорфины (а-, b- и у-), 2. динорфины (а и в), 3. неоэндорфины (альфа и бета), 4. энкефалины (метионин- и лейцин-). В аденогипофизе образуются эндорфины, динорфины и неоэндорфины. Энкефалины синтезируются в разных отделах мозга (прежде всего таламуса и гипоталамуса) и клетками слизистой оболочки кишечника.
Выделяют четыре семейства эндогенных опиатов: 1. эндорфины (а-, b- и у-), 2. динорфины (а и в), 3. неоэндорфины (альфа и бета), 4. энкефалины (метионин- и лейцин-). В аденогипофизе образуются эндорфины, динорфины и неоэндорфины. Энкефалины синтезируются в разных отделах мозга (прежде всего таламуса и гипоталамуса) и клетками слизистой оболочки кишечника.
 Механизм обезболивающего действия опиатов Опиаты соединяются с ноцицепторами и препятствуют соединению с ними нейротрансмиттера (субстанции P). Опиаты вызывают пресинаптическое торможение и уменьшают выделение дофамина, ацетилхолина, субстанции P, а также простагландинов. Опиаты вызывают угнетение аденилатциклазной системы и торможение выделения медиаторов.
Механизм обезболивающего действия опиатов Опиаты соединяются с ноцицепторами и препятствуют соединению с ними нейротрансмиттера (субстанции P). Опиаты вызывают пресинаптическое торможение и уменьшают выделение дофамина, ацетилхолина, субстанции P, а также простагландинов. Опиаты вызывают угнетение аденилатциклазной системы и торможение выделения медиаторов.
 Альтернативные механизмы обезболивания: 1. Вазопрессин, ангиотензин, окситоцин, соматостатин, нейротензин оказывают анальгетический эффект без активации опиатной системы. 2. Холинэргическая система усиливает морфийную систему, а ее блокада ослабляет.
Альтернативные механизмы обезболивания: 1. Вазопрессин, ангиотензин, окситоцин, соматостатин, нейротензин оказывают анальгетический эффект без активации опиатной системы. 2. Холинэргическая система усиливает морфийную систему, а ее блокада ослабляет.
 ОСТРАЯ БОЛЬ "ВОРОТНАЯ" ТЕОРИЯ Р. МЕЛЬЗАКА И П. УОЛЛА. Роль ворот, пропускающих ноцицептивные импульсы вверх, играют Т-клетки substantio gelatinosa заднего рога, осуществляющие пресинаптическое торможение терминалей, выделяющих субстанцию Р. В этих условиях болевые импульсы в ЦНС не проходят, и боль не возникает. При увеличении притока ноцицептивной афферентации происходит ингибирование клеток substantio gelatinosa. Это снимает тормозный эффект ее нейронов на ноцицептивные терминали и возникает боль вследствие облегчения передачи болевых импульсов в мозг.
ОСТРАЯ БОЛЬ "ВОРОТНАЯ" ТЕОРИЯ Р. МЕЛЬЗАКА И П. УОЛЛА. Роль ворот, пропускающих ноцицептивные импульсы вверх, играют Т-клетки substantio gelatinosa заднего рога, осуществляющие пресинаптическое торможение терминалей, выделяющих субстанцию Р. В этих условиях болевые импульсы в ЦНС не проходят, и боль не возникает. При увеличении притока ноцицептивной афферентации происходит ингибирование клеток substantio gelatinosa. Это снимает тормозный эффект ее нейронов на ноцицептивные терминали и возникает боль вследствие облегчения передачи болевых импульсов в мозг.
") ХРОНИЧЕСКАЯ БОЛЬ ► При длительном повреждении тканей (воспаление, переломы, опухоли и т. д. ) постоянная болевая информация, вызывая резкую активацию гипоталамуса и гипофиза, САС, лимбических образований мозга, сопровождается более сложными и продолжительными изменениями со стороны психики, поведения, эмоциональных проявлений, отношения к окружающему миру (уход в боль). ► Хроническая боль возникает в результате подавления тормозных механизмов, особенно на уровне задних рогов спинного мозга и таламуса. Под влиянием экзогенных и эндогенных факторов в определенных структурах ЦНС возникают генераторы патологически усиленного возбуждения (ГПУВ), вызывающие повышение возбудимости других нейронов.
ХРОНИЧЕСКАЯ БОЛЬ ► При длительном повреждении тканей (воспаление, переломы, опухоли и т. д. ) постоянная болевая информация, вызывая резкую активацию гипоталамуса и гипофиза, САС, лимбических образований мозга, сопровождается более сложными и продолжительными изменениями со стороны психики, поведения, эмоциональных проявлений, отношения к окружающему миру (уход в боль). ► Хроническая боль возникает в результате подавления тормозных механизмов, особенно на уровне задних рогов спинного мозга и таламуса. Под влиянием экзогенных и эндогенных факторов в определенных структурах ЦНС возникают генераторы патологически усиленного возбуждения (ГПУВ), вызывающие повышение возбудимости других нейронов.
 объясняются в основном дефицитом афферентной информации, в результате") Фантомные боли (боли в ампутированных конечностях) объясняются в основном дефицитом афферентной информации, в результате чего тормозное влияние Т-клеток на уровне рогов спинного мозга снимается, а любая афферентация из области заднего рога воспринимается как болевая.
Фантомные боли (боли в ампутированных конечностях) объясняются в основном дефицитом афферентной информации, в результате чего тормозное влияние Т-клеток на уровне рогов спинного мозга снимается, а любая афферентация из области заднего рога воспринимается как болевая.
 Отраженная боль Афференты внутренних органов и кожи связаны с одними и теми же нейронами заднего рога спинного мозга, которые дают начало спинноталамическому тракту. Поэтому афферентация, идущая от внутренних органов (при их поражении), повышает возбудимость и соответствующего дерматома, что воспринимается как боль в этом участке кожи.
Отраженная боль Афференты внутренних органов и кожи связаны с одними и теми же нейронами заднего рога спинного мозга, которые дают начало спинноталамическому тракту. Поэтому афферентация, идущая от внутренних органов (при их поражении), повышает возбудимость и соответствующего дерматома, что воспринимается как боль в этом участке кожи.
 Дерматомеры Отражённая боль обычно возникает в структурах, развивающихся из одного и того же эмбрионального сегмента, или дерматомера. Этот принцип называется «правилом дерматомера» . (сердце и левая рука , яичко и почки с мочеточниками).
Дерматомеры Отражённая боль обычно возникает в структурах, развивающихся из одного и того же эмбрионального сегмента, или дерматомера. Этот принцип называется «правилом дерматомера» . (сердце и левая рука , яичко и почки с мочеточниками).
 Положение зон Геда внутренних органов и их соотношение с дерматомами
Положение зон Геда внутренних органов и их соотношение с дерматомами
 Проецируемая боль - возникает при повреждении крупного нервного ствола в зоне его иннервации. Например, при ударе по локтевому нерву в месте, где он проходит под кожей, вызывает неприятные болезненные ощущения в областях, снабжаемых нервом. Это происходит потому, что активация афферентных нервных волокон у локтя проецируется в соответствующие области ЦНС.
Проецируемая боль - возникает при повреждении крупного нервного ствола в зоне его иннервации. Например, при ударе по локтевому нерву в месте, где он проходит под кожей, вызывает неприятные болезненные ощущения в областях, снабжаемых нервом. Это происходит потому, что активация афферентных нервных волокон у локтя проецируется в соответствующие области ЦНС.
 Нарушения функций организма при боли 1. Нарушение сна, сосредоточенности, полового влечения, повышенная раздражительность. 2. При хронической интенсивной боли резко уменьшается двигательная активность человека. Больной находится в состоянии депрессии, повышается болевая чувствительность в результате снижения болевого порога. 3. Небольшая боль учащает, а очень сильная замедляет дыхание вплоть до его остановки. 4. Может увеличиться ЧСС, системное АД, развиться спазм периферических сосудов. Кожные покровы бледнеют, а если боль непродолжительна, спазм сосудов сменяется их расширением, что проявляется покраснением кожи.
Нарушения функций организма при боли 1. Нарушение сна, сосредоточенности, полового влечения, повышенная раздражительность. 2. При хронической интенсивной боли резко уменьшается двигательная активность человека. Больной находится в состоянии депрессии, повышается болевая чувствительность в результате снижения болевого порога. 3. Небольшая боль учащает, а очень сильная замедляет дыхание вплоть до его остановки. 4. Может увеличиться ЧСС, системное АД, развиться спазм периферических сосудов. Кожные покровы бледнеют, а если боль непродолжительна, спазм сосудов сменяется их расширением, что проявляется покраснением кожи.
 5. 6. 7. 8. 9. Изменяется секреторная и двигательная функция ЖКТ. Сначала возбуждается СНС, этому усиление секреторной активности, а затем за счет активации ПСНС уменьшается секреция, замедляется моторика желудка и кишечника, возможна рефлекторная олиго- и анурия. При очень резкой боли появляется угроза развития шока. Биохимические изменения проявляются в виде повышения потребления кислорода, распада гликогена, гипергликемии, гиперлипидемии. Кардиалгии и головные боли сочетаются с подъемом АД, температуры тела, тахикардией, диспепсией, полиурией, повышенным потоотделением, тремором, жаждой, головокружением. Постоянным компонентом реакции на болевое воздействие является гиперкоагуляция крови.
5. 6. 7. 8. 9. Изменяется секреторная и двигательная функция ЖКТ. Сначала возбуждается СНС, этому усиление секреторной активности, а затем за счет активации ПСНС уменьшается секреция, замедляется моторика желудка и кишечника, возможна рефлекторная олиго- и анурия. При очень резкой боли появляется угроза развития шока. Биохимические изменения проявляются в виде повышения потребления кислорода, распада гликогена, гипергликемии, гиперлипидемии. Кардиалгии и головные боли сочетаются с подъемом АД, температуры тела, тахикардией, диспепсией, полиурией, повышенным потоотделением, тремором, жаждой, головокружением. Постоянным компонентом реакции на болевое воздействие является гиперкоагуляция крови.
 Главные методы облегчения боли
Главные методы облегчения боли
 Виды обезболивания Все многообразие вариантов анестезии можно разделить на два вида: ► Общее обезболивание предполагает использование средств и методов, воздействующих на центральные нервные структуры. ► Проводниковая анестезия действует на пути, проводящие болевую информацию, блокирует проведение этой информации.
Виды обезболивания Все многообразие вариантов анестезии можно разделить на два вида: ► Общее обезболивание предполагает использование средств и методов, воздействующих на центральные нервные структуры. ► Проводниковая анестезия действует на пути, проводящие болевую информацию, блокирует проведение этой информации.
 Методы общего обезболивания можно разделить на: 1. Методы общего обезболивания, не сопровождающиеся выключением коры головного мозга и сознания (применяется редко из-за трудности дозировки нейролептиков в сочетании с анальгетиками и/или кетамином). 2. Методы, которые сопровождаются выключением коры головного мозга и сознания больного. Эти методы носят название - наркоз.
Методы общего обезболивания можно разделить на: 1. Методы общего обезболивания, не сопровождающиеся выключением коры головного мозга и сознания (применяется редко из-за трудности дозировки нейролептиков в сочетании с анальгетиками и/или кетамином). 2. Методы, которые сопровождаются выключением коры головного мозга и сознания больного. Эти методы носят название - наркоз.
 Наркоз Название наркоза и его характер определяются основным анестетиком. Если наркотический и анальгетический эффекты достигаются с помощью нескольких веществ, то такой наркоз называется комбинированным. В настоящее время он применяется в большинстве случаев. Не нужно путать комбинированную анестезию с многокомпонентным наркозом.
Наркоз Название наркоза и его характер определяются основным анестетиком. Если наркотический и анальгетический эффекты достигаются с помощью нескольких веществ, то такой наркоз называется комбинированным. В настоящее время он применяется в большинстве случаев. Не нужно путать комбинированную анестезию с многокомпонентным наркозом.
 Многокомпонентный наркоз предполагает одновременное использование фармакологических агентов, воздействующих на разные нервные структуры: 1. Модулирующие и блокирующие болевой сигнал в зоне рецепторов и проводящих путей – результат – анальгезия. 2. Организующие диссоциацию связей между диэнцефальными образованиями и корой головного мозга – результат – гипнотический эффект. 3. Блокирующие вегетативные центры (гл. образом симпатические) – результат – снижение уровня вегетативных реакций. 4. Блокирующие нервно-мышечное проведение – результат –релаксация поперечно-полосатой мускулатуры.
Многокомпонентный наркоз предполагает одновременное использование фармакологических агентов, воздействующих на разные нервные структуры: 1. Модулирующие и блокирующие болевой сигнал в зоне рецепторов и проводящих путей – результат – анальгезия. 2. Организующие диссоциацию связей между диэнцефальными образованиями и корой головного мозга – результат – гипнотический эффект. 3. Блокирующие вегетативные центры (гл. образом симпатические) – результат – снижение уровня вегетативных реакций. 4. Блокирующие нервно-мышечное проведение – результат –релаксация поперечно-полосатой мускулатуры.
 2.") Опиатные пептиды 1. в ЦНС выполняют роль медиаторов и модуляторов (подавление болевых ощущений) 2. участвуют в регуляции адаптивного поведения и реакций организма на стресс, 3. модулируют секрецию гормонов адено- и нейрогипофиза (соматотропина, пролактина, гонадотропинов, вазопрессина). 4. регулируют процесс всасывания в ЖКТ, 5. угнетают секрецию панкреатического сока и соляной кислоты в желудке.
Опиатные пептиды 1. в ЦНС выполняют роль медиаторов и модуляторов (подавление болевых ощущений) 2. участвуют в регуляции адаптивного поведения и реакций организма на стресс, 3. модулируют секрецию гормонов адено- и нейрогипофиза (соматотропина, пролактина, гонадотропинов, вазопрессина). 4. регулируют процесс всасывания в ЖКТ, 5. угнетают секрецию панкреатического сока и соляной кислоты в желудке.
 ГОЛОВНАЯ БОЛЬ Головная боль является разновидностью отражённой боли, воспринимаемой как болевое ощущение, возникающее на поверхности головы. Многие виды болей возникают от болевых стимулов внутри черепа, другие — от раздражителей, расположенных снаружи черепа.
ГОЛОВНАЯ БОЛЬ Головная боль является разновидностью отражённой боли, воспринимаемой как болевое ощущение, возникающее на поверхности головы. Многие виды болей возникают от болевых стимулов внутри черепа, другие — от раздражителей, расположенных снаружи черепа.
 Головные боли внутричерепного происхождения Сам мозг полностью лишен болевой чувствительности. Поэтому причины болей: 1. Давление на венозные синусы, окружающие мозг 2. Растяжение твёрдой мозговой оболочки. Особенно чувствительны структуры средней мозговой артерии. 3. Воспаление мозговых оболочек - менингеальные боли - наиболее тяжёлый вид головных болей, отражающихся по всей поверхности головы. 4. Снижении давления спинномозговой жидкости
Головные боли внутричерепного происхождения Сам мозг полностью лишен болевой чувствительности. Поэтому причины болей: 1. Давление на венозные синусы, окружающие мозг 2. Растяжение твёрдой мозговой оболочки. Особенно чувствительны структуры средней мозговой артерии. 3. Воспаление мозговых оболочек - менингеальные боли - наиболее тяжёлый вид головных болей, отражающихся по всей поверхности головы. 4. Снижении давления спинномозговой жидкости
 5. Эмоциональное напряжение приводит к корковой депрессии, которая вызывает локальное накопление ионов калия в ткани мозга, инициирующее сосудистый спазм и приступы мигрени. 6. Алкогольная боль вызвана прямым токсическим раздражающим действием ацетальдегида на мозговые оболочки.
5. Эмоциональное напряжение приводит к корковой депрессии, которая вызывает локальное накопление ионов калия в ткани мозга, инициирующее сосудистый спазм и приступы мигрени. 6. Алкогольная боль вызвана прямым токсическим раздражающим действием ацетальдегида на мозговые оболочки.
 Головные боли внечерепного происхождения 1. Спазм мышц, прикреплённых к черепу и плечевому поясу - боль поверхности головы напоминает внутричерепную боль. 2. Раздражение носовой полости и придаточных пазух носа - боль фронтальной поверхности головы небольшой интенсивности. 3. При нарушениях зрения, при сильных сокращениях ресничной мышцы - рефлекторный спазм лицевых и наружных глазных мышц и появление головной боли.
Головные боли внечерепного происхождения 1. Спазм мышц, прикреплённых к черепу и плечевому поясу - боль поверхности головы напоминает внутричерепную боль. 2. Раздражение носовой полости и придаточных пазух носа - боль фронтальной поверхности головы небольшой интенсивности. 3. При нарушениях зрения, при сильных сокращениях ресничной мышцы - рефлекторный спазм лицевых и наружных глазных мышц и появление головной боли.
 Нейрогенная боль Такая боль имеет ряд особенностей: 1. характеризуется как тупая, пульсирующая, давящая, обжигающая и стреляющая. 2. частичная потеря чувствительности. 3. характерны вегетативные расстройства, такие как снижение кровотока, гипер и гипогидроз в болевой области. 4. она не мешает засыпанию пациента, но возможно просыпание от сильной боли. 5. она невосприимчива к опиатам в обычных анальгетических дозах (механизм отличен от опиоид-чувствительной боли).
Нейрогенная боль Такая боль имеет ряд особенностей: 1. характеризуется как тупая, пульсирующая, давящая, обжигающая и стреляющая. 2. частичная потеря чувствительности. 3. характерны вегетативные расстройства, такие как снижение кровотока, гипер и гипогидроз в болевой области. 4. она не мешает засыпанию пациента, но возможно просыпание от сильной боли. 5. она невосприимчива к опиатам в обычных анальгетических дозах (механизм отличен от опиоид-чувствительной боли).