Microelements_2.ppt
- Количество слайдов: 44



Фізіологічна роль мікроелементів у рослин доц. Баранов В.

Значення МЕ • З часів Римської імперії і до 1804 р(опублікована праця Соссюра по живленню рослин) врожай пшениці в Європі складав 4 -7 ц/га, • через 100 років – врожайність збільшилась у 3 -4 рази. • Зараз із застосуванням МЕ та регуляторів окремі сорти дають 60 ц/га

Етапи розвитку вчення про МЕ • 1 етап • Французький вчений Ролен – 1869 –Мп на ріст плісньових грибів і у1870 – Zn на ріст A. niger. Тімірязев - Zn на ріст вищих рослин • 1885 -1890 – Петербург Медико-Хірургічна академія – Боткін, Зайковський – вплив Cu, Fe, Mn, Ni на організм людей і тварин • 1897 –Габрієль Бертран- Mn є кофактором оксидаз –вперше вказав, що МЕ можуть бути компонентом ферментів • Вчення про МЕ в цей час називають “фізіологія подразливості” – вважали, що вони навіть при стимуляції росту не є необхідними для рослин. • Чому так ? ( 1. в дослідах використовувались погано очищені макроелементи; 2. При аналізах грунту не враховувалось, що МЕ сльніше зв*язуються орг частиною грунту, ніж макроелементи)

Етапи розвитку вчення про МЕ • 2 етап • 1919 рік – роботи Вернадського – вчення про біогеохімічні “провінції” • 1919 - Штейнберг – розробив методику очищення солей від Zn і додавання його до A. niger збільшило його ріст у 2000 разів • Далі – роботи Мак-Харга з Mn, Уорінга з B, Сомер і Ліпман з Zn. Cu • Мічурін – Проростки мигдалю при обробці Mn за 1 рік виросли до 180 см і на другий рік зацвіли і дали плоди( звичайно цвіте на 6 -8 рік)
 • 1. Організм не може завершити свій")
Критерії оцінки потреби МЕ організму(Арнон, Стоут, 1939) • 1. Організм не може завершити свій життєвий цикл без цього МЕ • 2. Дія елемента на організм є прямою • 3. Дія цього елемента специфічна і він не може бути замінений іншим • (Поправка до пункту 3 -Мо в азотфіксації може замінятись ванадієм, карбоангідраза стимулюється різними МЕ, etc)

Фізіологічна роль мікроелементів Дані елементи входять до складу кофакторів і простетичних груп ферментів і контролюють такі життєво важливі процеси, як фотосинтез, дихання, перетворення речовин, ріст і розвиток рослин, виконуючи тим самим метаболічну функцію. Крім того, в іонному стані мікроелементи активізують роботу різних ферментних систем клітин і виконують регуляторну функцію. В той же час кожен мікроелемент відрізняється певною специфікою дії.

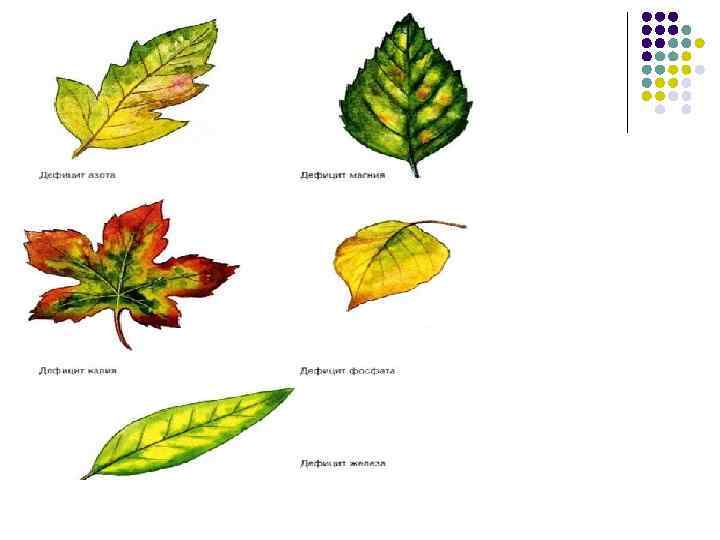
Напівмікроелементи l l l Залізо Fe 2+, Fe 3+ Середній вміст у грунтах коло 3%, а в рослинах 150 -160 мг на кг сухої маси Стимулює синтез хлорофілів, впливає на окисно-відновні процеси, ріст рослин за рахунок впливу на синтез ауксинів Переносник електронів при диханні, фотосинтезі, відновленні нітрогену і сульфуру. Входить до складу каталази, пероксидази, цитохромних систем Залізо не реутилізується Концентрується у нетоварній масі врожаю – у кукурудзи 44% є у коренях, в зерні -15%, залишок знаходиться у листках, стеблах качанах і їх обгортках

Ознаки дефіциту заліза l l l Тёмно-зелене забарвлення листків, при затримці росту надземних частин рослин і коренів, темно-коричневі до пурпурових листки на деяких рослинах (наприклад «бронзова хвороба» рису або тютюну) Сповільнення синтезу попередників хлорофілу Поява біло-жовтих плям між жилками, пізніше лист біліє повністю Ознаки нестачі зустрічаються рідко, в основному на бідних гумусом лужних грунтах. На кислих грунтах рослини частіше страждають від надлишку заліза.
 необхідний натрій. Це")
Напівмікроелементи l Натрій l Для росту окремих рослин засолених грунтів (галофітів) необхідний натрій. Це проявляється у рослин С 4 и САМ типу. У цих рослин показана необхідність натрію для регенерації ФЕП при карбоксилюванні. Дефіцит натрію у них приводить до хлорозу и некрозів, а також тормозить розвиток квітів. Натрій потребують і С 3 -рослини в малих кількостях. Показано, что цей елемент покращує ріст розтягом і выконує осморегулюючу функцію, подібно калію. Сприятливий вплив здійснює натрій на ріст цукрового буряку та ціанобактерій. l l l

Напівмікроелементи l l l l Хлор Вміст у рослинах відносно невеликий – насіння – 0. 02 -0. 07%на суху масу, солома і гичка – 0. 1 -0. 4%, стебла соняшника-1%, листки тютюну-до 3%, бульби картоплі- 0. 13%, листках буряка – 2. 5%, коренеплодах буряка – 0. 75%. Аніон хлору легко поступає у рослини і сприяє поглинанню катіонів, але інгібує поглинання аніонів, особливо фосфору. У великих кількостях пригнічує ріст, при нестачі також Йон хлору в клітинах частково зв*язаний з білками, але більша частина знаходиться у клітинному соці. Легко переміщується по рослині і так само як і оксиди сірки може поступати через листки Як правило рослини, які страждають від надлишку хлору, страждають і від надлишку кальцію. Бере участь у осмотичних процесах, нейтралізує заряди в мембранах, бере участь у реакції виділення кисню при фотосинтезі й у поділі клітин. При надлишку хлору у рослин спостерігається уповільнений ріст, загальний хлороз, поникання верхівок листків, погіршується якість продукції.

Напівмікроелементи l Кремній l За вмістом в земній корі на 2 місці після О 2. Є у всіх рослинах. В зерні - 0. 02 -0. 06%, соломі злакових -1. 33. 6%, у рису ще більше. Покращує поглинання фосфору, вочевидь за рахунок інгібування фосфатаз. Підвищує стійкість до хворіб та ушкодження комахами. Для росту діатомових водоростей потрібен кремній. Він покращує ріст деяких злаків, таких, як рис і кукурудза. Збільшує стійкість до вилягания, оскільки входить в склад клітинних стінок. Хвощі потребують кремній для проходження життєвого циклу. Однак і інші види аккмулюють досить кремнію - прискорюються темп росту і продуктивність. В гідратованій формі Si 02 кремний накопичується в ЕР, кліт-х стінках э межклітинному просторі. Може утворювати комплекс з поліфенолами і в цій формі замість лигніну служить для укріплення клітинних стінок. l l l l

Фізіологічна роль мікроелементів Мікроелементи Мідь Cu 2+ Стимулює біосинтез хлорофілу, антоціанів, сприяє синтезу білка і переносу електронів між ФС ІІ і ФС І. Впливає на інгібітори росту, підвищує стійкість рослини до посухи, морозу. Затримка росту і цвітіння, побіління у злаків кінчиків листків; пустоколосиця вівса, ячменю, у плодових – хлороз, некроз і суховершинність, ушкоджуються генеративні органи. Манган Mn 2+ Бере участь у відновленні нітратів до аміаку, у фотосинтезі (фотоліз води), відновленні СО 2, сприяє відтоку цукрів з листків. Активізує ферменти циклу Кребса, сприяє росту клітин. Ушкоджуються ферменти пероксидаза, ауксиноксидаза. Порушується співвідношення елементів мінерального Цинк Zn 2+ Активізує ферменти синтезу нуклеїнових кислот, а також дегідрогенази, ізомерази, ферменти синтезу триптофану Порушується синтез нуклеїнових кислот, гальмується поділ клітин, ріст рослин, утворюються дрібні розеткові листки, знижується морозовитривалість. Гальмується синтез триптофану.

Манган l Марганець поступає у виді йонів Мп 2+. Середній вміст в рослинах 0, 001 %. В клітинах марганець знаходиться в разніж степенях окислення (Мп 2+, Мп 3+, Мп 4+ ). Характеризується високим показником ОВП, з чим пов*язано його значення в реакціях окислення. Необхідний для фотосинтезу - входить в склад активного центру кисеньвидільного комплексу фотосистеми II и здійснює фотоліз води і виділення кисню: Приймає участь в відновленні С 02, підтримує структуру хлоропластів.

манган l У відсутність Мп хлорофілл швидко руйнується на світлі. Активує більше 35 ферментів в реакціях окислення-відновлення, декарбоксилювання і гідролізу. циклу Кребса, що пояснює значення для процесу дихання, особливо його аеробної фази. Приймає участь у відновленні нитратів до аміаку, Зв*язаний з синтезом білку через регуляцію активності ДНК-полімерази и РНКполімерази. Активує фермент окислення фітогормону ауксину.

цинк l l l l l Цинк поступає у вигляді виде йонів Zn 2+. Середній вміст в рослинах - 0, 002%. Цинк не бере участі у окисно -відновних реакціях, оскільк не міняє ступеню окислення. Входит в склад більше 30 ферментів, в т. ч. фосфатази, карбоангідрази, алкогольдегідрогенази, РНК-полимерази та ін. Поступлення цинку пасивне – додавання інгібіторів дихання не впливало на поступлення Слабо реутилізується В білках підтримує 3 -мірну структуру При дефіциті порушується поділ, кількість клітин зменшується, мітохондрії видовжуються і згортаються у спіраль, хлоропласти зменшуються або руйнуються. При дефіциті зменшується вміст глюкози і крохмалю, збільшується вміст органічних кислот При дефіциті зростав вміст амінокислот, нітратів, вміст глютаміну у 7 разів, аспарагіну у 50 разів. Дефіцит більше відбивається на утворенні насіння, ніж на процесах росту Збільшення дози цинку від 0. 05 до 0. 15 мг/л середовища вегетативна маса зросла у 2 рази, а врожай квітів і насіння конюшини у 100 разів.

Типова ознака дрібнолистяність При дефіциті цинку ріст пагонів послаблюється. Ламкі листки з плямами між жилками стають мілкими, асимметричними, гострозубчастими з металевим блиском. Грона рихлі з дрібними ягодами. Причиною нестачі частіше всього є надлишок удобрення фосфатами. Оприскування сульфатом цинку може послабит дефіцит.

l l l Медь поступає у вигляді йона Сu 2+ або Сu+. Середній вміст в рослинах - 0, 0002%. Мідь входить в склад ряду ферментних систем групи оксидаз, таких як ПФО, аскорбатоксидаза, цитохромоксидаза. В цих ферментах мідь з*єднана з білком через SH-группы. Поліфенолоксидаза і аскорбатоксидаза - окиснюють феноли і аскорбінову кислоту, а цитохромоксидаза входить в склад дихального ланцюга мітохондрій. мідь активує нітратредуктазу, а також протеази. Більша частина міді (75% від вмісту в листках) концентрується в хлоропластах, де входить в склад пластоціаніну. Мідь, як і залізо та манган, здатна до зворотнього окиснення та відновлення: Сu 2+ + е —> Сu+. тому пластоціанін приймає участь в переносі электронов від ФСII до ФСI. При дефициті міді зменшується активність першої фотосистеми. Дефіцит спостерігається на супіщаних і торфових грунтах – хвороби пшениці – череззерниця, біла “чума”

Фізіологічна роль мікроелементів Назва елемента Доступна форма Основні функції в процесі життєдіяльності рослини Ознаки прояву недостатності елемента Молібден Mo. O 42 - Входить до складу ферментів, що каталізують відновлення азоту, а також нітратів до нітритів. Сприяє переносу електронів в окисно-відновних реакціях, нагромадженню аскорбінової кислоти Затримка розвитку та росту, з'являються світлі листки, хлороз, некроз. Слабко розвиваються бульбочки на коренях бобових. Бор BO 33 - Синтез урацилу. Фенольний обмін. Нагромадження фенолів. Сприяє розвитку генеративних органів. Гальмування розвитку Сприяє засвоєнню мінеральних речовин. генеративних органів, відмирання пагонів, у цукрового буряка розвивається хвороба – гнилизна сердечка Хлор Cl-

молібден l l l Молібден поступає у вигляді аніону Мо 04 2 -. Вміст у рослинах 0, 0005— 0, 002%. Молібден входить в склад більше 20 ферментів, виконуючи не тільки каталітичну, але і структурну функцію. Молібден разом з залізом входить в склад нітрогенази у вигляді Mo-Fe-белок и приймає участь в фіксації азоту атмосфери мікроорганізмами. При нестачі молибдену зменшується синтез білку при одночасному падінні вмісту амінокислот та амідів. Порушення в азотному обміні особливо проявляються на фоні живлення рослин нітратами, так як молібден входить в активний центр фермента— нітратредуктази. При відновленні нітратів молібден діє як переносник электронів від ФАД до нітрату, при цьому N 03 - переходит в N 02 -, а Мо 5+ — в Мо 6+. цей фермент утворюється, коли в середовищі є нітрати і молібден. Активність нітратредуктази зростає в 10 разів і більше при живленні нітратами, порівнюючи з аміаком, При нестачі молібдену різко падає вміст аскорбінової кислоти. Спостерігаються порушення фосфорного обміну. Із здатністю молібдену до комплексоутворення пов*язаний його вплив на стабілізацію структури нуклеїнових кислот.

бор l l l l Бор поступає у вигляді аніону борної кислоти — Н 3 В 03 3 -. Вміст у рослинах 0, 0001%. бор, на відміну від більшості інших Ме, не входить в склад ні одного фермента и не є активатором ферментів. Велике значення має його здатність утворювати комплексні сполуки з простими цукрами, полівуглеводами, спиртами, фенолами та ін. . Тобто бор впливає на швидкість ферментативних реакцій через субстрати, на які діють ферменти. бор у комплексі з вуглеводами здійснює вплив на клітинну оболонку, регулюючи орієнтацію міцелл целюлози. При дефіциті бору, спостерігається швидка втрата еластичності оболонок клітин, що пов*язано з жорсткою оріентацією міцелл целюлози. Бор грає роль в підтримці структури мембран. При нестачі інгібується активність Н+-помпи, збільшується проникність мембран. Порушується синтез НК Видимі ознаки – опадання зав*язей, квітів і плодів.

бор l l l Комплекси цукрів з бором — цукроборати легше проникають через мембрани і швидше пересуваються по рослині. При нестачі бору цукри накопичуються в листках, і їх відтік різко тормозиться. В боронедостатніх рослинах заторможений процес амінування органічних кислот. Бор може виступати як інгибітор активності ряду ферментів, в першу чергу катализуючих утворення фенольних сполук. При нестачі бору накопичуються кофейна і хлорогенова кислоти - інгібітори росту рослин. Бор посилює ріст пилкових трубок, проростання пилку.

Нестача бору
")
кобальт l l Кобальт знаходиться в тканинах рослин в йонній (Со 2+, Со 3+) і комплексній формі. Вміст у рослинах 0, 00002%. Особливо необхідний бобовим рослинам, оскільки приймає участь в фіксації атмосферного азоту. Входить в склад кобаламіну (вітамін В 12 і його похідних), который синтезується бактеріями в бульбочках бобових рослин, а також входить в склад ферментів у азотфіксуючих організмів, приймає участь в синтезі метіоніну, ДНК и деленні клітин бактерій. При дефициті кобальту подавляєтся синтез леггемоглобіну, знижуєтся синтез білку, и зменшується розмір бактероїдів. Встановлена потреба в кобальті для вищих рослин, не здатних до азотфіксації. Показано вплив кобальту на функціонування фотосинтетичного аппарату, синтез білку, його зв*язок з ауксиновим обміном. Потреба в ньому дуже мала.

Нікель l l Нікель поступає в рослини у виді йона Ni 2+, але може также знаходитись у виді Ni+ и Ni 3+, Роль нікелю для вищих рослин як мікроэлемента була доведена недавно. До цього нікель вважали необхідним мікроелементом багатьох бактерій. У вищих рослин нікель входить в склад ферменту уреази, який відповідає за реакцію розкладу сечовины. Показано, що в рослинах, забеспечених нікелем, активність уреази вища і відповідно нищий вміст сечовини порівнюючи з незабеспеченими. Нікель активує ряд ферментів, в т. ч. нітратредуктазу, здійснює стабілізуючий вплив на структуру рибосом.

Поглинання МЕ l l l l l Поглинання звичайно проходить при дуже низьких концентраціях в розчинах. Поглинання сильно залежить від концентрації в розчині, особливо при низькому її рівні. Швидкість його сильно залежить від концентрациії Н+ та інших йонів. Интенсивність міняеться в залежності від виду рослини і стадії розвитку. Процеси поглинання чуттєві до температури, аерації, ОВП Поглинання може бути вибірковим. Накопичення йонов может бути протилежним їх градієнту концентрацій в грунті. В кругообігу МЕ між коренямии і середовищем важливу роль грає микориза. Головний шлях поступлення мікроелементів в растение – це абсорбція коренями,
 и")
Поглинання коренями l l l l Поглинання МЕ коренями може бути пасивним (неметаболічним) и активним (метаболичним). Пасивне поглинання проходить шляхом дифузії йонів з зовнішнього розчину в ендодерму коренів. При активному поглинанні необходна затрата енергії і воно направлене проти хімічного градіенту. Найбільш доступні рослинам ті мікроелементи, які адсорбовані на глинистих мінералах, тоді як фіксовані на оксидах і зв*язані мікроорганізмами є менш доступні. В поглинанні мікроелементів коренями приймають участь такі процеси: катіонний обмін з кореневою системою; переніс всередину клітин хелатоуиворюючими сполуками; дія ризосфери. йони та інші сполуки, які выділяють корені в оточуюче середовище, здійснюють вплив на поглинання коренями інших сполук

Поглинання листками- фоліарне поглинання l l фоліарне поглинання складається з двох фаз – неметаболічне проникнення через кутикулу, яке в цілому головним шляхом поступлення, и метаболічних процесів, якими пояснюється накопичення елементів, протипежне дії градієнтів концентрації. Друга фаза процессов відповідає за перенесення йонів через плазматичні мембрани в протоплазму клітин. МЕ, поглинені листками, можуть переноситись в інші тканини, включаючи і корені, где їх надлишок може відкладатись у запас. Швидкість руху МЕ в тканинах сильно змінюється в залежності від органу рослини, його віку і природи елемента. Частина МЕ, захоплена листками, может вымыватись з дощами.

Антагонізм йонів l l l Антагонізм виникає тоді, коли сумісна фізіологічна дія одного або більше елементів менше суми дії елементів, взятих окремо, а синергізм – коли сумісна дія більше. Антагоністичні ефекти частіше всього реалізуються двома способами: макрокомпонент може інгібувати поглинання мікроэлемента, або, навпаки, мікроелемент інгібує поглинання макрокомпонента. Ці реакції є досить частими для фосфатів, але спостерігалась інгібуюча дія інших іонів на поглинання елементів живлення. Для практичного застосування наибільш важнлива антагоністична дія Ca и Р на такі небеспечні для здоров*я людини важкі метали, як Be, Cd, Pb и Ni.

Макроелемент Антагонізм мікроелементами з Ca Al, Ba, Be, Cd, Co, Cr, Cs, Cu, Fe, Li, Mn, Ni, Pb, Sr, Cu, Mn, Zn Zn Mg Al, Be, Ba, Cr, Mn, F, Zn, Niа, Al, Zn Coа, Cuа, Feа P Al, As, B, Be, Cd, Cr, Cu, F, Fe, Al, B, Cu, F, Fe, Hg, Mo, Mn, Ni, Rb, Se, Si, Sr, Mo, Mn, Zn Zn K Al, B, Hg, Cd, Cr, F, Mn, Mo, Rb S As, Ba, Fe, Mo, Pb, Se Fб, Fe N B, F, Сu B, Cu, Fe, Mo Cl Br, I Синергізм

Дальній транспорт йонів l l l l Перенесення йонів в тканинах і органах рослин включає декілька процесів: Рух по ксилемі; Рух по флоемі; зберігання, накопичення і перехід в нерухомий стан. Хелатоутворюючі ліганди наибільш важливі для переносу катіонів в рослинах. Однак на рухомість металів в тканинах впливають також р. Н, ОВП, конкуренція між катіонами, гідроліз, полімеризація, утворення нерозчинних солей (фосфатів, оксалатів та інших. ) В цілому дальній транспорт МЕ в рослинах залежить від діяльності сосудистих тканин (ксилеми і флоемы) и частково зв*язаний з інтенсивністю транспірації. Хімічні форми МЕ в виділеннях флоеми різні для різних елементів.

Біологічна доступність МЕ l Наибільш достовірні методи встановлння доступності МЕ в грунтах – це методи, засновані на визначенні концентрацій елементів в грунтових розчинах, а не на визначенні запасу розчинних та/або здатних до обміну мікроелементів. При визначенні біодоступності МЕ дуже важливі специфічні властивості рослин. Вони досить сильно змінюються в залежності від умов грунту і стану рослин. Для эфективної оцінки запасу біологно доступних МЕ потрібно одночасно використовувати методики, засновані на вивченні аналізів стану грунту і вмісту в ньому МЕ і на даних анализу рослин.

МЕ в структурах клітини і біомолекулах l l l Входять в склад скелету рослин – Si, Fe, рідко Ba и Sr. Зв*язані в різні мілкі молекули, в тому числі антибіотики и порфірины – As, Br, Cu, Co, F, Fe, Hg, I, Se. Si и Мо Зв*язані с великими молекулами, головним чином з протеїнами, в тому числі энзимы, з каталітичними властивостями – Co, Cr, Cu, Fe, Mn, Mo, Se, Ni, Zn. Фиксовані у великих молекулах, включаючи накопичення, переніс або невідомі функції – Cd, Co, Cu, Fe, Hg, I, Mn, Ni, Se, Zn. Зв*язані с органелами (мітохондріями, хлоропластами, деякими ензиматичними системами) – Cu, Fe, Mn, Mo, Zn.

Токсичність и толерантність l l l l Метаболічні порушення викликаються не тільки нестачею МЕ, але і їх надлишком. Головні реакції, зв*язані з токсичною дією надлишку МЕ: Зміна проникності клітинних мембран – Ag, Au, Br, Cd, Cu, F, Hg, I, Pb, UO 2. Реакції тіольних груп з катіонами – Ag, Hg, Pb. Конкуренція з життєво важливими метаболітами – As, Sb, Se, Te, W, F. Велику спорідненість до фосфатних групп і активних центрів у АДФ и АТФ мають Al, Be, Sc, Y, Zr, лантаноїды і, вочевидь, всі важкі метали. Заміщення життєво важливих йонів (головним чином макрокатіонів) – Cs, Li, Rb, Se, Sr. Захоплення в молекулах позицій, які займають важливі функціональні групи, такі, як фосфат і нітрат – арсенат, фторид, бромат, селенат, теллурат, вольфрамат.

Токсичність йонів l l l Ряди мікроелементів за ступенем їх токсичності різні для кожного типу експериментів і кожної рослини, але досить добре корелюють з такими факторами: Електровід*ємність двохвалентних йонов; Здобуток розчинності сульфідів; Стійкість хелатів; біологічна доступність. Наибільш токсичними якк для вищих рослин, так і для ряду мікроорганізмів є Hg, Cu, Ni, Pb, Co, Cd, и, вочевидь, також Ag, Be, и Sn.

толерантність – це здатність зберігати життєдіяльність в умовах надлишку МЕ в середовищі, в основному в грунті. l l l Практичні завдання, які стосуються толерантних до металів організмів, можуть бути пов*язані з такими проблемами: мікробіологічне походження родовищ руд металів; кругообіг металів в середовищі; геоботанічні методи пошуку корисних копалин, мікробіологічна екстракція металів з збідненых руд; вырощування рослин на токсичних відходах; мікробіологічна очистка стічных вод;

Істинна толерантність пов*язана з внутрішніми факторами. Вона не являє собою певний єдиний механізм, а включає в себе кілька метаболічних процесів: l l l l селективне поглинання йонов; понижена проникність мембран або інші відміни в їх структурі функціях; іммобілізацію йонів в коренях, листках і насінні; Видалення йонов з метаболізму шляхом відкладеніня в фіксованих та/або нерозчинних формах; Зміна характеру метаболізму – посилення дії ензиматичних систем, які інгібуються, зростання вмісту антагоністичних метаболітів або відновлення ланцюгів метаболізму за рахунок пропуску інгібованої позиції; адаптацію до заміщення фізіологічного елемента токсичним у энзимі; Видалення йонів з рослин при вимиванні через листки, соковиділенні, скиданні листків і видаленні через корені.

МІКРОДОБРИВА l l l В принципі це ті самі солі, які використовуються у лабораторіях, але у мізерних концентраціях. В різних грунтах вміст МЕ неоднаковий, причому слід розрізняти валовий вміст і рухомі форми – валовий вміст може бути великим, але мало рухомих форм – тоді рослина відчуває нестачу по МЕ. МЕ використовуються як: 1. додаток до основних біогенних елементів(добрив), 2. ними опудрюють насіння перед посівом 3 -й шлях - позакоренева підкормка.

МІКРОДОБРИВА l l l При внесенні в грунт більшість мікроелементів дають в кількості 2 -3 кг/га, при опудрюванні насіння приблизно 25 -50 г МЕ/ центнер зерна, причому майже завжди додають у 4 -5 разів більше тальку, для рівномірності попадання МЕ на насіння. При позакореневому підживленні на 1 га дають 100 -300 г МЕ розчиненого у 300 -400 літрах води, тобто 50 -100 мг/л.

Марганцеві добрива l l l – Марганець – елемент зі змінною валентністю, бере участь в окисновідновних реакціях. На аміачному фоні живлення веде себе як сильний окислювач, а на нітратному – як відновник. гранульований суперфосфат з додаванням 10 -15% марганцевого шламу, що відповідає 1, 8 -2, 6% оксиду марганцю. Mn. SO 4 x 5 H 2 O- кристали світло-сірого кольору, містить 22, 8% марганцю.
. Входить в склад")
Молібденові добрива l l - Різка нестача спостерігається на кислих грунтах(Передкарпаття). Входить в склад ферментів азотного живлення – нітрогенази і нітратредуктази. Активують діяльність бульбочкових бактерій (нітрогеназу). Молібденізований суперфосфат- має приблизно 20% Р 2 О 5 та 0, 1% молібдену. Подвійний суперфосфат – 43% Р 2 О 5 і 0, 2% молібдену. (NH 4)2 Mo. O 4 - кристали білого кольору - містить коло 49% молібдену. Технічний молібдат амонію-натрію - (NH 4)2 Mo. O 4 + Na 2 MO 4 – біла з жовтим відтінком сіль – містить коло 35% молібдену – в звязку меншим вмістом молідену дозу застосування відповідно збільшують, порівнюючи з молібдатом.

Борні добрива l l l – нестача бору особливо відчувається на дерново-підзолистих грунтах і торфяниках. При вапнуванні кислих грунтів потреба рослин у борі зростає і їх ефективність збільшується. Гранульований боратовий суперфосфат – вміст бору не менше 0, 3 -0, 5% бору. Бормагнійсульфат – містить 13% борної кислоти(2, 27% бору) та 14% окису магнію – найбільш ефективний на легких піщанистих грунтах, де мало магнію. Борна кислота – 17% бору – для передпосівної обробки насіння використовують 0, 5% розчин.

Цинкові добрива l l l – Zn. SO 4 x 7 H 2 O – має 22% цинку. Цинкові полімікродобрива - порошок темно -сірого кольору, 19, 6% окису цинку, 17, 4 силікатного цинку, 21, 1 оксиду заліза, а також мідь, молібден, марганець і алюміній. Позакореневе підживлення – 0, 02 – 0, 05% сульфату цинку. При обробці плодових дерев концентрацію збільшують до 1 -5%.

ДЯКУЮ ЗА УВАГУ !
Microelements_2.ppt