лекция 6-2012.ppt
- Количество слайдов: 52



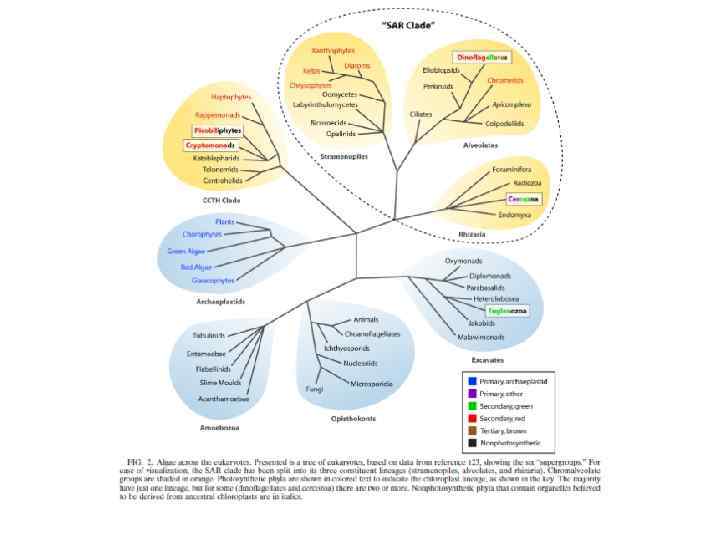
Филогения эукариот SARH

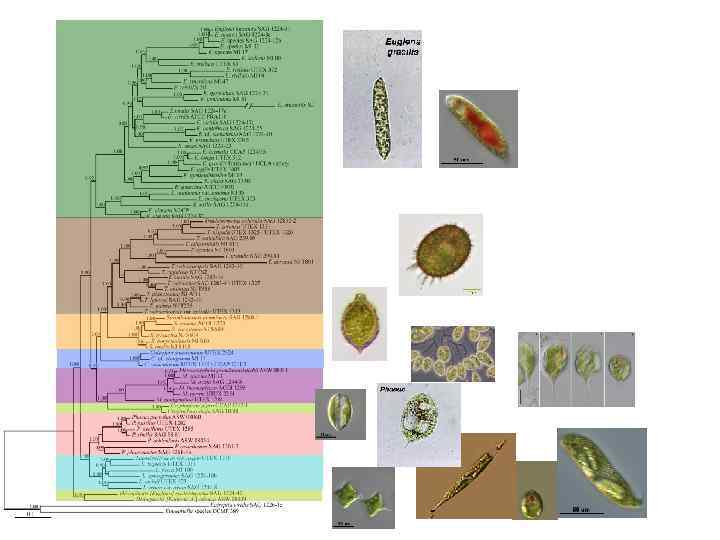
Euglenozoa Phylogeny based on molecular phylogenetic analyses primarily of ribosomal RNA genes and nucleus-encoded protein genes (Maslov et al. 1999; Закрытый митоз с внутриядерным веретеном Кристы митохондрий в форме теннисных ракеток, дисковидные В типе характерно два жгутика: вперед направлен спинной (дорсальный) жгутик и назад направлен брюшной (вентральный) жгутик В типе характерен жгутиковый аппарат, состоящий из трех микротрубочковых корешков: дорсального, промежуточного и вентрального Укрепленный микротрубочками вентральный или передний аппарат для питания (Mt. R карман) Гетероморфные параксонемные тяжи – относительно толстый жгутик Трубчатые экструсомы Simpson et al. , 2002, 2006; Simpson and Roger 2004; Moreira et al. 2004; Breglia et al. 2007). • •
 1. Одноклеточные и колониальные представители с монадным типом дифференциации")
Общая характеристика отдела Euglenida (Euglenophyta) 1. Одноклеточные и колониальные представители с монадным типом дифференциации таллома
 2. Обычно два гетеродинамичных жгутика, которые отходят от основания")
Общая характеристика отдела Euglenida (Euglenophyta) 2. Обычно два гетеродинамичных жгутика, которые отходят от основания жгутикового кармана (глотки). У большинства фотоавтотрофных и осмотрофных видов имеется видимый (жгутик для плавания) и один невидимый жгутик, который редуцирован в длину и не выходит из жгутикового кармана. У некоторых видов есть два или больше жгутика, а у немногих отсутствуют оба.
 3. Жгутики с параксиальными (параксонемными) тяжами: тяж дорсального жгутика")
Общая характеристика отдела Euglenida (Euglenophyta) 3. Жгутики с параксиальными (параксонемными) тяжами: тяж дорсального жгутика имеет трубчатую структуру, а в вентральном жгутике – решеткоподобную. Три микротрубочковых корешка: дорсальный, вентральный и промежуточный корешок. Жгутиковый карман у фотоавтотрофных видов изменен в резервуар и канал.
 4. Фотоавтотрофные представители реагируют на свет и его интенсивность,")
Общая характеристика отдела Euglenida (Euglenophyta) 4. Фотоавтотрофные представители реагируют на свет и его интенсивность, используя стигму или глазок и фоточувствительное вздутие при основании видимого жгутика.
 5. Хлоропласты окружены тремя мембранами, тилакоиды собраны по три.")
Общая характеристика отдела Euglenida (Euglenophyta) 5. Хлоропласты окружены тремя мембранами, тилакоиды собраны по три. Пиреноиды могут присутствовать, исключая Discoplastis, Phacus and Lepocinclis. Хлорофиллы a и b.
 6. Митохондрии с дисковидными ( в форме теннисных ракеток)")
Общая характеристика отдела Euglenida (Euglenophyta) 6. Митохондрии с дисковидными ( в форме теннисных ракеток) кристами.
 7. Ядро с постоянно конденсированными хромосомами и ядрышком.")
Общая характеристика отдела Euglenida (Euglenophyta) 7. Ядро с постоянно конденсированными хромосомами и ядрышком.
.")
8. Эвкариоворы, осмотрофные и фотоавтотрофные эвгленовые запасают парамилон (бета 1, 3 глюкан).
 9. Выделительные органеллы в форме трубчатых экструсом, мукоцист и")
Общая характеристика отдела Euglenida (Euglenophyta) 9. Выделительные органеллы в форме трубчатых экструсом, мукоцист и слизистых телец 10. Сложный аппарат Гольджи. 11. Пресноводные представители имеют сократительные вакуоли, связанные с резервуаром.
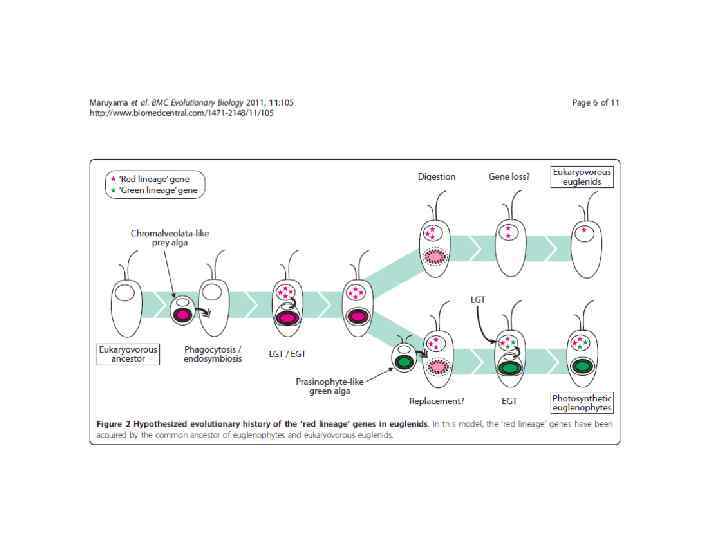
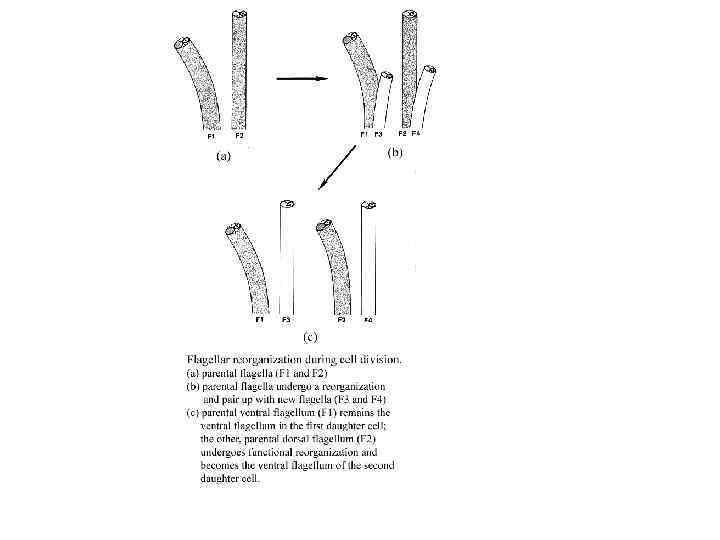
 12. Размножение вегетативное. Цитокинез идет с продольной бороздой деления.")
Общая характеристика отдела Euglenida (Euglenophyta) 12. Размножение вегетативное. Цитокинез идет с продольной бороздой деления.
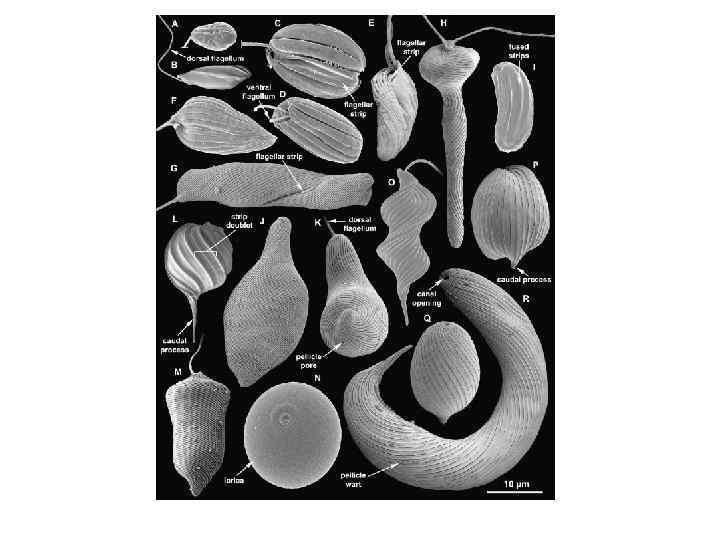
 13. Лучшая синапоморфная особенность группы – наличие пелликулы, состоящей")
Общая характеристика отдела Euglenida (Euglenophyta) 13. Лучшая синапоморфная особенность группы – наличие пелликулы, состоящей из S-образных белковых полос, лежащих ниже плазмолеммы, микротрубочек и цистерн эндоплазматического ретикулума. Пелликулярные полосы ориентированы продольно у бактериоворов и по спирали у эвкариоворов, фотоавтотрофов и осмотрофов. Полосы вторично продольные у некоторых регидных фотоавтотрофов (например, Phacus) и осмотрофов (например, Menoidium).

• Разнообразные и динамические способы передвижения, включая метаболию, скольжение по субстрату и плавание.

Среди эвгленовых встречаются представители с разными пищевыми стратегиями: Rhabdomonas – осмотрофы; Entosiphion развивает фаготрофный аппарат для поглощения бактерий, а Peranema – для поглощения эукариотных клеток; многие эвгленовые (например, Euglena, Trachelomonas, Phacus и др. ) – фотоавтотрофы. Способность к фотосинтезу была утеряна у Euglena longa (=Astasia longa), E. quartana (=Khawkinea quartana), E. hyalina, E. viridis hyalina, Phacus ocellatus (=Hylophacus ocellatus).
 корешка.")
• Аппарат для питания состоит из кармана, укрепленного микротрубочками от вентрального (брюшного) корешка. Карман для питания у многих бактериоворов и у эвкариоворов еще более укреплен жесткими тяжами и лопастями. Карман у фотоавтотрофных и осмотрофных представителей сильно редуцирован.
 образовывали токсины, обладающие ихтиотоксичностью. В 2009")
2 вида Euglena (E. sanguinea and E. granulata) образовывали токсины, обладающие ихтиотоксичностью. В 2009 году был идентифицирован эвгленофицин


Царство Alveolatae Cromera velia

Hypothetical tree of Apicomplexa. Three principal parasitic groups are coloured and their life cycle indicated, as well as Cryptosporidium that likely emerged from within gregarines. Numbers on branches and thickness indicates diversity (i. e. named species). © 2011 Jan Slapeta

Апикомплексы заболевание возбудитель • Малярия • Токсоплазмоз • Бабезиоз крупного рогатого скота • Лихорадка Восточного побережья крупного рогатого скота • Кокцидиоз цыплят • Криптоспоридиоз • Plasmodium spp • Toxoplasma gondii • Babesia spp. • Theileria spp. • Eimeria spp. • Cryptosporidium parvum

Геном апикопласта кодирует зо белков

Общая характеристика отдела Dinophyta 1. Преимущественно одноклеточные монадные представители с дорсо-вентральным строением

Общая характеристика отдела Dinophyta 2. Подвижные клетки имеют два жгутика, отличающихся как по морфологии, так и по длине. Поперечный жгутик длинный, уплощенный, имеет спиральнозакрученную аксонему и параксиальный тяж. Продольный жгутик короткий, обычного строения. Оба жгутика несут тонкие волоски.

Общая характеристика отдела Dinophyta 3. Хлоропласты окрашены в желто-бурый цвет, благодаря тому, что дополнительные пигменты перидинин, диадиноксантин и диноксантин маскируют зеленые хлорофиллы а и с2. Основной каротиноидный пигмент - перидинин, он поглощает световую энергию в синезеленой области спектра (470550 нм) и вместе с хлорофиллом образует водорастворимый перидинин - хлорофилл - белковый светособирающий комплекс. Пластиды с перидинином имеют трехмембранную оболочку и трехтилакоидные ламеллы, у них отсутствует опоясывающая ламелла.

Наиболее часто встречающийся тип пластид у динофитов – перединин содержащие, окруженные тремя мембранами. В этих пластидах обнаружена форма II Ру. Бис. Ко, известная также для некоторых бактерий, состоящая только из 2 -х больших субъединиц и кодирующаяся ядерным геномом. У ряда динофитовых с такими пластидами хлоропластный геном сильно редуцирован (осталось менее 20 действующих генов) и пластидная ДНК фрагментирована на отдельные кольцевые фрагменты 2 -3 кб, содержащие по одному гену. Такая форма хлоропластной ДНК и максимальная передача хлоропластных генов в ядро уникальна для водорослей. В хлоропластах встречаются пиреноиды различной формы. 4. Пресноводные формы запасают преимущественно крахмал, откладываемый в цитоплазме, а морские – липиды и стеролы.
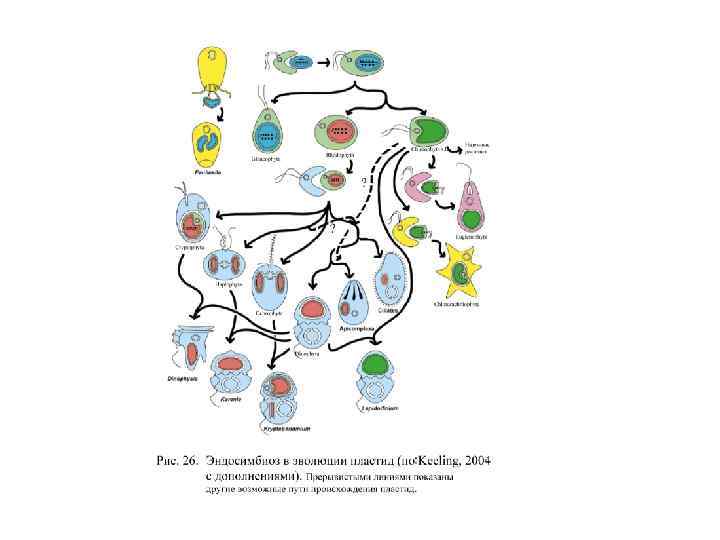
 photosynthetic organelles represent")
• Haptophyte-derived plastids: In the haptophore clade (Karenia, Karlodinium, Takayama) photosynthetic organelles represent real plastids - genes for many of their photosynthetic proteins have been shown to be encoded in the cell’s nucleus (Ishida and Green 2002, Patron et al. 2006, Nosenko et al. 2006). These type of endosymbiosis involving a secondary alga being engulfed by a eukaryote, is called ‘tertiary endosymbiosis’ (see Chromalveolate introductory page). Dinophysis mitra takes a haptophyte as kleptochloroplast (Koike et al. 2005). • • Prasinophyte-derived ‘plastids’: Permanent green chloroplasts in the genus Lepidodinium (Watanabe et al. 1991, Elbrächter and Schnepf 1996, Hansen et al. 2007). It is not known whether genetic material has moved from the endosymbiont into the host’s nucleus (although this seems likely). • • Diatom-derived photosynthetic endosymbionts: Some dinoflagellate species (e. g. , Durinskia baltica, Kryptoperidinium foliaceum, Peridinium quinquecorne) house nearly complete, permanent diatom endosymbionts with chloroplasts and nucleus (e. g. , Dodge 1971, Horiguchi and Pienaar 1991, 1994, Schnepf and Elbrächter 1999). This makes these host cells binucleate. Diatomophore dinoflagellates are culturable, they have been kept for years in culture collections. Nevertheless, it is interesting to note that non-photosynthetic, uninucleate versions of at least some of these species also exist in the wild, suggesting that the endosymbiosis is recent. It is not known whether any genes have moved from one nucleus to the other in diatomophore species. • • Dictyophyte-derived plastids: One dinoflagellate species (Podolampas bipes) houses complete, permanent dictyophycean endosymbionts with chloroplasts and nucleus (Schnepf and Elbrächter 1999, Schweikert and Elbrächter 2004). • • Cryptophyte-kleptochloroplasts: Most species of the genus Dinophysis harbour either complete cryptomonads or just their plastids (Schnepf and Elbrächter 1988, 1999, Janson 2004).

Общая характеристика отдела Dinophyta 5. В монадных клетках может присутствовать глазок, расположенный в хлоропласте или в цитоплазме. Глазки разнообразного строения Простейший глазок- это просто масса глобул, лежащих свободно в цитоплазме и не окруженных мембраной. Такой тип стигмы обнаружен у Woloszynskia coronata. Другой тип стигмы – это ряд глобул, локализованных в хлоропласте, например у Peridinium westii.

Третий тип представлен двумя рядами глобул, окруженных тройной мембраной у Kryptoperidinium foliaceum. Полагают, что такой глазок – сильно редуцированный перидинин – содержащий хлоропласт. Более сложный тип глазка, который встречается только у динофлагеллат, присутствует у Erythropsis и Nematodinium. У Erythropsis стигма состоит из пигментного тела и стекловидной линзы, преломляющей свет. Линзовидное тело представляет собой группу строго упорядоченных пузырьков. У Nematodinium помимо линзовидного тела в основании стигмы находится ретиноид - чашеподобное образование, содержащее зоны определенным образом ориентированных фибрилл и зернистого содержимого. На самом дне ретиноида в один ряд расположены пигментные глобулы.
 Erythropsis. (B) Eye organelle")
Eye organelle of the unicelllular dinoflagellates Erythropsis and Warnowia. (A) Erythropsis. (B) Eye organelle of Erythropsis. (C) Warnowia. (D) Eye organelle of Warnowia. (E) Nucleus and eye organelle of Warnowia. (F) Birefringence, the retina-like structure detected in polarized light in Warnowia. (G) Ultrastructure of the eye organelle of Warnowia. (H) Ultrastructure of the retinalike structure with stacked membranes and large pigment granules. A–F courtesy of Makiko Seimiya and Jean and Colette Febvre; G–H from Greuet 1969.

Общая характеристика отдела Dinophyta 6. Присутствует уникальное ядро – динокарион, его хромосомы находятся в конденсированном состоянии на протяжении всего клеточного цикла и содержат незначительное количество гистонов. Отсутствуют нуклеосомы. Митоз закрытый. Веретено деления внеядерное. Микротрубочки веретена проходят в каналах, ограниченных ядерной мембраной. Огромный геном.

Скручивание ДНК в хромосомах динофит ДНК динофитовых имеет уникальное азотистое основание – 5 – гидроксиметилурацил, замещающее 12 – 70 % тимина и обнаруженное только у бактерий. Вдобавок, ДНК динофлагеллат содержит 5 – метилцитозин и необычный N 6 – метиладенин.

Число хромосом у динофитов может варьировать от 12 -18 у Prorocentrum balticum, до 260 -280 у Ceratium hirundinlla и 500 -600 у отдельных видов Gymnodinium. Не исключено, что виды с большим числом хромосом могут быть полиплоидами, хотя имеются данные, опровергающие это мнение.

Динокарион имеется не у всех динофитовых. Так, у синдиниевых, являющихся внутриклеточными паразитами, ядра обычные эукариотные. В вегетативных клетках бластодиниевых и ноктилюковых также содержится простое эукариотное ядро, для которого характерно наличие деконденсированных хромосом в интерфазе. В то же время, в клетках их других стадий жизненного цикла, например в гаметах, присутствует динокарион. В ядре свободноживущего Oxyrrhis marina хромосомы менее конденсированы, чем в динокарионе, они имеют гистоны и во время митоза формируется внутриядерное веретено деления.

Особенности митоза

Общая характеристика отдела Dinophyta 7. Размножение чаще всего происходит продольным делением клетки пополам, встречается бесполое размножение зооспорами или апланоспорами. У некоторых видов описан половой процесс.

Жизненные циклы - гаплобионтный с зиготической редукцией, и только у Noctiluca - диплобионтный с гаметической редукцией, возможно, диплобионтный жизненный цикл у Pyrocystis lunula.

Амфиесма динофитов 8. Клеточный покров – амфиесма

9. Митохондрии с трубчатыми кристами 10. В клетках имеются характерные образования – пузулы Dinophyta

Стебельки – небольшие подвижные пальцевидные выросты, расположенные у основания жгутиков в борозде некоторых динофитов. Полагают, что у фаготрофных представителей они выполняют сенсорные функции, вовлеченные в процессы питания, а также функции прикрепления к субстрату. Щупальца – более удлиненные образования, известные у Noctiluca, Pronoctiluca, Pavillardia и др. У гетеротрофных представителей с их помощью происходит захват пищи и отправление ее к клеточному рту (цитостому). Поршень – более сложное устройство, имеющее вид полого цилиндрического выроста, с помощью которого пища подтягивается к цитостому. Он способен быстро удлиняться (в 15 раз может превышать длину тела), расширяться, а затем быстро сужаться и сокращаться. Поршень делает отверстие в клетке, которой питается, и ее цитоплазма через поршень движется в клетку динофлагеллаты. Такой поршень встречается у Gymnodinium fungiforme, зооспор Pfiesteria piscicida, фотосинтезирующей динофлагеллаты Amphidinium cryophilum.
 Protoperidinium depressum feeding on a chain of diatoms")
An illustration of the dinoflagellate (Dn) Protoperidinium depressum feeding on a chain of diatoms (Dt) using a pallium, a retractile organelle that spreads over the long spines of diatoms so that the contents can be digested. Illustration by D. M. Jacobson (reproduced from Jacobson, 1987). Вуаль – псевдоподиальная мембрана, которая выходит из борозды вблизи жгутиковой поры и имеет вид ловчей сети, по размерам превышающей размеры самой клетки. Отдельные крупные клетки или колониальные диатомеи прилипают к этой сети, которая обволакивает их, и происходит высасывание содержимого добычи. Продолжительность такого внеклеточного пищеварения – 20 – 30 минут. Такая вуаль известна для некоторых представителей родов Protoperidinium, Oblea и Diplopsalis, причем виды Protoperidinium питаются преимущественно диатомеями и динофлагеллатами, а другие, помимо диатомей, могут использовать гаптофитов, празинофициевых и криптофитов.
, так и в пресных водах. Свободноживущие,")
-12. Встречаются как в морских (около 90% видов), так и в пресных водах. Свободноживущие, паразиты, симбионты. Вызывают красные приливы.
.")
7% известных динофлагеллят – паразиты водных организмов (рыбы, ракообразные, радиолярии, инфузории, диатомеи, другие динофлагелляты). Большинство паразитических видов относится к порядкам Syndiniales и Blastodiniales

• Уникальная особенность, известная среди фотосинтезирующих организмов только у динофлагелляат, - биолюминесценция (тип свечения живых организмов, при котором энергия химических реакций превращается в световую). В основе биолюминисценции лежит реакция окисления люциферина с помощью фермента люциферазы. Она обнаружена примерно у 30 морских фотосинтезирующих видов динофлагеллят, относящихся к родам Gonyaulax, Protogonyaulax, Pyrodinium, Pyrocystis, Ceratium, и у некоторых бесцветных (Noctiluca и Protoperidinium). Причем, у одного и того же вида существуют биолюминесцентные и нелюминисцентные штаммы. Биолюминесцентные динофлагеллаты имеют сферические внутриклеточные структуры, называемые сцинтиллоны. Диаметр этих частиц - 0, 5 мкм, они располагаются по переферии клетки и окружены мембраной. Сцинтиллоны возникают в аппарате Гольджи и содержат люциферин, люциферазу и люциферин-связанные белки, причем, р. Н среды влияет на способность люциферина связываться с белками или реагировать с люциферазой.

Токсины динофит
лекция 6-2012.ppt