

 Ферменты 3
Ферменты 3
 Содержание: 1. Механизм регуляции активности ферментов. Аллостерическая регуляция. 2. Характеристика аллостерических ферментов. Виды ингибирования( обратимое, необратимое, конкурентное, неконкурентное, бесконкурентное) 3. Регуляция активности путем химической модификации ферментов. 4. Изоферменты. 5. Изменение активности ферментов в онтогенезе.
Содержание: 1. Механизм регуляции активности ферментов. Аллостерическая регуляция. 2. Характеристика аллостерических ферментов. Виды ингибирования( обратимое, необратимое, конкурентное, неконкурентное, бесконкурентное) 3. Регуляция активности путем химической модификации ферментов. 4. Изоферменты. 5. Изменение активности ферментов в онтогенезе.
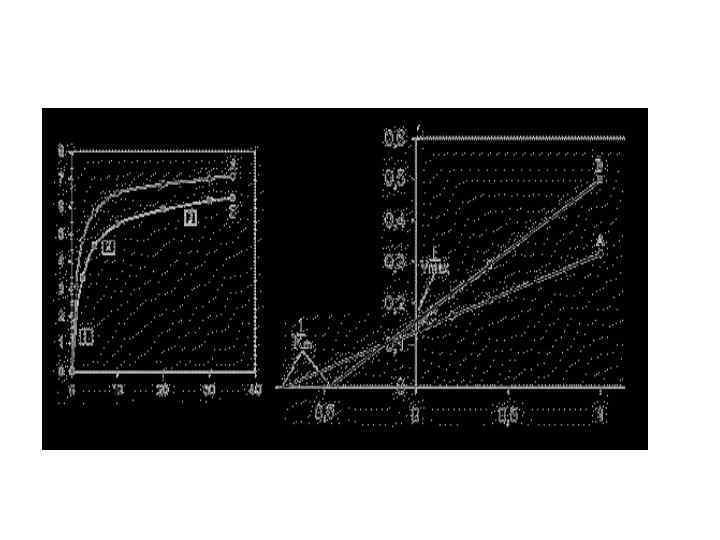
 • При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика всех ферментов основана на принципах Михаэлиса и Ментен. • Кинетика многих ферментов не подчиняется принципам кинетики Михаэлиса и Ментен
• При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика всех ферментов основана на принципах Михаэлиса и Ментен. • Кинетика многих ферментов не подчиняется принципам кинетики Михаэлиса и Ментен
 • У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов, протекающих в клетке, имеется ряд недостатков : • -узкий диапазон допустимых концентраций субстратов в клетке, способных для удовлетворения потребности, быстро реагировать на условия; • - потребность в тонкой регуляции концентраций большинства метаболитов для сохранения нормальной жизнедеятельности.
• У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов, протекающих в клетке, имеется ряд недостатков : • -узкий диапазон допустимых концентраций субстратов в клетке, способных для удовлетворения потребности, быстро реагировать на условия; • - потребность в тонкой регуляции концентраций большинства метаболитов для сохранения нормальной жизнедеятельности.
 • Поэтому природа вынуждена обратиться к "кооперативным" системам, в которых маленькие изменения в одном параметре, например концентрации ингибитора, вызывают большие изменения в скорости. Существует большое семейство ферментов, которые представляют собой кооперативные системы. Это аллостерические ферменты
• Поэтому природа вынуждена обратиться к "кооперативным" системам, в которых маленькие изменения в одном параметре, например концентрации ингибитора, вызывают большие изменения в скорости. Существует большое семейство ферментов, которые представляют собой кооперативные системы. Это аллостерические ферменты
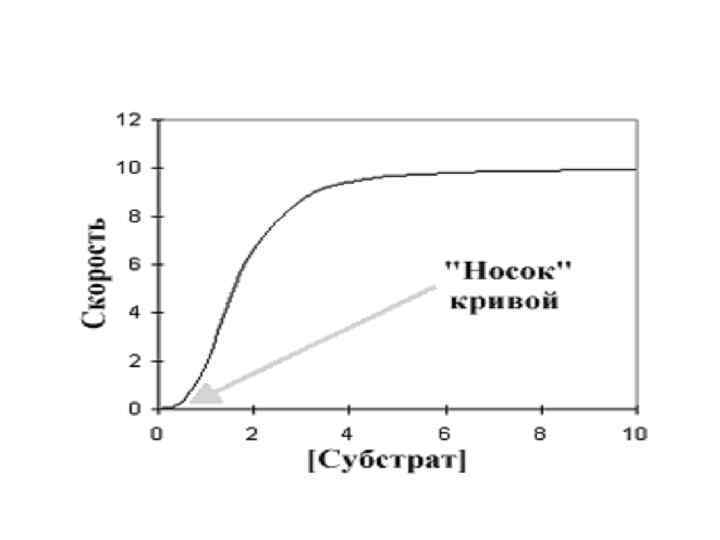
 • Графически результат работы такой кооперативной системы (график зависимости скорости реакции от концентрации субстрата) выражается не гиперболой, а S-образной сигмоидной кривой (сигмоидальные кривые всегда сдвинуты вправо в сравнении с гиперболой
• Графически результат работы такой кооперативной системы (график зависимости скорости реакции от концентрации субстрата) выражается не гиперболой, а S-образной сигмоидной кривой (сигмоидальные кривые всегда сдвинуты вправо в сравнении с гиперболой
 • Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся центра связывания лигандов (субстраты, ингибиторы и т. д), которые функционально взаимодействуют друг с другом. Связывание лиганда, с одним центром изменяет свойства другого (их). • Большая часть аллостерических белков – аллостерические ферменты, но некоторые белки, типа гемоглобина, выполняют и другие функции.
• Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся центра связывания лигандов (субстраты, ингибиторы и т. д), которые функционально взаимодействуют друг с другом. Связывание лиганда, с одним центром изменяет свойства другого (их). • Большая часть аллостерических белков – аллостерические ферменты, но некоторые белки, типа гемоглобина, выполняют и другие функции.
лиганда, белком, предшествующим связыванию другого лиганда.") • Кооперативность - это модификация константы связывания (Кs)лиганда, белком, предшествующим связыванию другого лиганда. Константы связывания - подобны Ks для субстрата или Ki для ингибитора, и являются в основном константами диссоциации белка и лиганда. Это указывает на силу связывания, или сродство белка и лиганда.
• Кооперативность - это модификация константы связывания (Кs)лиганда, белком, предшествующим связыванию другого лиганда. Константы связывания - подобны Ks для субстрата или Ki для ингибитора, и являются в основном константами диссоциации белка и лиганда. Это указывает на силу связывания, или сродство белка и лиганда.
 • Км обычно принимается как константа связывания субстрата, поскольку ее проще измерять, чем Ks. Понятие кооперативность означает, что связывание одного лиганда с белком, либо увеличит, либо уменьшит способность белка, связывать вторую молекулу лиганда. Если модификация увеличивает способность связывания (или сродство), это называют положительной кооперативностью. Если способность связывания снижается - это отрицательная кооперативность.
• Км обычно принимается как константа связывания субстрата, поскольку ее проще измерять, чем Ks. Понятие кооперативность означает, что связывание одного лиганда с белком, либо увеличит, либо уменьшит способность белка, связывать вторую молекулу лиганда. Если модификация увеличивает способность связывания (или сродство), это называют положительной кооперативностью. Если способность связывания снижается - это отрицательная кооперативность.
 Значения Vmax и Km определяются при экстраполяции линий до пересечения с осью абсцисс и ординат. Как видно исследуемый фермент отличается по значению Км от Км контрольного фермента более высокими значениями, что может свидетельствовать о более низком сродстве этого фермента к субстрату и возможном дефекте фермента
Значения Vmax и Km определяются при экстраполяции линий до пересечения с осью абсцисс и ординат. Как видно исследуемый фермент отличается по значению Км от Км контрольного фермента более высокими значениями, что может свидетельствовать о более низком сродстве этого фермента к субстрату и возможном дефекте фермента
 • Два лиганда один из которых влияет на связывание другого, могут быть химически идентичны, например, одна молекула субстрата, изменяет связывание другой молекулы субстрата. Такое взаимодействие называют гомотропным эффектом. Если они химически различаются, например, влияние ингибитора на связывание субстрата, тогда это - гетеротропный эффект.
• Два лиганда один из которых влияет на связывание другого, могут быть химически идентичны, например, одна молекула субстрата, изменяет связывание другой молекулы субстрата. Такое взаимодействие называют гомотропным эффектом. Если они химически различаются, например, влияние ингибитора на связывание субстрата, тогда это - гетеротропный эффект.
 У аллостерических ферментов особые свойства: • Полимерная структура • Сигмоидная ( в отличие от гиперболической для не аллостерических) форма кривой зависимости скорости реакции от концентрация субстрата • Существование эффекторов • Двухфазный ответ на конкурентные ингибиторы • Потеря аллостерических свойств при денатурации
У аллостерических ферментов особые свойства: • Полимерная структура • Сигмоидная ( в отличие от гиперболической для не аллостерических) форма кривой зависимости скорости реакции от концентрация субстрата • Существование эффекторов • Двухфазный ответ на конкурентные ингибиторы • Потеря аллостерических свойств при денатурации
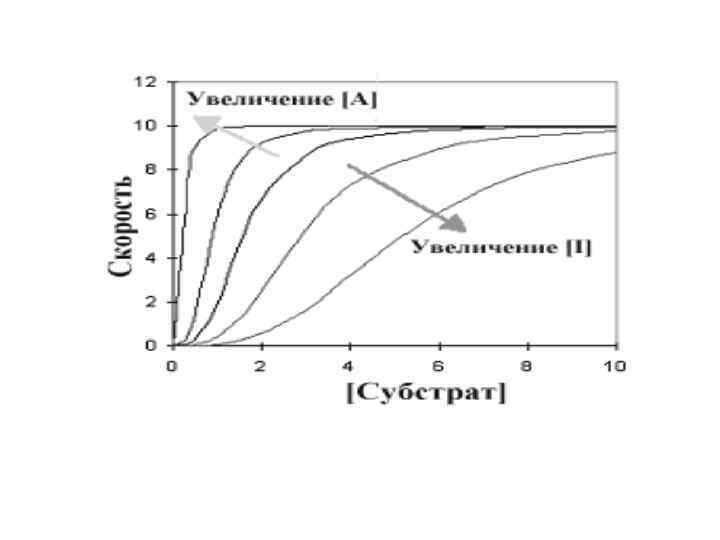
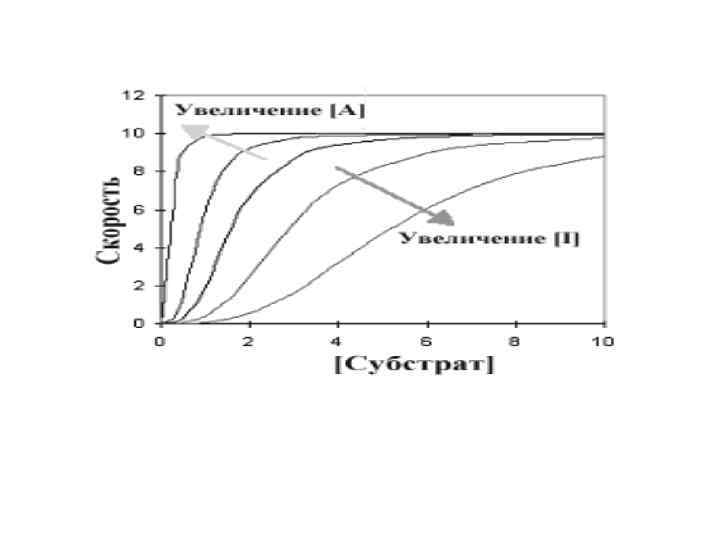
 • Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора. В присутствии активатора (А) скорость реакции повышается при любой данной концентрации субстрата, в то время как ингибитор уменьшает скорость реакции. Интересны и изменения общей формы кривой по сравнению с центральной линией.
• Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора. В присутствии активатора (А) скорость реакции повышается при любой данной концентрации субстрата, в то время как ингибитор уменьшает скорость реакции. Интересны и изменения общей формы кривой по сравнению с центральной линией.
 • Ингибитор увеличил сигмовидную форму, удлиняя «носок» кривой, в то время как активатор оказывал противоположный эффект. При более высокой концентрации активатора график в целом приобретает характер гиперболы. Это указывает на то, что аллостерический ингибитор увеличивает уровень субстратной кооперативности, в то время как активатор уменьшает его.
• Ингибитор увеличил сигмовидную форму, удлиняя «носок» кривой, в то время как активатор оказывал противоположный эффект. При более высокой концентрации активатора график в целом приобретает характер гиперболы. Это указывает на то, что аллостерический ингибитор увеличивает уровень субстратной кооперативности, в то время как активатор уменьшает его.
 • Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента. Денатурация вызывается рядом факторов, включая высокую температуру, экстремальные значения р. Н и химические денатурирующие реактивы типа мочевины.
• Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента. Денатурация вызывается рядом факторов, включая высокую температуру, экстремальные значения р. Н и химические денатурирующие реактивы типа мочевины.
 Инггибиторы бывают обратимые и необратимые • Вещества со свойствами ингибиторов ферментов можно грубо разделить на обратимые и необратимые. Обратимые ингибиторы связываются с ферментом, используя слабые связи, подобные тем, которые используются ферментом в связывании субстрата. Эти связи формируются быстро, но также быстро и легко разрушаются
Инггибиторы бывают обратимые и необратимые • Вещества со свойствами ингибиторов ферментов можно грубо разделить на обратимые и необратимые. Обратимые ингибиторы связываются с ферментом, используя слабые связи, подобные тем, которые используются ферментом в связывании субстрата. Эти связи формируются быстро, но также быстро и легко разрушаются
 • Следствием такого связывания обратимого ингибитора является эффективное мгновенное действие, но после удаления ингибитора фермент сохраняет свою активность. Ингибитор находится в равновесии с ферментом, формируя комплекс ингибитора фермента: Е +I EI (реакция обратима).
• Следствием такого связывания обратимого ингибитора является эффективное мгновенное действие, но после удаления ингибитора фермент сохраняет свою активность. Ингибитор находится в равновесии с ферментом, формируя комплекс ингибитора фермента: Е +I EI (реакция обратима).
 Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с ферментом, формируя прочные, обычно ковалентные связи: E+I = EI (реакция необратима).
Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с ферментом, формируя прочные, обычно ковалентные связи: E+I = EI (реакция необратима).
. Это соединение вошло") • Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение вошло в историю энзимологии как соединение использовавшееся для исследования роли химических групп в в структуре активного центра.
• Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение вошло в историю энзимологии как соединение использовавшееся для исследования роли химических групп в в структуре активного центра.
 ДИПФФ ковалентно связывается с гидроксильной группой серина и если эта группа важна в катализе реакции, фермент терял свою активность. Эти исследования позволило выявить группу ферментов, в активном центре которых активную роль играет серин (сериновые протеазы).
ДИПФФ ковалентно связывается с гидроксильной группой серина и если эта группа важна в катализе реакции, фермент терял свою активность. Эти исследования позволило выявить группу ферментов, в активном центре которых активную роль играет серин (сериновые протеазы).
 является необратимым ингибитором циклооксигеназы- фермента участвующего в синтезе") Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента участвующего в синтезе простагландинов.
Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента участвующего в синтезе простагландинов.
 Ингибитором синтеза протеогликанов стенки бактерий является пенициллин, структура которого напоминает Dаланин, встраиваемый в структуру протеогликанов. Связываясь с активным центром фермента бактерии благодаря своей схожести с переходным состоянием промежуточного продукта в активном центре, пенициллин образует ковалентную связь и тормозит работу фермента
Ингибитором синтеза протеогликанов стенки бактерий является пенициллин, структура которого напоминает Dаланин, встраиваемый в структуру протеогликанов. Связываясь с активным центром фермента бактерии благодаря своей схожести с переходным состоянием промежуточного продукта в активном центре, пенициллин образует ковалентную связь и тормозит работу фермента
 Различают два механизма конкурентного торможения. 1. Конкурентное торможение путем связывания активного центра. 2. Конкурентное торможение путем изменения конформации фермента.
Различают два механизма конкурентного торможения. 1. Конкурентное торможение путем связывания активного центра. 2. Конкурентное торможение путем изменения конформации фермента.
 Если связывание субстрата с АЦ фермента предшествует изменению конформации центра связывания ингибитора, то последний не связывает фермент.
Если связывание субстрата с АЦ фермента предшествует изменению конформации центра связывания ингибитора, то последний не связывает фермент.
 И субстрат и ингибитор не могут одновременно связаться с ферментом. В этом виде конкурентного торможения ингибитор может иметь любую химическую структуру, поскольку они связываются с различными участками фермента.
И субстрат и ингибитор не могут одновременно связаться с ферментом. В этом виде конкурентного торможения ингибитор может иметь любую химическую структуру, поскольку они связываются с различными участками фермента.
 Конкурентные ингибиторы не влияют на Vmax, они понижают Км. Оказывая эффект на Km, тем самым оказывают эффект на Vmax Наиболее часто приводимый пример конкурентного ингибирования - это использование малоновой кислоты для торможение дегидрогеназы янтарной кислоты. Наиболее близким структурным аналогом сукцината является малоновая кислота.
Конкурентные ингибиторы не влияют на Vmax, они понижают Км. Оказывая эффект на Km, тем самым оказывают эффект на Vmax Наиболее часто приводимый пример конкурентного ингибирования - это использование малоновой кислоты для торможение дегидрогеназы янтарной кислоты. Наиболее близким структурным аналогом сукцината является малоновая кислота.
 Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр на ферменте. Учитывая обратимость реакции, избыток янтарной кислоты снимет действие малоновой кислоты.
Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр на ферменте. Учитывая обратимость реакции, избыток янтарной кислоты снимет действие малоновой кислоты.
 Конкурентное торможение путем изменения конформации фермента. В отличие от классического варианта, ингибитор связывается не с активным центром, а со специальным центром, связывающим ингибитор, который расположен вдали от активного центра. Связывание ингибитора вызывает изменение пространственной структуры (изменение конформации) в области активного центра, которое не позволяет присоединиться субстрату.
Конкурентное торможение путем изменения конформации фермента. В отличие от классического варианта, ингибитор связывается не с активным центром, а со специальным центром, связывающим ингибитор, который расположен вдали от активного центра. Связывание ингибитора вызывает изменение пространственной структуры (изменение конформации) в области активного центра, которое не позволяет присоединиться субстрату.
 • Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования, ацетилирования, фосфорилирования (роль гормонов, АЦ-комплекса, ц. АМФ, ц. ГМФ, ионов Ca).
• Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования, ацетилирования, фосфорилирования (роль гормонов, АЦ-комплекса, ц. АМФ, ц. ГМФ, ионов Ca).
 Активность фермента можно изменить путем ковалентной модификации его структуры. Ковалентная модификация структуры ферментов может быть обратимой и необратимой.
Активность фермента можно изменить путем ковалентной модификации его структуры. Ковалентная модификация структуры ферментов может быть обратимой и необратимой.
 Гормональная регуляция активности ферментов Осуществляется при: • -функционировании АЦ комплекса, • - с помощью вторичных посредников • ц. АМФ, ц. ГМФ, Са-Кальмодулина, ИТФ, • ДАГ, NO, олигоаденилатами.
Гормональная регуляция активности ферментов Осуществляется при: • -функционировании АЦ комплекса, • - с помощью вторичных посредников • ц. АМФ, ц. ГМФ, Са-Кальмодулина, ИТФ, • ДАГ, NO, олигоаденилатами.
 Регуляция активности ферментов Гормональная регуляция осуществляется на генетическом уровне путем обратного фосфорилирования, и осуществляется при: -функционировании АЦ комплекса, - с помощью вторичных посредников- ц. АМФ, ц. ГМФ, Са-Кальмодулина, ИТФ, - ДАГ, NO, олигоаденилатами.
Регуляция активности ферментов Гормональная регуляция осуществляется на генетическом уровне путем обратного фосфорилирования, и осуществляется при: -функционировании АЦ комплекса, - с помощью вторичных посредников- ц. АМФ, ц. ГМФ, Са-Кальмодулина, ИТФ, - ДАГ, NO, олигоаденилатами.
 Аденилатциклазный комплекс • В ходе этого процесса образуется небелковое соединение - ц. АМФ, ц. АМФ - внутриклеточный гормон (вторичный посредник) яв-ся аллостерическим регулятором большого числа протеинкиназ. ц. АМФ образуется из АТФ под действием ферментааденилатциклазы
Аденилатциклазный комплекс • В ходе этого процесса образуется небелковое соединение - ц. АМФ, ц. АМФ - внутриклеточный гормон (вторичный посредник) яв-ся аллостерическим регулятором большого числа протеинкиназ. ц. АМФ образуется из АТФ под действием ферментааденилатциклазы
 В ходе этого процесса образуется небелковое соединение - ц. АМФ, ц. АМФ - внутриклеточный гормон (вторичный посредник) яв-ся аллостерическим регулятором большого числа протеинкиназ. ц. АМФ образуется из АТФ под действием ферментааденилатциклазы.
В ходе этого процесса образуется небелковое соединение - ц. АМФ, ц. АМФ - внутриклеточный гормон (вторичный посредник) яв-ся аллостерическим регулятором большого числа протеинкиназ. ц. АМФ образуется из АТФ под действием ферментааденилатциклазы.
 См. мультимедийную презентацию. Сигнальная трансдукция.
См. мультимедийную презентацию. Сигнальная трансдукция.
 Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует с поверхностью клетки, где расположены рецепторы (Rs и Ri), белки узнающие и связывающие гормон. Гормональный сигнал поступает на АЦ через белки посредники (Gs и Gi), которые активируются в условиях присоединения ГТФ.
Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует с поверхностью клетки, где расположены рецепторы (Rs и Ri), белки узнающие и связывающие гормон. Гормональный сигнал поступает на АЦ через белки посредники (Gs и Gi), которые активируются в условиях присоединения ГТФ.
 Уровень, образовавшийся под действием АЦ, ц. АМФ определяется не только активностью АЦ, но и активностью фосфодиэстераз, которые циклизируют ц. АМФ до АМФ. Кроме ц. АМФ, существуют ц. ГМФ, ц. УМФ, ц. ЦМФ.
Уровень, образовавшийся под действием АЦ, ц. АМФ определяется не только активностью АЦ, но и активностью фосфодиэстераз, которые циклизируют ц. АМФ до АМФ. Кроме ц. АМФ, существуют ц. ГМФ, ц. УМФ, ц. ЦМФ.
 Наибольшее значение имеет ц. ГМФ. Она образуется под действием гуанилатциклазы, расположенной как в наружной мембране, так и внутри клетки, ц. ГМФ единственный фермент, который реагирует на концентрацию Н 2 О 2 и активируется под действием продуктов перекисного окисления.
Наибольшее значение имеет ц. ГМФ. Она образуется под действием гуанилатциклазы, расположенной как в наружной мембране, так и внутри клетки, ц. ГМФ единственный фермент, который реагирует на концентрацию Н 2 О 2 и активируется под действием продуктов перекисного окисления.
 ц. ГМФ оказывает эффекты противоположные ц. АМФ находится в тесном контакте с ионами Ca 2+: высокая концентрация ц. АМФ в возбудимых тканях приводит к высокой [Ca 2+] и стимуляции клетки. И наоборот, резкое уменьшение [Ca 2+] тормозит АЦ и снижает уровень ц. АМФ.
ц. ГМФ оказывает эффекты противоположные ц. АМФ находится в тесном контакте с ионами Ca 2+: высокая концентрация ц. АМФ в возбудимых тканях приводит к высокой [Ca 2+] и стимуляции клетки. И наоборот, резкое уменьшение [Ca 2+] тормозит АЦ и снижает уровень ц. АМФ.
 Аденилатциклазный механизм
Аденилатциклазный механизм
 Некоторые клеточные ответы на действие гормонов, опосредуемое циклическим АМФ Гормон Клетки-мишени Главный ответ Тиротропин Тироциты Синтез и секреция тироксина Кортикотропин Клетки пучковой и сетчатой зоны коры надпочечников Секреция кортизола Лютропин Клетки Лейдига яичников Секреция прогестерона Адреналин Миоциты, гепатоциты Распад гликогена Паратгормон Остеокласты Резорбция кости Адреналин Р-клетки водителя ритма сердца Увеличение частоты сокращений Вазопрессин Эпителиоциты канальцев нефрона почки Реабсорбция воды Адреналин, кортикотропин, глюкагон, тиротропин Адипоциты Расщепление триацилглицеролов
Некоторые клеточные ответы на действие гормонов, опосредуемое циклическим АМФ Гормон Клетки-мишени Главный ответ Тиротропин Тироциты Синтез и секреция тироксина Кортикотропин Клетки пучковой и сетчатой зоны коры надпочечников Секреция кортизола Лютропин Клетки Лейдига яичников Секреция прогестерона Адреналин Миоциты, гепатоциты Распад гликогена Паратгормон Остеокласты Резорбция кости Адреналин Р-клетки водителя ритма сердца Увеличение частоты сокращений Вазопрессин Эпителиоциты канальцев нефрона почки Реабсорбция воды Адреналин, кортикотропин, глюкагон, тиротропин Адипоциты Расщепление триацилглицеролов
 ИФ 3 обеспечивает повышение уровня кальция
ИФ 3 обеспечивает повышение уровня кальция
 Белки, регулируемые Са 2+ и кальмодулином Аденилат циклаза Са 2+/кальмодулин, зависимая протеинкиназа ЦАМФ фосфодиэстераза Са 2+ зависимый Na+ канал ЦАМФ зависимый канал (зрение) Са 2+ канал эндоплазматической сети Киназа легкой цепи миозина Кальциневрин (фосфопротеинфосфатаза 2 В) НАДН киназа ц. ГМФ зависимый Na+, Ca 2+ канал (колбочки) Синтаза оксида азота Са 2+ АТФаза плазм. мембран ИФ-3 -киназа РНК хеликаза (р68) ЦГМФ - зависимая протеинкиназа легких цепей миозина киназа фосфорилазы фосфолипаза А 2
Белки, регулируемые Са 2+ и кальмодулином Аденилат циклаза Са 2+/кальмодулин, зависимая протеинкиназа ЦАМФ фосфодиэстераза Са 2+ зависимый Na+ канал ЦАМФ зависимый канал (зрение) Са 2+ канал эндоплазматической сети Киназа легкой цепи миозина Кальциневрин (фосфопротеинфосфатаза 2 В) НАДН киназа ц. ГМФ зависимый Na+, Ca 2+ канал (колбочки) Синтаза оксида азота Са 2+ АТФаза плазм. мембран ИФ-3 -киназа РНК хеликаза (р68) ЦГМФ - зависимая протеинкиназа легких цепей миозина киназа фосфорилазы фосфолипаза А 2
 Сигнальные роли ДАГ
Сигнальные роли ДАГ
 Если ИФ 3 повышает концентрацию ионов Са 2+ в цитозоле, другой продукт расщепления фосфатидилинозитол -4, 5 - дифосфата – ДАГ – остается в плазматической мембране и вызывает совершенно иные молекулярные эффекты. У него есть две "сигнальные" роли: он может гидролизоваться дальше с образованием арахидоновой кислоты, необходимой для синтеза простагландинов и родственных им медиаторов липидной природы, или способен активировать специфическую протеинкиназу, которая затем фосфорилирует ряд белков с различными функциями в клетке-мишени.
Если ИФ 3 повышает концентрацию ионов Са 2+ в цитозоле, другой продукт расщепления фосфатидилинозитол -4, 5 - дифосфата – ДАГ – остается в плазматической мембране и вызывает совершенно иные молекулярные эффекты. У него есть две "сигнальные" роли: он может гидролизоваться дальше с образованием арахидоновой кислоты, необходимой для синтеза простагландинов и родственных им медиаторов липидной природы, или способен активировать специфическую протеинкиназу, которая затем фосфорилирует ряд белков с различными функциями в клетке-мишени.
 или С–киназой, так как активность") • Фермент, активируемый ДАГ, называется протеинкиназой С (ПКС) или С–киназой, так как активность его зависит от уровня Са 2+ в цитозоле клетки. Полипептидная цепь этого фермента содержит четыре консервативных домена и пять вариабельных областей. Консервативные области включают АТФ-, Са 2+ - , диацилглицерол- и субстратсвязывающий домены. При низком внутриклеточном уровне Са 2+ и отсутствии диацилглицерола протеинкиназа С находится в цитоплазме в неактивном состоянии.
• Фермент, активируемый ДАГ, называется протеинкиназой С (ПКС) или С–киназой, так как активность его зависит от уровня Са 2+ в цитозоле клетки. Полипептидная цепь этого фермента содержит четыре консервативных домена и пять вариабельных областей. Консервативные области включают АТФ-, Са 2+ - , диацилглицерол- и субстратсвязывающий домены. При низком внутриклеточном уровне Са 2+ и отсутствии диацилглицерола протеинкиназа С находится в цитоплазме в неактивном состоянии.
 • Связывание диацилглицерола изменяет конформацию протеинкиназы С, что сопровождается повышением ее сродства к ионам Са 2+ и липидам. Это приводит к связыванию протеинкиназы С с цитоплазматической поверхностью плазматической мембраны и переводу фермента в активное состояние. Активация С -киназы кратковременна, так как через несколько секунд диацилглицерол фосфорилируется до фосфатидной кислоты или расщепляется с высвобождением арахидоновой кислоты.
• Связывание диацилглицерола изменяет конформацию протеинкиназы С, что сопровождается повышением ее сродства к ионам Са 2+ и липидам. Это приводит к связыванию протеинкиназы С с цитоплазматической поверхностью плазматической мембраны и переводу фермента в активное состояние. Активация С -киназы кратковременна, так как через несколько секунд диацилглицерол фосфорилируется до фосфатидной кислоты или расщепляется с высвобождением арахидоновой кислоты.
 • С-киназа, активированная диацилглицеролом и Са 2+, переносит концевую фосфатную группу с АТФ на специфические сериновые или треониновые остатки белков-мишеней, которые в разных клетках различны. Например, во многих животных клетках С-киназа фосфорилирует и тем самым активирует Na+/H+ обменный насос плазматической мембраны, контролирующий внутриклеточный р. Н.
• С-киназа, активированная диацилглицеролом и Са 2+, переносит концевую фосфатную группу с АТФ на специфические сериновые или треониновые остатки белков-мишеней, которые в разных клетках различны. Например, во многих животных клетках С-киназа фосфорилирует и тем самым активирует Na+/H+ обменный насос плазматической мембраны, контролирующий внутриклеточный р. Н.
 • Концентрация С-киназы выше всего в нейронах головного мозга, где, помимо, прочего она фосфорилирует ионные каналы нейронов и, изменяя их проницаемость, может влиять на возбудимость этих клеток. • В некоторых клетках активация С - киназы усиливает транскрипцию определенных генов.
• Концентрация С-киназы выше всего в нейронах головного мозга, где, помимо, прочего она фосфорилирует ионные каналы нейронов и, изменяя их проницаемость, может влиять на возбудимость этих клеток. • В некоторых клетках активация С - киназы усиливает транскрипцию определенных генов.
 Взаимосвязь ц. АМФ и ИФ 3 зависимых путей
Взаимосвязь ц. АМФ и ИФ 3 зависимых путей
 Изоферменты, их природа, биологическая роль, строение ЛДГ. • Изоферменты - это группа родственных ферментов, катализирующих одну и ту же реакцию. Они происходят из одного предшественника за счет дупликациии гена с последующей мутацией образуемых аллелей. Они отличаются между собой:
Изоферменты, их природа, биологическая роль, строение ЛДГ. • Изоферменты - это группа родственных ферментов, катализирующих одну и ту же реакцию. Они происходят из одного предшественника за счет дупликациии гена с последующей мутацией образуемых аллелей. Они отличаются между собой:
 Изоферменты
Изоферменты
 скоростью катализа; 2) направлением катализируемой реакции; 3) условиями протекания реакции; 4) чувствительностью") 1) скоростью катализа; 2) направлением катализируемой реакции; 3) условиями протекания реакции; 4) чувствительностью к регуляторам, факторам среды. (Более или менее устойчивы к ингибиторам); 5) сродством к субстрату; 6) особенностями структуры молекулы, ее ИЭТ, Mr, размерами и зарядом.
1) скоростью катализа; 2) направлением катализируемой реакции; 3) условиями протекания реакции; 4) чувствительностью к регуляторам, факторам среды. (Более или менее устойчивы к ингибиторам); 5) сродством к субстрату; 6) особенностями структуры молекулы, ее ИЭТ, Mr, размерами и зарядом.
 • Тетрамерный фермент, состоящий из двух кодируемых генетически субъединиц А и В может быть представлен пятью изоферментами А 4, АЗВ 1, А 2 В 2, А 1 ВЗ и В 4. Каждый из изоферментов может катализировать одинаковую реакцию, но отличается по своим свойствам, что важно при использовании этих ферментов в разных условиях. Таким путем Природа «экономит» генетический материал, создавая многообразие.
• Тетрамерный фермент, состоящий из двух кодируемых генетически субъединиц А и В может быть представлен пятью изоферментами А 4, АЗВ 1, А 2 В 2, А 1 ВЗ и В 4. Каждый из изоферментов может катализировать одинаковую реакцию, но отличается по своим свойствам, что важно при использовании этих ферментов в разных условиях. Таким путем Природа «экономит» генетический материал, создавая многообразие.
 Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма. Изоферменты обеспечивают межорганную связь, например, в процессе мышечной деятельности. В миокарде и печени существуют различные изоферменты ЛДГ, которые обеспечивают метаболизм лактата:
Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма. Изоферменты обеспечивают межорганную связь, например, в процессе мышечной деятельности. В миокарде и печени существуют различные изоферменты ЛДГ, которые обеспечивают метаболизм лактата:
 ЛДГ 4, и 5 в печени: ПВК лактат ЛДГ 1, и 2 в сердце: лактат ПВК ЛДГ - олигомерный фермент, состоящий из 4 -х субъединиц 2 типов. H (heart) и M (muscle).
ЛДГ 4, и 5 в печени: ПВК лактат ЛДГ 1, и 2 в сердце: лактат ПВК ЛДГ - олигомерный фермент, состоящий из 4 -х субъединиц 2 типов. H (heart) и M (muscle).
 Существует 5 изоферментных форм: HHHH HHHM HHMM HMMM MMMM H 4 H 3 M H 2 M 2 HM 3 M 4 ЛДГ 1, ЛДГ 2, ЛДГ 3, ЛДГ 4, ЛДГ 5. Поскольку H-протомеры несут более выраженный отрицательный заряд, то изофермент H 4 (ЛДГ 1) будет мигрировать при электрофорезе с наибольшей скоростью к аноду. С наименьшей скоростью к аноду будет двигаться М 4.
Существует 5 изоферментных форм: HHHH HHHM HHMM HMMM MMMM H 4 H 3 M H 2 M 2 HM 3 M 4 ЛДГ 1, ЛДГ 2, ЛДГ 3, ЛДГ 4, ЛДГ 5. Поскольку H-протомеры несут более выраженный отрицательный заряд, то изофермент H 4 (ЛДГ 1) будет мигрировать при электрофорезе с наибольшей скоростью к аноду. С наименьшей скоростью к аноду будет двигаться М 4.
 Остальные изоферменты занимают промежуточное положение. Изоферменты ЛДГ локализованы в различных тканях: ЛДГ 1, 2 мозг, аэробные ткани (миокард). ЛДГ 3 лейкозные клетки. ЛДГ 4, 5 анэробные ткани: мышечная, скелетная. Изоферменты появляются на различных этапах онтогенеза и реализуют программу индивидуального развития. Изоферментный профиль меняется в процессе развития. При патологиях имеется существенный изоферментный сдвиг.
Остальные изоферменты занимают промежуточное положение. Изоферменты ЛДГ локализованы в различных тканях: ЛДГ 1, 2 мозг, аэробные ткани (миокард). ЛДГ 3 лейкозные клетки. ЛДГ 4, 5 анэробные ткани: мышечная, скелетная. Изоферменты появляются на различных этапах онтогенеза и реализуют программу индивидуального развития. Изоферментный профиль меняется в процессе развития. При патологиях имеется существенный изоферментный сдвиг.
 Изменение активности ферментов в онтогенезе. Онтогенез человека развивается по определенной генетической программе, которая записана на уровне ткани, всего организма в гипоталамусе. 1. Внутриутробный период. Характеризуется высокой активностью ферментов синтеза белка, липидов, происходит увеличение массы организма. Плод находится в анаэробных условиях и для метаболизма характерно анаэробная направленность. Основной источник энергии - жирные кислоты, поступающие из организма матери; ЖК также выполняют строительную функцию (фосфолипиды мембран).
Изменение активности ферментов в онтогенезе. Онтогенез человека развивается по определенной генетической программе, которая записана на уровне ткани, всего организма в гипоталамусе. 1. Внутриутробный период. Характеризуется высокой активностью ферментов синтеза белка, липидов, происходит увеличение массы организма. Плод находится в анаэробных условиях и для метаболизма характерно анаэробная направленность. Основной источник энергии - жирные кислоты, поступающие из организма матери; ЖК также выполняют строительную функцию (фосфолипиды мембран).
, т. к. ткани плода не способны к") Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода не способны к ГНГ, и идет на развитие ЦНС. 2. Пренатальный период. Характеризуется изменением активности ферментов, происходит подготовка организма к пребыванию в аэробной среде. Изменяется спектр гемоглобина, уменьшается его сродство к кислороду, изменяется активность митохондриальных ферментов
Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода не способны к ГНГ, и идет на развитие ЦНС. 2. Пренатальный период. Характеризуется изменением активности ферментов, происходит подготовка организма к пребыванию в аэробной среде. Изменяется спектр гемоглобина, уменьшается его сродство к кислороду, изменяется активность митохондриальных ферментов
 3. Грудной Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно, но примерно до двух лет основным источником энергии является все же липиды, причиной чего является соматотропин. (Гормон роста). 4. Ранний дошкольный период. С 3 -х до 5 -и лет. В этот период клетки начинают питаться углеводами. Происходит стабилизация обмена и интенсивная миелизация нервных волокон. 5. Школьный и пубертантный период. Обмен веществ модулируется под действием половых гормонов
3. Грудной Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно, но примерно до двух лет основным источником энергии является все же липиды, причиной чего является соматотропин. (Гормон роста). 4. Ранний дошкольный период. С 3 -х до 5 -и лет. В этот период клетки начинают питаться углеводами. Происходит стабилизация обмена и интенсивная миелизация нервных волокон. 5. Школьный и пубертантный период. Обмен веществ модулируется под действием половых гормонов
 6. Зрелый. Происходит стабилизация массы тела, репродуктивного гомеостаза. После 3540 лет основным источником энергии являются опять липиды, что связано с ослаблением чувствительности тканей к Гл и изменение гормонального фона: гиперстресс (увеличивается уровень гормонов) заставляет клетку работать на пределе, т. е. использовать в качестве энергии жиры.
6. Зрелый. Происходит стабилизация массы тела, репродуктивного гомеостаза. После 3540 лет основным источником энергии являются опять липиды, что связано с ослаблением чувствительности тканей к Гл и изменение гормонального фона: гиперстресс (увеличивается уровень гормонов) заставляет клетку работать на пределе, т. е. использовать в качестве энергии жиры.
; Регуляция конститутивными") Активная регуляция ферментов возможна - на геномном уровне ( лактозный оперон); Регуляция конститутивными и индуцибельными ферментами. Синтез первых не зависит от индуктора, т. к. есть определенный уровень фермента, даже при отсутствии индуктора- это базовый уровень. Индуцибельные ферменты это ферменты ЦСМ, цитохром Р-450, бета-ГОМГ-Ко. А-редуктаза. Биосинтез аргиназы увеличивается при повышенном потреблении белков и ускорении ЦСМ. Биосинтез липазы возрастает при голодании, обеспечивая мышцы, в том числе и миокард жирными кислотами.
Активная регуляция ферментов возможна - на геномном уровне ( лактозный оперон); Регуляция конститутивными и индуцибельными ферментами. Синтез первых не зависит от индуктора, т. к. есть определенный уровень фермента, даже при отсутствии индуктора- это базовый уровень. Индуцибельные ферменты это ферменты ЦСМ, цитохром Р-450, бета-ГОМГ-Ко. А-редуктаза. Биосинтез аргиназы увеличивается при повышенном потреблении белков и ускорении ЦСМ. Биосинтез липазы возрастает при голодании, обеспечивая мышцы, в том числе и миокард жирными кислотами.
 Роль ПКС в регуляции транскрипции
Роль ПКС в регуляции транскрипции
 Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию генов. В одном, Скиназа активирует протеинкиназный каскад, приводяций к фосфорилированию митогенактивируемой протеинкиназы (МАП – киназы), которая фосфорилирует и активирует ген-регуляторный белок Elk-1.
Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию генов. В одном, Скиназа активирует протеинкиназный каскад, приводяций к фосфорилированию митогенактивируемой протеинкиназы (МАП – киназы), которая фосфорилирует и активирует ген-регуляторный белок Elk-1.
 и") Elk-1 связан с короткой последовательностью ДНК (обозначаемой serum response element – SRE) и ассоциирован с другим ДНК - связывающим белком (обозначаемым serum response factor – SRF).
Elk-1 связан с короткой последовательностью ДНК (обозначаемой serum response element – SRE) и ассоциирован с другим ДНК - связывающим белком (обозначаемым serum response factor – SRF).
 В другом пути, активация С-киназы приводит к фосфорилированию ингибиторного белка Ik-B, что сопровождается высвобождением из комплекса ген-регуляторного белка NF – k. B, который мигрирует из цитозоля в ядро и активирует транскрипцию соответствующего гена.
В другом пути, активация С-киназы приводит к фосфорилированию ингибиторного белка Ik-B, что сопровождается высвобождением из комплекса ген-регуляторного белка NF – k. B, который мигрирует из цитозоля в ядро и активирует транскрипцию соответствующего гена.
 Существует группа соединений, среди которых наиболее хорошо изучены форболовые эфиры, которые являются мощными активаторами ПКС. Они действуют подобно ДАГ как вторичные посредники, но в отличие от естественного ДАГ, они разрушаются медленно. Постоянно активируя ПКС, эти синтетические вещества вмешиваются в нормальную регуляцию клеточного роста и деления и служат факторами, стимулирующими образование опухолей. .
Существует группа соединений, среди которых наиболее хорошо изучены форболовые эфиры, которые являются мощными активаторами ПКС. Они действуют подобно ДАГ как вторичные посредники, но в отличие от естественного ДАГ, они разрушаются медленно. Постоянно активируя ПКС, эти синтетические вещества вмешиваются в нормальную регуляцию клеточного роста и деления и служат факторами, стимулирующими образование опухолей. .
 Метаболизм ферментов • Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов в тканях поддерживается их постоянным синтезом и распадом. В результате гибели клеток ферменты попадают в крвь, где при участии протеаз или в клетках РЭС подвергаются деградации.
Метаболизм ферментов • Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов в тканях поддерживается их постоянным синтезом и распадом. В результате гибели клеток ферменты попадают в крвь, где при участии протеаз или в клетках РЭС подвергаются деградации.
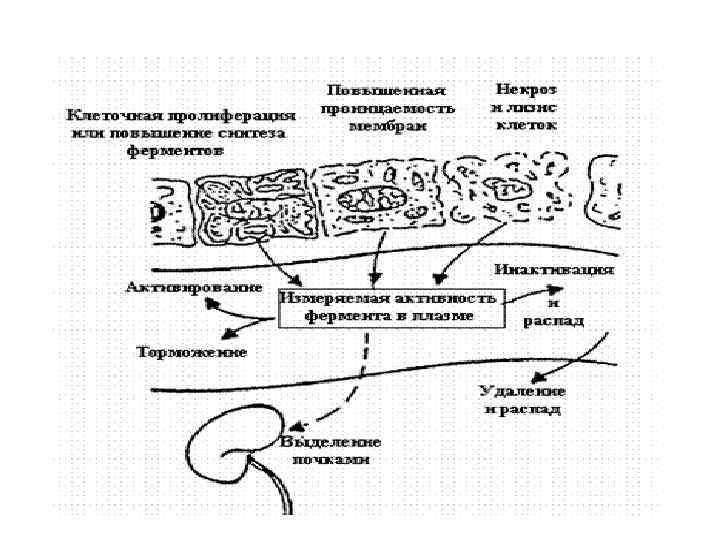
 Имея высокую молекулярную массу, большая часть ферментов не экскретируется с мочой. Ферменты экскретируемые с желчью -ЩФ, ГГТП, 5 - нуклеотидаза, аминотрансферазы.
Имея высокую молекулярную массу, большая часть ферментов не экскретируется с мочой. Ферменты экскретируемые с желчью -ЩФ, ГГТП, 5 - нуклеотидаза, аминотрансферазы.
 Активность фермента Активность - это изменение количества субстрата под влиянием фермента в единицу времени. Под изменением субстрата понимают снижающееся в единицу времени количество субстрата или же увеличивающееся количество продукта.
Активность фермента Активность - это изменение количества субстрата под влиянием фермента в единицу времени. Под изменением субстрата понимают снижающееся в единицу времени количество субстрата или же увеличивающееся количество продукта.
 Понятие "активность фермента" по сути дела идентична понятию "скорость ферментативной" реакции. Ферментативная активность выражается в единицах активности. В связи с существованием различных систем единиц исчисления введена интернациональная (стандартная) единица активности. Она носит символ "U" (unit-единица) и определяется как 1 мкмоль субстрата/мин. В системе СИ в качестве единицы
Понятие "активность фермента" по сути дела идентична понятию "скорость ферментативной" реакции. Ферментативная активность выражается в единицах активности. В связи с существованием различных систем единиц исчисления введена интернациональная (стандартная) единица активности. Она носит символ "U" (unit-единица) и определяется как 1 мкмоль субстрата/мин. В системе СИ в качестве единицы
 В системе СИ в качестве единицы ферментативной активности используют "катал" (kat). Катал определяется как 1 моль/сек. 1 kat = 1 моль/сек.
В системе СИ в качестве единицы ферментативной активности используют "катал" (kat). Катал определяется как 1 моль/сек. 1 kat = 1 моль/сек.
 Размерность её слишком велика, на практике пользуются меньшими кратными значениями, начиная с нанокатала (нкат). Это одна миллиардная катала или 10 -9 кат. В сравнении с международной единицей следующее уравнение 1 U = 16, 67 нкат
Размерность её слишком велика, на практике пользуются меньшими кратными значениями, начиная с нанокатала (нкат). Это одна миллиардная катала или 10 -9 кат. В сравнении с международной единицей следующее уравнение 1 U = 16, 67 нкат
 В практике лабораторий широко пользуются понятием удельная активность. Для этого число cтандартных единиц пересчитывают на какую-либо единицу сравнения. Это может быть мг белка в пробе или объем исследуемой биологической жидкости. Определение активности ферментов широко распространено в любой современной клинической лаборатории.
В практике лабораторий широко пользуются понятием удельная активность. Для этого число cтандартных единиц пересчитывают на какую-либо единицу сравнения. Это может быть мг белка в пробе или объем исследуемой биологической жидкости. Определение активности ферментов широко распространено в любой современной клинической лаборатории.
 При исследовании кинетики реакций используется и такое понятие как молекулярная активность. Она показывает, сколько молекул субстрата в секунду превращаются в продукт 1 молекулой фермента и используется для сравнительной характеристики активности нескольких ферментов.
При исследовании кинетики реакций используется и такое понятие как молекулярная активность. Она показывает, сколько молекул субстрата в секунду превращаются в продукт 1 молекулой фермента и используется для сравнительной характеристики активности нескольких ферментов.
 Пример для вычисления активности фермента Исходные данные: 25. 0 x 10 -3 моль л--1 пептидасубстрата, объем реакционной смеси 2. 5 мл, 0. 50 µг химотрипсина[1] Через 10 мин: 18. 6 x 10 -3 моль л--1 пептида -субстрата, Объем реакционной смеси 2. 5 мл, 0. 50 µг химотрипсина.
Пример для вычисления активности фермента Исходные данные: 25. 0 x 10 -3 моль л--1 пептидасубстрата, объем реакционной смеси 2. 5 мл, 0. 50 µг химотрипсина[1] Через 10 мин: 18. 6 x 10 -3 моль л--1 пептида -субстрата, Объем реакционной смеси 2. 5 мл, 0. 50 µг химотрипсина.
 Изменение активности ферментов у больного ИМ в разные сроки заболевания
Изменение активности ферментов у больного ИМ в разные сроки заболевания
 Номенклатура и классификация ферментов • Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий его положение в классификации: первая цифра характеризует класс фермента, вторая –подкласс и третья подподкласс.
Номенклатура и классификация ферментов • Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий его положение в классификации: первая цифра характеризует класс фермента, вторая –подкласс и третья подподкласс.
 Шифр КФК и ее место в классификации ферментов
Шифр КФК и ее место в классификации ферментов
 Каждый подподкласс представляет собой список ферментов. Порядковый номер фермента в этом списке – четвертая цифра кода. Например- показан шифр креатинфосфокиназы – КФ. 2. 7. 3. 2. Этот фермент катализирует реакцию фосфорилирования креатина. Систематическое название фермента АТФ: креатинфосфотрансфераза. Рабочее название этого фермента креатинкиназа или креатинфофокиназа
Каждый подподкласс представляет собой список ферментов. Порядковый номер фермента в этом списке – четвертая цифра кода. Например- показан шифр креатинфосфокиназы – КФ. 2. 7. 3. 2. Этот фермент катализирует реакцию фосфорилирования креатина. Систематическое название фермента АТФ: креатинфосфотрансфераза. Рабочее название этого фермента креатинкиназа или креатинфофокиназа
 1. 1. 0. 0. Действуют на СН-ОН группы") Оксидоредуктазы (1. 0. 0. 0. ) 1. 1. 0. 0. Действуют на СН-ОН группы доноров 1. 1. 1. 0. НАД+ или НАДФ+ в качестве акцепторов 1. 1. Алкогольдегидрогеназа 1. 14. 0. 0. Действуют на парные доноры при включении в один из них кислорода 1. 14. 15. 0. Один из доноров восстановленный железо -серный белок и включение одного атома кислорода 1. 14. 15. 1. Цитохром Р-450 1. 14. 15. 5. Кортикостерон 18 -монооксигеназа
Оксидоредуктазы (1. 0. 0. 0. ) 1. 1. 0. 0. Действуют на СН-ОН группы доноров 1. 1. 1. 0. НАД+ или НАДФ+ в качестве акцепторов 1. 1. Алкогольдегидрогеназа 1. 14. 0. 0. Действуют на парные доноры при включении в один из них кислорода 1. 14. 15. 0. Один из доноров восстановленный железо -серный белок и включение одного атома кислорода 1. 14. 15. 1. Цитохром Р-450 1. 14. 15. 5. Кортикостерон 18 -монооксигеназа
 2. 1. 0. 0. Переносят одноуглеродные группы 2.") Трансферазы (2. 0. 0. 0. ) 2. 1. 0. 0. Переносят одноуглеродные группы 2. 1. 1. 0. Метилтрансферазы 2. 1. 1. 1. Никотинамид метилтрансфераза 2. 1. 1. 45. Тимидилат синтаза 2. 3. 0. 0. Ацилтрансферазы 2. 3. 1. 6. Холинацетил трансфераза
Трансферазы (2. 0. 0. 0. ) 2. 1. 0. 0. Переносят одноуглеродные группы 2. 1. 1. 0. Метилтрансферазы 2. 1. 1. 1. Никотинамид метилтрансфераза 2. 1. 1. 45. Тимидилат синтаза 2. 3. 0. 0. Ацилтрансферазы 2. 3. 1. 6. Холинацетил трансфераза
 3. 1. 0. 0. Действуют на эфирные связи") Гидролазы (3. 0. 0. 0. ) 3. 1. 0. 0. Действуют на эфирные связи 3. 1. 1. 0. Гидролазы эфиров карбоновых кислот 3. 1. 1. 17. Ацетилхолинэстераза 3. 2. 1. 0. Гликозидгидролазы 3. 2. 1. 1. a-амилаза 3. 2. 1. 2. b-амилаза 3. 4. 0. 0. Действуют на пептидные связи 3. 4. 21. 0. Сериновые протеазы 3. 4. 21. 1. Химотрипсин 3. 4. 21. 4. Трипсин 3. 4. 21. 5. Тромбин
Гидролазы (3. 0. 0. 0. ) 3. 1. 0. 0. Действуют на эфирные связи 3. 1. 1. 0. Гидролазы эфиров карбоновых кислот 3. 1. 1. 17. Ацетилхолинэстераза 3. 2. 1. 0. Гликозидгидролазы 3. 2. 1. 1. a-амилаза 3. 2. 1. 2. b-амилаза 3. 4. 0. 0. Действуют на пептидные связи 3. 4. 21. 0. Сериновые протеазы 3. 4. 21. 1. Химотрипсин 3. 4. 21. 4. Трипсин 3. 4. 21. 5. Тромбин
 4. 1. 0. 0. Углерод-углерод лиазы 4. 1. 1. 0.") Лиазы(4. 0. 0. 0) 4. 1. 0. 0. Углерод-углерод лиазы 4. 1. 1. 0. Карбокси лиазы 4. 1. 1. 1. Пируватдекарбоксилаза 4. 2. 0. 0. Углерод-кислород-лиазы 4. 2. 1. 0. Гидролиазы 4. 2. 1. 11. Енолаза 4. 2. 1. 12. Фосфоглюконатдегидраза
Лиазы(4. 0. 0. 0) 4. 1. 0. 0. Углерод-углерод лиазы 4. 1. 1. 0. Карбокси лиазы 4. 1. 1. 1. Пируватдекарбоксилаза 4. 2. 0. 0. Углерод-кислород-лиазы 4. 2. 1. 0. Гидролиазы 4. 2. 1. 11. Енолаза 4. 2. 1. 12. Фосфоглюконатдегидраза
 6. 1. 0. 0. Образуют С-О связи 6.") Лигазы (6. 0. 0. 0) 6. 1. 0. 0. Образуют С-О связи 6. 1. 1. 0. Образуют молекулы аминоацилт. РНК и родственные им соединения. 6. 1. 1. 1. Тирозил-т. РНК синтаза 6. 5. 0. 0. Образуют фосфоэфирные связи 6. 5. 1. 1. ДНК-лигаза (АТФ -зависимая) 6. 5. 1. 2. ДНК-лигаза (НАД+-зависимая)
Лигазы (6. 0. 0. 0) 6. 1. 0. 0. Образуют С-О связи 6. 1. 1. 0. Образуют молекулы аминоацилт. РНК и родственные им соединения. 6. 1. 1. 1. Тирозил-т. РНК синтаза 6. 5. 0. 0. Образуют фосфоэфирные связи 6. 5. 1. 1. ДНК-лигаза (АТФ -зависимая) 6. 5. 1. 2. ДНК-лигаза (НАД+-зависимая)
 Ферменты 4
Ферменты 4