УМКД_ФР(клетка) (2).ppt
- Количество слайдов: 100



Федеральное государственное образовательное учреждение высшего и профессионального образования «Сибирский федеральный университет» кафедра физиологии растений и биотехнологии ФИЗИОЛОГИЯ РАСТЕНИЙ (иллюстративное сопровождение дисциплины) направление 020200 -биология Красноярск 2007

Лекция 1. Введение, цели и задачи, предмет и методы изучения, история План лекции 1. Физиология растений – наука о жизнедеятельности растительного организма. 2. История становления физиологии растений как науки. 3. Предмет, цели и задачи курса. 4. Методы исследования. 5. Место зеленого растения в экономике природы. 6. Население Земли и пищевые ресурсы 1. 1 Физиология растений

Физиология растений – наука о жизнедеятельности растительного организма fizrast. ru/skachat. html Физиология растений. Н. Д. растений. Н. И. Алехина, Ю. В. Якушкина Е. Ю. Балнокин. Под ред. Бахтенко. И. П. Ермакова. Физиология растений - С. С. Медведев. 2003 Основы биохимии растений - Филипцова Физиология Г. Г. , Смолич И. И. растений – растений - С. С. Кузнецов В. В. , Дмитриева Г. А. Медведев. 2013 1. 4 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история

Физиология растений – наука о жизнедеятельности растительного организма Зарубежные учебные издания в областях Физиологии растений 1. 5 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история

Физиология растений – наука о жизнедеятельности растительного организма Центральный журнал Материалы Международной конференции 1. 6 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история

Физиология растений – наука о жизнедеятельности растительного организма Скандинавские страны Япония Россия США Австралия Индия Малайзия Израиль Эмблемы Обществ физиологов растений и смежных областей 1. 2 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история

Место зеленого растения в экономике природы. Место растений в современной систематике (Grzebyk, al. , J. Phycol. 39, 259– 267 (2003) 1. 7 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история

Место зеленого растения в экономике природы. СО 2 + 2 Н 2 О + h СН 2 О + О 2 С 6 Н 12 О 6 + О 2 6 СО 2 + 6 Н 2 О + 686 ккал Круговорот углерода 1. 8 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история
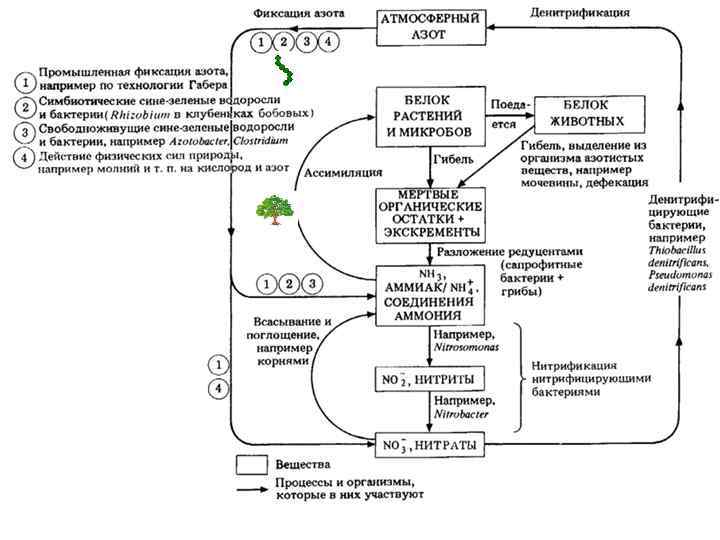
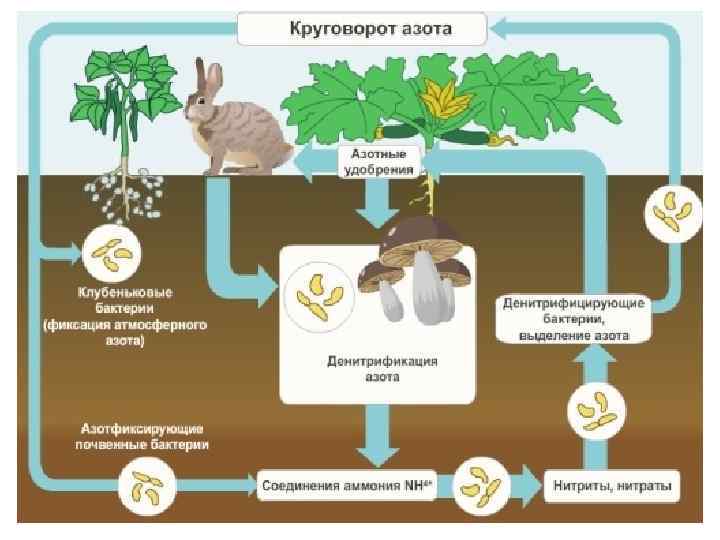

Место зеленого растения в экономике природы. 1. 10 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история

Место зеленого растения в экономике природы. Фитопланктон и растения путем фотосинтеза создают органические вещества, которые служат питанием для животных. В естественных экосистемах около 90% растительной органики потребляют микроорганизмы с размерами тела менее 1 мм. Насекомые и другие мелкие животные с размерами до 1 см используют менее 10% органической продукции, а на долю крупных животных приходится примерно 1% органики. В норме в этот 1% должен укладываться и человек. Однако человечество существенно превысило естественный лимит, и с учетом земледелия, скотоводства и лесозаготовок доля человека в потреблении органики составляет около 10% (пунктирная линия). 1. 11 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история
 предположил, что")
Население Земли и пищевые ресурсы Томас Роберт Мальтус (1766 - 1834) предположил, что люди размножаются быстрее, чем растут средства существования, и что если рост населения ничем не сдерживается, то каждые 25 -30 лет население будет удваиваться. 1. 12 Лекция 1. Введение, цели и задачи, предмет и методы изучения, история

Лекция 2. Физиология растительной клетки. Клетка как осмотическая система. План лекции 1. Общая схема организации растительной клетки. Функции органоидов. 2. Методы исследования растительных клеток. 3. Основные закономерности поглощения воды клеткой. 4. Химический потенциал воды и водный потенциал клетки. Растительная клетка - осмотическая система. 2. 1 Физиология растений

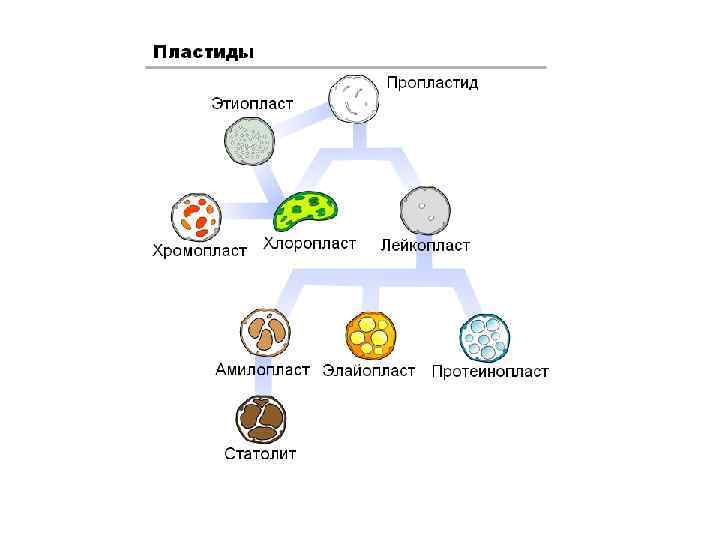
Общая схема организации растительной клетки. Функции органоидов. Строение растительной клетки 2. 2 Лекция 2. Физиология растительной клетки. Клетка как осмотическая система.

Нобел, 1970 Физиология растительной клетки.
 и плазмолиза (справа)")
Протопласты растительных клеток Клетки растений в состоянии тургора (слева) и плазмолиза (справа)

Quick look: A vacuole is a membrane-enclosed fluid filled sac found in the cells of plants including fungi. Vacuoles can be large organelles occupying between 30% and 90% of a cell by volume. Vacuoles appear to have three main functions, they: contribute to the rigidity of the plant using water to develop hydrostatic pressure store nutrient and non-nutrient chemicals break down complex molecules. Вакуоль представляет собой изолированный мембранной, заполненный жидкостью мешочек. Вакуоли найдены в клетках растений, а также в клетках грибов. Вакуоли занимают от 30% до 90% объема клетки. Три основные функции вакуолей: Необходимы для создания гидростатического давления (тургора) в клетки; вместилище питательных и неорганических химических веществ; Разрушение сложных молекул.
 водный")
Содержимое вакуоли — клеточный сок — представляет собой слабокислый (p. H 2 -5) водный раствор различных органических и неорганических веществ. По химическому составу и консистенции клеточный сок существенно отличается от протопласта. Эти различия связаны с избирательной проницаемостью тонопласта, выполняющего барьерную функцию. Большинство органических веществ, содержащихся в клеточном соке, относится к группе эргастических продуктов метаболизма протопласта. В зависимости от потребностей клетки они могут накапливаться в вакуоли в значительных количествах либо полностью исчезать. Наиболее обычны различные углеводы, играющие роль запасных энергетических веществ, а также органические кислоты. Вакуоли семян нередко содержат и белки- протеины. Растительные вакуоли часто служат местом концентрации разнообразных вторичных метаболитов — полифенольиых соединений: флавоноидов, антоцианов, таннидов и азотсодержащих веществ — алкалоидов, В клеточном соке растворены также многие неорганические соединения. Иногда вакуоли разрушают токсичные или ненужные клетке вещества. Обычно это выполняется специальными небольшими вакуолями, содержащими соответствующие ферменты. Такие вакуоли получили название лизосомных.
 storage. Passage")
Plant vacuoles serve as the primary intracellular compartments for inorganic phosphate (Pi) storage. Passage of Pi across vacuolar membranes plays a critical role in buffering the cytoplasmic Pi level against fluctuations of external Pi and metabolic activities. Here we demonstrate that the SPX- MFS proteins, designated as PHOSPHATE TRANSPORTER 5 family (PHT 5), also named Vacuolar Phosphate Transporter (VPT). Our findings show that a group of Pi transporters in vacuolar membranes regulate cytoplasmic Pi homeostasis and are required for fitness and plant growth.

Включения — это компоненты клетки, представляющие собой отложения веществ, временно выведенных из обмена, или конечные его продукты. Большинство включений видимы в световой микроскоп и располагаются либо в гиалоплазме и органоидах, либо в вакуоли. Существуют жидкие и твердые включения. К образованию включений ведет избыточное накопление веществ. Очень часто в виде включений откладываются запасные питательные вещества. Главнейшее и наиболее распространенное из них — полисахарид крахмал. Крахмал злаков, клубней картофеля, ряда тропических растений — важнейший источник углеводов в рационе человека. Первичный ассимиляционный крахмал образуется только в хлоропластах. Ночью, когда фотосинтез прекращается, ассимиляционный крахмал ферментативно гидролизуется до сахаров и транспортируется в другие части растения.
, картофеля (2), молочая (3), герани (4),")
Различные типы крахмальных зерен: у овса (1), картофеля (2), молочая (3), герани (4), фасоли (5), кукурузы (6) и пшеницы (7) Сложные алейроновые зерна в семенах клещевины: 1 — зерно, 2 — оболочка, 3 — кристаллоид, 4 — глобоид

Кристаллы щавелевокислого кальция в вакуолях растительных клеток: 1 - крахмальные зерна, 2 - друзы кристаллов, 3 - ядро клетки, 4 - цитоплазма.


Электронная микрофотография, на которой видны целлюлозные волокна в отдельных слоях клеточной стенки зеленой морской водоросли Chaetomorpha melagonium. Толщина целлюлозных микрофибрилл составляет 20 нм. Для получения контрастного изображения произведено напыление сплавом платины с золотом

Microtubule railroad. Microtubules that lie beneath the plant cell's plasma membrane serve as tracks for membrane-associated cellulose synthases (rosette structures) to travel along. As it moves, the synthase deposits a cellulose microfibril into the adjacent cell wall. It is not yet known if there is a direct link between cellulose synthase and microtubule. Science 9 June 2006: Vol. 312 no. 5779 pp. 1482 - 1483 DOI: 10. 1126/science. 1128903 PERSPECTIVE PLANT SCIENCEMicrotubules Make Tracks for Cellulose Clive Lloyd
 в клетках эпидермиса и мезофилла в листьях")
Модель, демонмтрирующая последовательность дестабилизации сети микротрубочек (МТ) в клетках эпидермиса и мезофилла в листьях арабидопсиса. Процессы сборки и разборки микротрубочек (микрофиламентов) (MTs) – неотъемлемое свойство всех клеток. Несмотря на то, что процесс «старения» листьев хорошо документирован, роль цитоскелета (МТ) в этом явлении остается неизвестной. Процесс старения листьев у Arabidopsis thaliana вызывали естественным путем или искусственно, используя затенение. МТ исследовали в клетках эпидермиса и клетках мезофилла. Зарегистрировали разрушение сети МТ кроме замыкающих клеток устьиц.
")
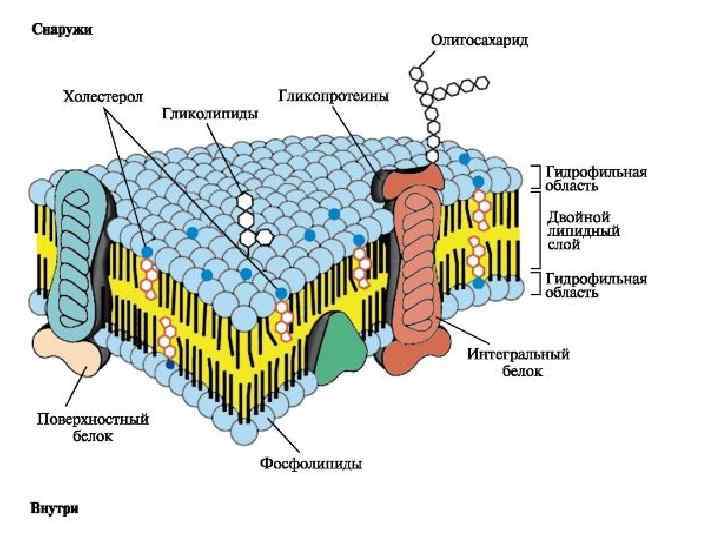
Мембранная система растительной клетки (L. A. Staehelin, 1997)
")
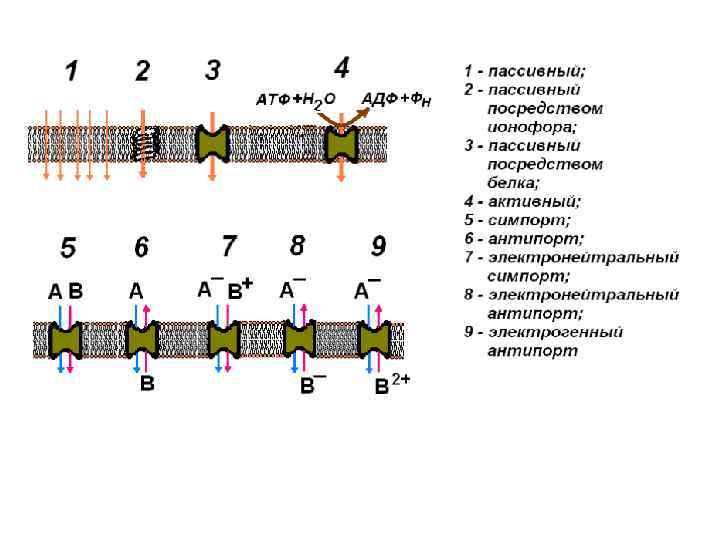
Типы транспорта веществ через клеточные мембраны растений (самостоятельно)
 растительной клетки (С. С. Медведев, 2004)")
Примеры ионного транспорта через плазмалемму (ПЛ) растительной клетки (С. С. Медведев, 2004)
 отвечают на внеклеточные стимулы (митогены) и")
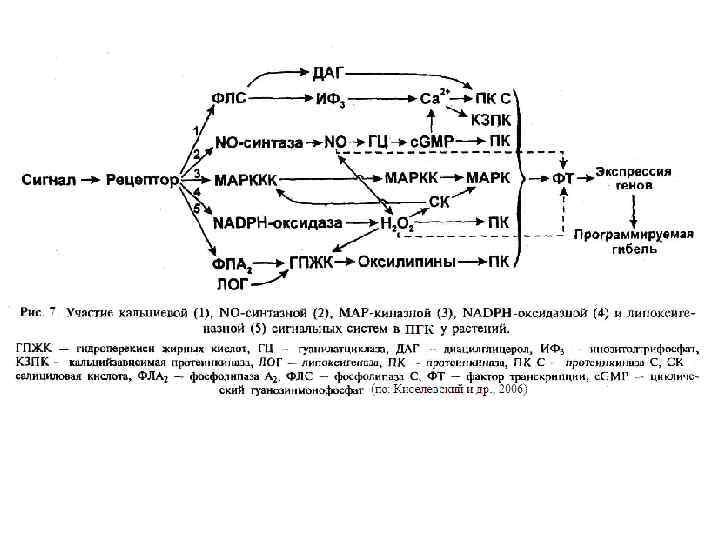
Митоген-активируемые киназы МАРK (КФ 2. 7. 11. 24) отвечают на внеклеточные стимулы (митогены) и регулируют многие клеточные процессы (экспрессию генов, деление, дифференцировку и апоптоз). МАРК вовлечены в работу многих неядерных белков — продуктов онкогенов. Внеклеточные стимулы ведут к активации МАРК через сигнальный каскад, который состоит из МАРК, МАРКК (МАР 2 К) и МАРККК (МАР 3 К). МАР 3 К активируется внеклеточными стимулами и фосфорилирует МАР 2 К, затем МАР 2 К фосфорилированием активирует МАРК. Такой сигнальный МАРK-каскад консервативен для эукариот от дрожжей до млекопитающих.

MAP-киназный каскад Три семейства, киназ MAPK это серин- треонин протеинкиназы. Они в конечном счете меняют спектр транскрипции в ответ на внешний стимул. Достигнуть этого изменения MAPK может тремя разными способами: 1. MAPK может сама транслоцироваться в ядро и там фосфорилировать факторы транскрипции. Так фосфорилируются известные факторы белок с-Myc и белок Elk-1. 2. 2. МАРК может фосфорилировать фактор транскрипции в цитоплазме. После этого фактор проникает в ядро и там активирует транскрипцию определенных генов. 3. МАРК может фосфорилировать ингибитор фактора транскрипции с которым этот фактор связан. Когда фактор связан он неактивен. При фосфорилировании ингибитора фактор освобождается и таким образом активируется. Вот так в самых общих чертах выглядит путь передачи сигнала от ростового фактора. Каскад трех протеинкиназ очень универсально используется в различных цепях передачи сигнала и высоко консервативен в эволюции [ Canman ea 1996 ]. Важно также представлять себе что сигнал, приходящий к поверхности клеток, внутри клетки амплифицируется благодаря существованию каскада. Даже очень слабые сигналы благодаря этому оказываются достаточными, чтобы клетка на них реагировала. Повышение активности МАР-киназ или MAPK происходит под действием РКС [ Azzi A. ea, 1992 ], находящихся "downstream"от c-Raf (онкобелок Raf действует непосредственно "downstream" от Ras и участвуюет в передаче сигнала на ген c-fos [ Angel P. and Karin M. , 1991 , Hunter T. , 1991 ) [ Davis R. J. , 1993 ]. Так как МАР-киназа при этом фосфорилируется одновременно по тирозину и серину, что не может осуществляться РКС, то между PKC и МАР-киназой существуют промежуточные звенья - киназы МАР-киназы ( МАРКК ) с двойной специфичностью (к тирозину и серину) и активирующие ее киназы МАР-киназы ( МАРККК ) [ Davis R. J. , 1993 , Gomez N. ea, 1992 ] (рис. 13) Некоторые изоформы PKC могут выступать в роли MAP киназ. В настоящее время каскад реакций MAP-киназ разделен на две подгруппы, одна из них контролируется протеинкиназным каскадом, передающим митогенные сигналы ( ERK ), а вторая связана с протеинкиназным каскадом, активируемым стрессом ( SAPK ), под контролем которого находятся ингибирование клеточного роста и воспалительные реакции. Роль МАРККК играет хорошо известный белок RAF.
 лауреат Нобелевской премии по медицине 2003")
Аквапорины Peter Agre (профессор биохимии университет Дж. Хопкинса) лауреат Нобелевской премии по медицине 2003 года. Начиная с 1991 года разрабатывает универсальные для всех живых организмов мембранные каналы, которые участвуют в транспорте воды.

В настоящее время существует более 200 различных аквапоринов. В растениях , аквапорины сгруппированы в пять подсемейств: Характерные белки плазматической мембраны (Plasma membrane Intrinsic Protein – PIP) Характерные белки тонопласта, Tonoplast Intrinsic Protein – TIP Характерные нодулин-26 -подобные белки, Nodulin-26 like Intrinsic Protein (NIP) – кроме воды переносят катионы и анионы. Малые основные внутренние белки, Small basic Intrinsic Protein (SIP) Внутренний Белок X - ( XIP )

Схема образования поры для транспорта воды. Активация аквапоры происходит за счет объединения двух петель: В и Е. Структура поры названа «hourgllass» песочные часы.
. 1 -6 — шесть транс")
Структурная модель аквапорина растений (С. С. Медведев, 2004). 1 -6 — шесть транс мембранных α- спиралей, НВ и НЕ —две короткие α - спирали. Лекция 3. Водный режим растений. Функция и формы воды в растениях. Поглощение воды растением. 3. 4

In situ локализация транскриптов аквапорина Os. PIP 2; 7 у 4 - недельных ростков риса (sense probe (A), or antisense probe (B–E)). A и B поперечный срез листа. C – продольный срез кончика корня, включая меристематическую зону, зону растяжения и корневой чехлик. D и E поперечный сред корня. а – клетки мезофилла; b – эпидермис и экзодерма; c - эндодерма; d - эпидермис; e - экзодерма; f – корневой волосок.

Нобел, 1970 Физиология растительной клетки.

Зависимость «натяжение» – «деформация» для клеточной стенки Обратимые упругие свойства материалов, в том числе и клеточных стенок описывает модуль Юнга (выражается в единицах давления): Модуль Юнга для чистой целлюлозы ~ 105 кг/см 2. Это 5% модуля Юнга стали. Для клеточной стенки нитчатой водоросли Nitella его величина 0, 07× 105 кг/см 2. Тургорное давление (Р), существующее внутри живой растительной клетки, приводит к деформации клетки (изменение линейных размеров тканей растений в зависимости от Р можно увидеть в эксперименте с клубнем картофеля, цветоносом одуванчика, работы устьиц и др. )

r = 0. 5 mm Продольное натяжение r = 5 мкм P= 6 бар (1 бар= 0, 987 атм) σL= r P/ 2 r σL= 300 бар; σT= 600 бар М. Ю. = 7000 кг/см 2 Тангенциальное Относительное удлинение: натяжение σT= r P/ r L/L = 306/7000 = 0. 044 или 4, 4% Относительное увеличение диаметра ? ?

Свойства воды В кристаллическом состоянии все молекулы воды связаны водородными связями. При плавлении (? ) разрушается около 15% всех в. с. При 25 С неразрушенными остаются 80% в. с. Испарение воды при этой температуре требует 10, 5 ккал/моль (уникальное свойство воды). Потеря тепла при транспирации – главный механизм регуляции температуры у растения (? )

Поверхностное натяжение и капиллярное поднятие воды Когезия – притяжение между молекулами воды в жидкой фазе; Адгезия – притяжение между жидкой водой и твердой фазой. К<А Р (сила тяжести) = f(σ ) (σ -поверхностное натяжение). r 2 h g=2 r σ cos h= 2 σ cos /r g В капилляре при хорошем смачивании 0, σ = 72, 8 дин/см, = 0, 988 г/см 3, g=980 см/с2 : h=0. 149/r Радиус сосуда ксилемы 20 мкм, высота капиллярного поднятия воды 0, 75 м.
 диффузии Диффузия эффективна на короткое расстояние, но не")
Расчет времени (tc = ½) диффузии Диффузия эффективна на короткое расстояние, но не обеспечивает транспорт на большое расстояние. Первый закон Фика: По мере диффузии вещества от точки старта концентрационный градиент Δcs уменьшается, а диффузионный поток замедляется. Время, необходимое для перемещение в точку пространства, в которой концентрация составит ½ от максимальной (в точке старта) (tc = ½) определяют из уравнения:
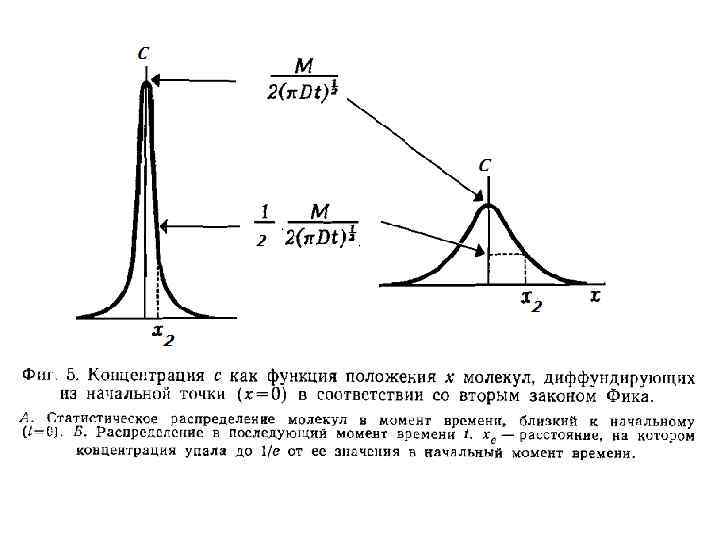
, Б в пределах сосуда ксилемы")
Численные примеры: А – в пределах клетки (50 мкм), Б в пределах сосуда ксилемы (1 м) А Б

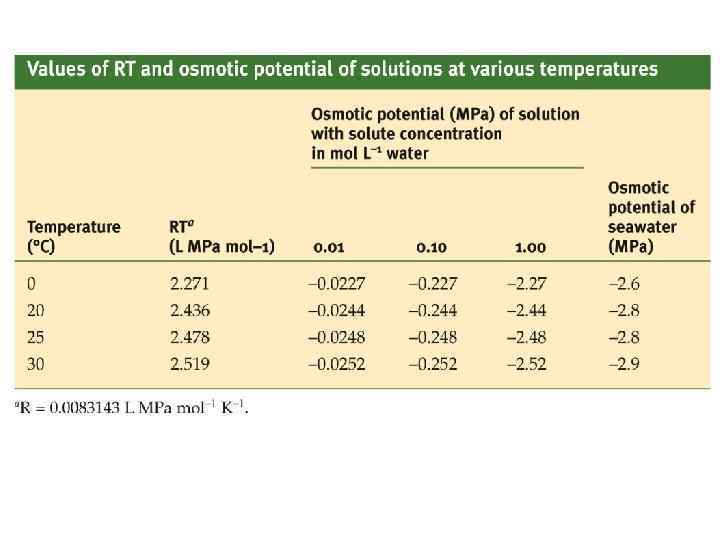
Студенты, изучающие компоненты водного потенциала, сталкиваются с различными обозначениями для одних и тех же величин. Для этого достаточно сравнить два выражения: P имеет то же значение, что и Ψp. Это гидростатическое давление раствора, которое сможет быть как положительным в паренхимной клетке и отрицательным в клетке ксилемы. Символ называют осмотическим давлением и это отрицательное значение для Ψs. Осмотическое давление с точки зрения физхимика, зоолога и др. это влияние растворенных веществ на свободную энергию воды. Ψs = – Термин осмотический потенциал некорректен.

Химический потенциал воды Химический потенциал - один из видов потенциальной энергии, обладает свойством аддитивности. Активность воды и осмотическое давление Уравнение В. Пфеффера, 1877

Матричный потенциал Водный потенциал – дефицит давления диффузии или сосущая сила. Р – гидрастотическое давление или тургорое давление. - осмотическое давление

Соотношение Бойля - Вант-Гоффа Объем тела, окруженного полупроницаемой мембраной, меняется в зависимости от осмотического давления окружающего раствора. Изменение объема тела в ответ на изменение осмотического давления описывает соотношение Бойля – Вант-Гоффа: Где 0 – осмотическое давление снаружи V – объем b – неактивный объем nj – число молей в активном объеме j – осмотический коэффициент n – кажущееся число осмотически-активных веществ.

Measuring Water Potential Plant scientists have expended considerable effort in devising accurate and reliable methods for evaluating the water status of a plant. Four instruments that have been used extensively to measure Ψw , Ψs , and Ψp are described here: psychrometer, pressure chamber, cryoscopic osmometer, and pressure probe. Psychrometer (Ψw measurement) Psychrometry (the prefix "psychro-" comes from the Greek word psychein, "to cool") is based on the fact that the vapor pressure of water is lowered as its water potential is reduced. Psychrometers measure the water vapor pressure of a solution or plant sample, on the basis of the principle that evaporation of water from a surface cools the surface.

One psychrometric technique, known as isopiestic psychrometry, has been used extensively by John Boyer and coworkers (Boyer and Knipling 1965) and is illustrated in Web Figure 3. 6. A. Investigators make a measurement by placing a piece of tissue sealed inside a small chamber that contains a temperature sensor (in this case, a thermocouple) in contact with a small droplet of a standard solution of known solute concentration (known Ψs and thus known Ψw). If the tissue has a lower water potential than that of the droplet, water evaporates from the droplet, diffuses through the air, and is absorbed by the tissue. This slight evaporation of water cools the drop. The larger the difference in water potential between the tissue and the droplet, the higher the rate of water transfer and hence the cooler the droplet. If the standard solution has a lower water potential than that of the sample to be measured, water will diffuse from the tissue to the droplet, causing warming of the droplet. Measuring the change in temperature of the droplet for several s olutions of known Ψw makes it possible to calculate the water potential of a solution for which the net movement of water between the droplet and the tissue would be zero (Web Figure 3. 6. B) signifying that the droplet and the tissue have the same water potential.
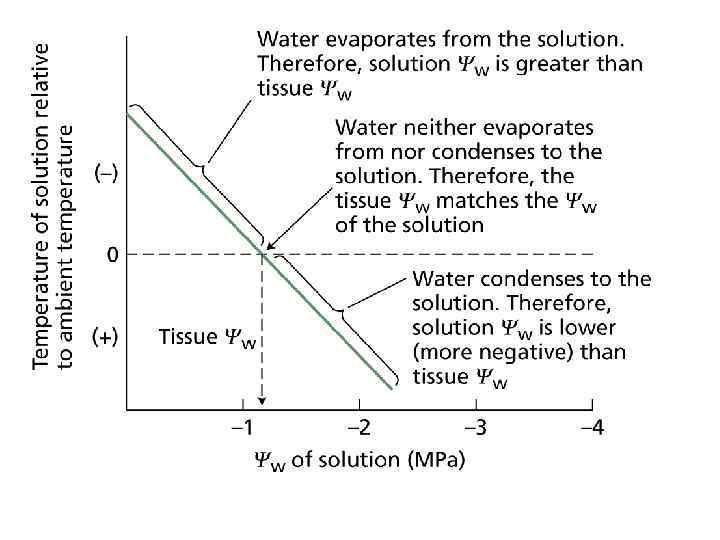

Psychrometers can be used to measure the water potentials of both excised and intact plant tissue. Moreover, the method can be used to measure the Ψs of solutions. This can be particularly useful with plant tissues. For example, the Ψw of a tissue is measured with a psychrometer, and then the tissue is crushed and the Ψs value of the expressed cell sap is measured with the same instrument. By combining the two measurements, researchers can estimate the turgor pressure that existed in the cells before the tissue was crushed (Ψp = Ψw – Ψs). A major difficulty with this approach is the extreme sensitivity of the measurement to temperature fluctuations. For example, a change in temperature of 0. 01°C corresponds to a change in water potential of about 0. 1 MPa. Thus, psychrometers must be operated under constant temperature conditions. For this reason, the method is used primarily in laboratory settings. There are many variations in psychrometric technique; interested readers should consult Brown and Van Haveren 1972, Slavik 1974, and Boyer 1995.
 A relatively quick method for estimating the water potential of")
Pressure chamber (Ψw measurement) A relatively quick method for estimating the water potential of large pieces of tissues, such as leaves and small shoots, is by use of the pressure chamber. This method was pioneered by Henry Dixon at Trinity College, Dublin, at the beginning of the twentieth century, but it did not come into widespread use until P. Scholander and coworkers at the Scripps Institution of Oceanography improved the instrument design and showed its practical use (Scholander et al. 1965). In this technique, the organ to be measured is excised from the plant and is partly sealed in a pressure chamber (Web Figure 3. 6. C). Before excision, the water column in the xylem is under tension. When the water column is broken by excision of the organ (i. e. , its tension is relieved allowing its Ψp to rise to zero), water is pulled rapidly from the xylem into the surrounding living cells by osmosis. The cut surface consequently appears dull and dry. To make a measurement, the investigator pressurizes the chamber with compressed gas until the distribution of water between the living cells and the xylem conduits is returned to its initial, pre-excision, state. This can be detected visually by observing when the water returns to the open ends of the xylem conduits that can be seen in the cut surface. The pressure needed to bring the water back to its initial distribution is called the balance pressure and is readily detected by the change in the appearance of the cut surface, which becomes wet and shiny when this pressure is attained.

The pressure chamber method for measuring plant water potential. The diagram at left shows a shoot sealed into a chamber, which may be pressurized with compressed gas. The diagrams at right show the state of the water columns within the xylem at three points in time: (A) The xylem is uncut and under a negative pressure, or tension. (B) The shoot is cut, causing the water to pull back into the tissue, away from the cut surface, in response to the tension in the xylem. (C) The chamber is pressurized, bringing the xylem sap back to the cut surface. (Click image to enlarge. )

The pressure chamber is often described as a tool to measure the tension in the xylem. However, this is only strictly true for measurements made on a non-transpiring leaf or shoot (for example, one that has been previously enclosed in a plastic bag). When there is no transpiration, the water potential of the leaf cells and the water potential in the xylem will come into equilibrium. The balancing pressure measured on such a non-transpiring shoot is equal in magnitude but opposite in sign to the pressure in the xylem (Ψp). Because the water potential of our non-transpiring leaf is equal to the water potential of the xylem, one can calculate the water potential of the leaf by adding together Ψp and Ψs of the xylem, provided one collects a sample of xylem sap for determination of Ψs. Luckily Ψs of the xylem is usually small (> – 0. 1 MPa) compared to typical midday tensions in the xylem ( Ψp of – 1 to – 2 MPa). Thus, correction for the Ψs of the xylem sap is frequently omitted. Balancing pressure measurements of transpiring leaves are more difficult to interpret. The fact that water is flowing from the xylem to the leaf means that differences in water potential must exist. When the transpiring leaf or shoot is cut off, the tension in the xylem is instantly relieved and water is drawn into the leaf cells until the water potentials of the xylem and the leaf cells come into equilibrium. Because the total volume of the leaf cells is much larger than the volume of sap in the xylem, this equilibrium water potential will be heavily weighted towards that of the leaf. Thus, any measurement of the balancing pressure on such a leaf or shoot will result in a value that is approximately the water potential of the leaf, rather than the tension of the xylem. (To be exact, one would have to add the Ψs of the xylem sap to the negative of the balancing pressure to get the leaf water potential. ) One can explore the differences between the water potential of the xylem and the water potential of a transpiring leaf by comparing balancing pressures measured on covered (i. e. , non-transpiring) versus uncovered (transpiring) leaves. Pressure chamber measurements provide a quick and accurate way of measuring leaf water potential. Because the pressure chamber method does not require delicate instrumentation or temperature control, it has been used extensively under field conditions (Tyree and Hammel 1972). For a more complete description of theory and operation of the pressure chamber see Boyer, 1995.
 The cryoscopic osmometer measures the osmotic potential of a solution")
Cryoscopic osmometer (Ψs measurement) The cryoscopic osmometer measures the osmotic potential of a solution by measuring its freezing point. Solutions have colligative properties that collectively depend on the number of dissolved particles and not on the nature of the solute. For example, solutes reduce the vapor pressure of a solution, raise its boiling point, and lower its freezing point. The specific nature of the solute does not matter. One of the colligative properties of solutions is the decrease in the freezing point as the solute concentration increases. For example, a solution containing 1 mol of solutes per kilogram of water has a freezing point of – 1. 86°C, compared with 0°C for pure water. Various instruments can be used to measure the freezing-point depression of solutions (for two examples, see Prager and Bowman 1963, and Bearce and Kohl 1970). With a cryoscopic osmometer, solution samples as small as 1 nanoliter (10 – 9 L) are placed in an oil medium located on the temperature-controlled stage of a microscope (Web Figure 3. 6. D). The very small sample size allows sap from single cells to be measured and permits rapid thermal equilibration with the stage. To prevent evaporation, the investigator suspends the samples in oil-filled wells in a silver plate (silver has high thermal conductivity). The temperature of the stage is rapidly decreased to about – 30° C, which causes the sample to freeze. The temperature is then raised very slowly, and the melting process in the sample is observed through the microscope. When the last ice crystal in the sample melts, the temperature of the stage is recorded (note that the melting and freezing points are the same). It is straightforward to calculate the solute concentration from the freezing-point depression; and from the solute concentration (cs), Ψs is calculated as –RTcs. This technique has been used to measure droplets extracted from single cells (Malone and Tomos 1992).

A cryoscopic osmometer measures the concentration of total dissolved solutes by measuring the freezing- point depression of a solution. (A) Very small liquid samples are loaded onto the temperature- controlled stage of a microscope. (B) When the temperature is quickly reduced, the samples supercool and freeze. (C) Slowly warming the stage causes the samples to thaw. The temperature at which the last ice crystal melts provides a measure of the melting point of the sample. (Click image to enlarge. )
 If a cell were as large as a watermelon or")
Pressure probe (Ψp measurement) If a cell were as large as a watermelon or even a grape, measuring its hydrostatic pressure would be a relatively easy task. Because of the small size of plant cells, however, the development of methods for direct measurement of turgor pressure has been slow. Using a micromanometer, Paul Green at the University of Pennsylvania developed one of the first direct methods for measuring turgor pressure in plant cells (Green and Stanton 1967). In this technique, an air-filled glass tube sealed at one end is inserted into a cell (Web Figure 3. 6. E). The high pressure in the cell compresses the trapped gas, and from the change in volume one can readily calculate the pressure of the cell from the ideal gas law (pressure × volume = constant). This method works only for cells of relatively large volume, such as the giant cell of the filamentous green alga Nitella. For smaller cells, the loss of cell sap into the glass tube is sufficient to deflate the cell and this yields artifactually low pressures.

Use of the micromanometer, a pressure probe, to measure cell turgor pressure. Nitella cells (which are particularly large—about 100 mm in diameter and many centimeters long) were used for these measurements. (After Green 1968. )

For higher plant cells, which are several orders of magnitude smaller in volume than Nitella, a more sophisticated device, the pressure probe , was developed by Ernest Steudle, Ulrich Zimmermann, and their colleagues in Germany (Husken et al. 1978). This instrument is similar to a miniature syringe (Web Figure 3. 6. F). A glass microcapillary tube is pulled to a fine point and is inserted into a cell. The microcapillary is filled with silicone oil, a relatively incompressible fluid that can be readily distinguished from cell sap under a microscope. When the tip of the microcapillary is first inserted into the cell, cell sap begins to flow into the capillary because of the initial low pressure of that region. Investigators can observe such movement of sap under the microscope and counteract it by pushing on the plunger of the device, thus building up a pressure. In such fashion the boundary between the oil and the cell sap can be pushed back to the tip of the microcapillary. When the boundary is returned to the tip and is held in a constant position, the initial volume of the cell is restored and the pressure inside the cell is exactly balanced by the pressure in the capillary. This pressure is measured by a pressure sensor in the device. Thus the hydrostatic pressure of individual cells may be measured directly.
. The primary advantage of this")
Diagram of the simplest pressure probe (not to scale). The primary advantage of this method over the one shown in Web Figure 3. 6. E is that cell volume is minimally disturbed. Minimal disturbance is of great importance for the tiny cells that are typical of higher plants, in which loss of even a few picoliters (10– 12 L) of fluid can substantially reduce turgor pressure. (Click image to enlarge. )

This method has been used to measure Ψp and other parameters of water relations in cells of both excised and intact tissues of a variety of plant species (Steudle 1993). The primary limitation of this method is that some cells are too small to measure. Furthermore, some cells tend to leak after being stabbed with the capillary, and others plug up the tip of the capillary, thereby preventing valid measurements. The pressure probe has also been adapted to measure positive and negative values of Ψpin the xylem (Heydt and Steudle 1991). However, technical problems with cavitation (see textbook Chapter 4) limit the measurement of negative Ψp by this technique.

The Matric Potential In discussions of soils, seeds, and cell walls, one often finds reference to yet another component of Ψw: the matric potential (Ψm). The matric potential is used to account for the reduction in free energy of water when it exists as a thin surface layer, one or two molecules thick, adsorbed onto the surface of relatively dry soil particles, cell walls, and other materials. The matric potential does not represent a new force acting on water, because the effect of surface interactions can theoretically be accounted for by an effect on Ψs and Ψp (see Passioura 1980; Nobel 1999). In dry materials, however, this surface interaction effect often cannot easily be separated into Ψp and Ψs components in dry materials, so they are frequently bulked together and designated as the matric potential. It is generally not valid to add Ψm to independent measurements of Ψs and Ψp to arrive at a total water potential. This is particularly true for water inside hydrated cells and cell walls, where matric effects are either negligible or they are accounted for by a reduction in Ψp. For instance, the negative pressure in water held by cell wall microcapillaries at the evaporative surfaces of leaves, discussed on p. 75 of the textbook, is sometimes described as a wall matric potential. Care is needed to avoid inconsistencies when accounting for this physical effect in definitions of Ψp, Ψs, and Ψm (Passioura 1980).

Wilting and Plasmolysis is the separation of plant cell cytoplasm from the cell wall as a result of water loss. It is unlikely to occur in nature, except in severe conditions. Plasmolysis is induced in the laboratory by immersing a plant cell in a strongly saline or sugary solution, so that water is lost by osmosis. If onion epidermal tissue is immersed in a solution of calcium nitrate, cells rapidly lose water by osmosis and the protoplasm of the cells shrinks (Web Figure 3. 8. A). This occurs because the calcium and nitrate ions freely permeate the cell wall and encounter the selectively permeable plasma membrane. The large vacuole in the center of the cell originally contains a dilute solution with much lower osmotic pressure than that of the calcium nitrate solution on the other side of the membrane. The vacuole thus loses water and becomes smaller. The space between the cell membrane and the cell wall enlarges and the plasma membrane and the protoplasm within it contract to the center of the cell. Strands of cytoplasm extend to the cell wall because of plasma membrane-cell wall attachment points. Plasmolysed cells die unless they are transferred quickly from the salt or sugar solution to water.

Plasmolysis of an epidermis cell of Allium cepa after addition of calcium nitrate. Both chloroplast and vacuole become unevenly shaped. Strands of cytoplasm remain between the shrinking protoplasm and the neighboring cells. (Courtesy of Dr. Peter v. Sengbusch, University of Hamburg. )

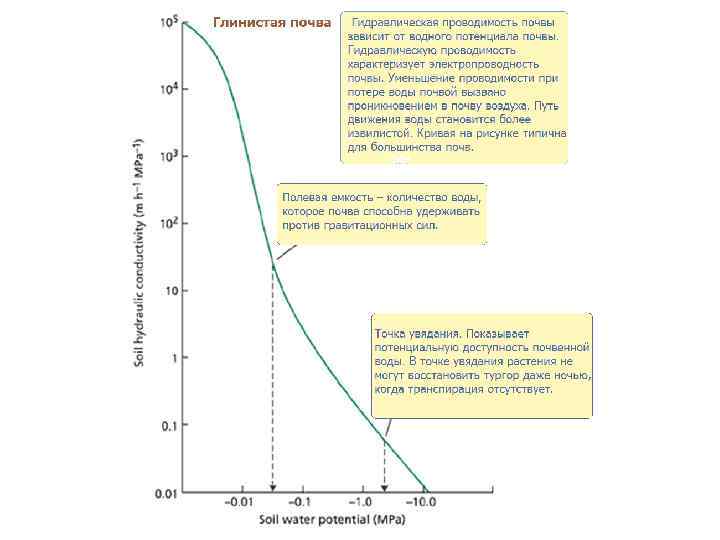
Понятие гидравлической проводимости Рассмотрим клетку с начальным водным потенциалом равным – 0. 2 MPa, погруженной в чистую воду. Вода будет двигаться в клетку за счет движущей силы ΔΨw = 0. 2 Mpa. С какой скоростью перемещается вода? Скорость зависит от гидравлической проницаемости мембраны (Lp) и движущей силы. Гидравлическая проницаемость выражает как вода может двигаться через мембрану. Она выражается в единицах объема воды, отнесенных к времени и к движущей силе: m 3 m– 2 s– 1 MPa– 1 or m s– 1 MPa– 1). Гидравлическая проводимость мембраны 10– 6 m s– 1 MPa– 1. Поток (Jv) можно определить по формуле:

Следует обратить внимание, что рассмотрена идеальная мембрана, т. е. проницаемая только для воды (но, не для растворенных веществ). Также считают, что составляющие водного потенциала (ΔΨs и ΔΨp ) равнозначны по отношению к потоку воды. Реальная (неидеальная) мембрана требует более сложного уравнения и разделения водных потоков, которые зависят от ΔΨs и by ΔΨp (Nobel 1999). В рассмотренном примере Jv выражается как 0. 2 x 10– 6 m s– 1. Это начальная максимальная скорость транспорта. По мере роста водного потенциала скорость падает (падает движущая сила (ΔΨw)) по экспоненциальному закону.

Водные потоки в растении зависят от градиента водного потенциала и от величины гидравлической проводимости тканей (клеток). По аналогии с законом Ома поток воды (Jw) в системе почва - растение - атмосфера, можно представить в виде гидравлической цепи:

В целом растении можно выделить три основных последовательно соединенных гидравлических сопротивления: R 1 - участок от поверхности клеток эпидермиса до сосудов ксилемы, R 2 - сопротивление элементов ксилемы, R 3 - сопротивление при испарении воды в атмосферу из листьев. Поскольку , а R (гидравлическое сопротивление) – величина обратная kw (гидравлическая проводимость), имеем: , где - суммарный градиент водного потенциала всей цепи, R - суммарное сопротивление всех элементов цепи.


Корневая система растения: 1 -общий вид, 2 -поперечный разрез, 3 и 4 –корневые волоски, 5 – кончик корня
")
Радиальный путь воды в корне (С. С. Медведев, 2004)

Схема основных путей перемещения воды и неорганических ионов из почвы через эпидерму и кору в ксилему. Вода движется в основном по апопласту, пока не достигнет эндодермы, где апопластное движение перекрывается поясками Каспари. Пояски Каспари заставляют воду на пути к ксилеме пересечь плазматические мембраны и протопласты эндодермальных клеток. Пройдя сквозь плазматическую мембрану на внутренней поверхности эндодермы, вода может снова пойти по апопластному пути до полостей элементов ксилемы. Неорганические ионы активно поглощаются эпидермальными клетками и затем перемещаются по симпласту через кору в паренхимные клетки, из которых они перекачиваются в элементы ксилемы.

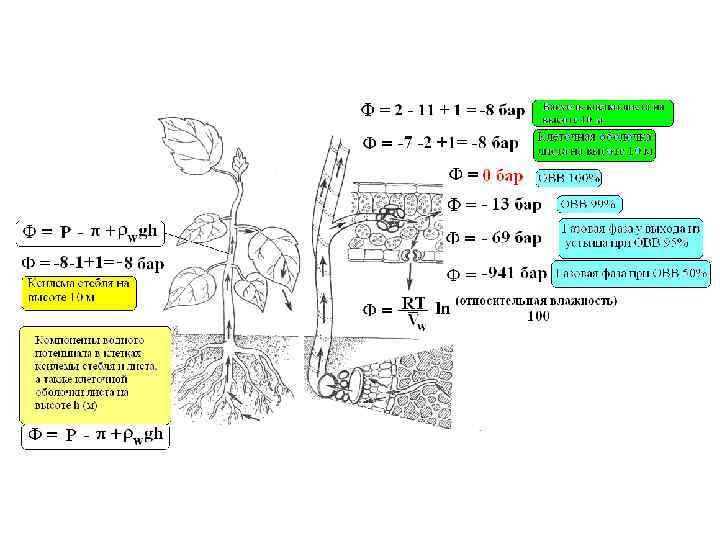
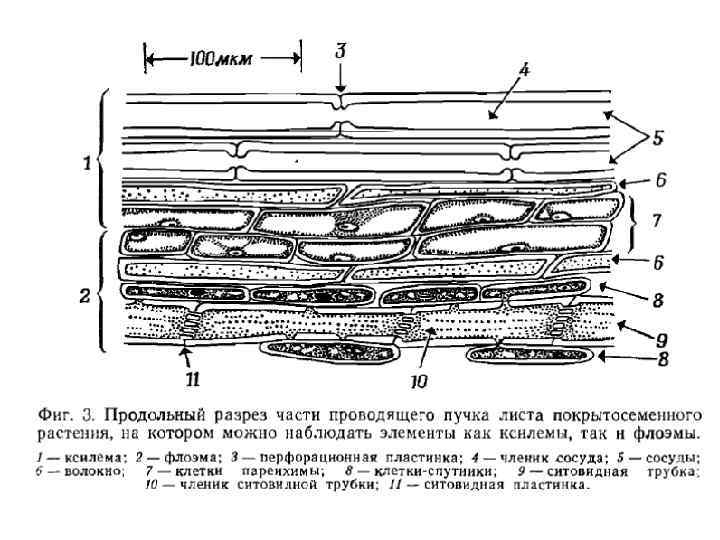

Движение воды по сосудам ксилемы происходит в соответствии с законом Пуазейля Ламинарное движение жидкости в цилиндре пропорционально квадрату его радиуса, градиенту гидростатического давления и обратно пропорционально вязкости раствора

Средний поток водного раствора в ксилеме транспирирующего дерева составляет 0, 1 см/сек или 3, 6 м/час. (Как можно измерить скорость движения воды ? ) При радиусе элемента ксилемы 20 мкм и вязкости 0, 010 П (Пуаз) градиент давления составляет: -0, 002 бар/см или -0, 2 бар/м При заданной скорости движения воды через поры клеточной оболочки с радиусом 5 нм потребуется градиент давления -30000 бар/см.

Скорость, с которой вода проходит вверх по стволу дерева зависит от типа дерева и транспирации. У деревьев с толстыми сосудами (r от 100 до 200 мкм), пиковые скорости от 4 до 13 мм с-1. Деревья с тонкими сосудами (r от 25 до 75 мкм) имеют более низкие пиковые скорости, от 0, 3 до 1, 7 мм с -1. Для промера мы будем величину скорости 4 мм с-1 для ксилемного транспорта и 40 мкм в качестве радиуса сосуда. Это высокая скорость для такого узкого сосуда, поэтому оценка градиента давления может оказаться завышенной.



Концентрация водяного пара в насыщенном воздухе в зависимости от температуры воздуха.

Натяжение воды , содержащейся в клеточной оболочке клеток листа. Главный вклад в общий водный потенциал клеточной стенки ( кл. ст. ) вносит матричный потенциал. Для количественных оценок используют уравнение для силы поверхностного натяжения в цилиндре со смачиваемыми стенками в условиях адгезии: Поверхностное натяжение σ воды составляет 7, 28× 10 -5 бар/см, промежутки между фибриллами целлюлозы клеточной оболочки достигают 10 нм, угол стремится к 0. Натяжение или отрицательное гидростатическое давление составит 291 бар. Для водного потенциала при ОВВ 95% равному -69 бар, достаточно, чтобы радиус капилляров был в пределах 20 -25 нм. В расчеты может внести коррективы концентрация раствора. Тогда диаметр капилляров снова придется уменьшить.

Поперечный срез листовой пластинки

Транспирация водяного пара. Градиенты Транспирация из листьев зависит от двух основных факторов: разница в концентрации паров воды между воздушными промежутками листьев и наружного воздуха и, диффузионного resisitance (г) этого пути. Эта концепция транспирации аналогичен потоку электронов в электрической цепи. Действительно, электрическим аналогом широко используется в качестве модели для потери паров воды от листа. В этом аналоге, сопротивления, связанные с каждой стороны пути; Основными из них являются сопротивление устьичной поры (rs) и сопротивление за счет слоя неперемешиваемого воздуха на поверхности листа (rb) (так называемый пограничный слой). Интенсивность транспирации (Е, в моль · м-2 с-1) может быть связано с диффузионным сопротивлением (R, в см-1) по следующей формуле:
 Листья в полевых условиях при 20")
Схема испарения воды чрез малые отверстия (правило Стефана) Листья в полевых условиях при 20 градусах Цельсия и скорости ветра 5 км час могут испарять 0, 02 г/см 2 час Определение количества устьиц на единицу поверхности листа. Различия верхней и нижней сторон листа.
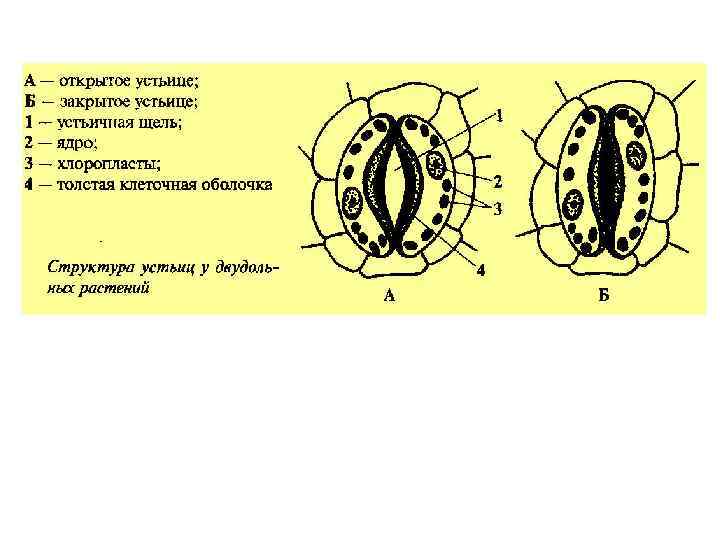
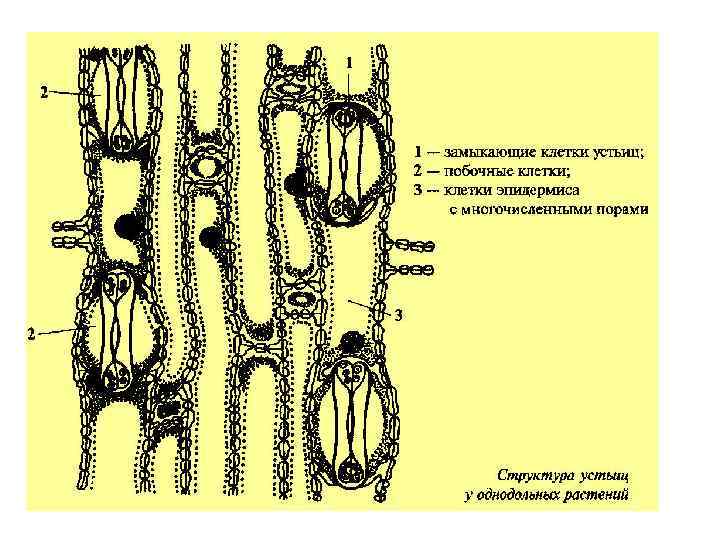
 А- -анионы, К+ - катионы, в основном, калия")
Схема работы устьиц (Taiz, Zeiger, 1998) А- -анионы, К+ - катионы, в основном, калия

Чем меньше относительная влажность атмосферного воздуха, тем ниже его водный потенциал. Если водный потенциал воздуха меньше водного потенциала подустьичных полостей, то молекулы воды испаряются нару- жу. Основным фактором, влияющим на открывание и закрывание устьиц, является содержание воды в листе, в том числе и в замыкающих клетках устьиц. Высокая оводненность замыкающих клеток приводит к открыванию устьиц. При недостатке воды замыкающие клетки выпрямляются – и устьичная щель закрывается. Кроме того, по мере увеличения водного дефицита в тканях растения повышается концентрация ингибитора роста абсцизовой ки- слоты. Она подавляет деятельность Н+ -насосов в плазмалемме замыкающих клеток, которые участвуют в Н+ /К+ обмене, вследствие чего снижается их тургор и устьица закрываются. Абсцизовая кислота также ингибирует синтез фермента α-амилазы, что приводит к снижению гидролиза крахмала, поэтому сосущая сила замыкающих клеток уменьшается – и устьица закрываются.
УМКД_ФР(клетка) (2).ppt