Молекулярная биология.pptx
- Количество слайдов: 52



ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯРОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ -МСХА ИМЕНИ К. А. ТИМИРЯЗЕВА Презентация на тему: Важнейшие открытия в генетике и молекулярной биологии. Выполнил студент 305 группы зооинженерного факультета Чистяков Александр.

ВАЖНЕЙШИЕ ОТКРЫТИЯ ГЕНЕТИКИ И МОЛЕКУЛЯРНОЙ БИОЛОГИИ 1865 – открытие Г. Менделем факторов наследственности 1868 г - открытие ДНК швейцарским врачом Ф. Мишером из спермы лосося. Он назвал его нуклеином 1871 – Ч. Дарвин опубликовал книгу «Происхождение человека и половой отбор» 1881 – Кассель получил аденин, гуанин , сахар и фосфорную кислоту из нуклеина 1900 – формальное рождение генетики. Независимая публикация статей Г. де Фриза, К. Корренса и Э. Чермака с изложением основных законов генетики

1902 – В, Саттон и Т. Бовери создают хромосомную теорию наследственности 1905 – У. Бетсон предложил название генетика 1910 – Т. Морган установил, что гены находятся в хромосомах. А. Кассель получил Н. П. За установление того, что в состав нуклеиновой кислоты входят аденин, гуанин , цитозин и тимин

1922 – Н. И. Вавилов сформулировал закон гомологических рядов о параллелизме в изменчивости родственных групп растений 1934 – Белозерский обнаружил ДНК в растениях 1931 – Барбара Мак-Клинток продемонстрировала явление кроссинговера 1938 – Ф. Гриффит открыл феномен трансформации (невирулентный штамм пневмококков превращался в вирулентный при смешивании с убитым вирулентным штаммом)

1940 - Дж. Бидл и Э. Татум сформулировали теорию – один ген – один фермент 1944 – Эвери из Рокфеллеровского института (США) показал, что за феномен трансформации отвечает ДНК 1950 – Э. Чаргафф сформулировал правило- количество А равно Т, количество Г равно Ц. Барбара Мак-Клинток показала существование транспозонов. (Н. П. В 1983 г) 1953 - Джеймс Уотсон и Френсис Крик открыли двойную спираль ДНК (Н. П. 1962 г). 60 – 70 годы 19 века – сформулированы канонические представления в молекулярной биологии

1951 – Р. Франклин и М. Уилкинс получили рентгенограмму ДНК 1953 - Джеймс Уотсон и Френсис Крик создали структурную модель ДНК - двойную спираль ДНК (Н. П. 1962 г). 1956 г- А. Корнберг открыл ДНК-полимеразу. Iи механизм биологического синтеза ДНК и РНК ( Н. П. Совместно с С. Очоа 1959 г) 1958 – М. Мезельсон и Ф. Сталь – полуконсервативный механизм репликации ДНК 1961 – начата расшифровка генетического кода. Н. П. 1968 г М. У. Ниренберг, Р. У. Холли, Х. Г. Корана 1962 – Дж. Гердон клонировал лягушку 60 – 70 годы 19 века – сформулированы канонические представления в молекулярной биологии

1969 – Х. Г. Корана синтезировал химическим путем ген 1970 – открытие обратной транскриптазы. Н. П. 1975 Г. Темин и Д. Балтимор 1972 –Пол Берг и Г. Бойер – получили первые рекомбинантные ДНК. Н. П. 1980. Заложены основы генной Инженерии 1973 - С. Коэн и Г. Бойер - стратегия переноса генов в бактериальные клетки 1974 – С. Милстайн и Г. Келер технология получения моноклональных антител. Н. П. вместе с Н. Ернев 1984

1975 – С. Тонегава показал расположение генов иммуноглобулинов в ДНК эмбриональных и лимфоинных клеток (Н. П. 1987) 1977 – У. Гилберт, А. Максам и Ф. Сенгер - быстрые методы секвенирования ДНК. (Н. П. 1980) 1976 – компания Genentech – перенос гена инсулина человека в бактериальную клетку 1985 – К. Б. Мюллис – ПЦР 1988 – начало международного проекта «Геном человека» 1995 – определена нуклеотидная последовательность Haemophilus influenzae. Становление геномики
 Я. Вильмут клонировал овечку")
1997 – открытие прионов С. Прузинер ( Н. П. 2000) Я. Вильмут клонировал овечку Долли 1998 – определена нуклеотидная последовательность ДНК Caenorabditis elegans. Феномен РНК-интерференции 1999 – полностью секвенирована 21 хромосома человека Клонирование мыши и коровы. Открытие оксида азота как регулятора 2000 – Черновой вариант генома человека и дрозофилы. Клонирование свиньи 2003 - завершение программы «Геном человека»



Нуклеиновые кислоты РНК ДНК Нуклеотид Остаток фосфорной кислоты Пентоза Азотистое основание

Нуклеотид 5’ 1’ Фосфат 3’ Азотистое основание (А или Г или Т или Ц или У) Пентоза (рибоза или дезоксирибоза) Тиминовый нуклеотид

Азотистые основания Пиримидиновые Пуриновые Аденин Цитозин Гуанин Тимин Урацил А Г ДНК РНК Ц У Т
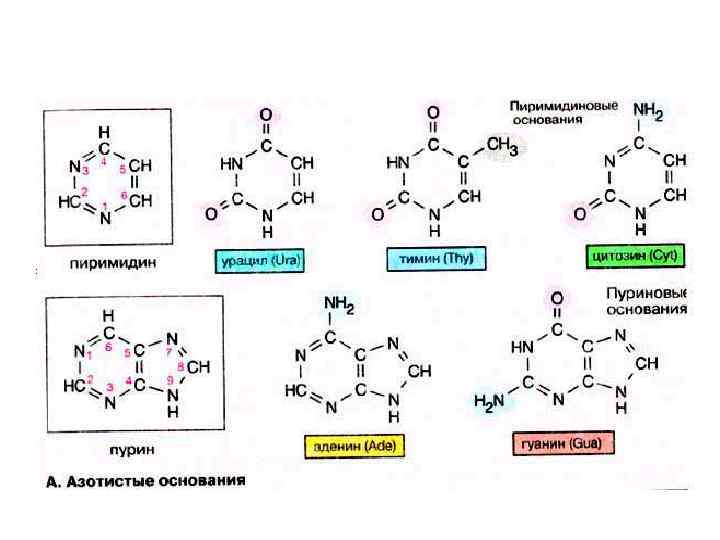

аденин гуанин тимин цитозин
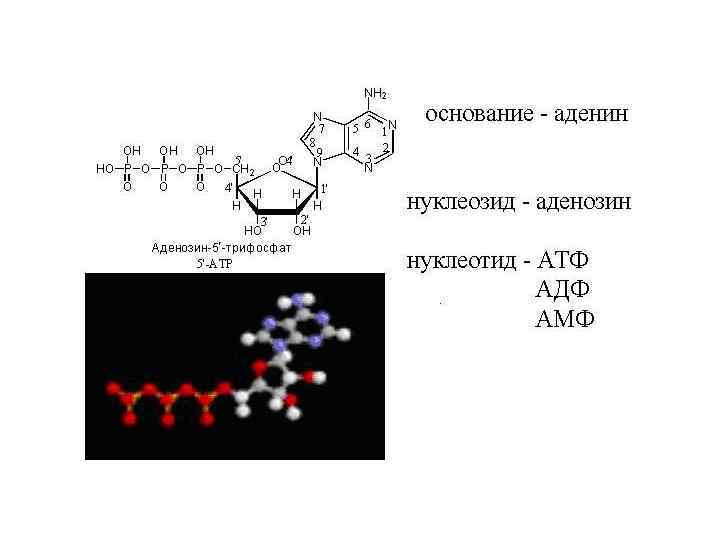
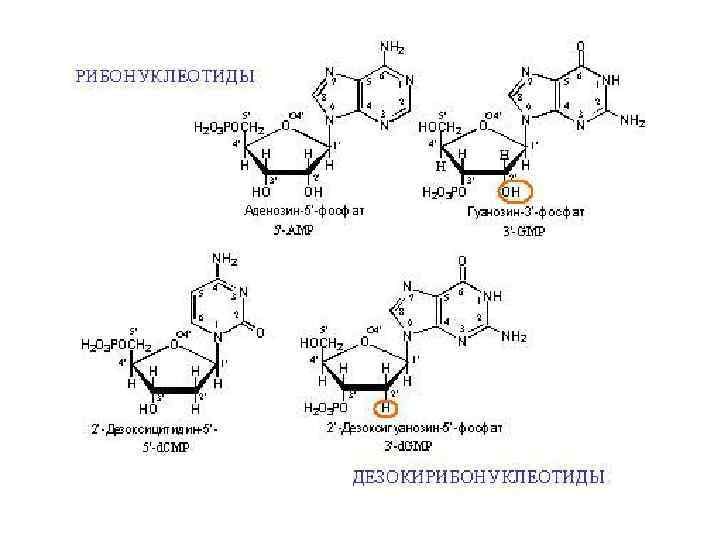

Функции нуклеотидов v мономеры нуклеиновых кислот v. Носители химической энергии в их легкогидролизуемой фосфоангидридной связи (АТФ) v. С другими группами образуют коэнзимы (Со. А, НАДН) v. Сигнальные молекулы с. АМР
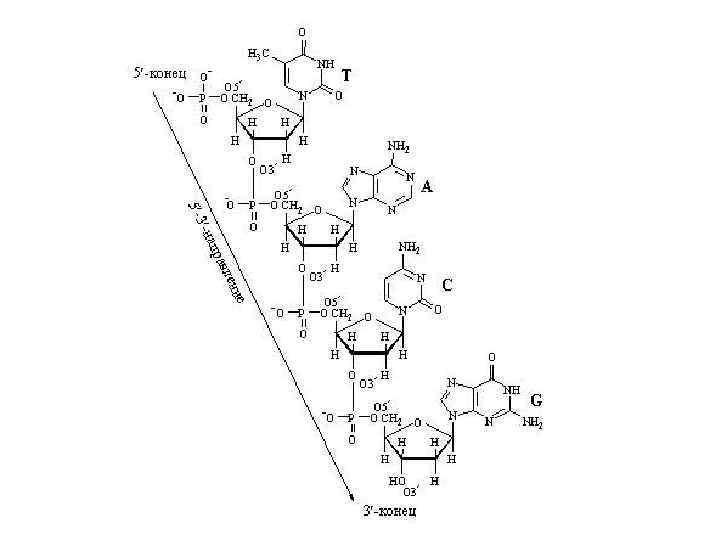

Цепь ДНК А 5’ 3’ 5’ Ц 3’ – 3’ 5’-фосфодиэфирная связь Динуклеотид

Пары оснований Принцип строения ДНК Водородные связи А=Т Г=Ц !
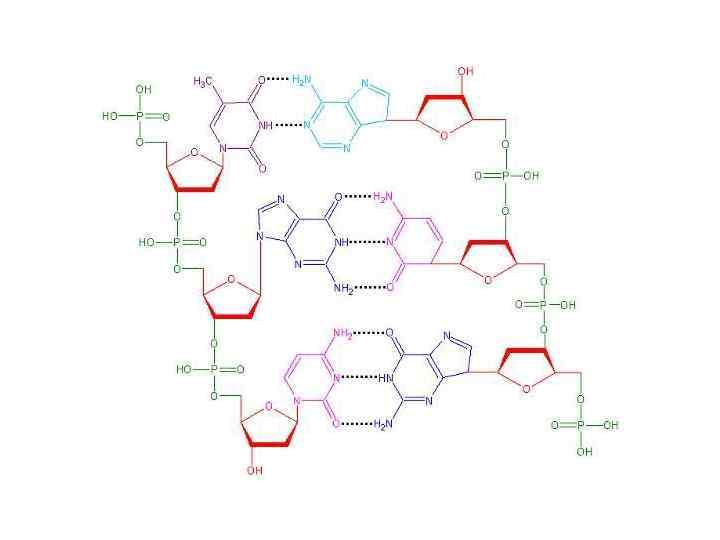

Правила Чаргаффа 1. 2. 3. 4. Количество А =Т Количество Г=Ц А+Г=Т=Ц кол-во пуринов=кол-ву пиримидинов Соотношение А+Ц: Г+Т разное у разных ДНК

ДНК РНК

Большая бороздка Малая бороздка

А Формы ДНК В Z
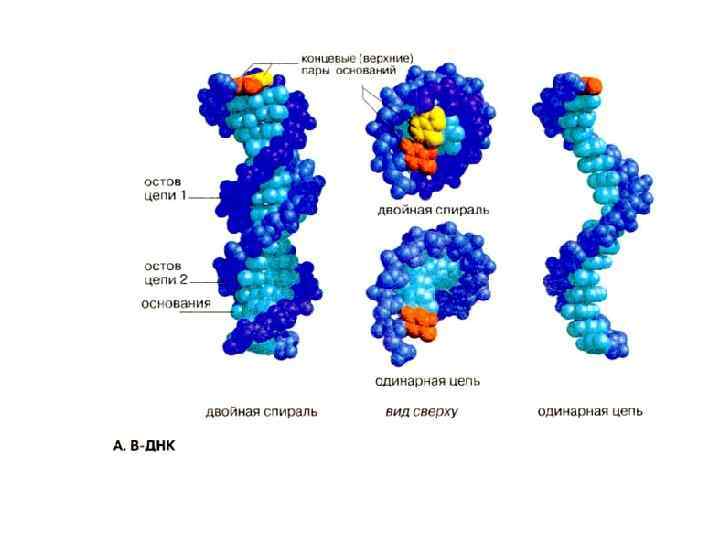
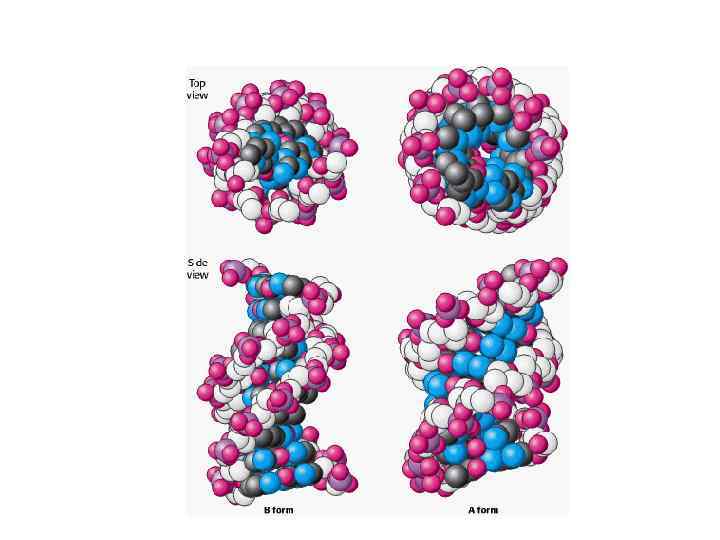

Физические параметры ДНК Диаметр – 2 нм Расстояние между парами нуклеотидов – 0, 34 нм 1 поворот спирали – 10 пар оснований Длина ДНК в самой длинной чел хромосоме – 8 см, во всех хромосомах одной клетки 2 м Длина ДНК в 1 млрд раз больше ширины В организме человека 5. 10 13 – 10 14 клеток поэтому Общая длина ДНК у человека 10 11 км, т. е. В 1000 раз больше, чем расстояние от Земли до Солнца. В гаплоидном наборе человека 3, 2 млрд пар нуклеотидов В ядре 7 пг ДНК
 и")
Молекулярная структура хромосом - гигантские нуклеопротеидные комплексы из ДНК, белков (гистоны и негистоновые) и РНК 1) Нуклеосомы – 200 п. н. на поверхности октамера гистонов (Н 2 А, Н 2 В, Н 3 и Н 4) 2) ДНП фибриллы – 30 нм- за счет ассоциации октамеров нуклеосом с гистоном Н 1. В одном нуклеомере 6 - 10 нуклеосом. Степень компактизации – 40 3) Петли – по 10 -60 тыс. п. н. в виде розетки с фиксированной плотной центральной зоной. Это нклеомер. Хромомер, содержит 20 -40 нуклеомеров. 4) Домены – хромомеры образуют хромонемы

Нуклеосома состоит из ядра - участка ДНК 147 п. н. закрученной 1 2/3 витка вокруг гистонового октамера Октамер включает по 2 молекулы Н 2 А, Н 2 В, Н 3 и Н 4 внутри и по разные концы торчат димеры Н 2 А/Н 2 В. На входе и выхоле нуклеосомы соединяются с ДНК по одному Н 1.

Топология ДНК в хромосомах Некодируюшая ДНК участвует в формировании и модуляции макроструктуры хромосомы – гетеро- и эухроматина При переносе гена в область гетерохроматина наблюдается его инактивация (эффект положения) Отдельные участки некодирующей ДНК могут использоваться как места посадки белков-скрепок, обеспечивающих различную упаковку хроматина

Компактность генома эукариот Компактность - другое принципиальное отличие генома эукариот от прокариотического генома. При средней разнице размеров геномов на 3 порядка, линейные размеры эукариотических хромосом соизмеримы с длиной ДНК прокариот. Выделяют, по крайней мере, 4 уровня компактизации ДНК. При этом нить ДНК "укорачивается" в 10000 раз. Это все равно, что нить, длиной с Останкинскую башню (500 м), уложить в спичечный коробок (5 см). Два первых уровня компактизации эукариотического генома обеспечиваются гистонами.

Общая характеристика гистонов • Гистоны - основные белки. Все они обогащены лизином и аргинином - положительно заряженными аминокислотами. Выделяют 5 фракций гистонов. Нарабатывается их очень много - 60 млн. молекул каждой фракции на клетку. • Все гистоны, кроме Н 1, черезвычайно консервативны в эволюционном отношении (у коровы и клевера разница в Н 2 А всего в одну аминокислоту!). Следовательно, эти белки выполняют принципиальную функцию, которая у всех эукариот обеспечивается одинаково. • Любая мутация в гистоновых генах летальна. • Н 1 - очень вариабельная фракция. Этот гистон различен не только у видов, но даже у одного организма, в зависимости от стадий онтогенеза. • В гистонах лизин и аргинин располагаются кластерами. Положительно заряженные аминокислоты гистонов обеспечивают электростатические взаимодействия с ДНК. Гидрофобная часть необходима для взаимодействия гистонов между собой.

Четыре уровня компактизации ДНК 1. Нуклеосомный уровень. В основе нуклеосомы лежит гистоновый октамер. Расположение гистонов не случайно. Каждая молекула представлена дважды. Они образуют кор (серцевину) нуклеосомы. На кор наматывается ДНК - 1. 75 левых витка спирали. Нуклеосомой называется повторяющийся структурный элемент хроматина, содержащий гистоновый октамер и ~180 п. н. ДНК. Непосредственно с октамером контактирует 145 п. н. и 20 -30 -40 п. н. между нуклеосомными корами. Нуклеосомный уровень упаковки свойственен всей эукариотической ДНК, он дает укорочение в 7 раз. Диаметр увеличивается с 20 Å до 110 Å. Гистоновые октамеры "скользят" по ДНК.
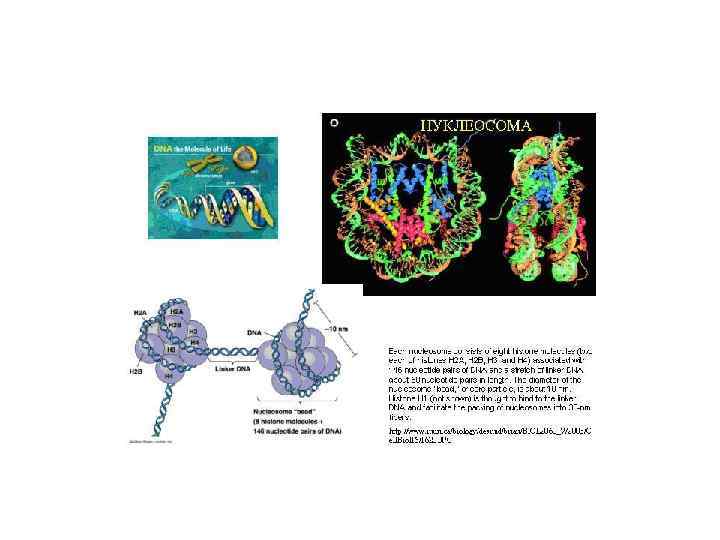

Нуклеосомы

2. Супербидный, или соленоидный, уровень. • Фактически обеспечивается Н 1 гистоном. Н 1 взаимодействует с октамерами, сближает их, и еще на него наматывется ДНК. Образуется супербид. • Происходит сокращение линейного размера ДНК в 6 -10 раз. Диаметр увеличивается до 300Å. • Этот уровень компактизации, как и первый, не зависит от первичной структуры ДНК.

3. Петлевой уровень. • Обеспечивается негистоновыми белками. Они узнают определенные последовательности ДНК и связываются с ними и другом, образуя петли по 20 -80 тыс. п. н. • Петля обеспечивает экспрессию гена, т. е петля является не только структурным, но и функциональным образованием. • Есть участки, в которых нет петель. Укорочение за счет петель проходит в 20 -30 раз. Образуются и петлевые домены. Диаметр увеличивается до 700Å.

Петли ДНК

4. Метафазная хромосома. • Метафазная хромосома уже удвоена. Она состоит из двух хроматид. Каждая из них содержит одну молекулу ДНК. Сюда входят белки ядерной мембраны, серия белковых нитей, сопряженных с ядерной оболочкой и пронизывающих все ядро. • Чем больше в геноме А-Т пар, тем больше возможностей для изменения вторичной структуры ДНК. При суперспирализации ДНК А-Т богатые участки плавятся в первую очередь.
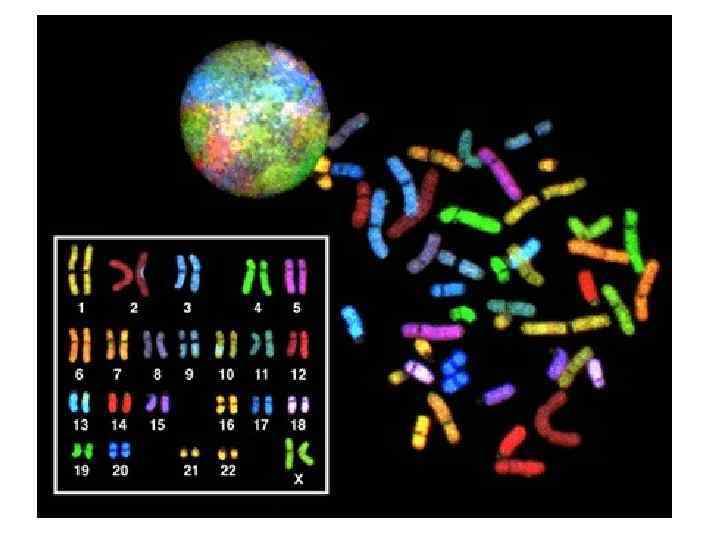


В метафазной хромосоме размер ДНК уменьшен в 8000 раз

Пространственная организация хромосомных доменов в ядре Петли ДНК закреплены на ядерном матриксе. Имеют участки origin и связаны с топоизомеразой II Участки прикрепления имеют MAR элементы (matrix Associated region) и регуляторные элементы генов – промоторы, энхансеры, сайленсеры, инсуляторы.

Политенные хромосомы

Функция ДНК: хранение и передача генетической информации в ряду поколений

Функции РНК: 1. Перенос информации от ДНК к месту синтеза белка 2. Транспорт аминокислот к месту синтеза белка 3. Создание структур для синтеза белка 4. Ферментативная активность

РНК Пентоза рибоза Азотистые основания А У Г Ц Цепь одна
 и")
Молекулярная структура хромосом - гигантские нуклеопротеидные комплексы из ДНК, белков (гистоны и негистоновые) и РНК 1) Нуклеосомы – 200 п. н. на поверхности октамера гистонов (Н 2 А, Н 2 В, Н 3 и Н 4) 2) ДНП фибриллы – 30 нм- за счет ассоциации октамеров нуклеосом с гистоном Н 1. В одном нуклеомере 6 - 10 нуклеосом. Степень компактизации – 40 3) Петли – по 10 -60 тыс. п. н. в виде розетки с фиксированной плотной центральной зоной. Это нклеомер. Хромомер, содержит 20 -40 нуклеомеров. 4) Домены – хромомеры образуют хромонемы

Топология ДНК в хромосомах Некодируюшая ДНК участвует в формировании и модуляции макроструктуры хромосомы – гетеро- и эухроматина При переносе гена в область гетерохроматина наблюдается его инактивация (эффект положения) Отдельные участки некодирующей ДНК могут использоваться как места посадки белков-скрепок, обеспечивающих различную упаковку хроматина
Молекулярная биология.pptx