

фармакология рецепторов.pptx
- Количество слайдов: 52
 Фармакология рецепторов ВЫПОЛНИЛИ: СТУДЕНТК И 3 КУРС А КУЗН ЕЦОВА ЛЮДМИЛА И ХУДА ДА ТОВ А ГУЛЬВЕР Д
Фармакология рецепторов ВЫПОЛНИЛИ: СТУДЕНТК И 3 КУРС А КУЗН ЕЦОВА ЛЮДМИЛА И ХУДА ДА ТОВ А ГУЛЬВЕР Д
 Рецепторы-белковые образования в организме, предназначенные для взаимодействия с эндогенными соединениями, нейромедиаторами. При нервном импульсе медиатор (вещество) выделяется и воздействует на рецептор
Рецепторы-белковые образования в организме, предназначенные для взаимодействия с эндогенными соединениями, нейромедиаторами. При нервном импульсе медиатор (вещество) выделяется и воздействует на рецептор

 Взаимодействие лиганда с рецептором
Взаимодействие лиганда с рецептором
 Взаимодействие лиганда с рецептором Модель кинетики взаимодействия по E. J. Ariens, A. J. Beld (1977) Обозначим: L лиганд (любое вещ-во, способное связаться с рецептором); R рецептор в исходном состоянии; R рецептор в возбужденном состоянии; R рец-р в рефрактерном состоянии (десенситизация); KX константа скорости прямой реакции; K X константа скорости обратной реакции. . Тогда:
Взаимодействие лиганда с рецептором Модель кинетики взаимодействия по E. J. Ariens, A. J. Beld (1977) Обозначим: L лиганд (любое вещ-во, способное связаться с рецептором); R рецептор в исходном состоянии; R рецептор в возбужденном состоянии; R рец-р в рефрактерном состоянии (десенситизация); KX константа скорости прямой реакции; K X константа скорости обратной реакции. . Тогда:
 Взаимодействие лиганда с рецептором Свойства лиганда опеределяет только судьба лиганд-рецепторного комплекса LR: K 2 >> K-1 K 2 << K-1 K 2 = K-1 = 0
Взаимодействие лиганда с рецептором Свойства лиганда опеределяет только судьба лиганд-рецепторного комплекса LR: K 2 >> K-1 K 2 << K-1 K 2 = K-1 = 0
 Строение «типового» химического синапса
Строение «типового» химического синапса
, 2, Н Угнетение: 1") Пресинаптические рецепторы: действие на освобождение медиатора Стимуляция: 1 (в НМС), 2, Н Угнетение: 1 (в ВГ), 2, D 2, M, опиатные
Пресинаптические рецепторы: действие на освобождение медиатора Стимуляция: 1 (в НМС), 2, Н Угнетение: 1 (в ВГ), 2, D 2, M, опиатные
 Пресинаптические рецепторы: действие на обратный захват медиатора
Пресинаптические рецепторы: действие на обратный захват медиатора
 Главные механизмы действия рецепторов
Главные механизмы действия рецепторов
 Фосфолипазный механизм
Фосфолипазный механизм
 Signal transduction
Signal transduction
 АДРЕНЕРГИЧЕСКИЙ СИНАПС
АДРЕНЕРГИЧЕСКИЙ СИНАПС
 Адренорецептор это конформационно- лабильная генетически детерминированная гликопротеиновая молекула. Его функция состоит, в специфическом узнавании молекул медиатора и трансформации этого сигнала с участием ионов и циклических нуклеотидов на другие биоструктуры клетки
Адренорецептор это конформационно- лабильная генетически детерминированная гликопротеиновая молекула. Его функция состоит, в специфическом узнавании молекул медиатора и трансформации этого сигнала с участием ионов и циклических нуклеотидов на другие биоструктуры клетки
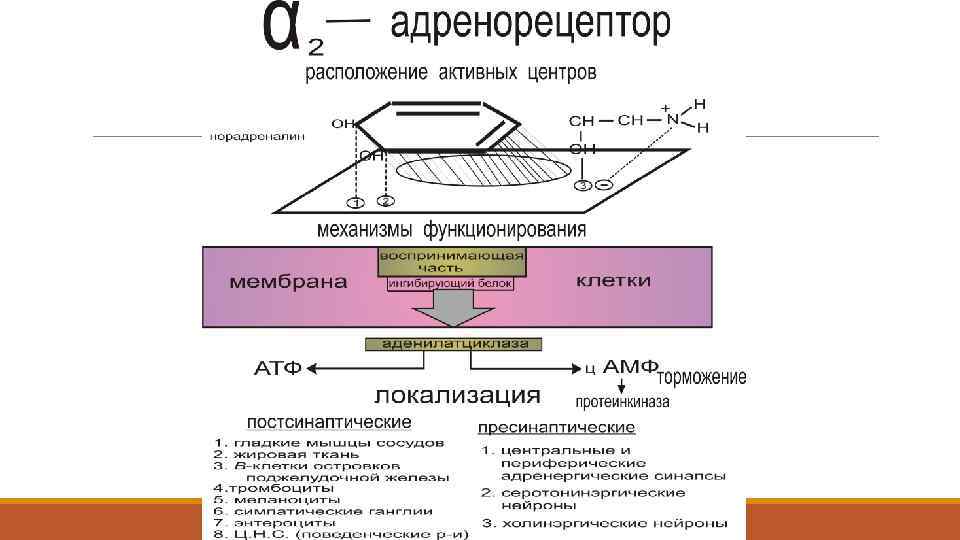
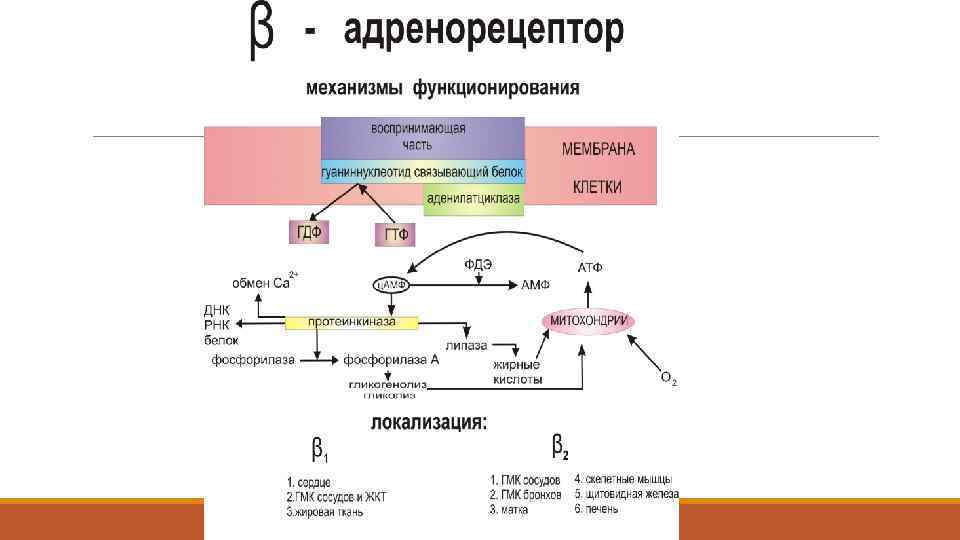
 адренорецепторы Мембранный белок 62 k. Da. Структура в настоящее время известна. различают α и β -типы АР, которые делятся на подтипы: α 1, α 2 - и β 1 , β 2 АР
адренорецепторы Мембранный белок 62 k. Da. Структура в настоящее время известна. различают α и β -типы АР, которые делятся на подтипы: α 1, α 2 - и β 1 , β 2 АР
 Молекула α адренорецептора содержит 7 мембранно-связанных участков β-адренорецептор- тоже 7 участков, но в виде спирали:
Молекула α адренорецептора содержит 7 мембранно-связанных участков β-адренорецептор- тоже 7 участков, но в виде спирали:
 Распределение адренорецепторов: l Пресинаптические: ЦНС- регулируют высвобождение медиаторов Постсинаптические 2 Постсинаптические 1: n Гладкие мышцы сосудов, Радиальная мышца радужки Миокард Печень Селезенка ЦНС Гладкие мышцы сосудов Поджелудочная железа Жировые клетки Симпатические ганглии Тромбоциты ЦНС
Распределение адренорецепторов: l Пресинаптические: ЦНС- регулируют высвобождение медиаторов Постсинаптические 2 Постсинаптические 1: n Гладкие мышцы сосудов, Радиальная мышца радужки Миокард Печень Селезенка ЦНС Гладкие мышцы сосудов Поджелудочная железа Жировые клетки Симпатические ганглии Тромбоциты ЦНС
 Пресинаптические -адренорецепторы: опосредуют высвобождение медиаторов из нервных окончаний Постсинаптические 1 рецепторы: Сердце Печень и скелетная мускулатура Жировая ткань ЖКТ Постсинаптичекие 2 рецепторы: Артериолы Гладкие мышцы бронхов Матка -клетки поджелудочной железы Щитовидная железа Клетки Лейдига
Пресинаптические -адренорецепторы: опосредуют высвобождение медиаторов из нервных окончаний Постсинаптические 1 рецепторы: Сердце Печень и скелетная мускулатура Жировая ткань ЖКТ Постсинаптичекие 2 рецепторы: Артериолы Гладкие мышцы бронхов Матка -клетки поджелудочной железы Щитовидная железа Клетки Лейдига
 Эффекты возбуждения альфаадренорецепторов Повышение тонуса сосудов- увеличение артериального давления. Сокращение радиальной мышцы радужки - расширение зрачка.
Эффекты возбуждения альфаадренорецепторов Повышение тонуса сосудов- увеличение артериального давления. Сокращение радиальной мышцы радужки - расширение зрачка.
 Стимуляция бета 1 адренорецепторов ионы кальция, поступающие через кальциевые каналы, активируют вход кальция из саркоплазматического ретикулума кардиомиоцитов. Связывая тормозной комплекс тропонин-тропомиозин, ионы кальция способствуют взаимодействию актина и миозина.
Стимуляция бета 1 адренорецепторов ионы кальция, поступающие через кальциевые каналы, активируют вход кальция из саркоплазматического ретикулума кардиомиоцитов. Связывая тормозной комплекс тропонин-тропомиозин, ионы кальция способствуют взаимодействию актина и миозина.
 Стимуляция бета-1 адренорецепторов сердца Основные эффекты: - повышение силы сокращений, - повышение частоты сокращений, - облегчение атриовентрикулярной проводимости, - повышение автоматизма волокон проводящей системы. повышение силы сокращений,
Стимуляция бета-1 адренорецепторов сердца Основные эффекты: - повышение силы сокращений, - повышение частоты сокращений, - облегчение атриовентрикулярной проводимости, - повышение автоматизма волокон проводящей системы. повышение силы сокращений,
 Стимуляция бета - 2 адренорецепторов Основные эффекты: - расширение кровеносных сосудов, - расслабление гладких мышц бронхов, - снижение тонуса и ритмической сократительной активности миометрия, - гликонеогенолиз.
Стимуляция бета - 2 адренорецепторов Основные эффекты: - расширение кровеносных сосудов, - расслабление гладких мышц бронхов, - снижение тонуса и ритмической сократительной активности миометрия, - гликонеогенолиз.
 α 2 + Gi-белки Аденилатциклаза") Норадреналин β + GS-белки + Аденилатциклаза ц. АМФ (↑) α 2 + Gi-белки Аденилатциклаза ц. АМФ (↓) Протеинкиназы α 1 + Gq-белки + Фосфолипаза С IP 3(↑), DAG(↑)
Норадреналин β + GS-белки + Аденилатциклаза ц. АМФ (↑) α 2 + Gi-белки Аденилатциклаза ц. АМФ (↓) Протеинкиназы α 1 + Gq-белки + Фосфолипаза С IP 3(↑), DAG(↑)
 Адренорецепторы: возбуждения эффекты
Адренорецепторы: возбуждения эффекты
 Адренорецепторы: возбуждения эффекты
Адренорецепторы: возбуждения эффекты
 (+) 2 S, 3 R, 5 S - Мускарин-алкалоид мухомора") Подтипы холинорецепторов М-холинорецептор (мускариночуствительный) (+) 2 S, 3 R, 5 S - Мускарин-алкалоид мухомора красного и некоторых других грибов. H-холинорецептор (никотинчуствительный) гликопротеид Мм 50000. БЕЛЯНИН М. Л. 28
Подтипы холинорецепторов М-холинорецептор (мускариночуствительный) (+) 2 S, 3 R, 5 S - Мускарин-алкалоид мухомора красного и некоторых других грибов. H-холинорецептор (никотинчуствительный) гликопротеид Мм 50000. БЕЛЯНИН М. Л. 28
 1. Молекулярное строение н-холинорецепторов Получение н-холинорецепторов в чистом виде стало возможным благодаря наличию маркера - альфа-бунгаротоксина, полипептида, выделенного из яда змей. Исследования очищенного препарата н-холинорецепторов позволили установить, что они имеют гликопротеидную природу и состоят из пяти субъединиц: На эффекторных клетках н-холинорецепторы расположены трансмембранно: они “прошивают” всю толщу мембраны, выступая над ней и частично углубляясь внутрь клетки. Субъединицы н-холинорецептора образуют канал-ионофор диаметром приблизительно 6, 5 А, проницаемый для Nа + и Са+. Взаимодействие ацетилхолина с н-холинорецептором и образование между ними комплекса приводит к открытию канала, входу Na+ в клетку, деполяризации постсинаптической мембраны и возбуждению эффекторной клетки. Проведение одного нервного импульса соответствует открытию 8 тысяч каналов. Время существования комплекса ацетилхолина с нхолинорецептором составляет 0, 1 мс, после чего ацетилхолин диссоциирует с рецептора и гидролизуется ацетилхолинэстеразой. Несмотря на отсутствие медиатора, канал остается открытым и функционирует 1 мс. Одна молекула н-холинорецептора связывает 2 молекулы ацетилхолина, или 2 молекулы альфа-бунгаротоксина, что позволяет предположить наличие узнающих центров на альфа-субъединицах. Участки, связывающие ацетилхолин, имеют 2 центра: анионный, вступающий в электростатическое взаимодействие с катионной головкой медиатора, и эстерофильный, образующий водородные связи и способный вступать в диполь-дипольное взаимодействие с карбонильной группой и кислородом сложноэфирной связи ацетилхолина. Обнаружено, что н-холинорецепторы гетерогенны, поскольку неодинаково реагируют на агонисты (никотин) и антагонисты (ганглиоблокаторы и курареподобные средства). Различают рецепторы «мышечного» типа, нейрональные, «ганглионарного» и эпителиально-нейронального типа. БЕЛЯНИН М. Л. 29
1. Молекулярное строение н-холинорецепторов Получение н-холинорецепторов в чистом виде стало возможным благодаря наличию маркера - альфа-бунгаротоксина, полипептида, выделенного из яда змей. Исследования очищенного препарата н-холинорецепторов позволили установить, что они имеют гликопротеидную природу и состоят из пяти субъединиц: На эффекторных клетках н-холинорецепторы расположены трансмембранно: они “прошивают” всю толщу мембраны, выступая над ней и частично углубляясь внутрь клетки. Субъединицы н-холинорецептора образуют канал-ионофор диаметром приблизительно 6, 5 А, проницаемый для Nа + и Са+. Взаимодействие ацетилхолина с н-холинорецептором и образование между ними комплекса приводит к открытию канала, входу Na+ в клетку, деполяризации постсинаптической мембраны и возбуждению эффекторной клетки. Проведение одного нервного импульса соответствует открытию 8 тысяч каналов. Время существования комплекса ацетилхолина с нхолинорецептором составляет 0, 1 мс, после чего ацетилхолин диссоциирует с рецептора и гидролизуется ацетилхолинэстеразой. Несмотря на отсутствие медиатора, канал остается открытым и функционирует 1 мс. Одна молекула н-холинорецептора связывает 2 молекулы ацетилхолина, или 2 молекулы альфа-бунгаротоксина, что позволяет предположить наличие узнающих центров на альфа-субъединицах. Участки, связывающие ацетилхолин, имеют 2 центра: анионный, вступающий в электростатическое взаимодействие с катионной головкой медиатора, и эстерофильный, образующий водородные связи и способный вступать в диполь-дипольное взаимодействие с карбонильной группой и кислородом сложноэфирной связи ацетилхолина. Обнаружено, что н-холинорецепторы гетерогенны, поскольку неодинаково реагируют на агонисты (никотин) и антагонисты (ганглиоблокаторы и курареподобные средства). Различают рецепторы «мышечного» типа, нейрональные, «ганглионарного» и эпителиально-нейронального типа. БЕЛЯНИН М. Л. 29
 Строение Н-холинорецептора
Строение Н-холинорецептора
 Локализация Н - холинорецепторов I Пресинаптические: в постганглионарных волокон симпатической и парасимпатической НС (высвобождение н. а). В ЦНС: полосатое тело (высвобождение дофамина и серотонина) II Постсинаптические: 1. Вегетативные ганглии СНС и ПНС. 2. Надпочечники (мозговое вещество). 3. Скелетные мышцы. 4. Синокаротидная зона. 5. ЦНС: кора г. м. , нейрогипофиз, клетки Реншоу и др.
Локализация Н - холинорецепторов I Пресинаптические: в постганглионарных волокон симпатической и парасимпатической НС (высвобождение н. а). В ЦНС: полосатое тело (высвобождение дофамина и серотонина) II Постсинаптические: 1. Вегетативные ганглии СНС и ПНС. 2. Надпочечники (мозговое вещество). 3. Скелетные мышцы. 4. Синокаротидная зона. 5. ЦНС: кора г. м. , нейрогипофиз, клетки Реншоу и др.
 хроно- и дромотропный") М-холинорецепторы Эффекты: М 1: ЦНС, усиление моторики ЖКТ М 2: (-) хроно- и дромотропный эффекты М 3: миоз, усиление секреции, спазм гладких мышц, релаксация сфинктеров М 4, М 5: ЦНС
М-холинорецепторы Эффекты: М 1: ЦНС, усиление моторики ЖКТ М 2: (-) хроно- и дромотропный эффекты М 3: миоз, усиление секреции, спазм гладких мышц, релаксация сфинктеров М 4, М 5: ЦНС
 D 1 -ДР находятся на переферии (расширение") Дофаминовые рецепторы (D 1 и D 2) D 1 -ДР находятся на переферии (расширение сосудов, релаксация мышц) D 2 -ДР центральное действие Нарушение обмена дофамина - шизофрения БЕЛЯНИН М. Л. 33
Дофаминовые рецепторы (D 1 и D 2) D 1 -ДР находятся на переферии (расширение сосудов, релаксация мышц) D 2 -ДР центральное действие Нарушение обмена дофамина - шизофрения БЕЛЯНИН М. Л. 33
 РЕЦЕПТОРЫ ДОФАМИНА
РЕЦЕПТОРЫ ДОФАМИНА
 Локализуются") Серотониновые рецепторы 5 -НТ 1, 5 -НТ 2, 5 -НТ 3 (5 -hydroxytryphtamine) Локализуются в ЦНС и в периферических органах. Возбуждение приводит к повышению АД, сокращению сосудов, повышение свертываемости крови Индопан – агонист 5 -НТ 1 рецепторов БЕЛЯНИН М. Л. 35
Серотониновые рецепторы 5 -НТ 1, 5 -НТ 2, 5 -НТ 3 (5 -hydroxytryphtamine) Локализуются в ЦНС и в периферических органах. Возбуждение приводит к повышению АД, сокращению сосудов, повышение свертываемости крови Индопан – агонист 5 -НТ 1 рецепторов БЕЛЯНИН М. Л. 35
 РЕЦЕПТОРЫ СЕРОТОНИНА
РЕЦЕПТОРЫ СЕРОТОНИНА
 РЕЦЕПТОРЫ СЕРОТОНИНА
РЕЦЕПТОРЫ СЕРОТОНИНА
 РЕЦЕПТОРЫ СЕРОТОНИНА СИОЗС - антидепрессанты Антагонисты 5 -НТ в ЦНС – нейролептики и бензодиазепины “D”-блокаторы: противомигренозные средства (суматриптан, лизурид) 5 -НТ 3 -блокаторы – противорвотные (без седации!): ◦ Ондансетрон (зофран): 150 мкг/кг ◦ Гранисетрон: 10 -40 мкг/кг Трамадол: слабый опиоид, снижает выброс и обратный захват 5 -НТ
РЕЦЕПТОРЫ СЕРОТОНИНА СИОЗС - антидепрессанты Антагонисты 5 -НТ в ЦНС – нейролептики и бензодиазепины “D”-блокаторы: противомигренозные средства (суматриптан, лизурид) 5 -НТ 3 -блокаторы – противорвотные (без седации!): ◦ Ондансетрон (зофран): 150 мкг/кг ◦ Гранисетрон: 10 -40 мкг/кг Трамадол: слабый опиоид, снижает выброс и обратный захват 5 -НТ
 ГАМК-рецепторы По современным представлениям, ГАМКа рецептор является олигомерным гликопротеидным комплексом, имеющим по крайней мере 2 вида субъединиц: а и р. Молекулярная масса комплекса для различных областей головного мозга крыс составляет 220 -270 к. Да. Полагают, что а-субъединица связывает бензодиазепины, В-субъединица - ГАМК и мусцимол. Установлена молекулярная масса а и В субъединиц. Она равна 53 к. Да и 57 к. Да, соответственно. Предполагают, что обе субъединицы имеют частично гомологичную последовательность аминокислот. Это может отражать общность кодирующего их гена.
ГАМК-рецепторы По современным представлениям, ГАМКа рецептор является олигомерным гликопротеидным комплексом, имеющим по крайней мере 2 вида субъединиц: а и р. Молекулярная масса комплекса для различных областей головного мозга крыс составляет 220 -270 к. Да. Полагают, что а-субъединица связывает бензодиазепины, В-субъединица - ГАМК и мусцимол. Установлена молекулярная масса а и В субъединиц. Она равна 53 к. Да и 57 к. Да, соответственно. Предполагают, что обе субъединицы имеют частично гомологичную последовательность аминокислот. Это может отражать общность кодирующего их гена.
 ГАМК-рецепторы Локализованы на пресинаптической мембране, выполняют тормозную функцию (препятствуют проведению возбуждения путем блокировки выделения нейромедиаторов). Различают несколько подтипов (А, В, С). ГАМК - гамма-аминомасляная кислота (4 аминобутановая кислота) – тормозной медиатор ЦНС.
ГАМК-рецепторы Локализованы на пресинаптической мембране, выполняют тормозную функцию (препятствуют проведению возбуждения путем блокировки выделения нейромедиаторов). Различают несколько подтипов (А, В, С). ГАМК - гамма-аминомасляная кислота (4 аминобутановая кислота) – тормозной медиатор ЦНС.
 Модель ГАМК В - рецептора
Модель ГАМК В - рецептора
 Механизм работы ГАМК В рецептора Функциональные ГАМКB-рецепторы существуют в виде димера, расположенного в плазматической мембране. Субъединица R 1 связывает агонист ГАМК, субъединица R 2 активирует три пути передачи сигнала (пунктирные линии) через комплексы G-белков (Gαβγ). Gα активирует аденилатциклазу, приводя к увеличению концентрации аденозинмонофосфата (ц. АМФ); Gβγ взаимодействует с кальциевыми каналами, уменьшая поступления ионов кальция в клетку; также Gβγ активирует калиевые каналы, увеличивая количество ионов К+, выходящих из клетки.
Механизм работы ГАМК В рецептора Функциональные ГАМКB-рецепторы существуют в виде димера, расположенного в плазматической мембране. Субъединица R 1 связывает агонист ГАМК, субъединица R 2 активирует три пути передачи сигнала (пунктирные линии) через комплексы G-белков (Gαβγ). Gα активирует аденилатциклазу, приводя к увеличению концентрации аденозинмонофосфата (ц. АМФ); Gβγ взаимодействует с кальциевыми каналами, уменьшая поступления ионов кальция в клетку; также Gβγ активирует калиевые каналы, увеличивая количество ионов К+, выходящих из клетки.
 Гистаминовые рецепторы В организме существуют специфические рецепторы, для которых гистамин является естественным лигандом. В настоящее время различают три подгруппы гистаминовых (Н) рецепторов: Н 1 -, Н 2 - и Н 3 -рецепторы. H 1 рецепторы в гладких мышцах, эндотелии сосудов, ЦНС (постсинаптические). Эффекты: вазодилатация, спазм гладкой мускулатуры бронхов, раздвижение клеток эндотелия (и, как следствие, транссудации жидкости в околососудистое пространство, отек, крапивница. H 2 рецепторы. Стимуляция секреции желудочного сока. H 3 рецепторы (пресинаптические). Центральная и переферическая нервная система Подавление высвобождения нейромедиаторов.
Гистаминовые рецепторы В организме существуют специфические рецепторы, для которых гистамин является естественным лигандом. В настоящее время различают три подгруппы гистаминовых (Н) рецепторов: Н 1 -, Н 2 - и Н 3 -рецепторы. H 1 рецепторы в гладких мышцах, эндотелии сосудов, ЦНС (постсинаптические). Эффекты: вазодилатация, спазм гладкой мускулатуры бронхов, раздвижение клеток эндотелия (и, как следствие, транссудации жидкости в околососудистое пространство, отек, крапивница. H 2 рецепторы. Стимуляция секреции желудочного сока. H 3 рецепторы (пресинаптические). Центральная и переферическая нервная система Подавление высвобождения нейромедиаторов.
 РЕЦЕПТОРЫ ГИСТАМИНА
РЕЦЕПТОРЫ ГИСТАМИНА
 РЕЦЕПТОРЫ ГИСТАМИНА
РЕЦЕПТОРЫ ГИСТАМИНА
 РЕЦЕПТОРЫ ВАПП
РЕЦЕПТОРЫ ВАПП
 РЕЦЕПТОРЫ ВАПП
РЕЦЕПТОРЫ ВАПП
 Опиоидные рецепторы — разновидность рецепторов нервной системы, относящихся к рецепторам, сопряженным с G белком. Основной функцией опиоидных рецепторов в организме является регулирование болевых ощущений. При активации опиоидного рецептора ингибируется аденилатциклаза, которая играет важную роль при синтезе вторичного посредника ц. АМФ (c. AMP), а также осуществляется регулирование ионных каналов. Закрытие потенциал-зависимых кальциевых каналов в пресинаптическом нейроне приводит к уменьшению выброса возбуждающих нейромедиаторов (таких как глутаминовая кислота), а активация калиевых каналов в постсинаптическом нейроне приводит к гиперполяризации мембраны, что уменьшает чувствительность нейрона к возбуждающим нейромедиаторам БЕЛЯНИН М. Л. 48
Опиоидные рецепторы — разновидность рецепторов нервной системы, относящихся к рецепторам, сопряженным с G белком. Основной функцией опиоидных рецепторов в организме является регулирование болевых ощущений. При активации опиоидного рецептора ингибируется аденилатциклаза, которая играет важную роль при синтезе вторичного посредника ц. АМФ (c. AMP), а также осуществляется регулирование ионных каналов. Закрытие потенциал-зависимых кальциевых каналов в пресинаптическом нейроне приводит к уменьшению выброса возбуждающих нейромедиаторов (таких как глутаминовая кислота), а активация калиевых каналов в постсинаптическом нейроне приводит к гиперполяризации мембраны, что уменьшает чувствительность нейрона к возбуждающим нейромедиаторам БЕЛЯНИН М. Л. 48
 Схема опиоидных рецепторов БЕЛЯНИН М. Л. 49
Схема опиоидных рецепторов БЕЛЯНИН М. Л. 49
, δ (дельта) и κ (каппа), каждая") Различают три группы опиоидных рецепторов: μ (мю), δ (дельта) и κ (каппа), каждая из которых подразделяется ещё на несколько типов. Эффект анальгезии наблюдается при стимуляции любого типа рецепторов. Агонисты μ-рецепторов, кроме того, вызывают угнетение дыхания и седативный эффект, а агонисты κ-рецепторов — психотомиметические эффекты. Действие большинства опиоидных анальгетиков связяно со стимуляцией рецепторов μ-типа БЕЛЯНИН М. Л. 50
Различают три группы опиоидных рецепторов: μ (мю), δ (дельта) и κ (каппа), каждая из которых подразделяется ещё на несколько типов. Эффект анальгезии наблюдается при стимуляции любого типа рецепторов. Агонисты μ-рецепторов, кроме того, вызывают угнетение дыхания и седативный эффект, а агонисты κ-рецепторов — психотомиметические эффекты. Действие большинства опиоидных анальгетиков связяно со стимуляцией рецепторов μ-типа БЕЛЯНИН М. Л. 50

 — ионотропный рецептор глутамата, селективно связывающий N-метил-D-аспартат (NMDA). Участвует в процессах") NMDA-рецептор (NMDAR; НМДА-рецептор) — ионотропный рецептор глутамата, селективно связывающий N-метил-D-аспартат (NMDA). Участвует в процессах запоминания информации (синаптическая пластичность) Схема NMDA-рецептора 1. Клеточная мембрана 2. Канал, блокируемый магнием Mg 2+ 3. Сайт блокировки Mg 2+ 4. Сайт связывания галлюциногенов 5. Сайт связывания цинка Zn 2+ 6. Сайт связывания агонистов (глутамат) иили антагонистов (APV) 7. Сайты гликозилирования 8. Сайты связывания протонов 9. Сайты связывания глицина 10. Сайт связывания полиаминов 11. Внеклеточное пространство 12. Внутриклеточное пространство 13. Комплексная (сложная) субъединица БЕЛЯНИН М. Л. 52
NMDA-рецептор (NMDAR; НМДА-рецептор) — ионотропный рецептор глутамата, селективно связывающий N-метил-D-аспартат (NMDA). Участвует в процессах запоминания информации (синаптическая пластичность) Схема NMDA-рецептора 1. Клеточная мембрана 2. Канал, блокируемый магнием Mg 2+ 3. Сайт блокировки Mg 2+ 4. Сайт связывания галлюциногенов 5. Сайт связывания цинка Zn 2+ 6. Сайт связывания агонистов (глутамат) иили антагонистов (APV) 7. Сайты гликозилирования 8. Сайты связывания протонов 9. Сайты связывания глицина 10. Сайт связывания полиаминов 11. Внеклеточное пространство 12. Внутриклеточное пространство 13. Комплексная (сложная) субъединица БЕЛЯНИН М. Л. 52