Врожденный иммунитет.ppt
- Количество слайдов: 82



Факторы врожденного иммунитета профессор Бажукова Т. А. зав. каф. микробиологии, вирусологии и иммунологии
 иммунитет – это выработанная в")
Врожденный иммунитет • Врожденный (видовой, генетический, конституциональный, естественный, неспецифический) иммунитет – это выработанная в процессе филогенеза, передающаяся по наследству, присущая всем особям данного вида устойчивость к инфекционным агентам (или антигенам).

Факторы врожденного иммунитета 1. Покровный эпителий кожи и слизистых оболочек, обладающий колонизационной резистентностью. 2. Барьер лимфатических узлов 3. Кровь 4. Внутренние органы
")
Факторы врожденного иммунитета • Барьерфиксирующая функция кожи и слизистых (нормальная микрофлора, лизоцим, колонизационная резистентность) • Гуморальные факторы защиты: система комплемента, растворимые рецепторы к поверхностным структурам микроорганизмов (pattern-структуры), антимикробные пептиды, интерфероны • Клеточные факторы защиты (нейтрофилы, макрофаги, дендритные клетки, эозинофилы, базофилы, естественные киллеры-NК)

Барьеры кожи и слизистых • • • Кожа. Кератиноциты рогового слоя – это погибшие клетки, устойчивые к агрессивным химическим соединениям. На их поверхности отсутствуют рецепторы для адгезивных молекул микроорганизмов (исключение S. aureus, Pr. acnae, I. pestis) Продукты потовых и сальных желез: молочная кислота, ферменты, жирные кислоты, антибактериальные пептиды. Клетки Лангерганса и клетки Гринстейна (отростчатые эпидермоциты). Имеют миелоидное происхождение и относятся к дендритным клеткам. По функции эти клетки являются оппозитными. Клетки Лангерганса – АПК (индуцируют иммунный ответ). Клетки Гринстейна продуцируют цитокины, подавляющие иммунные реакции в коже. Резидентная микрофлора кожи
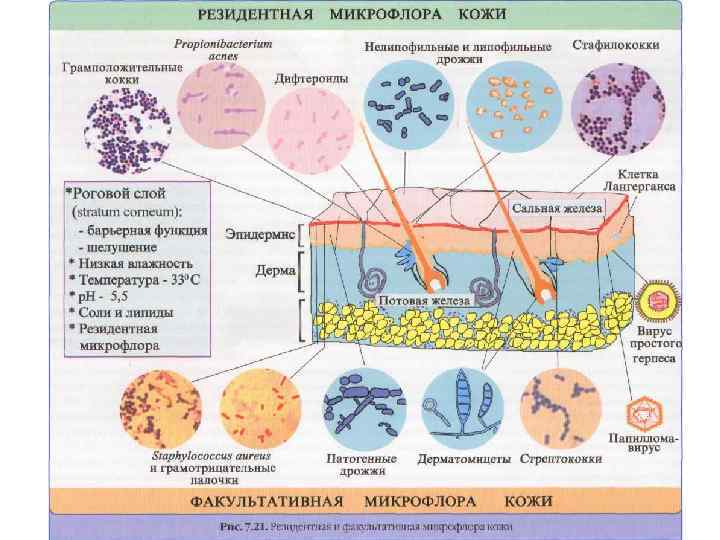

Барьеры кожи и слизистых Слизистые • М-клетки – система облегченного транспорта Аг к ИКК, транслокация бактерий и вирусов через эпителиальный барьер. • Ассоциация эпителиоцитов с лимфоидной тканью (слизистая респираторного тракта, ЖКТ и мочеполовой системы). • Резидентная микрофлора – колонизационная резистентность.

Колонизационная резистентность покровного эпителия • Нормальная микрофлора • Муцин – экранирует сайты адгезии и делает недоступными для бактерий. • Фагоцитирующие клетки и продукты их дегрануляции – продукты лизосом (лизоцим, пероксидаза, лактоферрин, дефензины, токсичные метаболиты кислорода, азота) • Химические и механические факторы – секреты слизистых. • Целенаправленные движения – перистальтика кишечника, реснички мерцательного эпителия. • Секреторные Ig. A

Гуморальные факторы 1. Система комплемента 2. Растворимые рецепторы для патогенов 3. Антимикробные пептиды 4. Семейство интерферонов

Система комплемента • Система комплемента многокомпонентная полиферментная самосорбирующаяся система сывороточных белков, которые в норме находятся в неактивном состоянии. • Комплемент является компонентом многих иммунологических реакций, направленных на освобождение организма от микробов и других чужеродных клеток и антигенов.

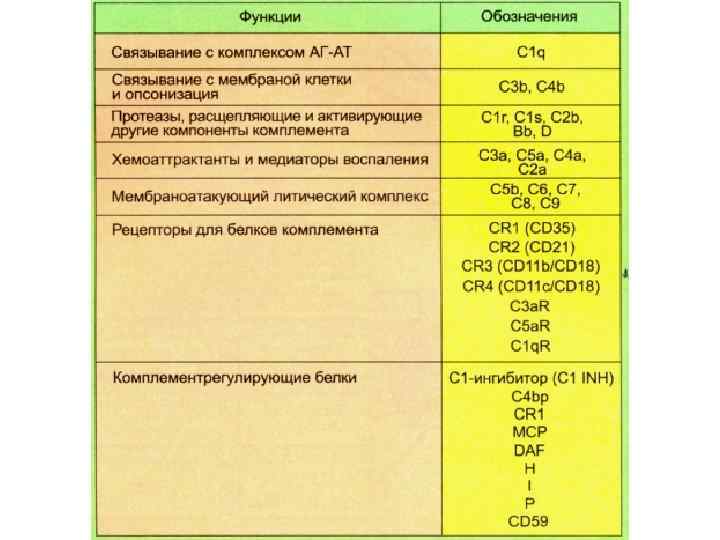
Система комплемента • Входят 20 белков, 9 компонентов: • C 1 -C 1 q, C 1 r, C 1 s; • C 2; • C 3 -C 3 a, C 3 b; • C 4; • C 5 – C 5 a, C 5 b; • C 6; C 7; • C 8; C 9 • Ключевым событием является его активация.

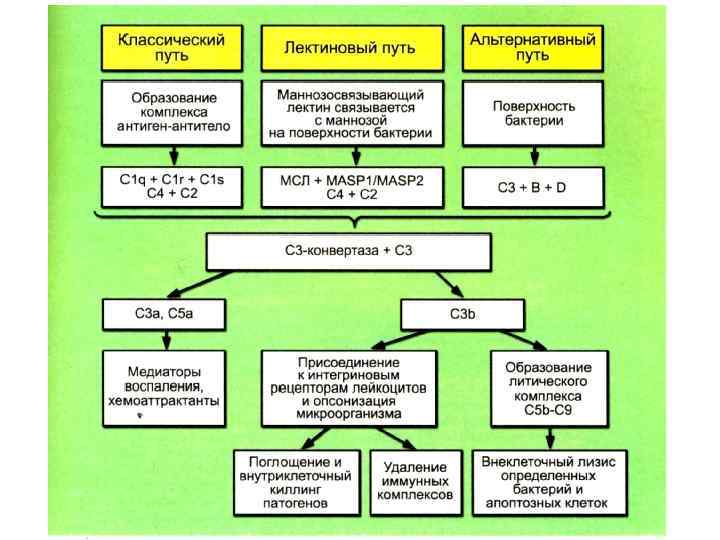
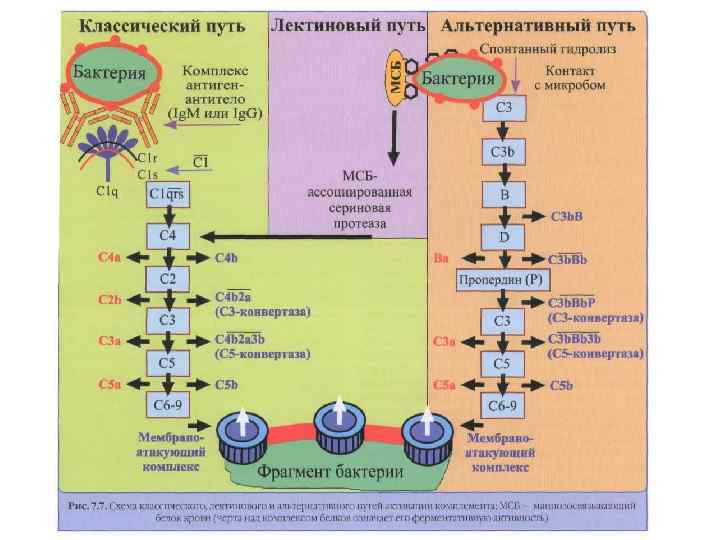
Пути активации системы комплемента 1. Классический – активатор комплекс Аг+Ат 2. Лектиновый – маннан связывающий протеин (лектин) 3. Альтернативный – активатор ЛПС и пептидогликан клеточной стенки ГОБ, вирусы связываются с белками B, D – протеины, Р – пропердин. Активация косплемента протекает в присутствии ионов Са и Mg.

Комплемент

• С 1 состоит из трёх молекул: С 1 q, С 1 r и С 1 s. С 1 q представляет собой структуру, содержащую 6 идентичных глобулярных головок и длинный коллагеновый хвост. • Хвост взаимодействуете двумя другими компонентами С 1: С 1 r и С 1 s обладающими протеазной активностью.

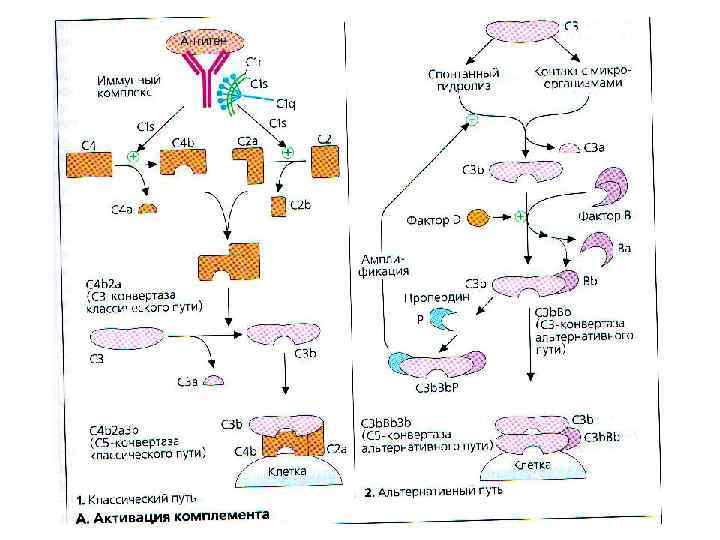
Классический путь активации комплемента • После присоединения АТ класса Ig. M или некоторых изотипов Ig. G к поверхности бактериальной клетки у молекулы АТ образуется участок, способный взаимодействовать с С 1 -компонентом комплемента. • Образуется комплекс С 1 q - С 1 r 2 С 1 s 2. Каждая головка С 1 q взаимодействует с одним участком Fc- фрагмента Ig. При соприкосновении двух и более головок с Ig происходит активация протеазы С 1 r.

Классический путь активации комплемента • C 1 r расщепляет С 4 компонент на С 4 а и С 4 b. • С 4 b ковалентно прикрепляется к поверхности микроорганизма.

Классический путь активации комплемента • C 1 s расщепляет С 2 на С 2 а и С 2 b. • С 2 b прочно прикрепляется к поверхности бактерии, образуя комплекс с С 4 b.

Классический путь активации комплемента • Комплекс С 2 b-С 4 b является активной СЗконвертазой, центральной фигурой классического, лектинового и альтернативного пути активации комплемента. • Протеазной активностью обладает в основном С 2 b. • С 3 -конвертаза расщепляет С 3 -компонент на СЗа и СЗb.

Классический путь активации комплемента • C 3 a остаётся во внеклеточной среде и является мощным хемоаттрактантом и медиатором воспаления (также как С 2 а и С 4 а). • C 3 b прочно присоединяется к поверхности клетки и инициирует соединение с мембраной клетки терминальных компонентов комплемента - С 5 и, С 6, С 7, С 8, С 9. • Образуется литический комплекс, который вызывает формирование пор в мембране клетки и её лизис.

Классический путь активации комплемента

Каскад, генерируемый С 5 конвертазой • • • а. При присоединении СЗb к С 3 конвертазе (С 2 b-С 4 b в классическом пути или СЗb-Вb в альтернативном пути) происходит образование С 5 - конвертазы, прочно прикреплённой к поверхности бактериальной клетки. б. С 5 -компонент расщепляется сериновой протеазой С 2 b в классическом или Вb в альтернативном пути, являющимися компонентами С 5 конвертазы. в. Продуктом расщепления С 5 являются два пептида: С 5 а и С 5 b. Первый поступает во внеклеточную среду и является хемоаттрактантом и медиатором воспаления. С 5 b может находиться во внеклеточной среде или прикрепляться к поверхности клетки и инициировать присоединение литического комплекса.

Образование литического мембраноатакующего комплекса • а. С 5 b-пептид инициирует образование литического комплекса. • б. С 5 b связывает С 6 и С 7. Происходит образование С 5 b-С 6 -С 7 комплекса и внедрение его в липидный слой клеточной стенки. • в. Молекула С 8 состоит из двух протеинов: С 8β и С 8α-γ. С 8β глубоко внедряется в липидный слой. • г. С 8α-γ индуцирует полимеризацию С 9, образуя при этом канал диаметром 100 А, что позволяет воде и солям свободно проходить через него. Нарушаются гомеостаз клетки и происходит её лизис.
 С 3 -конвертаза – спонтанный гидролиз с образованием С 3(H")
Альтернативный путь активации а) С 3 -конвертаза – спонтанный гидролиз с образованием С 3(H 2 O)-формы. б) расщепление С 3 на С 3 а и С 3 b присоединяется к поверхности клетки хозяина или бактерии, реагирует с фактором В, который фактором D расщепляется на Ba и Bb. в) Bb связывается с С 3 b, образуя Bb-С 3 b- конвертазу. г) инактивация Bb-С 3 b-конвертазы (CR 1 и DAF-decay-accelerating factor, MCP-membrane cofactor of proteolysis). Из плазмы поступает вытесняющий фактор H. Эти структуры вытесняют из комплекса Bb. CR 1, MCP и H расщепляют С 3 b протеазой I на неактивный i. С 3 b. д) Комплекс Bb-С 3 b стабилизируется пропердином (фактор Р) и начинает работать как классическая конвертаза С 3 (С 4 b-C 2 b), производя большое количество пептидов С 3 b и С 2 а.

Активация комплемента

Комплемент
 С 1 -ингибитор, связывается с С 1 s и")
Регуляция процесса активации комплемента а) С 1 -ингибитор, связывается с С 1 s и СIr и вытесняет их из комплекса с С 1 q. ограничивает время расщепления С 4 и С 2 эстеразой С 1 s. б) С 3 и С 4 взаимодействуют с клетками хозяина и патогена. DAF, MCP, CR 1, C 4 bp (C 4 -binding protein) используются для инактивации С 4 b, находящегося на поверхности клетки. Фрагмент С 4 b расщепляется протеазой I на 2 пептида: С 4 d и С 4 с. В расщеплении компонента СЗb решающую роль играет сывороточный фактор Н. Он имеет повышенный аффинитет к терминальным сиаловым кислотам. После присоединения фактора Н СЗb расщепляется на пептиды i. СЗb и С 3 dg. в) Инициатором образования литического комплекса является С 5 b. Формируется комплекс С 5 b-С 6 -С 7 -С 8. Присоединение С 9 вызывает образование каналов в мембране клетки. Этому взаимодействию мешает мембранный белок СD 59. Лизиса клетки не происходит.

Биологическая активность С’
")
Биологические эффекты активации комплемента • Образование мембрано-атакующего комплекса (С 5, 6, 7, 8, 9) и лизис клеток. • С 3 а-, С 4 а-и С 5 а – компоненты являются анафилотоксинами, индуцируют дегрануляцию тучных клеток и базофилов (медиторы воспаления). • С 5 а- хемоаттрактант для фагоцитов. • С 3 в- и С 4 в – опсонины, повышают адгезию иммунных комплексов с мембранами макрофагов, нейтрофилов и эритроцитов, усиливают фагоцитоз.

• Комплемент • • Активация комплемента приводит к появлению С 3 - конвертазы, которая превращает С 3 в СЗb, и эта конверсия — центральное событие всего каскада. В свою очередь, СЗb активирует цепочку концевых компонентов комплемента (С 5—С 9), образующих литический комплекс. При активации по классическому пути сначала антиген связывается со специфическими антителами и только затем происходит фиксация СЗ. В альтернативной активации антитела не участвуют. Она начинается ковалентным связыванием СЗb с гидроксильными группами на цитоплазматической мембране микробной клетки. Активация по альтернативному пути служит механизмом неспецифического врожденного иммунитета, тогда как классический путь представляет собой связующее звено между врожденным и приобретенным иммунитетом.
 активацию нейтрофилов, • 2) повышенную")
Комплемент • Анафилатоксин С 5 а вызывает • 1) активацию нейтрофилов, • 2) повышенную экспрессию ими молекул межклеточной адгезии, • 3) эмиграцию нейтрофилов и хемотаксис, • 4) активацию моноцитов и • 5) дегрануляцию тучных клеток, в результате которой происходит сокращение гладкой мускулатуры и повышение проницаемости сосудов.

Комплемент • Компонент С 3, связанный с бактериальной клеткой в виде СЗb или i. СЗb, • 1) взаимодействует с СR 1 эритроцитов, на которых бактерии транспортируются кровотоком, • 2) служит причалом для лизирующего мембрану комплекса на поверхности бактериальных клеток, • 3) сшивает рецепторы комплемента на фагоцитах, • 4) активирует фагоциты, стимулируя фагоцитоз, вспышку клеточного дыхания и бактерицидную активность.

Иммунологическое действие
 системного образования")
Комплемент • Активация комплемента может вызвать патологические реакции в результате • 1) системного образования анафилатоксинов (при септицемии, вызванной грамотрицательными бактериями), • 2) внедрения лизирующего комплекса в мембраны собственных клеток организма (при этом происходит активация клеток и высвобождение метаболитов арахидоновой кислоты, входящей в состав мембран) и • З) фиксации СЗ (привлекающего и активирующего тканевые и циркулирующие лейкоциты) на иммунных комплексах, локализованных в тканях.


Система комплемента

Растворимые рецепторы для патогенов • Белки крови связывающиеся с различными липидными и углеводными структурами микробной клетки (рattern-структурами). Обладают свойствами опсонинов и активируют комплемент. Белки острой фазы: • Пентраксины – высококонсервативные белки с пентамерной структурой: С-реактивный белок (CRB), сывороточный амилоид Р (SAP), пентраксин 3 (PTX 3). • Коллектины и фиколины- Коллектины - олигомерные белки, состоящие из углевод распознающего домена (CRD- лектиновый домен С-типа) и коллагеновой области (маннозосвязывающий лектин (MBL), сурфактантные белки (Sp-A, Sp-D). Фиколины – L, M, H. Углевод распознающий фибриногеноподобный домен. • Белки острой фазы, связывающие железо – трансферрин, гаптоглобин, гемопексин. Препятствуют размножению бактерий, нуждающихся в этих элементах.

Пентраксины

С-реактивный белок • СРБ- связывается с Сполисахаридом бактерий и усиливает фагоцитоз и активацию С 1 g фракции комплемента (классический путь). Синтез в печени и нарастает в ответ на ИЛ-6.

Коллектины и фиколины • Коллектины и фиколины присутствуют во всех биологических жидкостях человека. Осуществляют • опсонизацию бактерий, • активацию комплемента и • играют роль в защите организма от инфекций.
 MBL распознаёт поверхностные структуры, содержащие D-маннозу, L-фукозу,")
Функциональная активность MBL • Углеводраспознающий домен (CRD) MBL распознаёт поверхностные структуры, содержащие D-маннозу, L-фукозу, Nацетилглюкозамин на поверхности грамположительных, грамотрицательных бактерий, грибов, вирусов. Помимо бактерий, белки MBL, Sр-А и Sр-D распознают апоптотические клетки, ишемические ткани, трансформированные клетки, ДНК и др. • По своей структуре MBL похож на C 1 q-компонент комплемента. Находясь в димерной форме, MBL взаимодействует с двумя сериновыми протеазами МАSР- 1 и МАSP-2 (MBL-associated serine protease). Эти протеазы имеют сходство с C 1 r и С 1 q компонентами комплемента.

Функциональная активность MBL • Когда MBL присоединяется к поверхности бактериальной клетки, происходит активация ферментов МАSP которые могут стимулировать лектиновый и альтернативный пути активации комплемента. Лектиновый напоминает классический путь активации. При этом МАSP-2 расщепляет С 2 и С 4 на С 2 b и С 4 b, с образованием С 3 -конвертазы, а последняя — С 3 на СЗа и СЗb, прочно присоединяясь к клеточной стенке бактерии, может инициировать CD 11 b/СD 18 -опосредованный фагоцитоз или образование С 5 -С 9 -литического комплекса, что ведёт к элиминации возбудителя из организма.

Защитная функция MBL • MBL играет важную роль в противовирусной защите. • Этот белок, в частности, опсонизирует ВИЧ, эффективно связываясь с gр120, и предотвращает его взаимодействие с Т-клетками. • Важный защитный путь комплементзависимый лизис вирусинфицированных клеток.

Роль маннаносвязывающего белка в патологии Норма- MBL-0, 1 -50 мг/мл

Роль сурфактантов в защите легких от патогенов и аллергенов а. Эффективное усиление фагоцитоза происходит, если сурфактантом вначале обработать МФ или НФ, а затем добавить микроорганизмы. Это показано на модели взаимодействия М. pneumoniae с МФ и Е. coli с НФ. Такая ситуация соответствует естественному ходу развития инфекции, так как в норме сурфактанты находятся в тесном контакте с клетками дыхательных путей. б. Сурфактанты играют важную роль в предотвращении взаимодействия аллергенов с ТК и базофилами дыхательных путей. Связываясь с углеводными детерминантами аллергена, Sр-А препятствует его взаимодействию с Ig. E-рецепторами. Не происходит дегрануляции ТК или базофилов и выброса гистамина и других биологически активных веществ.

Пропердин • Пропердин – гамма-глобулин нормальной сыворотки. • Активация комплемента по альтернативному пути

Белки сыворотки крови • Бета- лизины – белки сыворотки крови, синтезируются тромбоцитами. Повреждение ЦПМ бактериальной клетки

Фибронектин

Фибронектин

Фибронектин • Фибронектин – белок плазмы и тканевых жидкостей, • синтезируется макрофагами. • Обеспечивает опсонизацию, экранирует дефекты эндотелия, • препятствует тромбообразованию.
 клеточной стенки бактерий")
Антимикробные пептиды 1. Лизоцим – фермент муромидаза вызывает гидролиз муреина (пептидокликана) клеточной стенки бактерий и их лизис. 2. Дефензины и кателицидины – пептиды, обладающие антимикробной активностью. Образуются эукариотическими клетками, содержат 13 -18 аминокислот. Известно около 500 разновидностей. Синтезируются макрофагами и нейтрофилами (αдефензины), а также эпителиальными клетками кишечника, легких, мочевого пузыря.

Антимикробные пептиды • Лизоцим – протеолитический фермент мурамидаза, синтезируется макрофагами и нейтрофилами • Механизм действия: разрушение гликопротеидов клеточной стенки бактерии • Лизис бактерий • Активация фагоцитоза

Семейство интерферонов • Интерферон –открыт в 1957 г Айзексом и Линдеманом при изучении интерференции вирусов (лат. inter-между, ferens-несущий). Интерференция – явление когда ткань инфицированная одним вирусом становится устойчивой к заражению другим вирусом, вызывается белком продуцируемым этими клетками. • Интерферон –гликопротеид. Выделяют интерфероны I и II типов.

Интерферон • α –интерферон лейкоциты • β - интерферон фибробласты • γ - интерферон – Тлимфоциты, макрофаги, ЕК. • Механизм действия: связывается с рецепторами клетки и блокирует синтез белка в клетке (препятствует размножению вирусов)

Интерфероны • • • I тип включает ИНФ α и β. α–интерферон - лейкоциты β- интерферон - фибробласты ζέ – интерферон – трофобласты λ-интерферон, κ-интерферон. Механизм действия ИНФ α и β: активация в клетке генов, блокирующих репродукцию вирусов (индукция синтеза протеинкиназы R, нарушение трансляции м. РНК и запуск апоптоза зараженной клетки через Bcl-2 и каспазазависимые реакции. Другой механизм – активация латентной РНКэндонуклеазы, вызывающей деструкцию вирусной НК.

Интерфероны • II тип включает γ-интерферон – продуцируется Т-лимфоциты и ЕК после Аг стимуляции.

Интерферон

Клетки-продуценты IFNα/β • п. ДК-плазмоцитоподобные ДК синтезируют IFNα/β в 1000 раз больше (ss. РНК, неметилированные бактериальные и вирусные Cp. G-последовательности) • м. ДК – миелоидные ДК (ds. РНК)

Функции интерферона α/β
 в")
Влияние IFNα/β на ДК и В-лимфоциты • IFNα/β вызывают превращение незрелых ДК (1) в зрелые (2). Усиление ими синтеза цитокинов, хемокинов, экспрессии молекул МНС, особенно 1 класса, костимуляторных молекул, экспрессии и секреции главных факторов выживания и активапии В 2 -клеток. - ВIys (BAFF) и Аргil. Эти лиганды, взаимодействуя с рецепторами В-клеток BAFFR и ТАСI, при участии цитокинов IL-10, TGFβ и IL 15, синтезируемых активированными ДК, вызывают у наивных В-клеток (3) переключение иммуноглобулиновых генов Сμ-Сγ и Сμ-Сα и их созревание в Ig. G (4)- и Ig. A(5)-плазмабласты. Этот процесс может происходить независимо от взаимодействия СD 40 -СD 40 L и его избыточная активация является одной из причин развития аутоиммунных заболеваний. IFNα/β вызывают активацию МФ. IFNα/β являются необходимыми цитокинами для созревания и пролиферации В 1 - клеток.

Влияние IFNα/β на Т-клетки • IFNα/β: способствуют выживанию и пролиферации Т-СD 4+ и СD 8+ • усиливают цитотоксические свойства СD 8+ Т-клеток, МФ и NК. • обладают мощным антипролиферативным и проапоптозным эффектом, усиливая зкспрессию проапоптотических молекул. • Наличие одновременно пролиферативных и антипролиферативных свойств связано с включением на различных этапах инфекционного процесса или адаптивного иммунного ответа различных регуляторных механизмов, определяющих чувствительность клетки к действию IFNα/β.

IFNγ • IFNγ- гомодимер, состоящий из двух полипептидных цепей. • Продуцентами IFNγ могут быть различные типы клеток. Немедленный ответ на внедрение патогена дают NKТклетки, NK-клетки, а также плазмацитоидные ДК и МФ и редко НФ. • При развитии адаптивного иммунитета мощными продуцентами IFNγ являются СD 8+ цитотоксические лимфоциты и СD 4+ Тh 1 -клетки. • В-лимфоциты могут также синтезировать IFNγ. • IFNγ оказывает существенный эффект практически на все клетки иммунной системы.

Функции IFNγ

Фагоциты

Рецепторы макрофагов

Механизм эндоцитоза

Механизм эндоцитоза

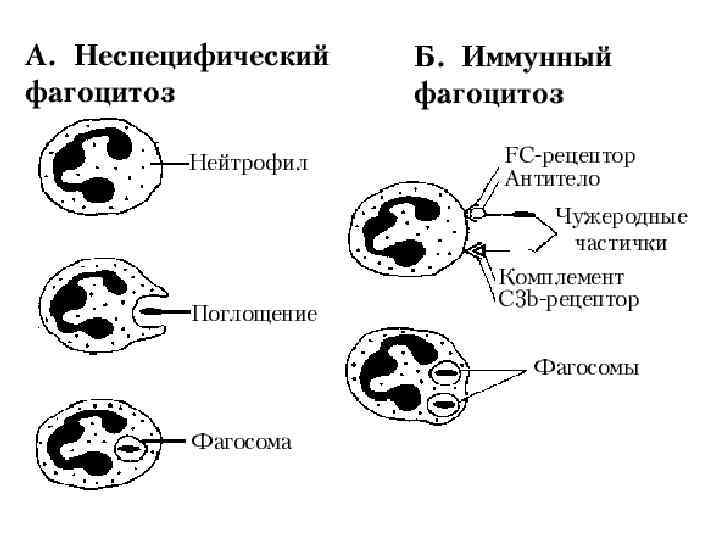
Фагоцитоз

Фагоцитоз

Механизмы действия фагоцитов
 частиц к")
Этапы фагоцитоза 1. 2. 3. 4. 5. Активация и хемотаксис Адгезия (прикрепление) частиц к поверхности фагоцита Поглощение частиц, их погружение в цитоплазму и формирование фагосомы Образование фаголизосомы Внутриклеточный киллинг и переваривание – активация лизосомальных гранул (кислороднезависимая бактерицидность), усиление потребления кислорода и глюкозы – окислительный взрыв с образованием токсических метаболитов кислорода и азота (перекись водорода, супероксиданион кислорода, гипохлорная кислота, пироксинитрит) – кислородзависимая система бактерицидности.

Миграция фагоцитов

Фагосома

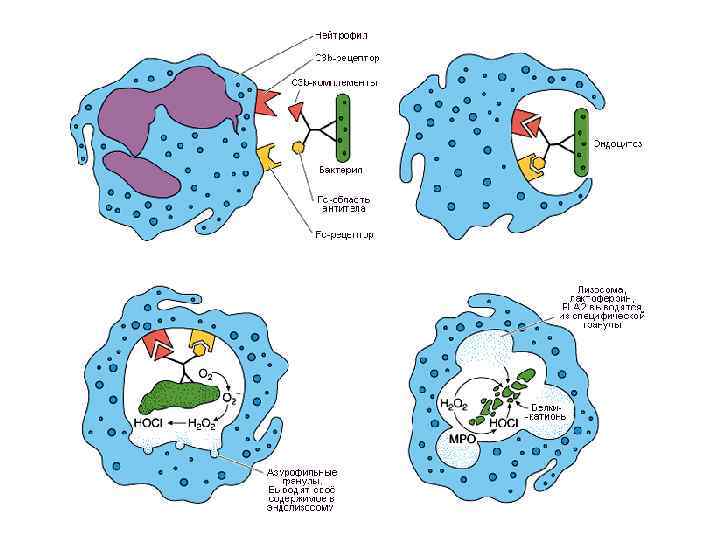

Незавершенный фагоцитоз
 – активация")
Эозинофилы • Слабая фагоцитарная активность • Защита от паразитов (гельминты и простейшие) – активация эозинофилов и выделение токсических продуктов из гранул (катионный белок; РНК-аза – образование мембранных каналов в оболочке паразита; пероксидаза – образование токсичных галидов; главный основной белок – полимеризуется в мембране паразита с образованием трансмембранных пор).

Действие NK-клеток на клетки – мишени CD 95 – маркер апоптоза клетки CD 16 – Fc. R, антителозависимая цитотоксичность CD 56 – активация цитотоксичности и продукции цитокинов CD 94 – ингибиция/ активация цитотоксичности
Врожденный иммунитет.ppt