Лекция_ЭВ_В-СОб_2.ppt
- Количество слайдов: 41



Эволюция выделения и водно-солевого обмена
 - совокупность процессов, обеспечивающих удаление из организма животных веществ, количества которых превышают")
Выделение (экскреция) - совокупность процессов, обеспечивающих удаление из организма животных веществ, количества которых превышают метаболические потребности. К ним относятся СО 2, Н 2 О, неорганические ионы, продукты белкового обмена и т. д. Водно-солевой обмен - совокупность процессов всасывания, распределения, потребления и выделения воды и солей в организме животных и человека. Водно-солевой обмен обеспечивает постоянство объема, осмотических концентраций, ионного состава и кислотнощелочного равновесия внутренней среды организма (гомеостаз)

Продукты белкового обмена и их выделение l l l Аммиак: высоко токсичен, легко растворим. выведение требует очень большого количества воды Аммониотелия – широко распространена у первичноводных животных: большинства п/в беспозвоночных, пресноводных костистых и двоякодышащих рыб; а также у крокодилов Мочевина: менее токсична, растворима выведение требует достаточно большого количества воды Уреотелия – среди беспозвоночных лишь у некоторых плоских червей, аннелид, моллюсков; среди позвоночных у хрящевых рыб, морских костистых рыб, амфибий, млекопитающих Мочевая кислота: мало токсична, плохо растворима выведение требует небольшого количества воды Урикотелия – у большинства наземных беспозвоночных; среди позвоночных у большинства рептилий, у птиц Теплота сгорания, к. Дж/моль: Аммиак – 378 Мочевина - 638 Мочевая кислота - 1932

Аспекты регуляции водно-солевого баланса l Волюморегуляция: поддержание постоянства объема жидкости (у одноклеточных – внутриклеточной, у многоклеточных – внеклеточной) l Ионорегуляция: поддержание постоянства концентрации отдельных ионов в циркулирующей жидкости l Осморегуляция: поддержание постоянства осмотического давления циркулирующей жидкости Процессы мочеобразования l l l Фильтрация Секреция Реабсорбция

Направления эволюции осмотического баланса Пойкилоосмия: внутренняя среда изоосмотична внешней, её осмолярность изменяется вслед за колебаниями осмолярности внешней среды. l Животные - осмоконформеры (большинство первичноводных морских беспозвоночных) l l l проницаемые покровы тела нет системной регуляции осмолярности ЦЖ возможны отличия ионного состава (т. е. наличие ионорегуляции) большинство - стеногалинные (жизнеспособны в пределах небольших колебаний солености воды) относительная эвригалинность достигается развитием изолирующих приспособлений (смыкание створок, выделение слизи) и внутриклеточных механизмов осмотической устойчивости (повышение содержания в цитоплазме связывающих воду белков, аминокислот, калия)

Направления эволюции осмотического баланса l ü ü ü Гомоосмия: при внешних колебаниях осмотического давления внутренняя среда сохраняет постоянную осмолярность, может быть гипо- или гиперосмотична по отношению к внешней среде. Животные – осморегуляторы: отчасти пресноводные беспозвоночные, наземные беспозвоночные, позвоночные l развивается в связи с освоением пресных вод и наземновоздушной среды l малопроницаемые покровы тела l системная регуляция осмолярности ЦЖ на основе активной секреции и реабсорбции ионов эпителиями жабр, ЖКТ и «органов выделения» l большинство - эвригалинные

ОДНОКЛЕТОЧНЫЕ: Контрактильная вакуоль l l У простейших одноклеточных организмов примитивным «выделительным» органоидом является контрактильная вакуоль. Контрактильная вакуоль – органоид, функция которого тесно связана с регуляцией количества воды и солей в организме. Инфузория туфелька: 7 – сократительная вакуоль. Образование контрактильной вакуоли у амебы.

l Скорость образования и опорожнения контрактильной вакуоли зависит от осмолярности окружающей среды. Инфузории Время между двумя опоражниван иями (в сек. ) Среда обитания Характеристика внешней среды по отношению к животному Очень гипотоническая То же Почти изотоническая То же Изотоническая Chilodon cucullulus 5 Пресная вода Euplotes Lagynus crassicollis 25 – 30 120 Пресная вода Морская вода Acineria incurvata Cryptochilum echini 360 – 720 1200 l l Морская вода Паразит морского ежа Контрактильная вакуоль – органоид, обеспечивающий волюморегуляцию Экскреция азотистых продуктов – через всю клеточную мембрану

МНОГОКЛЕТОЧНЫЕ l Выделение продуктов белкового обмена: диффузия через покровы тела и дыхательные поверхности (преобладает у первичноводных беспозвоночных и позвоночных) l нагнетание или фильтрация жидкости в канальцы выделительных органов l секреция в просвет выделительного канальца или кишечной трубки Роль органов выделения в экскреции азотистых продуктов возрастает с развитием малопроницаемых покровов, уреа- и урикотелии и выходом на сушу. l l Обеспечение водно-солевого баланса l l Пассивный транспорт воды по осмотическому (или гидростатическому) градиенту через эпителий покровов тела, жабр, кишечной трубки, выделительных органов Активный транспорт ионов: l l l Абсорбция (поглощение) ионов в пресной воде жабрами, кишечником Секреция ионов в соленой воде жабрами, кишечником, канальцами выделительных органов Реабсорбция канальцами выделительных органов, кишечнником Роль органов выделения в поддержании водно-солевого равновесия возрастает в связи с освоением пресных вод и суши


Выделительные системы многоклеточных l Контрактильная вакуоль - Губки l Нет специализированной системы – Кишечнополостные, Иглокожие l Вентральная железа - Круглые черви Нефридии (трубки эктодермального происхождения) l l Протонефридии l l l Метанефридии – Олигохеты Нефромиксии (трубки экто- мезодермального происхождения) - Полихеты Целомодукты (трубки мезодермального происхождения) l l Антеннальные и максилярные железы - Ракообразные Коксальные железы - Паукообразные Перикардиальная «почка» - Моллюски Почка позвоночных (нефрос – скопление целомодуктов) l l С пламенными клетками – Плоские черви, Немертины С бульбусами – Коловратки С соленоцитами – Полихеты, Приапулиды, Гастротрихи Головная почка (в эмбриогенезе) Туловищная почка - Анамнии Тазовая почка – Амниоты Мальпигиевы сосуды (трубчатые выросты на границе средней и задней кишки) - Насекомые

Протонефридиальная система Пламенная клетка ресничного червя: 1 — ядро; 2 — мерцательный жгутик. Протонефридиальная система Planaria maculata. А – общая схема. 1 – выводные протоки; 2 – отверстие общего выводного протока. В – при большом увеличении. 3 – выводные протоки пртонефридиальной системы с пламенными клетками на концах (4).

Протонефридиальная система Протонефридии турбеллярии: А-клетка с ресничным пламенем, Б-система протонефридиев; 1 -ядро; 2 -терминальная клетка; 3 -жгутики; 4 -трубчатая клетка; 5 -ядро трубчатой клетки; 6 -соединение клеток; 7 -отверстия; 8 -нефридиальные поры; 9 -нефридиальные каналы.

Протонефридиальная система l l От типичных протонефридиев несколько отличается выделительная система некоторых Polychaeta и Priapulida, у которых она состоит из сегментарно расположенных органов, кончающихся группой слепых вздутий, носящих название соленоцитов. Каждый соленоцит состоит из грушевидной клетки с внутриклеточным просветом, в который вдается длинный жгутик – гомолог «пламенных ресничек» , терминальных клеток типичных протонефридиев. Слепой конец протонефридия кольчатого червя с сидящими на нём соленоцитами: 1 — соленоциты; 2 — выделительная трубка протонефридия; 3 — ядро; 4 — жгутик соленоцита.

Выделительная система нематод

Метанефридиальная система Метанефридии: 1 воронка нефростома, 2 —клетки мерцательного эпителия, З — входная щель, 4— 7 — выделительный канал, 8 — нефропора.

Системы на основе целомодуктов Выделительные органы, развивающиеся из мезодермы, — так называемые целомодукты — имеются у моллюсков. У моллюсков, утративших сегментацию, имеется всего одна пара нефридиев. Перикардиальная полость моллюсков – остаток целомической полости. В нее через стенку желудочка фильтруется жидкость. В перикардиальную полость открывается нефростом, через который жидкость попадает в систему каналов – почечный мешок и мочеточники, и затем – наружу. В проксимальном канальце всасывается глюкоза, натрий и хлор. У морских форм в мочу выделяются ионы калия, магния и сульфаты. Конечная моча морских моллюсков изотонична гемолимфе, пресноводных – гипотонична.

Системы на основе целомодуктов У ракообразных выделительные органы — извитые трубки, начинающиеся слепым «целомическим» мешочком и открывающиеся наружу у основания антенн или максилл (отсюда названия «антеннальные» и «максиллярные» железы). Сходны выделительные органы ( «коксальные железы» ) у мечехвостов, открывающиеся у основания 5 -й пары ног, у паукообразных – 6 -й сегмент. Открывающиеся наружу выделительные органы, через которые выводятся легко растворимые продукты обмена, типичны для водных беспозвоночных. Антеннальная железа речного рака (в расправленном виде): 1 — целомический мешочек; 2 — «зелёный канал» ; 3 — промежуточный канал; 4 — «белый канал» ; 5 — мочевой пузырь; 6 — выводной проток; 7 — наружное отверстие железы.

Мальпигиевы сосуды – основной орган выделения Трахейнодышащих. У насекомых дополнительную функцию выделения выполняют почки накопления: клетки жирового тела и перикардиальные клетки вокруг сердца. Продукты обмена откладываются и в покровную кутикулу, что удаляется с линькой. Экологическ ая группа Количество мальпи гиевых телец Их значение Водные насекомые Осморегуляция по типу разведения Насекомые аридных зон 1 -6 -передняя кишка; 7 -9 -средняя кишка; 10 -задняя кишка; 11 -анус; 12 -мальпигиевы сосуды. до 150 1 -2 пары Экономия влаги в организме через обратное всасывание воды в гемолимфу

Эволюция выделительной системы Хордовых Ланцетник Выделительная система ланцетника – нефридиального типа. По строению близка к выделительной системе некоторых кольчатых червей. Нефридии у ланцетников – короткие изогнутые трубки (до 90 пар), расположенные в области глотки. Большая часть нефридия лежит в целоме. Здесь имеется ряд отверстий (они называются нефростомы), окруженных булавовидными клетками (соленоцитами). Головка соленоцита, представляющая тело клетки с ядром, переходит в длинный полый стебелёк, или трубочку, которая открывается в просвет главного канала нефридия. имеет тонкий каналец с мерцательным волоском, обеспечивающим ток целомической жидкости. Выводящий конец нефридия открывается в атриальную полость. Продукты метаболизма поступают в полость целома из капилляров, пронизывающих стенки целома, а затем за счет движения мерцательных волосков соленоцитов выводятся в атриальную полость и через атриопор – наружу. Нефридий ланцетника. Часть нефридия с семью соленоцитами (А) и каналец с нефростомами (Б): 1 -булавовидные клетки-соленоциты; 2 -отверстие в полость тела (целом); 3 -отверстие в атриальную полость; 4 -жаберная щель.

Возникновение и эволюция почки У позвоночных органами выделения служат типичные целомодукты, скопления которых образуют почки. От почек отходят мочеточники, которые впадают непосредственно в клоаку или в мочевой пузырь, открывающийся наружу мочеиспускательным каналом. В эволюционном ряду позвоночных, как и при индивидуальном развитии высших позвоночных, наблюдается последовательная смена трёх типов почек. Первыми возникают передние, или головные, почки, называемые также предпочками (пронефрос). Далее развиваются первичные, средние, или туловищные, почки — вольфовы тела (мезонефрос). Последними появляются вторичные, задние, или тазовые, почки (метанефрос).

Эволюционные преобразования системы выделения и водно-солевого обмена в связи с переходом позвоночных к наземному образу жизни. Схема эволюции почки у позвоночных животных (по Смиту). Сверху – наземные позвоночные (птицы, рептилии и млекопитающие); снизу – водные позвоночные (хрящевые и костные рыбы, амфибии); слева – пресноводные; справа – морские.


Рис. 1. Схематическое изображение эволюции сегментов нефрона почек позвоночных животных. 1 – клубочек; 2 – шейка; 3 – проксимальный извитой каналец; 4 - промежуточный сегмент; 5 – тонкий сегмент петли Генле; 6 – дистальный извитой каналец; 7 – начало собирательной трубочки; 8 – особый сегмент хрящевых рыб.

Круглоротые Целомодукты предпочки и первичной почки, обычно по паре на сегмент, возникают из нефротома. Часть нефротома, открывающаяся во вторичную полость тела, образует мерцательную воронку (нефростом). Против нефростомов предпочки в полости тела обычно развивается общий для всех воронок клубок кровеносных сосудов; через него фильтруются из крови жидкие продукты выделения (водные растворы солей), которые и поступают в воронки. Предпочка, имеющая сегментарное строение и тянущаяся вдоль всего туловища, сохраняется в продолжении всей жизни из круглоротых только у бделлостомы. У миног остатки пронефроса, образующие передний отдел почки, открываются в перикардиальную полость. У других позвоночных пронефрос служит лишь личиночным органом. Продольный разрез речной миноги: 1 — хорда, 2 — соединительнотканная оболочка хорды, 3 — мозговой череп, 4 — хрящи ротовой воронки, 5 — околосердечный хрящ, 6 — миомер, 7 — миосепта, 8 — мускулатура языка, 9 — головной мозг, 10 — спинной мозг, 11 — обонятельный мешок, 12 — питуитарный вырост, 13 — ротовая полость, 14 — пищевод, 15 — кишка, 16 — анальное отверстие, 17 — печень, 18 — жаберные мешки, 19 — дыхательная трубка, 19 а — внутренние отверстия жаберных мешков, 20 — предсердие, 21 — желудочек, 22 — венозная пазуха, 23 — брюшная аорта, 24 — почка, 25 — мочеточник, 26 — мочеполовой синус, 27 — половая железа, 28 — мочеполовой сосочек, 29 —- половая пора

Рыбы У всех рыб и земноводных во взрослом состоянии функционируют первичные почки. Канальцы их развиваются у эмбриона сходно с канальцами предпочки, отличаясь лишь тем, что открываются в готовый пронефрический проток, который получает с этого времени название первичнопочечного канала. У акуловых рыб этот канал расщепляется с переднего конца на два протока: вольфов канал, служащий мочеточником первичной почки, и мюллеров канал, вступающий в связь с остатками пронефроса и выполняющий у самок функцию яйцевода. Стереограмма головных и развивающихся первичных почек у позвоночных: 1 — аорта; 2 — вторичная полость тела; 3 — клубок; 4 — брыжейка; 5 — пронефрическая трубочка; 6 — пронефрический проток; 7 — полость нефротома (8); 9 — воронка; 10 — мезонефрическая трубочка; 11 — хорда; 12 — брюшина; 13 — миотом; 14 — спинной мозг.

Стереограмма сформированных первичных почек у позвоночных: 1 — аорта; 2 — вторичная полость тела; 3 — половой валик; 4 — клубочек; 5 — брыжейка; 6 — мальпигиево тельце; 7 — мезонефрическая трубочка; 8 — миотом; 9 — воронка; 10 — хорда; 11 — брюшина; 12 — вольфов проток; 13 — спинной мозг.
 и")
l У некоторых костных рыб почка утратила почечные клубочки (агломерулярная - бесклубочковая почка) и отфильтровывает из крови очень мало воды. А так как жидкости тела и кровь костных рыб гипотоничны по отношению к морской воде, то для восполнения воды морские костные рыбы пьют воду, а избыток солей удаляют с помощью специализированных солевых желез, расположенных в жабрах. l Акуловые рыбы решают эту проблему другим путем: аммиак, образующийся в результате обмена веществ, в их организме превращается в мочевину, накапливающуюся в крови, способствуя тем самым повышению осмотического давления внутри тела. Почки у акуловых выделяют гипертоническую мочу (они не пьют морскую воду), часть воды поступает осмотическим путем через покровы. l Пресноводные рыбы редко пьют воду, а их жабры поглощают соли путем активного переноса почки же выделяют обильную малоконцентрированную мочу для удаления излишка воды, которая всасывается через жабры и выстилку ротовой полости. l У проходных рыб происходит перестройка водно-солевого обмена при переходе из моря в реки и наоборот.

Морские хрящевые рыбы, достигнув изотонии со средой за счет накопления в тканях мочевины, выделяют небольшое количество мочи: всего от 2 до 50 мл/кг в сутки. Некоторые скаты и рыба-пила, не смотря на снижение осмотического давления и содержания мочевины, остаются гипертоничными по отношению к среде (опресненная вода). Им всегда грозит обводнение. В результате усиливается водно-выделительная функция почек (250 мл/кг) и адсорбция солей в почечных канальцах. Такой тип водно-солевого обмена и ионного равновесия со средой требует минимальных затрат энергии. Экологические группы внешней крови Содержание мочевины в крови, % 1, 25 -2, 60 среды Хрящевые рыбы: морские 1, 80 -2, 30 1, 90 -2, 30 пресноводные 0, 02 -0, 03 0, 90 -1, 20 0, 10 мочи 1, 87 -2, 3 0, 49 -0, 78

Пресноводные костные рыбы живут в гипотонической среде, поэтому вода постоянно проникает в организм через кожу, жабры, с пищей (Рис. ). В результате возникает угроза излишнего «обводнения» . Эта проблема была решена благодаря: l Высокому развитию фильтрационного аппарата почек (выведение избытка воды) – 50 -300 мл/кг за сутки; l Активной реабсорбцией солей в почечных канальцах (задержка солей); За 3 -4 часа поступающей извне водой заменяется вода организма. Морские костные рыбы в гипертонической среде находятся под угрозой «иссушения» вследствие потерь воды через кожу, жабры, с мочой и фекалиями. Возник новый механизм водно-солевого обмена: l Уменьшилось число клубочков в почках, а у части они исчезли полностью (агломерулярные почки) – выделение воды: 0, 5 -20 мл/кг в сутки; l Восстановление водно-солевого баланса происходит за счет употребления морской воды (40 -200 мл/кг в сутки); l Усилилась роль почечных канальцев, жабр и ЖКТ, клетки которых увеличили выведение избытка солей и продуктов азотистого обмена Экологические группы внешней среды крови Содержание мочевины в крови, % мочи Костистые рыбы: морские 1, 80 -2, 30 0, 67 -0, 91 0, 01 -0, 03 0, 09 -0, 90 пресноводные 0, 02 -0, 03 0, 42 -0, 70 0, 005 -0, 03 0, 07 -0, 08
 и морских (2) костистых рыб.")
Схема механизмов осморегуляции у пресноводных (1) и морских (2) костистых рыб.

Амфибии Парные компактные туловищные почки расположены по бокам позвоночного столба в задней части полости тела. У переднего края каждой почки располагаются жировые тела – резерв питательных веществ для формирования половых клеток. Структурно-функциональной единицей почек является нефрон (его строение и функционирование принципиально не отличается от такового у рыб). Основным продуктом азотного обмена у лягушки является мочевина, а у головастиков и Хвостатых амфибий, живущих в воде – аммиак. По внешнему краю каждой почки тянется тонкий канал – мочеточник (вольфов канал), открывающийся в клоаку. В клоаку открывается и мочевой пузырь. Моча накапливается в мочевом пузыре, концентрируется а затем выводится наружу. Органы размножения и выделения самки лягушки

Амфибии У Амфибий в связи с переходом к обитанию в наземно-воздушную среду происходили изменения в сторону экономии воды: l Дальнейшее совершенствование клубочковых процессов; l Интенсификация реабсорбции в почечных канальцах ( обратное всасывание ценных низкомолекулярных веществ, ионов натрия, пассивная реабсорбция воды); l Обратное всасывание воды в мочевом пузыре; l Большое значение приобретает кожа в поддержании водного обмена (Так, кожа населяющей сырые биотопы травяной лягушки с каждого см 3 испаряет до 30 мм 3 /час воды); l Огромную роль в водном обмене играет приспособительное поведение (активны ночью, в пасмурную погоду, используют различные укрытия в жаркие периоды). Экологическая группа Критические пределы обезвоживания, % Обитатели водоемов (протеи) 30 -35% Амфибии сырых биотопов (лягушки) 40 -50% Живущие на деревьях (некоторые квакши) 70%

Рептилии У амниот вторичная почка заменяет функционирующую в эмбриональном периоде первичную почку, которая у взрослых самок вообще дегенерирует, а у самцов несёт функцию семяпроводящего аппарата и называется придатком семенника, или эпидидимисом. l Эмбрионально вторичная почка образуется из специального мочеточника, вырастающего из заднего конца вольфова канала Этот мочеточник имеет на конце несколько конечных разветвлений, в которые впадают почечные канальцы, возникающие из нефрогенной ткани. Извитые канальцы вторичной почки никогда не имеют воронок и начинаются боуменовой капсулой. l Особенностью нефронов тазовых почек является полное отсутствие связи с полостью тела. У крокодилов и пресноводных черепах выделяется жидкая моча, у наземных Рептилий – твердая или кашицеобразная. Основным продуктом азотного обмена является мочевая кислота. Особенности выделительной системы настолько обеспечивают независимость рептилий от воды, что позволили им захватить совершенно засушливые области Земли l
. Есть")
В общем в сравнении с Амфибиями происходит увеличение числа нефронов (около 5 тыс). Есть существенная разница в строении нефронов внутри класса: • У форм, ведущих полуводный образ жизни (крокодилы, черепахи) имеются достаточно развитые сосудистые клубочки; • У чешуйчатых они развиты слабо и мочеотделение в этом случае идет в основном за счет секреции извитых канальцев. Т. о. скорость фильтрации у Рептилий значительно ниже, чем у Амфибий. Солевые железы рептилий – дополнительные органы выделения. Урикотеличность (выделение мочевой кислоты) у рептилий объясняется появлением «замкнутых» яиц, а т. ж. потребностью в экономии воды. Классы Скорость фильтрации, мл/(кг ч) Рептилии: черепахи, крокодилы чешуйчатые 5 -10 4 -5 Амфибии: бесхвостые 34 -35

Птицы Выделительная система птиц типичная для амниот. Почки компактные, сегментированные, заполняют углубления подвздошных костей. У большинства видов имеется петля Генле – тонкая вставка между проксимальной и дистальной частями извитого канальца. Петля Генле обеспечивает эффективное всасывание воды. Мочевого пузыря нет. Есть клоака.

Выделительная система птиц характеризуется следующими особенностями: l Крупная величина почек, в сравнении с рептилиями и даже млекопитающими, что стоит в прямой связи с интенсификацией обменных процессов птиц; l Количество нефронов исчисляется уже десятками тысяч; l Как и для рептилий, характерна редукция сосудистого клубочка; мочеотделение происходит в основном за счет секреторной деятельности извитых канальцев; l У морских птиц дополнительным органом выделения служат специальные солевые железы. l Урикотеличность, что появилась у рептилий закрепляется. Представитель Масса почки от общей массы тела, % скворец 1, 2 крачка 1, 6 крохаль 2, 6

Млекопитающие У Млекопитающих, как и у всех амниот, имеются тазовые почки. Рассмотрим подробнее строение выделительной системы на примере человека. l На разрезе почки выделяют два слоя: корковый (наружный) и мозговой (внутренний). l Корковое вещество содержит нефроны (в состав одной почки входит свыше миллиона нефронов). Мозговое вещество состоит из собирательных трубочек и кровеносных сосудов, собранных вместе в виде почечных пирамид. Верхушки пирамид, называемые сосочками, открываются в малые почечные чашечки. Малые почечные чашечки открываются в большие почечные чашечки, из которых моча поступает в почечную лоханку. Почечная лоханка образует расширенное устье мочеточника. Мочеточники дают начало мочевыводящим органам: они выходят из почки и впадают в мочевой пузырь. Мочевой пузырь через мочеиспускательный канал открывается наружу мочеиспускательным отверстием.
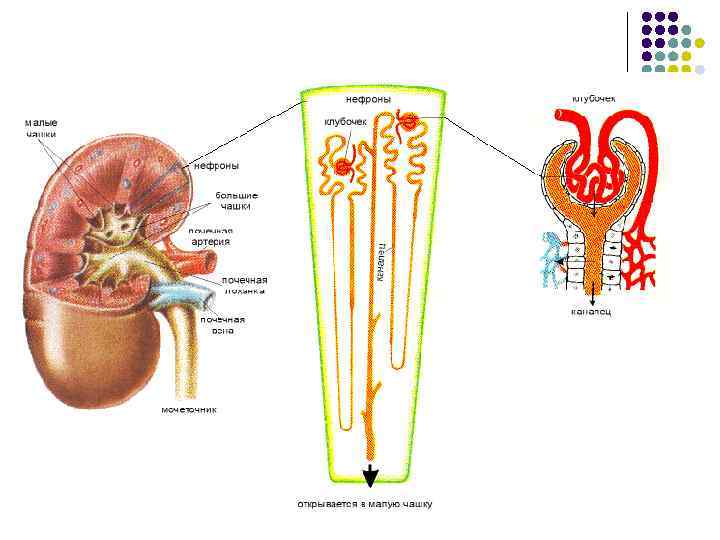

У Млекопитающих происходило дальнейшее совершенствование метанефроса. В сравнении с предыдущими классами у них произошли следующие преобразования: l l l Процесс фильтрации (180 л) и реабсорбции (178, 5 -179 л/сутки) протекают с высокой скоростью; У млекопитающих мозговой слой почки получает максимальное выражение из-за развития петли Генле; Очень большое количество почечных канальцев; Относительные размеры почек находятся в обратной зависимости от величины тела; Мочевинный тип обмена, что объясняется образованием плаценты. Представители различных классов Число почечных канальцев Млекопитающие: мышь кролик 10 тыс 285 тыс Амфибии: лягушка тритон 2 тыс 400 Классы Доля мочевой кислоты, % Доля мочевин ы, % Птицы 63 -80 1 -10 Млекопитающие 0, 1 -8 68 -91

Теория, объясняющая смену почек Наиболее вероятной теорией, объясняющей последовательную смену почек, является теория смены почечных канальцев. Сторонники этой теории считают, что у предков позвоночных была длинная почка, построенная по типу пронефроса. У их рыбообразных потомков на спинной стороне канальцев предпочки развились канальцы более совершенного типа — канальцы первичной почки, которые и вытеснили предпочечные канальцы на протяжении всего туловища, за исключением его переднего конца. У амниот канальцы вторичной почки, развивавшиеся в заднем отделе первичной почки, постепенно вытеснили последнюю.
Лекция_ЭВ_В-СОб_2.ppt